Novel Insights into the Regulatory Role of Nuclear Factor (Erythroid-Derived 2)-Like 2 in Oxidative Stress and Inflammation of Human Fetal Membranes



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
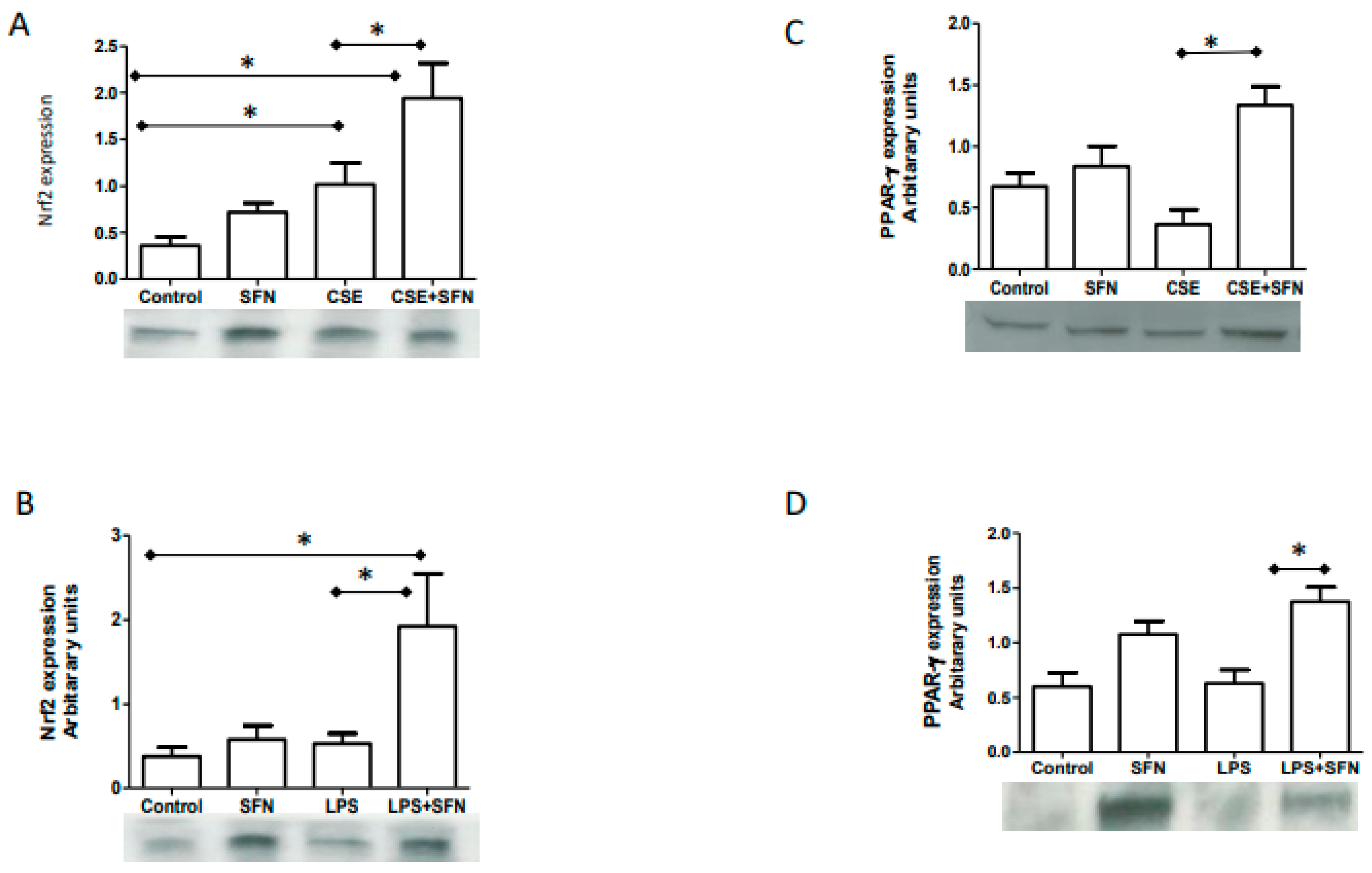
2.1. CSE and LPS Increase Nrf2 Expressions in Human Fetal Membranes
2.2. SFN Co-Treatment Augments CSE and LPS Response and Increase Nrf2
2.3. SFN Co-Treatment with CSE or LPS Increases PPARγ Expression
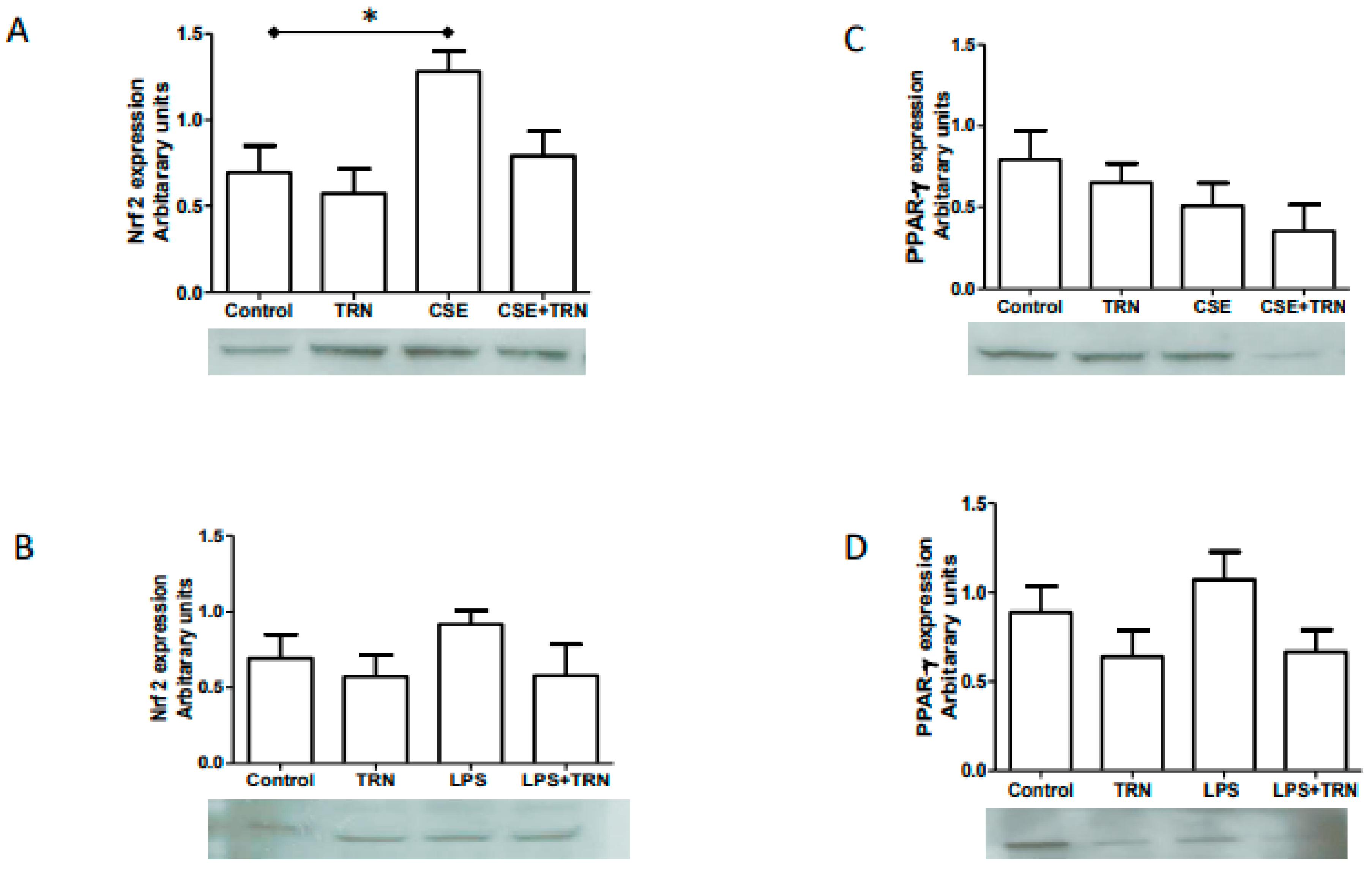
2.4. TRN Co-Treatment with CSE and LPS Did Not Change Nrf2 and PPARγ Expressions
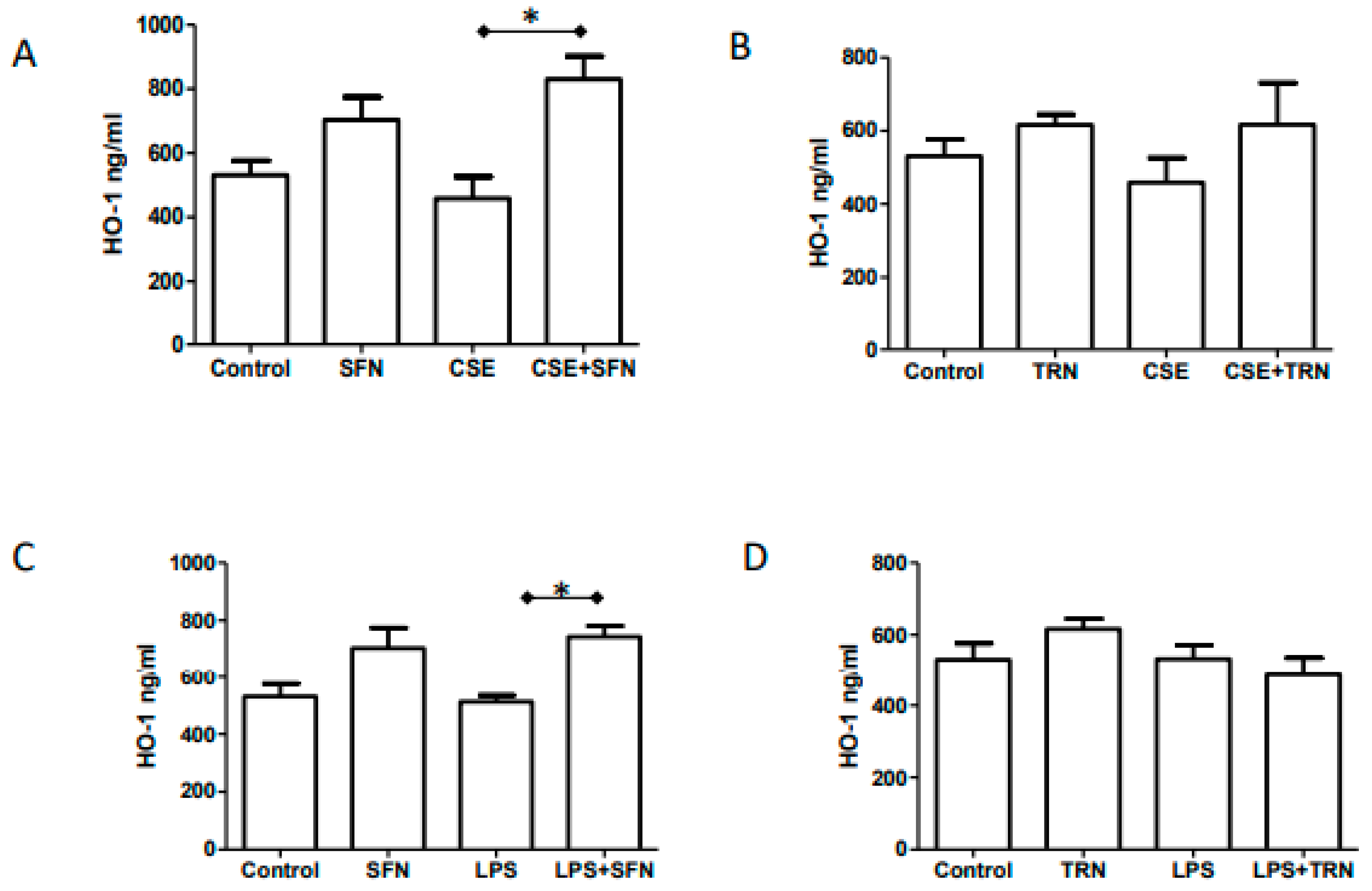
2.5. Endogenous HO-1 Levels are Increased by SFN Co-Treatment of CSE and LPS
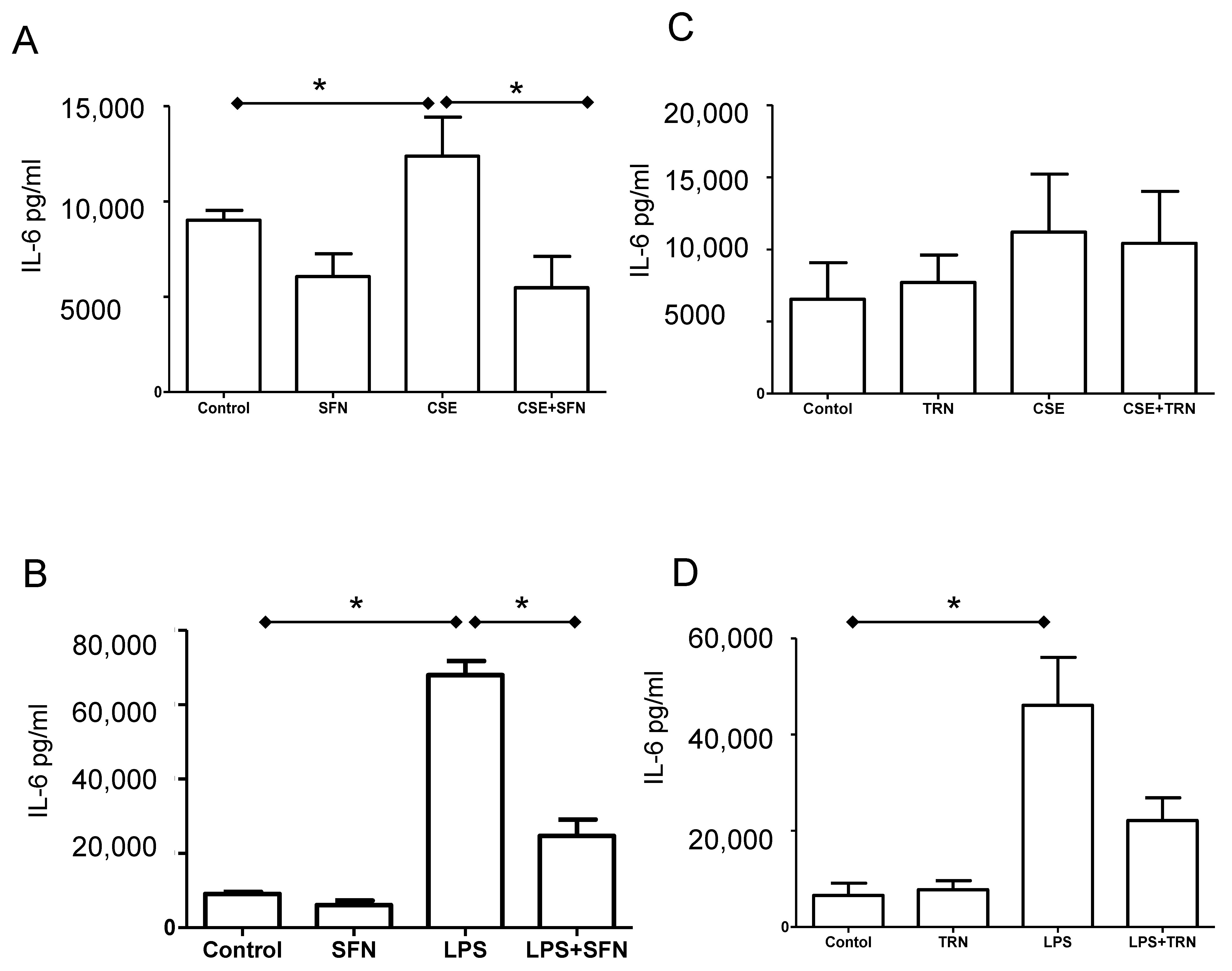
2.6. SFN Reduces IL-6 Release Induced by Both CSE and LPS
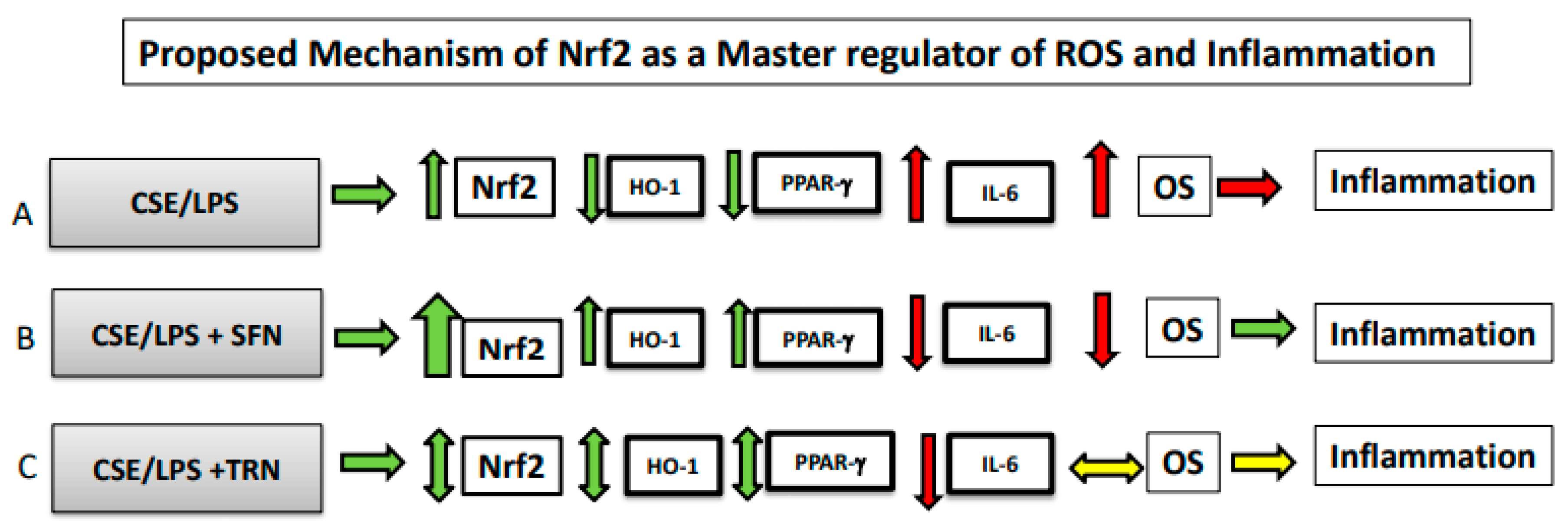
3. Discussion
4. Materials and Methods
4.1. IRB Ethics Committee Approval Statement
4.2. Tissue-Culture of Normal Term Fetal Membranes
4.3. Water-Soluble Cigarette Smoke Extract (CSE) Preparation and Stimulation of the Fetal Membranes with CSE and LPS
4.4. Treatment with SFN and TRN
4.5. Western Blot Analysis for Nrf2 and PPARγ
4.6. HO-1 ELISA
4.7. Luminex Assay to Determine Cytokine Concentration in Culture Supernatants
4.8. Statistical Analysis
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Beck, S.; Wojdyla, D.; Say, L.; Betran, A.P.; Merialdi, M.; Requejo, J.H.; Rubens, C.; Menon, R.; Van Look, P.F. The worldwide incidence of preterm birth: A systematic review of maternal mortality and morbidity. Bull. World Health Organ. 2009, 88, 31–38. [Google Scholar] [CrossRef]
- McCabe, E.R.; Carrino, G.E.; Russell, R.B.; Howse, J.L. Fighting for the next generation: US Prematurity in 2030. Pediatrics 2014, 134, 1193–1199. [Google Scholar] [CrossRef] [Green Version]
- Ananth, C.V.; Vintzileos, A.M. Epidemiology of preterm birth and its clinical subtypes. J. Matern. Neonatal Med. 2006, 19, 773–782. [Google Scholar] [CrossRef]
- Goldenberg, R.L.; Culhane, J.F.; Iams, J.D.; Romero, R. Epidemiology and causes of preterm birth. Lancet 2008, 371, 75–84. [Google Scholar] [CrossRef]
- Menon, R.; Fortunato, S.J. Infection and the role of inflammation in preterm premature rupture of the membranes. Best Pr. Res. Clin. Obstet. Gynaecol. 2007, 21, 467–478. [Google Scholar] [CrossRef]
- Menon, R.; Taylor, B.D. Exploring Inflammatory Mediators in Fetal and Maternal Compartments During Human Parturition. Obstet. Gynecol. 2019, 134, 765–773. [Google Scholar] [CrossRef]
