Lipopolysaccharide-Linked Enterobacterial Common Antigen (ECALPS) Occurs in Rough Strains of Escherichia coli R1, R2, and R4


Abstract
:1. Introduction
2. Results
2.1. Isolation and Purification of E. coli R1, R2, R3, R4, and O39 ECALPS-Derived Poly- and Oligosaccharides
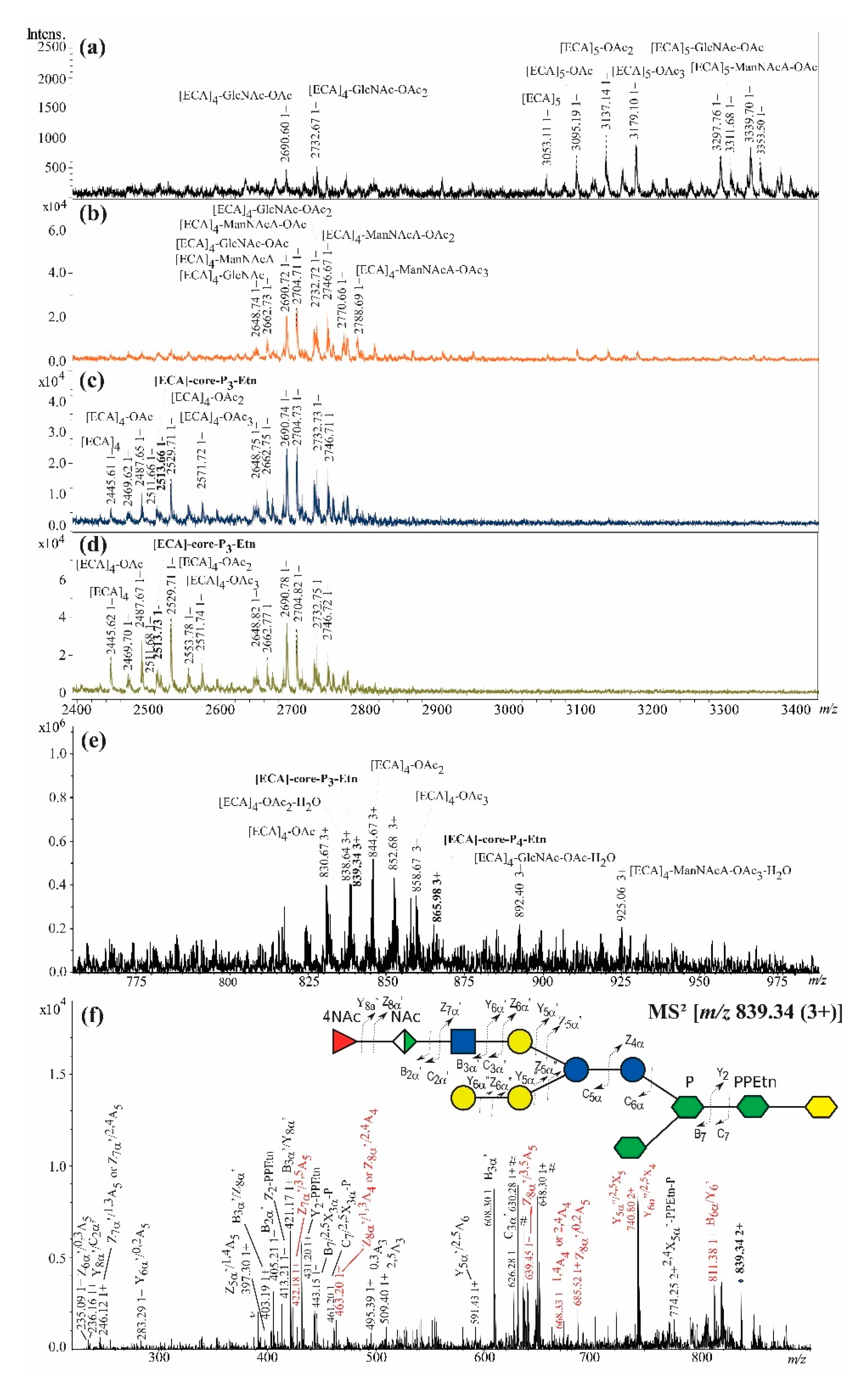
2.2. Rough Strains of E. coli R1, R2, and R4 Synthesize ECALPS
2.3. ECALPS is Absent in Rough E. coli R3 Lipooligosaccharide Preparation
3. Discussion
4. Materials and Methods
4.1. Bacteria and Culture Conditions
4.2. Preparation of LOS and Oligosaccharides
4.3. Electrospray Mass Spectrometry
4.4. Matrix-Assisted Laser Desorption/Ionization-Time of Flight Mass Spectrometry
4.5. NMR Spectroscopy
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| ECA | Enterobacterial common antigen |
| ECAPG | Phosphatidylglycerol-linked ECA |
| ECACYC | Cyclic ECA |
| ECALPS | Lipopolysaccharide-associated ECA |
| LPS | Lipopolysaccharide |
| LOS | Lipooligosaccharide |
| O-PS | O-specific polysaccharide |
| OS | Oligosaccharide |
| P | Phosphate group |
| PP | Pyrophosphate group |
| PPEtn | Pyrophosphorylethanolamine |
| OAc | O-acetyl group |
| ESI-IT MS | Electrospray ionization-ion trap mass spectrometry |
| MALDI-TOF MS | Matrix-assisted laser-desorption/ionization-time of flight mass spectrometry |
| NMR | Nuclear magnetic resonance |
| MS | Mass spectrometry |
References
- Makela, P.H.; Mayer, H. Enterobacterial common antigen. Bacteriol. Rev. 1976, 40, 591–632. [Google Scholar] [CrossRef]
- Lugowski, C.; Romanowska, E.; Kenne, L.; Lindberg, B. Identification of a trisaccharide repeating-unit in the enterobacterial common-antigen. Carbohyd. Res. 1983, 118, 173–181. [Google Scholar] [CrossRef]
- Valvano, M.A. Chapter 4—Genetics and Biosynthesis of Lipopolysaccharide. In Molecular Medical Microbiology, 2nd ed.; Tang, Y.-W., Sussman, M., Liu, D., Poxton, I., Schwartzman, J., Eds.; Academic Press: Boston, MA, USA, 2015; pp. 55–89. [Google Scholar] [CrossRef]
- Mitchell, A.M.; Srikumar, T.; Silhavy, T.J. Cyclic Enterobacterial Common Antigen Maintains the Outer Membrane Permeability Barrier of Escherichia coli in a Manner Controlled by YhdP. mBio 2018, 9. [Google Scholar] [CrossRef] [Green Version]
- Kunin, C.M.; Beard, M.V.; Halmagyi, N.E. Evidence for a Common Hapten Associated with Endotoxin Fractions of E. coli and other Enterobacteriaceae. Expe. Biol. Med. 1962, 111, 160–166. [Google Scholar] [CrossRef]
- Kunin, C.M.; Beard, M.V. Serological Studies of O Antigens of Escherichia Coli by Means of the Hemagglutination Test. J. Bacteriol. 1963, 85, 541–548. [Google Scholar] [CrossRef] [Green Version]
- Gozdziewicz, T.K.; Lugowski, C.; Lukasiewicz, J. First evidence for a covalent linkage between enterobacterial common antigen and lipopolysaccharide in Shigella sonnei phase II ECALPS. J. Biol. Chem. 2014, 289, 2745–2754. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gozdziewicz, T.K.; Lugowski, C.; Lukasiewicz, J. First evidence for a covalent linkage between enterobacterial common antigen and lipopolysaccharide in Shigella sonnei phase II ECALPS. J. Biol. Chem. 2018, 293, 11652–11653. [Google Scholar] [CrossRef] [PubMed]
- Vinogradov, E.V.; Van Der Drift, K.; Thomas-Oates, J.E.; Meshkov, S.; Brade, H.; Holst, O. The structures of the carbohydrate backbones of the lipopolysaccharides from Escherichia coli rough mutants F470 (R1 core type) and F576 (R2 core type). Eur. J. Biochem. 1999, 261, 629–639. [Google Scholar] [CrossRef] [PubMed]
- Holst, O. The structures of core regions from enterobacterial lipopolysaccharides—An update. FEMS Microbiol. Lett. 2007, 271, 3–11. [Google Scholar] [CrossRef] [PubMed]
- Muller-Loennies, S.; Lindner, B.; Brade, H. Structural analysis of deacylated lipopolysaccharide of Escherichia coli strains 2513 (R4 core-type) and F653 (R3 core-type). Eur. J. Biochem. 2002, 269, 5982–5991. [Google Scholar] [CrossRef] [PubMed]
