Ameliorated Autoimmune Arthritis and Impaired B Cell Receptor-Mediated Ca2+ Influx in Nkx2-3 Knock-out Mice

 ,
,  ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
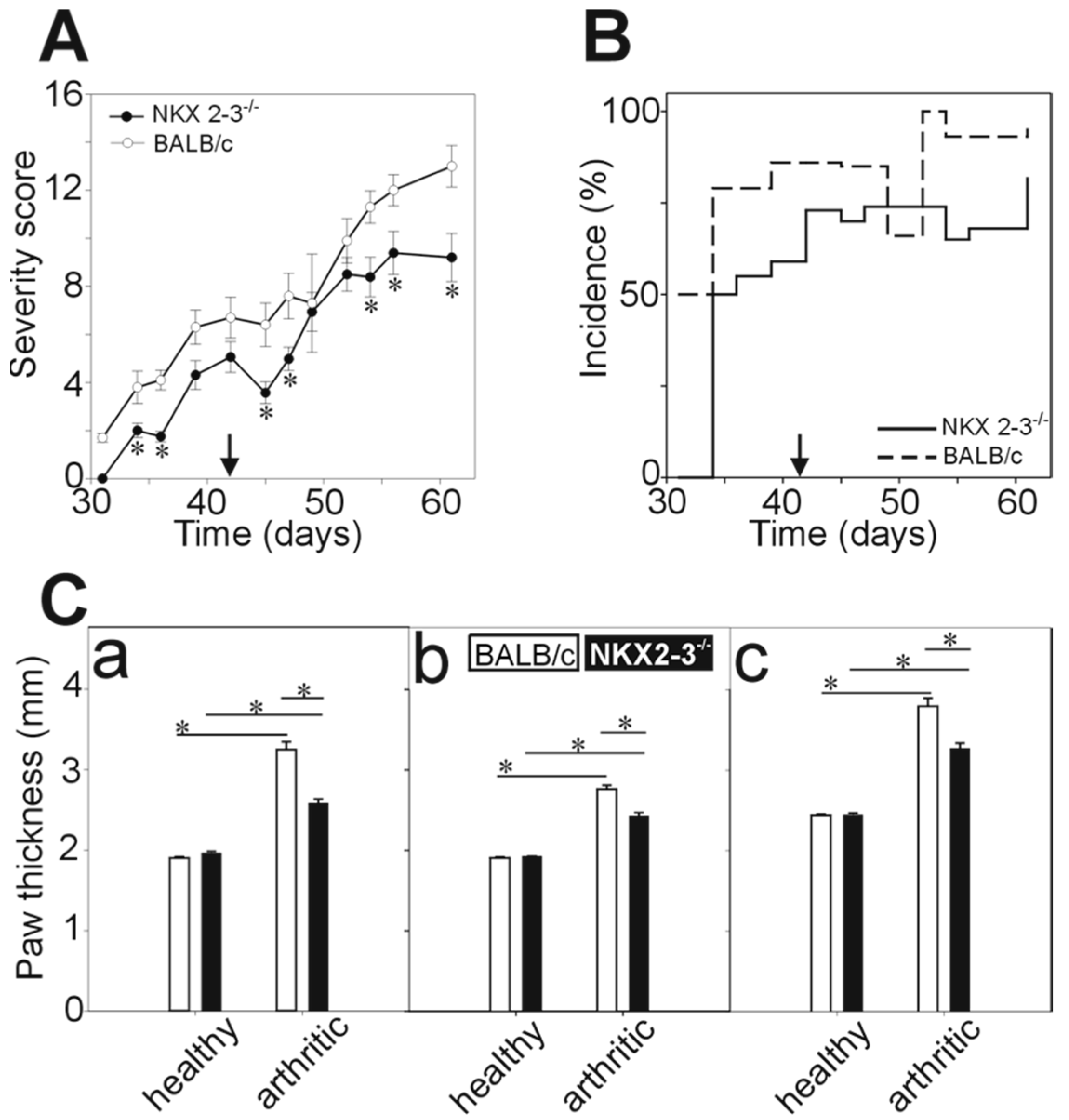
2.1. Decreased Severity and Incidence of rhG1-Induced Arthritis in Nkx2-3 Knock-Out Mice
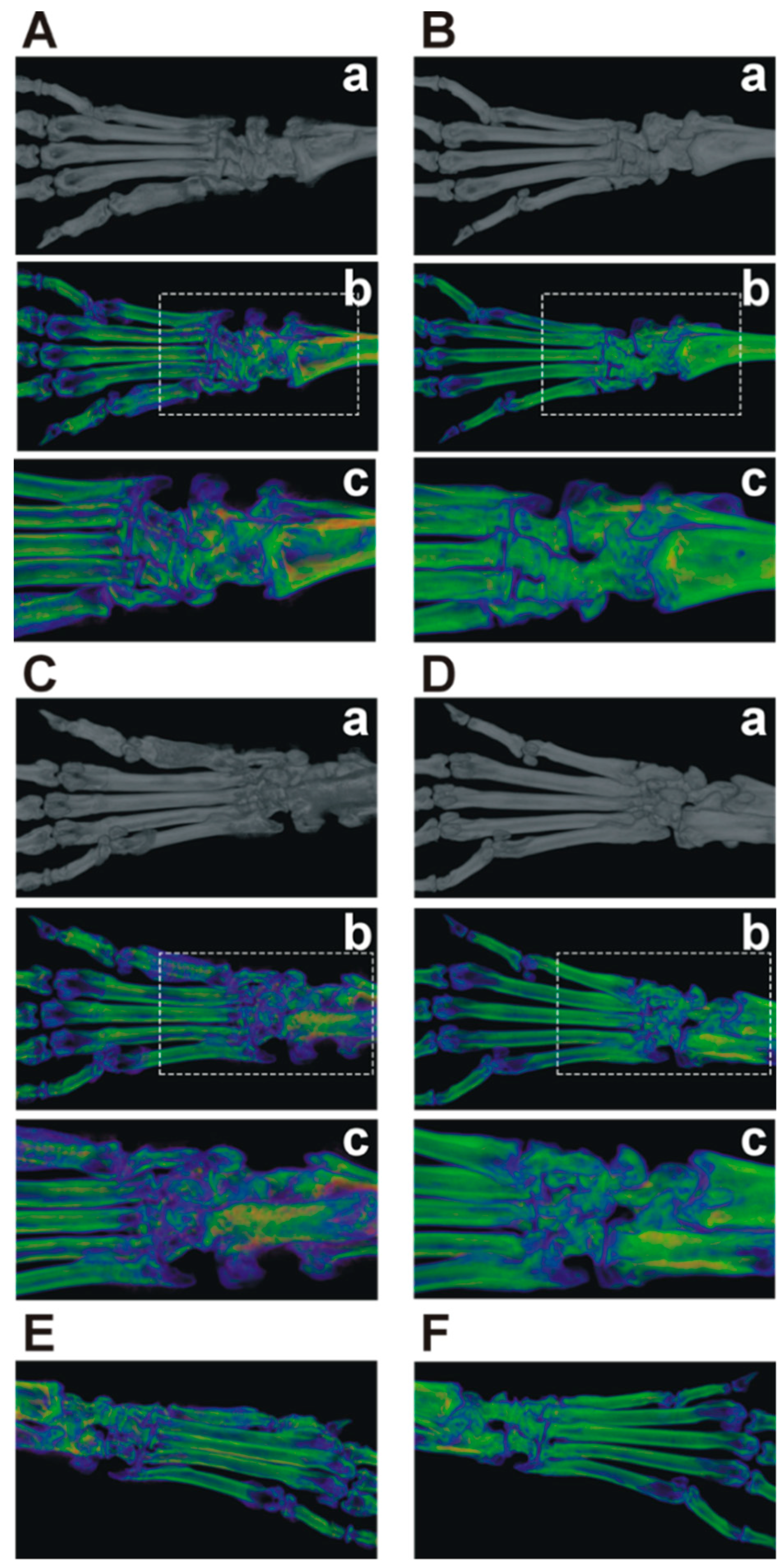
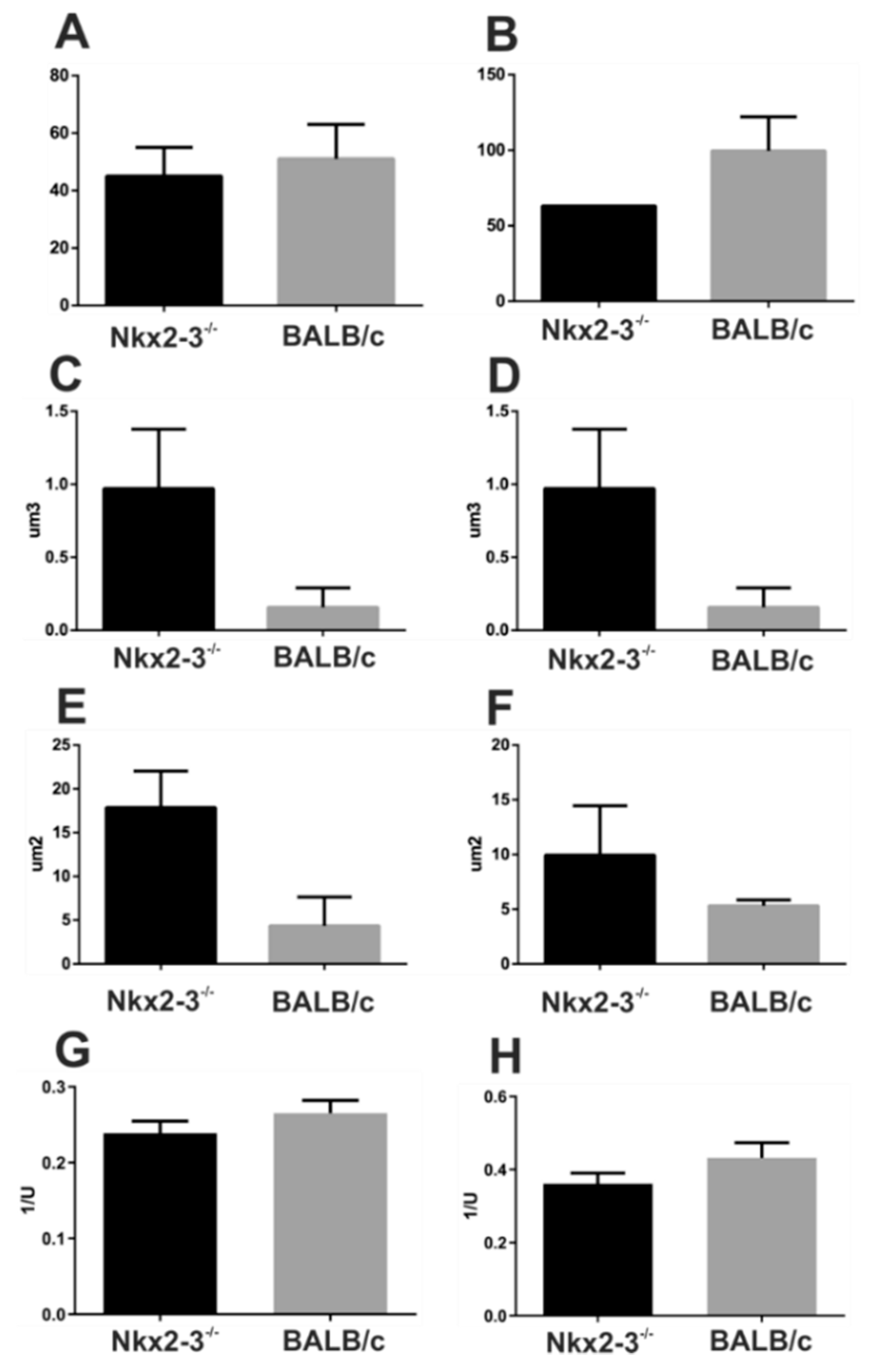
2.2. Micro-CT Imaging Confirmed Decreased Cartilage and Bone Destruction in Arthritic Nkx2-3 Knock-Out Mice
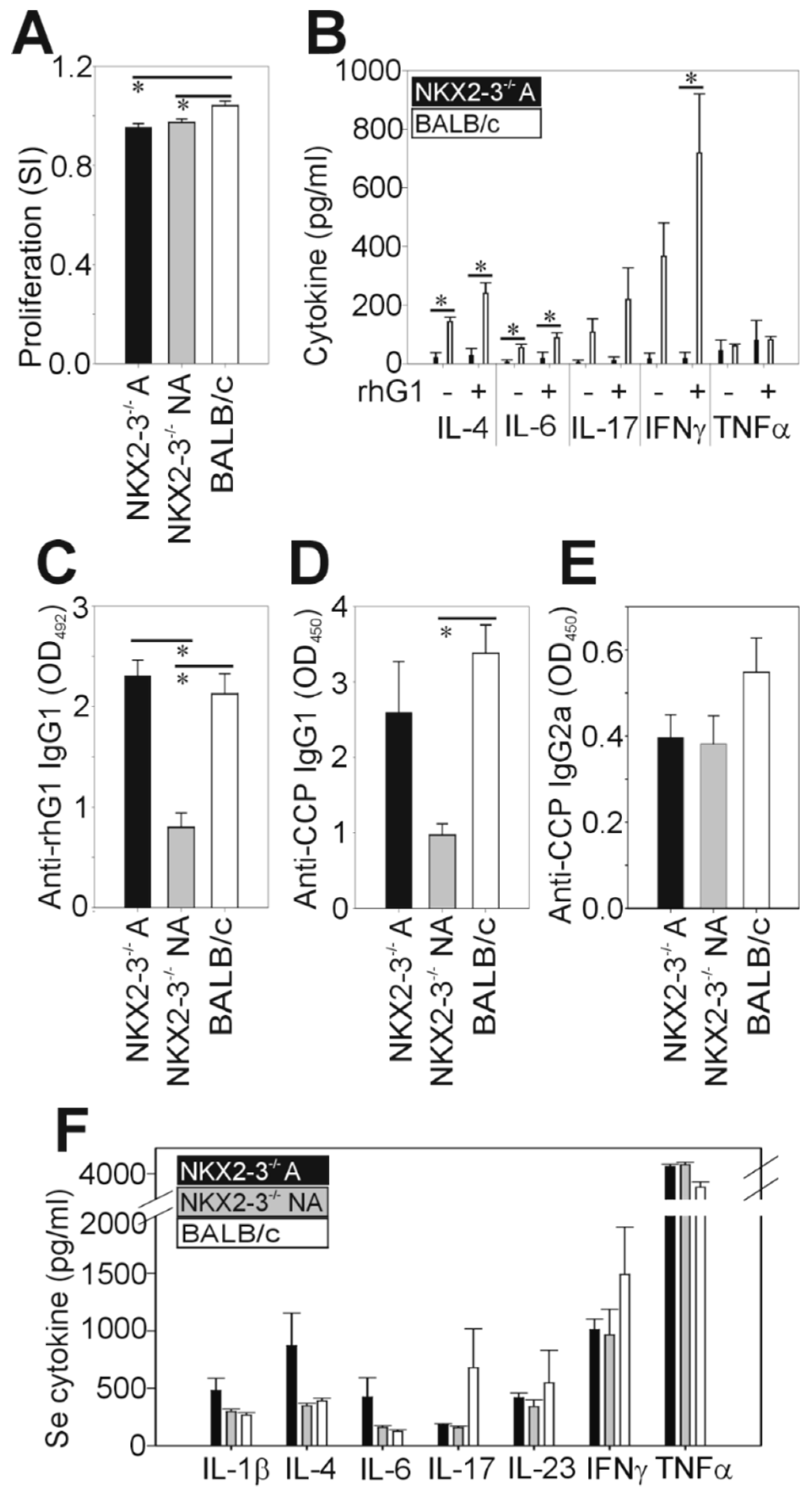
2.3. Comparison of the G1-Specific Immune Response between Nkx2-3−/− and BALB/c Mice
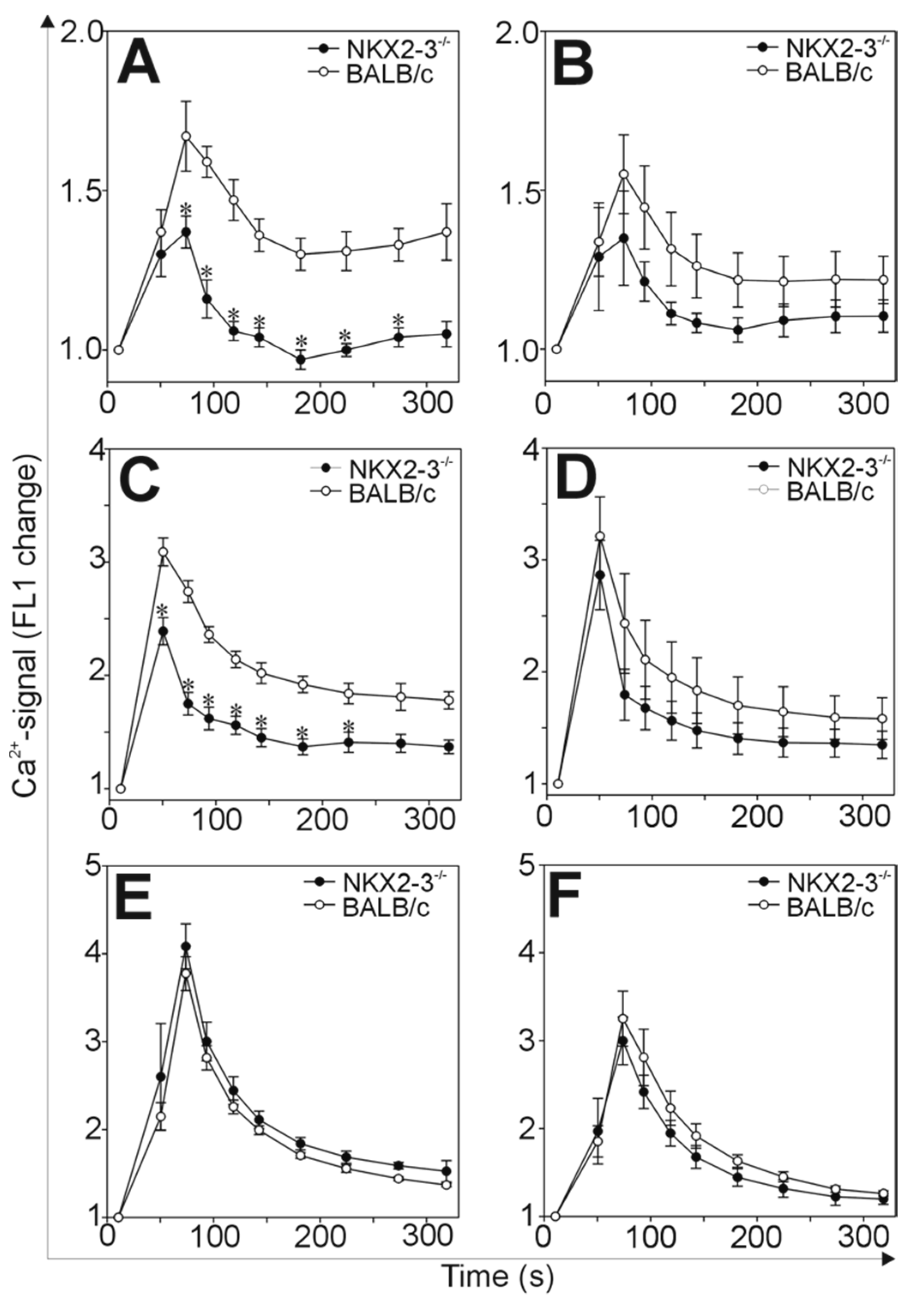
2.4. Comparison of the Ca2+-Signaling in B and T Cells of Nkx2-3−/− and BALB/c Mice
3. Discussion
4. Materials and Methods
4.1. Mice
4.2. Induction and Assessment of Recombinant Human G1-Induced Arthritis