- Park, J.S.; Park, C.W.; Lockwood, C.J.; Norwitz, E.R. Role of cytokines in preterm labor and birth. Minerva Ginecol. 2005, 57, 349–366. [Google Scholar]
- Vadillo-Ortega, F.; Estrada-Gutierrez, G. Role of matrix metalloproteinases in preterm labour. BJOG 2005, 112 (Suppl. S1), 19–22. [Google Scholar] [CrossRef]
- Jacobsson, B.; Mattsby-Baltzer, I.; Andersch, B.; Bokstrom, H.; Holst, R.M.; Nikolaitchouk, N.; Wennerholm, U.B.; Hagberg, H. Microbial invasion and cytokine response in amniotic fluid in a Swedish population of women with preterm prelabor rupture of membranes. Acta Obstet. Gynecol. Scand. 2003, 82, 423–431. [Google Scholar] [CrossRef]
- Cobo, T.; Kacerovsky, M.; Holst, R.M.; Hougaard, D.M.; Skogstrand, K.; Wennerholm, U.B.; Hagberg, H.; Jacobsson, B. Intra-amniotic inflammation predicts microbial invasion of the amniotic cavity but not spontaneous preterm delivery in preterm prelabor membrane rupture. Acta Obstet. Gynecol. Scand. 2012, 91, 930–935. [Google Scholar] [CrossRef]
- Kacerovsky, M.; Celec, P.; Vlkova, B.; Skogstrand, K.; Hougaard, D.M.; Cobo, T.; Jacobsson, B. Amniotic fluid protein profiles of intraamniotic inflammatory response to Ureaplasma spp. and other bacteria. PLoS ONE 2013, 8, e60399. [Google Scholar] [CrossRef] [Green Version]
- Bhat, G.; Peltier, M.R.; Syed, T.A.; Drobek, C.O.; Saade, G.; Menon, R. Fetal membrane biomarker network diversity and disease functions induced by intra-amniotic pathogens. Am. J. Reprod. Immunol. 2013, 69, 124–133. [Google Scholar] [CrossRef] [Green Version]
- Abrahams, V.M.; Potter, J.A.; Bhat, G.; Peltier, M.R.; Saade, G.; Menon, R. Bacterial modulation of human fetal membrane Toll-like receptor expression. Am. J. Reprod. Immunol. 2013, 69, 33–40. [Google Scholar] [CrossRef] [Green Version]
- Menon, R.; Peltier, M.R.; Eckardt, J.; Fortunato, S.J. Diversity in cytokine response to bacteria associated with preterm birth by fetal membranes. Am. J. Obstet. Gynecol. 2009, 201, 306.e1–306.e6. [Google Scholar] [CrossRef]
- Burton, G.J. Oxygen, the Janus gas; its effects on human placental development and function. J. Anat. 2009, 215, 27–35. [Google Scholar] [CrossRef] [Green Version]
- Agarwal, A.; Aponte-Mellado, A.; Premkumar, B.J.; Shaman, A.; Gupta, S. The effects of oxidative stress on female reproduction: A review. Reprod. Biol. Endocrinol. 2012, 10, 1–31. [Google Scholar] [CrossRef] [Green Version]