- Kunin, C.M. Separation, Characterization, and Biological Significance of a Common Antigen in Enterobacteriaceae. J. Exp. Med. 1963, 118, 565–586. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rinno, J.; Golecki, J.R.; Mayer, H. Localization of enterobacterial common antigen:immunogenic and nonimmunogenic enterobacterial common antigen-containing Escherichia coli. J. Bacteriol. 1980, 141, 814–821. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jensen, S.O.; Reeves, P.R. Deletion of the Escherichia coli O14:K7 O antigen gene cluster. Can. J. Microbiol. 2004, 50, 299–302. [Google Scholar] [CrossRef] [PubMed]
- Kuhn, H.M.; Meier-Dieter, U.; Mayer, H. ECA, the enterobacterial common antigen. FEMS Microbiol. Rev. 1988, 4, 195–222. [Google Scholar] [CrossRef]
- Whang, H.Y.; Mayer, H.; Schmidt, G.; Neter, E. Immunogenicity of the common enterobacterial antigen produced by smooth and rough strains. Infect. Immun. 1972, 6, 533–539. [Google Scholar] [CrossRef] [Green Version]
- Mayer, H.; Schmidt, G.; Whang, H.Y.; Neter, E. Biochemical basis of the immunogenicity of the common enterobacterial antigen. Infect. Immun. 1972, 6, 540–544. [Google Scholar] [CrossRef] [Green Version]
- Duda, K.A.; Duda, K.T.; Beczala, A.; Kasperkiewicz, K.; Radziejewska-Lebrecht, J.; Skurnik, M. ECA-immunogenicity of Proteus mirabilis strains. Arch. Immunol. Ther. Exp. (Warsz) 2009, 57, 147–151. [Google Scholar] [CrossRef] [Green Version]
- Romanowska, E.; Katzenellenbogen, E.; Lugowski, C.; Gamian, A.; Bogulska, M. Immunochemical characteristics of Shigella sonnei and serotype 6 Shigella flexneri lipopolysaccharides and enterobacterial common antigen. Arch. Immunol. Ther. Exp. (Warsz) 1978, 26, 249–254. [Google Scholar]
- Radziejewska-Lebrecht, J.; Kasperkiewicz, K.; Skurnik, M.; Brade, L.; Steinmetz, I.; Swierzko, A.S.; Muszynski, A. ECA-antibodies in antisera against R mutants of Yersinia enterocolitica O:3. Adv. Exp. Med. Biol. 2003, 529, 215–218. [Google Scholar] [CrossRef]
- Rabsztyn, K.; Kasperkiewicz, K.; Duda, K.A.; Li, C.M.; Lukasik, M.; Radziejewska-Lebrecht, J.; Skurnik, M. Characterization of anti-ECA antibodies in rabbit antiserum against rough Yersinia enterocolitica O:3. Biochemistry (Mosc) 2011, 76, 832–839. [Google Scholar] [CrossRef] [Green Version]
- Muszynski, A.; Rabsztyn, K.; Knapska, K.; Duda, K.A.; Duda-Grychtol, K.; Kasperkiewicz, K.; Radziejewska-Lebrecht, J.; Holst, O.; Skurnik, M. Enterobacterial common antigen and O-specific polysaccharide coexist in the lipopolysaccharide of Yersinia enterocolitica serotype O:3. Microbiology 2013, 159, 1782–1793. [Google Scholar] [CrossRef] [PubMed]
- Noszczynska, M.; Kasperkiewicz, K.; Duda, K.A.; Podhorodecka, J.; Rabsztyn, K.; Gwizdala, K.; Swierzko, A.S.; Radziejewska-Lebrecht, J.; Holst, O.; Skurnik, M. Serological characterization of the enterobacterial common antigen substitution of the lipopolysaccharide of Yersinia enterocolitica O:3. Microbiology 2015, 161, 219–227. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Islam, S.T.; Lam, J.S. Synthesis of bacterial polysaccharides via the Wzx/Wzy-dependent pathway. Can. J. Microbiol. 2014, 60, 697–716. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kalynych, S.; Morona, R.; Cygler, M. Progress in understanding the assembly process of bacterial O-antigen. FEMS Microbiol. Rev. 2014, 38, 1048–1065. [Google Scholar] [CrossRef] [Green Version]
- Whitfield, C. Biosynthesis of lipopolysaccharide O antigens. Trends Microbiol. 1995, 3, 178–185. [Google Scholar] [CrossRef]
- Varki, A.; Cummings, R.D.; Aebi, M.; Packer, N.H.; Seeberger, P.H.; Esko, J.D.; Stanley, P.; Hart, G.; Darvill, A.; Kinoshita, T.; et al. Symbol Nomenclature for Graphical Representations of Glycans. Glycobiology 2015, 25, 1323–1324. [Google Scholar] [CrossRef] [Green Version]
- Dell, A.; Oates, J.; Lugowski, C.; Romanowska, E.; Kenne, L.; Lindberg, B. The enterobacterial common-antigen, a cyclic polysaccharide. Carbohydr. Res. 1984, 133, 95–104. [Google Scholar] [CrossRef]
- Domon, B.; Costello, C.E. Structure elucidation of glycosphingolipids and gangliosides using high-performance tandem mass spectrometry. Biochemistry 1988, 27, 1534–1543. [Google Scholar] [CrossRef]
- Kuhn, H.M.; Neter, E.; Mayer, H. Modification of the lipid moiety of the enterobacterial common antigen by the “Pseudomonas factor”. Infect. Immun. 1983, 40, 696–700. [Google Scholar] [CrossRef] [Green Version]