4.3. Micro-CT
4.4. In Vitro Spleen Cell Culture
4.5. Antigen-Specific Proliferation Assay
4.6. ELISA Measurements
4.7. Ca2+ Signaling Measurements
4.8. Statistical Analysis
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| a-CCP | Anticyclic citrullinated peptide antibody |
| APCs | Antigen-presenting cells |
| Ca2+ | Calcium |
| CD | cluster of differentiation |
| DDA | Dimethyl-Dioctadecyl-ammonium adjuvant |
| DMEM | Dulbecco’s modified Eagle’s medium |
| ELISA | Enzyme-linked immunosorbent assay |
| FCS | Fetal calf serum |
| FSC | Forward scatter |
| GC | Germinal center |
| GIA | Recombinant human G1-induced arthritis |
| i.p | Intraperitoneal |
| IFN-γ | Interferon γ |
| IGH | Immunoglobulin heavy chain gene |
| IL | Interleukin |
| MADCAM-1 | Mucosal vascular addressin cell adhesion molecule 1 |
| MZ | Marginal zone |
| NFDM | Nonfat dry milk |
| NF-kB | Nuclear factor kappa-light-chain-enhancer of activated B cells |
| Nkx2-3 | Nirenberg-Kim (NK) 2 homeobox 3 |
| Nkx2-3−/− | Nkx2-3-deficient mouse (homozygous) |
| PBS | Phosphate buffer solution |
| PI3K | Phosphoinositide 3-kinases |
| PKB | Protein Kinase B |
| PTPN2 | Tyrosine-protein phosphatase nonreceptor type 2 |
| RA | Rheumatoid arthritis |
| rhG1 | Recombinant human G1 |
| RPMI | Roswell Park Memorial Institute Medium |
| SSC | Side scatter |
| TF | Transcription factor |
| Th | T helper cell |
| TNF-α | Tumor necrosis factor α |
| VEGF | Vascular endothelial growth factor |
Appendix A

References
- Scott, D.L.; Wolfe, F.; Huizinga, T.W.J. Rheumatoid arthritis. Lancet 2010, 376, 1094–1108. [Google Scholar] [CrossRef]