- Agarwal, A.; Gupta, S.; Sekhon, L.; Shah, R. Redox considerations in female reproductive function and assisted reproduction: From molecular mechanisms to health implications. Antioxid. Redox Signal. 2008, 10, 1375–1404. [Google Scholar] [CrossRef]
- Dennery, P.A. Effects of oxidative stress on embryonic development. Birth Defects Res. Part C Embryo Today Rev. 2007, 81, 155–162. [Google Scholar] [CrossRef]
- Burton, G.J.; Jauniaux, E. Oxidative stress. Best. Pract. Res. Clin. Obstet. Gynaecol. 2011, 25, 287–299. [Google Scholar] [CrossRef] [Green Version]
- Herway, C.; Kanninen, T.; Witkin, S.S.; Saade, G.; Fortunato, S.J.; Menon, R. Ethnic disparity in amniotic fluid levels of hyaluronan, histone H2B and superoxide dismutase in spontaneous preterm birth. J. Périnat. Med. 2013, 41, 277–282. [Google Scholar] [CrossRef]
- Woods, J.R., Jr. Reactive oxygen species and preterm premature rupture of membranes-a review. Placenta 2001, 22 (Suppl. A), S38–S44. [Google Scholar] [CrossRef]
- Rumbold, A.; Duley, L.; Crowther, C.A.; Haslam, R.R. Antioxidants for preventing pre-eclampsia. Cochrane Database Syst. Rev 2008, CD004227. [Google Scholar] [CrossRef]
- Sultana, Z.; Maiti, K.; Aitken, J.; Morris, J.; Dedman, L.; Smith, R. Oxidative stress, placental ageing-related pathologies and adverse pregnancy outcomes. Am. J. Reprod. Immunol. 2017, 77, e12653. [Google Scholar] [CrossRef] [Green Version]
- Menon, R. Oxidative stress damage as a detrimental factor in preterm birth pathology 14. Front. Immunol. 2014, 5, 567. [Google Scholar] [CrossRef]
- Menon, R. Initiation of human parturition: Signaling from senescent fetal tissues via extracellular vesicle mediated paracrine mechanism. Obstet. Gynecol. Sci. 2019, 62, 199–211. [Google Scholar] [CrossRef]
- Rossi, A.C.; Mullin, P.M. Prevention of pre-eclampsia with low-dose aspirin or vitamins C and E in women at high or low risk: A systematic review with meta-analysis. Eur. J. Obstet. Gynecol. Reprod. Biol. 2011, 158, 9–16. [Google Scholar] [CrossRef]
- Hauth, J.C.; Clifton, R.G.; Roberts, J.M.; Spong, C.Y.; Myatt, L.; Leveno, K.J.; Pearson, G.D.; Varner, M.W.; Thorp, J.M., Jr.; Mercer, B.M.; et al. Vitamin C and E supplementation to prevent spontaneous preterm birth: A randomized controlled trial. Obstet. Gynecol. 2010, 116, 653–658. [Google Scholar] [CrossRef] [Green Version]