- Schmidt, G.; Jann, B.; Jann, K. Genetic and immunochemical studies on Escherichia coli O14:K7:H. Eur. J. Biochem. 1974, 42, 303–309. [Google Scholar] [CrossRef]
- Schmidt, G.; Mannel, D.; Mayer, H.; Whang, H.Y.; Neter, E. Role of a lipopolysaccharide gene for immunogenicity of the enterobacterial common antigen. J. Bacteriol. 1976, 126, 579–586. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schmidt, G.; Mayer, H.; Makela, P.H. Presence of rfe genes in Escherichia coli: Their participation in biosynthesis of O antigen and enterobacterial common antigen. J. Bacteriol. 1976, 127, 755–762. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rick, P.D.; Wolski, S.; Barr, K.; Ward, S.; Ramsay-Sharer, L. Accumulation of a lipid-linked intermediate involved in enterobacterial common antigen synthesis in Salmonella typhimurium mutants lacking dTDP-glucose pyrophosphorylase. J. Bacteriol. 1988, 170, 4008–4014. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, B.; Furevi, A.; Perepelov, A.V.; Guo, X.; Cao, H.; Wang, Q.; Reeves, P.R.; Knirel, Y.A.; Wang, L.; Widmalm, G. Structure and genetics of Escherichia coli O antigens. FEMS Microbiol. Rev. 2019. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heinrichs, D.E.; Yethon, J.A.; Amor, P.A.; Whitfield, C. The assembly system for the outer core portion of R1- and R4-type lipopolysaccharides of Escherichia coli. The R1 core-specific beta-glucosyltransferase provides a novel attachment site for O-polysaccharides. J. Biol. Chem. 1998, 273, 29497–29505. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heinrichs, D.E.; Monteiro, M.A.; Perry, M.B.; Whitfield, C. The assembly system for the lipopolysaccharide R2 core-type of Escherichia coli is a hybrid of those found in Escherichia coli K-12 and Salmonella enterica. Structure and function of the R2 WaaK and WaaL homologs. J. Biol. Chem. 1998, 273, 8849–8859. [Google Scholar] [CrossRef] [Green Version]
- Gamian, A.; Romanowska, E.; Ulrich, J.; Defaye, J. The structure of the sialic acid-containing Escherichia coli O104 O-specific polysaccharide and its linkage to the core region in lipopolysaccharide. Carbohydr. Res. 1992, 236, 195–208. [Google Scholar] [CrossRef]
- Meredith, T.C.; Mamat, U.; Kaczynski, Z.; Lindner, B.; Holst, O.; Woodard, R.W. Modification of lipopolysaccharide with colanic acid (M-antigen) repeats in Escherichia coli. J. Biol. Chem. 2007, 282, 7790–7798. [Google Scholar] [CrossRef] [Green Version]
- Westphal, O.; Jann, K. Bacterial lipopolysacharides: Extraction with Phenol-Water and further applications of the procedure. Methods Carbohydr. Chem. 1965, 5, 83–89. [Google Scholar]
- Ceroni, A.; Maass, K.; Geyer, H.; Geyer, R.; Dell, A.; Haslam, S.M. GlycoWorkbench: A tool for the computer-assisted annotation of mass spectra of glycans. J. Proteome. Res. 2008, 7, 1650–1659. [Google Scholar] [CrossRef] [Green Version]
- Lee, W.; Tonelli, M.; Markley, J.L. NMRFAM-SPARKY: Enhanced software for biomolecular NMR spectroscopy. Bioinformatics 2015, 31, 1325–1327. [Google Scholar] [CrossRef] [PubMed] [Green Version]

 Fucose;
Fucose;  Mannuronic acid;
Mannuronic acid;  N-acetylglucosamine;
N-acetylglucosamine;  Glucose;
Glucose;  Galactose;
Galactose;  l,d-manno-Heptose;
l,d-manno-Heptose;  Kdo; NAc, N-acetyl group [27]. The interpretation of ions is shown in Table 1. The most informative ions are colored in red. The mark # stands for non-interpreted ions.
Fucose; Mannuronic acid; N-acetylglucosamine; Glucose; Galactose; l,d-manno-Heptose; Kdo; NAc, N-acetyl group [27]. The interpretation of ions is shown in Table 1. The most informative ions are colored in red. The mark # stands for non-interpreted ions.
Kdo; NAc, N-acetyl group [27]. The interpretation of ions is shown in Table 1. The most informative ions are colored in red. The mark # stands for non-interpreted ions.
Fucose; Mannuronic acid; N-acetylglucosamine; Glucose; Galactose; l,d-manno-Heptose; Kdo; NAc, N-acetyl group [27]. The interpretation of ions is shown in Table 1. The most informative ions are colored in red. The mark # stands for non-interpreted ions.