- Lin, Y.-J.; Anzaghe, M.; Schülke, S. Update on the Pathomechanism, Diagnosis, and Treatment Options for Rheumatoid Arthritis. Cells 2020, 9, 880. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kugyelka, R.; Kohl, Z.; Olasz, K.; Mikecz, K.; Rauch, T.A.; Glant, T.T.; Boldizsar, F. Enigma of IL-17 and Th17 cells in rheumatoid arthritis and in autoimmune animal models of arthritis. Mediat. Inflamm. 2016, 2016, 6145810. [Google Scholar] [CrossRef] [Green Version]
- Glant, T.T.; Mikecz, K.; Arzoumanian, A.; Poole, A.R. Proteoglycan-induced arthritis in BALB/c mice. Clinical features and histopathology. Arthritis Rheum. 1987, 30, 201–212. [Google Scholar] [CrossRef] [PubMed]
- Glant, T.T.; Radacs, M.; Nagyeri, G.; Olasz, K.; Laszlo, A.; Boldizsar, F.; Hegyi, A.; Finnegan, A.; Mikecz, K. Proteoglycan-induced arthritis and recombinant human proteoglycan aggrecan G1 domain-induced arthritis in BALB/c mice resembling two subtypes of rheumatoid arthritis. Arthritis Rheum. 