- Chappell, L.C.; Seed, P.T.; Kelly, F.J.; Briley, A.; Hunt, B.J.; Charnock-Jones, D.S.; Mallet, A.; Poston, L. Vitamin C and E supplementation in women at risk of preeclampsia is associated with changes in indices of oxidative stress and placental function. Am. J. Obstet. Gynecol. 2002, 187, 777–784. [Google Scholar] [CrossRef] [Green Version]
- Roberts, J.M.; Myatt, L.; Spong, C.Y.; Thom, E.A.; Hauth, J.C.; Leveno, K.J.; Pearson, G.D.; Wapner, R.J.; Varner, M.W.; Thorp, J.M., Jr.; et al. Vitamins C and E to prevent complications of pregnancy-associated hypertension. N. Engl. J. Med. 2010, 362, 1282–1291. [Google Scholar] [CrossRef] [Green Version]
- Rumbold, A.; Ota, E.; Nagata, C.; Shahrook, S.; Crowther, C.A. Vitamin C supplementation in pregnancy. Cochrane Database Syst. Rev. 2015, CD004072. [Google Scholar] [CrossRef]
- Rumbold, A.; Ota, E.; Hori, H.; Miyazaki, C.; Crowther, C.A. Vitamin E supplementation in pregnancy. Cochrane Database Syst. Rev. 2015, CD004069. [Google Scholar] [CrossRef]
- Lavu, N.; Richardson, L.; Radnaa, E.; Kechichian, T.; Urrabaz-Garza, R.; Sheller-Miller, S.; Bonney, E.; Menon, R. Oxidative stress-induced downregulation of glycogen synthase kinase 3 beta in fetal membranes promotes cellular senescencedagger. Biol. Reprod. 2019, 101, 1018–1030. [Google Scholar] [CrossRef] [PubMed]
- Menon, R.; Behnia, F.; Polettini, J.; Saade, G.R.; Campisi, J.; Velarde, M. Placental membrane aging and HMGB1 signaling associated with human parturition. Aging 2016, 8, 216–230. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Menon, R.; Boldogh, I.; Hawkins, H.K.; Woodson, M.; Polettini, J.; Syed, T.A.; Fortunato, S.J.; Saade, G.R.; Papaconstantinou, J.; Taylor, R.N. Histological evidence of oxidative stress and premature senescence in preterm premature rupture of the human fetal membranes recapitulated in vitro. Am. J. Pathol. 2014, 184, 1740–1751. [Google Scholar] [CrossRef]
- Menon, R.; Boldogh, I.; Urrabaz-Garza, R.; Polettini, J.; Syed, T.A.; Saade, G.R.; Papaconstantinou, J.; Taylor, R.N. Senescence of primary amniotic cells via oxidative DNA damage. PLoS ONE 2013, 8, e83416. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Menon, R.; Bonney, E.A.; Condon, J.; Mesiano, S.; Taylor, R.N. Novel concepts on pregnancy clocks and alarms: Redundancy and synergy in human parturition. Hum. Reprod. Update 2016, 22, 535–560. [Google Scholar] [CrossRef] [PubMed]