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Oligosaccharide Structure | Calculated Monoisotopic Mass (Da) | Observed Ion (m/z) | Calculated Ion (m/z) | Interpretation of the Ion |
|---|---|---|---|---|
| Fraction 3 (negative mode) | ||||
| [ECA]3-Glc3-Gal2-Hep3-Kdo-P2 | 3606.12 | 1201.15/nd | 1201.03/1802.05 | [M-3H]3−/[M-2H]2− |
| [ECA]3-Glc3-Gal2-Hep3-Kdo-P3-Etn | 3729.13 | 1242.15/nd | 1242.04/1863.56 | [M-3H]3−/[M-2H]2− |
| [ECA]3-Glc3-Gal2-Hep3-Kdo-P3-Etn | 3729.13 | 1235.81/nd | 1236.03/1854.55 | [M-H2O-3H]3−/[M-H2O-2H]2− |
| [ECA]3-Glc3-Gal2-Hep3-Kdo-P3-Etn | 3729.13 | 1249.47/nd | 1249.36/1874.54 | [M-3H, Na]3−/[M-2H, Na]2− |
| [ECA]3-Glc3-Gal2-Hep3-Kdo-P4-Etn | 3809.10 | 1268.81/nd | 1268.69/1903.54 | [M-3H]3−/[M-2H]2− |
| [ECA]3-Glc3-Gal2-Hep3-Kdo-P4-Etn | 3809.10 | 1276.45/nd | 1276.02/1914.53 | [M-3H, Na]3−/[M-2H, Na]2− |
| [ECA]4-Glc3-Gal2-Hep3-Kdo-P2 | 4213.35 | 1403.53/nd | 1403.44/2105.67 | [M-3H]3−/[M-2H]2− |
| [ECA]4-Glc3-Gal2-Hep3-Kdo-P3-Etn | 4336.35 | 1444.53/nd | 1444.44/2167.17 | [M-3H]3−/[M-2H]2− |
| Fraction 4 (negative mode) | ||||
| [ECA]2-Glc3-Gal2-Hep3-Kdo-P2 | 2998.90 | 998.63/1498.41 | 998.63/1498.44 | [M-3H]3−/[M-2H]2− |
| [ECA]2-Glc3-Gal2-Hep3-Kdo-P2 | 2998.90 | 992.63/1489.39 | 992.62/1489.44 | [M-H2O-3H]3−/[M-H2O-2H]2− |
| [ECA]2-Glc3-Gal2-Hep3-Kdo-P3-Etn | 3121.91 | 1039.63/1559.91 | 1039.63/1559.95 | [M-3H]3−/[M-2H]2− |
| [ECA]2-Glc3-Gal2-Hep3-Kdo-P3-Etn | 3121.91 | 1033.63/1550.88 | 1033.62/1550.94 | [M-H2O-3H]3−/[M-H2O-2H]2− |
| [ECA]2-Glc3-Gal2-Hep3-Kdo-P4-Etn | 3201.88 | 1066.28/nd | 1066.28/1599.93 | [M-3H]3−/[M-2H]2− |
| [ECA]3-Glc3-Gal2-Hep3-Kdo-P2 | 3606.12 | 1201.02/nd | 1201.03/1802.05 | [M-3H]3−/[M-2H]2− |
| [ECA]3-Glc3-Gal2-Hep3-Kdo-P3-Etn | 3729.13 | 1242.00/nd | 1242.04/1863.56 | [M-3H]3-/[M-2H]2− |
| Fraction 5 (negative or positive mode) | ||||
| [ECA]-Glc3-Gal2-Hep3-Kdo-P2 | 2391.68 | 796.25/1194.81 | 796.22/1194.83 | [M-3H]3−/[M-2H]2− |
| [ECA]-Glc3-Gal2-Hep3-Kdo-P2 | 2391.68 | 790.22/1185.80 | 790.21/1185.82 | [M-H2O-3H]3−/[M-H2O-2H]2− |
| [ECA]-Glc3-Gal2-Hep3-Kdo-P3 | 2471.64 | 822.89/1234.81 | 822.87/1234.82 | [M-3H]3−/[M-2H]2− |
| [ECA]-Glc3-Gal2-Hep3-Kdo-P3-Etn | 2514.69 | 837.25/1256.31 | 837.22/1256.34 | [M-3H]3−/[M-2H]2− |
| [ECA]-Glc3-Gal2-Hep3-Kdo-P3-Etn | 2514.69 | 831.23/1247.28 | 831.22/1247.33 | [M-H2O-3H]3−/[M-H2O-2H]2− |
| [ECA]-Glc3-Gal2-Hep3-Kdo-P3-Etn | 2514.69 | 844.56/1267.32 | 844.55/1267.32 | [M-3H, Na]3−/[M-2H, Na]2− |
| [ECA]-Glc3-Gal2-Hep3-Kdo-GlcN-P2 | 2552.75 | nd/1275.31 | 849.91/1275.37 | [M-3H]3−/[M-2H]2− |
| [ECA]-Glc3-Gal2-Hep3-Kdo-P4-Etn | 2594.65 | 863.88/1296.32 | 863.88/1296.32 | [M-3H]3−/[M-2H]2− |
| [ECA]2-Glc3-Gal2-Hep3-Kdo-P2 | 2998.90 | 998.63/1498.40 | 998.63/1498.44 | [M-3H]3−/[M-2H]2− |
| [ECA]2-Glc3-Gal2-Hep3-Kdo-P3-Etn | 3121.91 | 1039.59/nd | 1039.63/1559.95 | [M-3H]3−/[M-2H]2− |
| [ECA]-Glc3-Gal2-Hep3-Kdo-P3-Etn | 2514.69 | 839.25/1258.34 * | 839.24/1258.35 | [M+3H]3+/[M+2H]2+ |
| [ECA]-Glc3-Gal2-Hep3-Kdo-P3-Etn | 2514.69 | 833.25/nd | 833.23/1249.34 | [M-H2O+3H]3+/[M-H2O+2H]2+ |
| [ECA]-Glc3-Gal2-Hep3-Kdo-P3-Etn | 2514.69 | 846.57/1269.34 * | 846.56/1269.34 | [M+3H, Na]3+/[M+2H, Na]2+ |
| [ECA]-Glc3-Gal2-Hep3-Kdo-GlcN-P2 | 2552.75 | 851.92/1277.31 * | 851.92/1277.38 | [M+3H]3+/[M+2H]2+ |
| [ECA]-Glc3-Gal2-Hep3-Kdo-GlcN-P2-Etn | 2617.76 | 873.56/nd | 873.59/1309.89 | [M+3H, Na]3+/[M+2H, Na]2+ |