2011, 63, 1312–1321. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Horváth, Á.; Borbély, É.; Bölcskei, K.; Szentes, N.; Kiss, T.; Belák, M.; Rauch, T.; Glant, T.; Zákány, R.; Juhász, T.; et al. Regulatory role of capsaicin-sensitive peptidergic sensory nerves in the proteoglycan-induced autoimmune arthritis model of the mouse. J. Neuroinflamm. 2018, 15, 335. [Google Scholar] [CrossRef] [PubMed]
- Hanyecz, A.; Olasz, K.; Tarjanyi, O.; Nemeth, P.; Mikecz, K.; Glant, T.T.; Boldizsar, F. Proteoglycan aggrecan conducting T cell activation and apoptosis in a murine model of rheumatoid arthritis. Biomed. Res. Int. 2014, 2014, 942148. [Google Scholar] [CrossRef] [Green Version]
- Boldizsar, F.; Tarjanyi, O.; Nemeth, P.; Mikecz, K.; Glant, T.T. Th1/Th17 polarization and acquisition of an arthritogenic phenotype in arthritis-susceptible BALB/c, but not in MHC-matched, arthritis-resistant DBA/2 mice. Int. Immunol. 2009, 21, 511–522. [Google Scholar] [CrossRef] [Green Version]
- McInnes, I.B.; Schett, G. Pathogenetic insights from the treatment of rheumatoid arthritis. Lancet 2017, 389, 2328–2337. [Google Scholar] [CrossRef] [Green Version]
- Smolen, J.S.; Aletaha, D.; Barton, A.; Burmester, G.R.; Emery, P.; Firestein, G.S.; Kavanaugh, A.; McInnes, I.B.; Solomon, D.H.; Strand, V.; et al. Rheumatoid arthritis. Nat. Rev. Dis. Prim. 2018, 4, 18001. [Google Scholar] [CrossRef]
- Abbas, A.; Lichtman, A.; Pillai, S. Cellular and Molecular Immunology; Elsevier: Amsterdam, The Netherlands, 2017; Available online: https://www.elsevier.com/books/cellular-and-molecular-immunology/abbas/978-0-323-47978-3 (accessed on 15 June 2020).