- Jin, J.; Richardson, L.; Sheller-Miller, S.; Zhong, N.; Menon, R. Oxidative stress induces p38MAPK-dependent senescence in the feto-maternal interface cells. Placenta 2018, 67, 15–23. [Google Scholar] [CrossRef] [PubMed]
- Rangasamy, T.; Cho, C.Y.; Thimmulappa, R.K.; Zhen, L.; Srisuma, S.S.; Kensler, T.W.; Yamamoto, M.; Petrache, I.; Tuder, R.M.; Biswal, S. Genetic ablation of Nrf2 enhances susceptibility to cigarette smoke-induced emphysema in mice. J. Clin. Investig. 2004, 114, 1248–1259. [Google Scholar] [CrossRef]
- Olgun, N.S.; Arita, Y.; Hanna, M.; Murthy, A.; Tristan, S.; Peltier, M.R.; Hanna, N. Carbon monoxide attenuates bacteria-induced Endothelin-1 expression in second trimester placental explants. Placenta 2014, 35, 351–358. [Google Scholar] [CrossRef]
- Peltier, M.R.; Koo, H.C.; Gurzenda, E.M.; Arita, Y.; Klimova, N.G.; Olgun, N.; Hanna, N. Can Carbon Monoxide Prevent Infection-Mediated Preterm Birth in a Mouse Model? Am. J. Reprod. Immunol. 2013, 70, 31–37. [Google Scholar] [CrossRef]
- Lin, W.; Wu, R.T.; Wu, T.; Khor, T.O.; Wang, H.; Kong, A.N. Sulforaphane suppressed LPS-induced inflammation in mouse peritoneal macrophages through Nrf2 dependent pathway. Biochem. Pharmacol. 2008, 76, 967–973. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harvey, C.J.; Thimmulappa, R.K.; Sethi, S.; Kong, X.; Yarmus, L.; Brown, R.H.; Feller-Kopman, D.; Wise, R.; Biswal, S. Targeting Nrf2 signaling improves bacterial clearance by alveolar macrophages in patients with COPD and in a mouse model. Sci. Transl. Med. 2011, 3, 78ra32. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mukherjee, S.; Gangopadhyay, H.; Das, D.K. Broccoli: A unique vegetable that protects mammalian hearts through the redox cycling of the thioredoxin superfamily. J. Agric. Food Chem. 2008, 56, 609–617. [Google Scholar] [CrossRef] [PubMed]
- Ludwig, I.A.; Clifford, M.N.; Lean, M.E.; Ashihara, H.; Crozier, A. Coffee: Biochemistry and potential impact on health. Food Funct. 2014, 5, 1695–1717. [Google Scholar] [CrossRef]
- Arlt, A.; Sebens, S.; Krebs, S.; Geismann, C.; Grossmann, M.; Kruse, M.L.; Schreiber, S.; Schafer, H. Inhibition of the Nrf2 transcription factor by the alkaloid trigonelline renders pancreatic cancer cells more susceptible to apoptosis through decreased proteasomal gene expression and proteasome activity. Oncogene 2012, 32, 4825–4835. [Google Scholar] [CrossRef] [PubMed]