| [ECA]-Glc3-Gal2-Hep3-Kdo-P4-Etn | 2594.65 | 865.90/1298.32 * | 865.89/1298.33 | [M+3H]3+/[M+2H]2+ |
| Residue | Chemical Shifts (ppm) | Selected Inter-Residue NOE and 3JH,C Connectivities | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| H1 C1 | H2,(H3′) C2 | H3 C3 | H4 C4 | H5 C5 | H6,H6′ C6 | H7, H7′ C7 | H8,H8′ C8 | H1/C1 Connectivities to | Inter-Residue Atom/Residue | ||
| A | →5)-α-Kdop | nd | (2.27) 96.2 | 1.91 34.2 | 4.13 66.4 | 4.18 73.4 | 3.70 69.9 | 3.81 73.0 | 3.47, 3.94 64.9 | ||
| B | →3)-l-α-d-Hepp4PPEtn-(1→ | 5.21 100.2 | 4.02 71.8 | 4.09 78.6 | 4.64 72.4 | 4.24 72.1 | 4.11 69.4 | 3.73 63.9 | 4.18 | H5 of A | |
| C | →3,7)-l-α-d-Hepp4P-(1→ | 5.11 103.7 | 4.40 70.6 | 4.12 79.9 | 4.42 69.5 | 3.81 73.3 | 4.23 68.8 | 3.61, 3.77 68.5 | 4.09 78.6 | C3, H3 of B | |
| D | l-α-d-Hepp-(1→ | 5.00 100.3 | 3.94 70.9 | 3.89 71.5 | 3.86 67.0 | 3.63 72.0 | 4.05 69.6 | 3.64, 3.75 63.7 | 3.61, 3.77 a 68.5 | C7, H7,7′ of C | |
| E | →3)-α-d-Glcp-(1→ | 5.21 102.1 | 3.68 71.2 | 4.09 76.8 | 3.80 71.3 | 3.92 73.2 | 3.81, 3.92 60.7 | 4.12 a | H3 of C | ||
| F | →2,3)-α-d-Glcp-(1→ | 5.81 95.4 | 3.88 73.5 | 4.18 78.7 | 3.58 68.8 | 4.11 72.1 | 3.79, 4.00 61.1 | 4.09 76.8 | C3, H3 of E | ||
| G | →2)-α-d-Galp-(1→ | 5.63 92.1 | 4.00 73.2 | 4.20 68.9 | 3.99 70.9 | 4.15 72.2 | 3.76 62.1 | 3.88 a | H2 of F | ||
| H | α-d-Galp-(1→ | 5.32 96.6 | 3.86 69.2 | 3.96 70.2 | 4.00 70.2 | 4.14 72.1 | 3.75 62.1 | 4.00 | H2 of G | ||
| I | →3)-β-d-Glcp-(1→ | 4.74 103.2 | 3.39 73.7 | 3.70 85.5 | 3.50 69.0 | 3.45 76.3 | 3.75, 3.91 61.5 | 4.18 78.7 | C3, H3 of F | ||
| I’ b | β-d-Glcp-(1→ | 4.76 103.2 | 3.34 74.0 | 3.53 76.7 | 3.43 70.5 | 3.45 76.6 | 3.92, 3.74 61.5 | 4.18 78.7 | C3, H3 of F | ||
| J | →4)-β-d-GlcpNAc-(1→ | 4.79 102.4 | 3.76 56.4 | 3.75 72.8 | 3.70 79.6 | 3.55 75.2 | 3.73, 3.86 61.0 | 3.70 85.5 | C3, H3 of I | ||
| K | →4)-β-d-ManpNAcA-(1→ | 4.94 99.8 | 4.51 54.3 | 4.08 73.3 | 3.84 74.9 | 3.88 77.2 | - 175.1 | 3.70 a 79.6 | C4, H4 of J | ||
| L | α-d-Fucp4NAc-(1→ | 5.36 99.7 | 3.65 69.3 | 3.97 69.2 | 4.21 54.7 | 4.19 66.7 | 1.06 16.3 | 3.84 | H4 of K | ||
| PPEtn | 4.21 63.2 | 3.30 40.8 | |||||||||
| Oligosaccharide Structure | Calculated Monoisotopic Mass (Da) | Observed Ion (m/z) | Calculated Ion (m/z) | Interpretation of the Ion |
|---|---|---|---|---|
| Fraction 3 (negative mode) | ||||
| [ECA]5 | 3054.12 | nd/1526.06 | 1017.03/1526.05 | [M-3H]3−/[M-2H]2− |
| [ECA]5-OAc | 3096.13 | nd/1547.06 | 1031.04/1547.06 | [M-3H]3−/[M-2H]2− |
| [ECA]3-Glc3-GlcNAc-Gal-Hep3-Kdo-P3-Etn | 3770.16 | 1255.69/nd | 1255.71/1884.07 | [M-3H]3−/[M-2H]2− |
| [ECA]3-Glc3-GlcNAc-Gal-Hep3-Kdo-P4-Etn | 3850.12 | 1282.34/nd | 1282.37/1924.06 | [M-3H]3−/[M-2H]2− |
| [ECA]3-Glc3-GlcNAc-Gal-Hep3-Kdo-P4-Etn2 | 3893.17 | 1290.66/nd | 1290.71/1936.57 | [M-H2O-3H]3−/[M-H2O-2H]2− |
| [ECA]3-Glc3-GlcNAc-Gal-Hep3-Kdo-P4-Etn2 | 3893.17 | 1296.69/nd | 1296.72/1945.58 | [M-3H]3−/[M-2H]2− |
| [ECA]3-Glc3-GlcNAc-Gal-Hep3-Kdo-P4-Etn2 | 3893.17 | 1304.03/nd | 1304.04/1956.56 | [M-3H, Na]3−/[M-2H, Na]2− |
| [ECA]3-Glc3-GlcNAc-Gal-Hep3-Kdo-P5-Etn2 | 3973.13 | 1323.38/nd | 1323.37/1985.56 | [M-3H]3−/[M-2H]2− |
| [ECA]4-Glc3-GlcNAc-Gal-Hep3-Kdo-P2 | 4254.37 | 1417.06/nd | 1417.12/2126.18 | [M-3H]3−/[M-2H]2− |
| [ECA]4-Glc3-GlcNAc-Gal-Hep3-Kdo-P3-Etn | 4377.38 | 1458.08/nd | 1458.12/2187.68 | [M-3H]3−/[M-2H]2− |
| [ECA]4-Glc3-GlcNAc-Gal-Hep3-Kdo-P4-Etn2 | 4500.39 | 1499.07/nd | 1499.12/2249.19 | [M-3H]3−/[M-2H]2− |