- Crane, G.M.; Liu, Y.-C.; Chadburn, A. Spleen: Development, anatomy and reactive lymphoid proliferations. Semin. Diagn. Pathol. 2020. [Google Scholar] [CrossRef] [PubMed]
- Tsuji, F.; Yoshimi, M.; Katsuta, O.; Takai, M.; Ishihara, K.; Aono, H. Point mutation of tyrosine 759 of the IL-6 family cytokine receptor, gp130, augments collagen-induced arthritis in DBA/1J mice. BMC Musculoskelet. Disord. 2009, 10, 23. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, W.; Hegarty, J.P.; Berg, A.; Chen, X.; West, G.; Kelly, A.A.; Wang, Y.; Poritz, L.S.; Koltun, W.A.; Lin, Z. NKX2-3 transcriptional regulation of endothelin-1 and VEGF signaling in human intestinal microvascular endothelial cells. PLoS ONE 2011, 6, e20454. [Google Scholar] [CrossRef] [PubMed]
- Nagel, S.; Drexler, H.G. Deregulated NKL Homeobox Genes in B-Cell Lymphoma. Cancers 2019, 11, 1874. [Google Scholar] [CrossRef] [Green Version]
- Kerkhofs, C.; Williams, A.P.; Brunner, H.G.; Faust, S.N.; Rae, W.; Wurm, P.; Fockens, P.; Laass, M.; Kokke, F. Mutations in RPSA and NKX2—3 link development of the spleen and intestinal vasculature. Hum. Mutat. 2020, 41, 196–202. [Google Scholar] [CrossRef] [Green Version]
- Vojkovics, D.; Kellermayer, Z.; Kajtár, B.; Roncador, G.; Vincze, Á.; Balogh, P. Nkx2-3-A Slippery Slope From Development Through Inflammation Toward Hematopoietic Malignancies. Biomark. Insights 2018, 13. [Google Scholar] [CrossRef] [Green Version]
- Kellermayer, Z.; Vojkovics, D.; Dakah, T.A.; Bodó, K.; Botz, B.; Helyes, Z.; Berta, G.; Kajtár, B.; Schippers, A.; Wagner, N.; et al. IL-22-Independent Protection from Colitis in the Absence of Nkx2.3 Transcription Factor in Mice. J. Immunol. 2019, 202, 1833–1844. [Google Scholar] [CrossRef] [Green Version]
- Kuhbandner, K.; Hammer, A.; Haase, S.; Terbrack, E.; Hoffmann, A.; Schippers, A.; Wagner, N.; Hussain, R.Z.; Miller-Little, W.A.; Koh, A.Y.; et al. MAdCAM-1-Mediated Intestinal Lymphocyte Homing Is Critical for the Development of Active Experimental Autoimmune Encephalomyelitis. Front. Immunol. 2019, 10, 903. [Google Scholar] [CrossRef] [Green Version]
- Myint, P.K.; Park, E.J.; Gaowa, A.; Kawamoto, E.; Shimaoka, M. Targeted remodeling of breast cancer and immune cell homing niches by exosomal integrins. Diagn. Pathol. 2020, 15, 38. [Google Scholar] [CrossRef]
- Tarlinton, D.; Light, A.; Metcalf, D.; Harvey, R.P.; Robb, L. Architectural defects in the spleens of Nkx2-3-deficient mice are intrinsic and associated with defects in both B cell maturation and T cell-dependent immune responses. J. Immunol. 2003, 170, 4002–4010. [Google Scholar] [CrossRef] [Green Version]
- Pabst, O. NKX2.3 is required for MAdCAM-1 expression and homing of lymphocytes in spleen and mucosa-associated lymphoid tissue. EMBO J. 2000, 19, 2015–2023. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Parkes, M.; Barrett, J.C.; Prescott, N.J.; Tremelling, M.; Anderson, C.A.; Fisher, S.A.; Roberts, R.G.; Nimmo, E.R.; Cummings, F.R.; Soars, D.; et al. Sequence variants in the autophagy gene IRGM and multiple other replicating loci contribute to Crohn’s disease susceptibility. Nat. Genet. 2007, 39, 830–832. [Google Scholar] [CrossRef] [PubMed]
- Fisher, S.A.; Tremelling, M.; Anderson, C.A.; Gwilliam, R.; Bumpstead, S.; Prescott, N.J.; Nimmo, E.R.; Massey, D.; Berzuini, C.; Johnson, C.; et al. Genetic determinants of ulcerative colitis include the ECM1 locus and five loci implicated in Crohn’s disease. Nat. Genet. 2008, 40, 710–712. [Google Scholar] [CrossRef] [PubMed]