- Boettler, U.; Sommerfeld, K.; Volz, N.; Pahlke, G.; Teller, N.; Somoza, V.; Lang, R.; Hofmann, T.; Marko, D. Coffee constituents as modulators of Nrf2 nuclear translocation and ARE (EpRE)-dependent gene expression. J. Nutr. Biochem. 2011, 22, 426–440. [Google Scholar] [CrossRef] [PubMed]
- Eskenazi, B.; Stapleton, A.L.; Kharrazi, M.; Chee, W.Y. Associations between maternal decaffeinated and caffeinated coffee consumption and fetal growth and gestational duration. Epidemiology 1999, 10, 242–249. [Google Scholar] [CrossRef]
- McDonald, A.D.; Armstrong, B.G.; Sloan, M. Cigarette, alcohol, and coffee consumption and congenital defects. Am. J. Public Health 1992, 82, 91–93. [Google Scholar] [CrossRef] [Green Version]
- McDonald, A.D.; Armstrong, B.G.; Sloan, M. Cigarette, alcohol, and coffee consumption and prematurity. Am. J. Public Health 1992, 82, 87–90. [Google Scholar] [CrossRef]
- Santos, I.S.; Victora, C.G.; Huttly, S.; Morris, S. Caffeine intake and pregnancy outcomes: A meta-analytic review. Cad. Saude Publica 1998, 14, 523–530. [Google Scholar] [CrossRef]
- Silveira, M.F.; Santos, I.S.; Barros, A.J.; Matijasevich, A.; Barros, F.C.; Victora, C.G. Increase in preterm births in Brazil: Review of population-based studies. Rev. Saude Publica 2008, 42, 957–964. [Google Scholar] [CrossRef]
- Li, W.; Kong, A.N. Molecular mechanisms of Nrf2-mediated antioxidant response. Mol. Carcinog. 2009, 48, 91–104. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sinha, D.; Biswas, J.; Bishayee, A. Nrf2-mediated redox signaling in arsenic carcinogenesis: A review. Arch. Toxicol. 2013, 87, 383–396. [Google Scholar] [CrossRef] [PubMed]
- Srisook, K.; Kim, C.; Cha, Y.N. Molecular mechanisms involved in enhancing HO-1 expression: De-repression by heme and activation by Nrf2, the “one-two” punch. Antioxid. Redox Signal. 2005, 7, 1674–1687. [Google Scholar] [CrossRef] [PubMed]
- Jazwa, A.; Cuadrado, A. Targeting heme oxygenase-1 for neuroprotection and neuroinflammation in neurodegenerative diseases. Curr. Drug Targets 2010, 11, 1517–1531. [Google Scholar] [CrossRef]
- Feng, H.; Wang, L.; Zhang, G.; Zhang, Z.; Guo, W. Oxidative stress activated by Keap-1/Nrf2 signaling pathway in pathogenesis of preeclampsia. Int. J. Clin. Exp. Pathol. 2020, 13, 382–392. [Google Scholar]
- Zenclussen, M.L.; Casalis, P.A.; El-Mousleh, T.; Rebelo, S.; Langwisch, S.; Linzke, N.; Volk, H.D.; Fest, S.; Soares, M.P.; Zenclussen, A.C. Haem oxygenase-1 dictates intrauterine fetal survival in mice via carbon monoxide. J. Pathol. 2011, 225, 293–304. [Google Scholar] [CrossRef]