| Fraction 4 (negative mode) | ||||
| [ECA]4 | 2446.90 | nd/1222.44 | 814.63/1222.44 | [M-3H]3−/[M-2H]2− |
| [ECA]2-Glc3-GlcNAc-Gal-Hep3-Kdo-P3-Etn | 3162.94 | 1053.30/1580.47 | 1053.30/1580.46 | [M-3H]3−/[M-2H]2− |
| [ECA]2-Glc3-GlcNAc-Gal-Hep3-Kdo-P4-Etn | 3242.90 | 1079.66/1620.44 | 1079.96/1620.44 | [M-3H]3−/[M-2H]2− |
| [ECA]2-Glc3-GlcNAc-Gal-Hep3-Kdo-P4-Etn2 | 3285.94 | 1094.31/1641.94 | 1094.31/1641.96 | [M-3H]3−/[M-2H]2− |
| [ECA]2-Glc3-GlcNAc-Gal-Hep3-Kdo-P4-Etn2 | 3285.94 | 1101.94/1652.91 | 1101.63/1652.95 | [M-3H, Na]3−/[M-2H, Na]2− |
| [ECA]2-Glc3-GlcNAc-Gal-Hep3-Kdo-P5-Etn2 | 3365.91 | 1120.94/1681.90 | 1120.96/1681.95 | [M-3H]3−/[M-2H]2− |
| [ECA]3-Glc3-GlcNAc-Gal-Hep3-Kdo-P3 | 3727.12 | 1241.39/nd | 1241.36/1862.55 | [M-3H]3−/[M-2H]2− |
| [ECA]3-Glc3-GlcNAc-Gal-Hep3-Kdo-P3-Etn | 3770.16 | 1255.03/nd | 1255.71/1884.07 | [M-3H]3−/[M-2H]2− |
| [ECA]3-Glc3-GlcNAc-Gal-Hep3-Kdo-P4-Etn | 3850.12 | 1282.38/nd | 1282.37/1924.06 | [M-3H]3−/[M-2H]2− |
| [ECA]3-Glc3-GlcNAc-Gal-Hep3-Kdo-P4-Etn2 | 3893.17 | 1296.67/nd | 1296.72/1945.58 | [M-3H]3−/[M-2H]2− |
| Fraction 5 (negative/positive modes) | ||||
| Glc3-GlcNAc-Gal-Hep3-Kdo-P3 | 1905.45 | nd/951.73 | 634.14/951.72 | [M-3H]3−/[M-2H]2− |
| Glc3-GlcNAc-Gal-Hep3-Kdo-P3-Etn | 1948.49 | nd/973.27 | 648.49/973.24 | [M-3H]3−/[M-2H]2− |
| Glc3-GlcNAc-Gal-Hep3-Kdo-P4-Etn | 2028.46 | nd/1004.22 | 669.14/1004.21 | [M-H2O-3H]3−/[M-H2O-2H]2− |
| Glc3-GlcNAc-Gal-Hep3-Kdo-P4-Etn | 2028.46 | nd/1013.23 | 675.15/1013.22 | [M-3H]3−/[M-2H]2− |
| Glc3-GlcNAc-Gal-Hep3-Kdo-P4-Etn2 | 2071.50 | nd/1025.73 | 683.49/1025.73 | [M-H2O-3H]3−/[M-H2O-2H]2− |
| Glc3-GlcNAc-Gal-Hep3-Kdo-P4-Etn2 | 2071.50 | nd/1034.74 | 689.49/1034.74 | [M-3H]3−/[M-2H]2− |
| [ECA]-Glc3-GlcNAc-Gal-Hep3-Kdo-P3-Etn | 2555.71 | 850.88/1276.83 | 850.90/1276.85 | [M-3H]3−/[M-2H]2− |
| [ECA]-Glc3-GlcNAc-Gal-Hep3-Kdo-P4-Etn | 2635.68 | 871.57/1307.82 | 871.55/1307.82 | [M-H2O-3H]3−/[M-H2O-2H]2− |
| [ECA]-Glc3-GlcNAc-Gal-Hep3-Kdo-P4-Etn | 2635.68 | 877.63/1316.83 | 877.55/1316.83 | [M-3H]3−/[M-2H]2− |
| [ECA]-Glc3-GlcNAc-Gal-Hep3-Kdo-P4-Etn2 | 2678.72 | 885.92/1329.31 | 885.89/1329.35 | [M-H2O-3H]3-/[M-H2O-2H]2− |
| [ECA]-Glc3-GlcNAc-Gal-Hep3-Kdo-P4-Etn2 | 2678.72 | 891.88/1338.31 | 891.90/1338.35 | [M-3H]3−/[M-2H]2− |
| [ECA]-Glc3-GlcNAc-Gal-Hep3-Kdo-P4-Etn2 | 2678.72 | 899.25/1349.31 | 899.22/1349.34 | [M−3H, Na]3−/[M−2H, Na]2− |
| [ECA]-Glc3-GlcNAc-Gal-Hep3-Kdo-P5-Etn2 | 2758.69 | 912.56/1369.29 | 912.55/1369.33 | [M-H2O-3H]3−/[M-H2O-2H]2− |
| [ECA]-Glc3-GlcNAc-Gal-Hep3-Kdo-P5-Etn2 | 2758.69 | 918.53/1378.31 | 918.56/1378.34 | [M-3H]3−/[M-2H]2− |
| [ECA]-Glc3-GlcNAc-Gal-Hep3-Kdo-P5-Etn2 | 2758.69 | 925.89/1389.31 | 925.88/1389.32 | [M-3H, Na]3−/[M-2H, Na]2− |
| Glc3-GlcNAc-Gal-Hep3-Kdo-P3-Etn | 1948.49 | nd/975.20 | 650.50/975.25 | [M+3H]3+/[M+2H]2+ |
| Glc3-GlcNAc-Gal-Hep3-Kdo-P3-Etn | 1948.49 | nd/986.18 | 657.83/986.24 | [M+3H, Na]3+/[M+2H, Na]2+ |
| Glc3-GlcNAc-Gal-Hep3-Kdo-P4-Etn2 | 2071.50 | nd/1027.65 | 685.50/1027.75 | [M-H2O+3H]3+/[M-H2O+2H]2+ |
| Glc3-GlcNAc-Gal-Hep3-Kdo-P4-Etn2 | 2071.50 | nd/1036.74 | 691.51/1036.76 | [M+3H]3+/[M+2H]2+ |
| [ECA]-Glc3-GlcNAc-Gal-Hep3-Kdo-P3-Etn | 2555.71 | 852.88/1278.88 | 852.91/1278.86 | [M+3H]3+/[M+2H]2+ |
| [ECA]-Glc3-GlcNAc-Gal-Hep3-Kdo-P3-Etn | 2555.71 | 860.21/1289.84 | 860.24/1289.85 | [M+3H, Na]3+/[M+2H, Na]2+ |