- Zoete, V.; Grosdidier, A.; Michielin, O. Peroxisome proliferator-activated receptor structures: Ligand specificity, molecular switch and interactions with regulators. Biochim. Biophys. Acta 2007, 1771, 915–925. [Google Scholar] [CrossRef]
- Toth, B.; Hornung, D.; Scholz, C.; Djalali, S.; Friese, K.; Jeschke, U. Peroxisome proliferator-activated receptors: New players in the field of reproduction. Am. J. Reprod. Immunol. 2007, 58, 289–310. [Google Scholar] [CrossRef]
- Lappas, M.; Rice, G.E. Phospholipase A2 isozymes in pregnancy and parturition. Prostaglandins Leukot. Essent. Fatty Acids 2004, 70, 87–100. [Google Scholar] [CrossRef]
- Capece, A.; Vasieva, O.; Meher, S.; Alfirevic, Z.; Alfirevic, A. Pathway analysis of genetic factors associated with spontaneous preterm birth and pre-labor preterm rupture of membranes. PLoS ONE 2014, 9, e108578. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lappas, M.; Rice, G.E. Transcriptional regulation of the processes of human labour and delivery. Placenta 2009, 30 (Suppl. A), S90–S95. [Google Scholar] [CrossRef]
- Sykes, L.; Thomson, K.R.; Boyce, E.J.; Lee, Y.S.; Rasheed, Z.B.; MacIntyre, D.A.; Teoh, T.G.; Bennett, P.R. Sulfasalazine augments a pro-inflammatory response in interleukin-1beta-stimulated amniocytes and myocytes. Immunology 2015, 146, 630–644. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lim, R.; Barker, G.; Lappas, M. The transcription factor Nrf2 is decreased after spontaneous term labour in human fetal membranes where it exerts anti-inflammatory properties. Placenta 2015, 36, 7–17. [Google Scholar] [CrossRef]
- Chigusa, Y.; Kishore, A.H.; Mogami, H.; Word, R.A. Nrf2 Activation Inhibits Effects of Thrombin in Human Amnion Cells and Thrombin-Induced Preterm Birth in Mice. J. Clin. Endocrinol. Metab. 2016, 101, 2612–2621. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Li, M.; Li, N.; Liu, Z. Regulation of Keap-1/Nrf2 Signaling Pathway Is Activated by Oxidative Stress in Patients with Premature Rupture of Membranes. Med. Sci. Monit. 2020, 26, e921757. [Google Scholar] [CrossRef] [PubMed]
- Hong, F.; Freeman, M.L.; Liebler, D.C. Identification of sensor cysteines in human Keap1 modified by the cancer chemopreventive agent sulforaphane. Chem. Res. Toxicol. 2005, 18, 1917–1926. [Google Scholar] [CrossRef]
- McMahon, M.; Itoh, K.; Yamamoto, M.; Hayes, J.D. Keap1-dependent proteasomal degradation of transcription factor Nrf2 contributes to the negative regulation of antioxidant response element-driven gene expression. J. Biol. Chem. 2003, 278, 21592–21600. [Google Scholar] [CrossRef] [Green Version]
- Dutta, E.H.; Behnia, F.; Boldogh, I.; Saade, G.R.; Taylor, B.D.; Kacerovsky, M.; Menon, R. Oxidative stress damage-associated molecular signaling pathways differentiate spontaneous preterm birth and preterm premature rupture of the membranes. Mol. Hum. Reprod. 2016, 22, 143–157. [Google Scholar] [CrossRef] [Green Version]
- Halliwell, B.; Whiteman, M. Measuring reactive species and oxidative damage in vivo and in cell culture: How should you do it and what do the results mean? Br. J. Pharmacol. 2004, 142, 231–255. [Google Scholar] [CrossRef] [Green Version]
- Scholl, T.O.; Stein, T.P. Oxidant damage to DNA and pregnancy outcome. J. Matern. Fetal Med. 2001, 10, 182–185. [Google Scholar] [CrossRef] [PubMed]
- Mitra, S.; Boldogh, I.; Izumi, T.; Hazra, T.K. Complexities of the DNA base excision repair pathway for repair of oxidative DNA damage. Environ. Mol. Mutagen. 2001, 38, 180–190. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fortunato, S.J.; Menon, R.; Swan, K.F.; Lyden, T.W. Organ culture of amniochorionic membrane in vitro. Am. J. Reprod. Immunol. 1994, 32, 184–187. [Google Scholar] [CrossRef] [PubMed]
- Fortunato, S.J.; Menon, R.; Swan, K.F. Amniochorion: A source of interleukin-8. Am. J. Reprod. Immunol. 1995, 34, 156–162. [Google Scholar] [CrossRef] [PubMed]





© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Menon, R.; Peltier, M.R. Novel Insights into the Regulatory Role of Nuclear Factor (Erythroid-Derived 2)-Like 2 in Oxidative Stress and Inflammation of Human Fetal Membranes. Int. J. Mol. Sci. 2020, 21, 6139. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21176139
Menon R, Peltier MR. Novel Insights into the Regulatory Role of Nuclear Factor (Erythroid-Derived 2)-Like 2 in Oxidative Stress and Inflammation of Human Fetal Membranes. International Journal of Molecular Sciences. 2020; 21(17):6139. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21176139
Chicago/Turabian StyleMenon, Ramkumar, and Morgan R Peltier. 2020. "Novel Insights into the Regulatory Role of Nuclear Factor (Erythroid-Derived 2)-Like 2 in Oxidative Stress and Inflammation of Human Fetal Membranes" International Journal of Molecular Sciences 21, no. 17: 6139. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21176139