| [ECA]-Glc3-GlcNAc-Gal-Hep3-Kdo-P3-Etn | 2555.71 | nd/1300.75 | 867.57/1300.85 | [M+3H, 2Na]3+/[M+2H, 2Na]2+ |
| [ECA]-Glc3-GlcNAc-Gal-Hep3-Kdo-P4-Etn | 2635.68 | 879.50/1318.75 | 879.57/1318.85 | [M+3H]3+/[M+2H]2+ |
| [ECA]-Glc3-GlcNAc-Gal-Hep3-Kdo-P4-Etn2 | 2678.72 | 887.75/1331.28 | 887.91/1331.36 | [M-H2O+3H]3+/[M-H2O+2H]2+ |
| [ECA]-Glc3-GlcNAc-Gal-Hep3-Kdo-P4-Etn2 | 2678.72 | 893.75/1340.34 | 893.91/1340.37 | [M+3H]3+/[M+2H]2+ |
| [ECA]-Glc3-GlcNAc-Gal-Hep3-Kdo-P4-Etn2 | 2678.72 | 901.13/1351.34 | 901.24/1351.35 | [M+3H, Na]3+/[M+2H, Na]2+ |
| [ECA]-Glc3-GlcNAc-Gal-Hep3-Kdo-P5-Etn2 | 2758.69 | 920.76/1380.33 | 920.57/1380.35 | [M+3H]3+/[M+2H]2+ |
| Oligosaccharide Structure | Calculated Monoisotopic Mass (Da) | Observed Ion (m/z) | Calculated Ion (m/z) | Interpretation of the Ion |
|---|---|---|---|---|
| Fraction 25 (negative mode) | ||||
| [ECA]5-OAc | 3096.13 | 3095.19 | 3095.13 | [M-H]− |
| [ECA]5 | 3054.12 | 3053.11 | 3053.12 | [M-H]− |
| [ECA]5-OAc2 | 3138.14 | 3137.14 | 3137.14 | [M-H]− |
| [ECA]5-OAc3 | 3180.15 | 3179.10 | 3179.15 | [M-H]− |
| [ECA]5-GlcNAc-OAc | 3299.21 | 3297.76 | 3298.21 | [M-H]− |
| [ECA]5-ManNAcA-OAc | 3313.19 | 3311.68 | 3312.18 | [M-H]− |
| [ECA]5-ManNAcA-OAc2 | 3355.20 | 3353.50 | 3354.20 | [M-H]− |
| [ECA]4-GlcNAc-OAc2 | 2734.00 | 2732.67 | 2732.99 | [M-H]− |
| [ECA]4-GlcNAc-OAc | 2691.99 | 2690.60 | 2690.98 | [M-H]− |
| Fraction 26 (negative mode) | ||||
| [ECA]4-ManNAcA-OAc3 | 2789.99 | 2788.69 | 2788.98 | [M-H]− |
| [ECA]4-ManNAcA-OAc3 | 2789.99 | 2770.66 | 2770.97 | [M-H2O-H]− |
| [ECA]4-ManNAcA-OAc2 | 2747.98 | 2746.67 | 2746.97 | [M-H]− |
| [ECA]4-GlcNAc-OAc2 | 2734.00 | 2732.72 | 2732.99 | [M-H]− |
| [ECA]4-ManNAcA-OAc | 2705.97 | 2704.71 | 2704.96 | [M-H]− |
| [ECA]4-GlcNAc-OAc | 2691.99 | 2690.72 | 2690.98 | [M-H]− |
| [ECA]4-ManNAcA | 2663.96 | 2662.73 | 2662.95 | [M-H]− |
| [ECA]4-GlcNAc | 2649.98 | 2648.74 | 2648.97 | [M-H]− |
| Fraction 27 (negative mode) | ||||
| [ECA]4-ManNAcA-OAc2 | 2747.98 | 2746.71 | 2746.97 | [M-H]− |
| [ECA]4-GlcNAc-OAc2 | 2734.00 | 2732.73 | 2732.99 | [M-H]− |
| [ECA]4-GlcNAc-OAc | 2691.99 | 2690.74 | 2690.98 | [M-H]− |
| [ECA]4-ManNAcA | 2663.96 | 2662.75 | 2662.95 | [M-H]− |
| [ECA]4-GlcNAc | 2649.98 | 2648.75 | 2648.97 | [M-H]− |
| [ECA]4-OAc3 | 2572.93 | 2571.72 | 2571.92 | [M-H]− |
| [ECA]4-OAc2 | 2530.92 | 2529.71 | 2529.91 | [M-H]− |
| [ECA]-Gal3-Glc2-Hep3-Kdo-P3-Etn | 2514.69 | 2513.66 | 2513.68 | [M-H]− |
| [ECA]4-OAc | 2488.91 | 2487.65 | 2487.90 | [M-H]− |
| [ECA]4-OAc | 2470.90 | 2469.62 | 2469.89 | [M-H2O-H]− |
| [ECA]4 | 2446.90 | 2445.61 | 2445.89 | [M-H]− |
| Fraction 28 (negative/positive modes) | ||||
| [ECA]4-ManNAcA-OAc2 | 2747.98 | 2746.72 | 2746.97 | [M-H]− |
| [ECA]4-GlcNAc-OAc2 | 2734.00 | 2732.75 | 2732.99 | [M-H]− |
| [ECA]4-ManNAcA-OAc | 2705.97 | 2704.82 | 2704.96 | [M-H]− |
| [ECA]4-GlcNAc-OAc | 2691.99 | 2690.78 | 2690.98 | [M-H]− |
| [ECA]4-ManNAcA | 2663.96 | 2662.77 | 2662.95 | [M-H]− |
| [ECA]4-GlcNAc | 2649.98 | 2648.82 | 2648.97 | [M-H]− |
| [ECA]4-OAc3 | 2572.93 | 2571.74 | 2571.92 | [M-H]− |
| [ECA]4-OAc3 | 2554.92 | 2553.78 | 2553.91 | [M-H2O-H]− |
| [ECA]4-OAc2 | 2530.92 | 2529.71 | 2529.91 | [M-H]− |
| [ECA]-Gal3-Glc2-Hep3-Kdo-P3-Etn | 2514.69 | 2513.73 | 2513.68 | [M-H]− |
| [ECA]4-OAc2 | 2512.91 | 2511.68 | 2511.90 | [M-H2O-H]− |
| [ECA]4-OAc | 2488.91 | 2487.67 | 2487.90 | [M-H]− |
| [ECA]4-OAc | 2470.90 | 2469.70 | 2469.89 | [M-H2O-H]− |
| [ECA]4 | 2446.90 | 2445.62 | 2445.89 | [M-H]− |
| [ECA]4-OAc | 2488.91 | 830.67 | 830.64 | [M+3H]3+ |
| [ECA]4-OAc2 | 2530.92 | 838.64 | 838.65 | [M-H2O+3H]3+ |
| [ECA]4-OAc2 | 2530.92 | 844.67 | 844.65 | [M+3H]3+ |
| [ECA]-Gal3-Glc2-Hep3-Kdo-P3-Etn | 2514.69 | 839.34 | 839.24 | [M+3H]3+ |
| [ECA]4-OAc3 | 2572.93 | 858.67 | 858.65 | [M+3H]3+ |
| [ECA]4-OAc3 | 2572.93 | 852.68 | 852.65 | [M-H2O+3H]3+ |
| [ECA]-Gal3-Glc2-Hep3-Kdo-P4-Etn | 2594.65 | 865.98 | 865.89 | [M+3H]3+ |
| [ECA]4-GlcNAc-OAc | 2691.99 | 892.40 | 892.34 | [M-H2O+3H]3+ |
| [ECA]4-ManNAcA-OAc3 | 2771.98 | 925.06 | 925.00 | [M-H2O+3H]3+ |
| Oligosaccharide Structure | Calculated Monoisotopic Mass (Da) | Observed Ion (m/z) | Calculated Ion (m/z) | Interpretation of the Ion |
|---|---|---|---|---|
| [ECA]4-FucNAc-OAc3 | 2760.02 | 2759.94 | 2759.01 | [M-H]− |
| [ECA]4-FucNAc-OAc3 | 2760.02 | 2741.93 | 2740.99 | [M-H2O-H]− |
| [ECA]4-FucNAc-OAc2 | 2718.01 | 2717.90 | 2717.00 | [M-H]− |
| Glc3-Gal-GlcNAc-Hep3-Kdo-P3 | 1905.45 | 1904.49 | 1904.44 | [M-H]− |
| Glc3-Gal-GlcNAc-Hep3-Kdo-P3-Etn | 1948.49 | 1947.51 | 1947.48 | [M-H]− |
| Glc3-Gal-GlcNAc-Hep3-Kdo-P3-Etn | 1948.49 | 1929.53 | 1929.47 | [M-H2O-H]− |
| Glc3-Gal-GlcNAc2-Hep3-Kdo-P1 | 1948.60 | 1947.63 | 1947.59 | [M-H]− |
| Glc3-Gal-GlcNAc2-Hep3-Kdo-P1 | 1948.60 | 1929.60 | 1929.57 | [M-H2O-H]− |
| Glc3-Gal-GlcNAc2-Hep3-Kdo-P2 | 2028.66 | 2027.59 | 2027.55 | [M-H]− |
| Glc3-Gal-GlcNAc2-Hep3-Kdo-P2 | 2028.66 | 2009.58 | 2009.54 | [M-H2O-H]− |
| Glc3-Gal-GlcNAc2-Hep3-Kdo-P2-Etn | 2071.60 | 2070.68 | 2070.60 | [M-H]− |
| Glc3-Gal-GlcNAc2-Hep3-Kdo-P2-Etn | 207.160 | 2052.62 | 2052.58 | [M-H2O-H]− |
| Glc3-Gal-GlcNAc2-Hep3-Kdo-P3-Etn | 2151.57 | 2150.59 | 2150.56 | [M-H]− |
| Glc3-Gal-GlcNAc2-Hep3-Kdo-P3-Etn | 2151.57 | 2132.57 | 2132.55 | [M-H2O-H]− |
| Glc3-Gal-GlcNAc-Hep3-GlcN-Kdo-P1 | 1906.58 | 1887.63 | 1887.56 | [M-H2O-H]− |
| Glc3-Gal-GlcNAc-Hep3-GlcN-Kdo-P1 | 1906.58 | 1905.61 | 1905.58 | [M-H]− |
| Glc3-Gal-GlcNAc-Hep3-GlcN-Kdo-P2 | 1986.55 | 1967.57 | 1967.53 | [M-H2O-H]− |
| Glc3-Gal-GlcNAc-Hep3-GlcN-Kdo-P2 | 1986.55 | 1985.56 | 1985.54 | [M-H]− |
| Glc3-Gal-GlcNAc-Hep3-GlcN-Kdo-P2-Etn | 2029.59 | 2010.64 | 2010.57 | [M-H2O-H]− |
| Glc3-Gal-GlcNAc-Hep3-GlcN-Kdo-P2-Etn | 2029.59 | 2028.63 | 2028.59 | [M-H]− |
| Glc3-Gal-GlcNAc-Hep3-GlcN-Kdo-P3-Etn | 2109.56 | 2108.59 | 2108.55 | [M-H]− |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Maciejewska, A.; Kaszowska, M.; Jachymek, W.; Lugowski, C.; Lukasiewicz, J. Lipopolysaccharide-Linked Enterobacterial Common Antigen (ECALPS) Occurs in Rough Strains of Escherichia coli R1, R2, and R4. Int. J. Mol. Sci. 2020, 21, 6038. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21176038
Maciejewska A, Kaszowska M, Jachymek W, Lugowski C, Lukasiewicz J. Lipopolysaccharide-Linked Enterobacterial Common Antigen (ECALPS) Occurs in Rough Strains of Escherichia coli R1, R2, and R4. International Journal of Molecular Sciences. 2020; 21(17):6038. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21176038
Chicago/Turabian StyleMaciejewska, Anna, Marta Kaszowska, Wojciech Jachymek, Czeslaw Lugowski, and Jolanta Lukasiewicz. 2020. "Lipopolysaccharide-Linked Enterobacterial Common Antigen (ECALPS) Occurs in Rough Strains of Escherichia coli R1, R2, and R4" International Journal of Molecular Sciences 21, no. 17: 6038. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21176038