- Robles, E.F.; Mena-Varas, M.; Barrio, L.; Merino-Cortes, S.V.; Balogh, P.; Du, M.-Q.; Akasaka, T.; Parker, A.; Roa, S.; Panizo, C.; et al. Homeobox NKX2-3 promotes marginal-zone lymphomagenesis by activating B-cell receptor signalling and shaping lymphocyte dynamics. Nat. Commun. 2016, 7, 11889. [Google Scholar] [CrossRef] [PubMed]
- Lábadi, A.; Balogh, P. Differential preferences in serosal homing and distribution of peritoneal B-cell subsets revealed by in situ CFSE labeling. Int. Immunol. 2009, 21, 1047–1056. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kugyelka, R.; Prenek, L.; Olasz, K.; Kohl, Z.; Botz, B.; Glant, T.T.; Berki, T.; Boldizsár, F. ZAP-70 Regulates Autoimmune Arthritis via Alterations in T Cell Activation and Apoptosis. Cells 2019, 8, 504. [Google Scholar] [CrossRef] [Green Version]
- Holló, K.; Glant, T.T.; Garzó, M.; Finnegan, A.; Mikecz, K.; Buzás, E. Complex pattern of Th1 and Th2 activation with a preferential increase of autoreactive Th1 cells in BALB/c mice with proteoglycan (aggrecan)-induced arthritis. Clin. Exp. Immunol. 2000, 120, 167–173. [Google Scholar]
- Czömpöly, T.; Lábadi, A.; Kellermayer, Z.; Olasz, K.; Arnold, H.-H.; Balogh, P. Transcription factor Nkx2-3 controls the vascular identity and lymphocyte homing in the spleen. J. Immunol. 2011, 186, 6981–6989. [Google Scholar] [CrossRef] [Green Version]
- Buzás, E.I.; Végvári, A.; Murad, Y.M.; Finnegan, A.; Mikecz, K.; Glant, T.T. T-cell recognition of differentially tolerated epitopes of cartilage proteoglycan aggrecan in arthritis. Cell. Immunol. 2005, 235, 98–108. [Google Scholar] [CrossRef]
- O’Neill, S.K.; Shlomchik, M.J.; Glant, T.T.; Cao, Y.; Doodes, P.D.; Finnegan, A. Antigen-specific B cells are required as APCs and autoantibody-producing cells for induction of severe autoimmune arthritis. J. Immunol. 2005, 174, 3781–3788. [Google Scholar] [CrossRef]
- Gracey, E.; Vereecke, L.; McGovern, D.; Fröhling, M.; Schett, G.; Danese, S.; De Vos, M.; Van den Bosch, F.; Elewaut, D. Revisiting the gut–joint axis: Links between gut inflammation and spondyloarthritis. Nat. Rev. Rheumatol. 2020, 16, 415–433. [Google Scholar] [CrossRef] [PubMed]
- Mikecz, K.; Glant, T.T.; Poole, A.R. Immunity to cartilage proteoglycans in BALB/c mice with progressive polyarthritis and ankylosing spondylitis induced by injection of human cartilage proteoglycan. Arthritis Rheum. 1987, 30, 306–318. [Google Scholar] [CrossRef] [PubMed]
- Olasz, K.; Boldizsar, F.; Kis-Toth, K.; Tarjanyi, O.; Hegyi, A.; van Eden, W.; Rauch, T.A.; Mikecz, K.; Glant, T.T. T cell receptor (TCR) signal strength controls arthritis severity in proteoglycan-specific TCR transgenic mice. Clin. Exp. Immunol. 2012, 167, 346–355. [Google Scholar] [CrossRef] [PubMed]
- Cao, Y.; Brombacher, F.; Tunyogi-Csapo, M.; Glant, T.T.; Finnegan, A. Interleukin-4 regulates proteoglycan-induced arthritis by specifically suppressing the innate immune response. Arthritis Rheum. 2007, 56, 861–870. [Google Scholar] [CrossRef]
- Grosse, J.; Allado, E.; Roux, C.; Pierreisnard, A.; Couderc, M.; Clerc-Urmes, I.; Remen, T.; Albuisson, É.; De Carvalho-Bittencourt, M.; Chary-Valckenaere, I.; et al. ACPA-positive versus ACPA-negative rheumatoid arthritis: Two distinct erosive disease entities on radiography and ultrasonography. Rheumatol. Int. 2020, 40, 615–624. [Google Scholar] [CrossRef]
- Coutant, F. Pathogenic effects of anti-citrullinated protein antibodies in rheumatoid arthritis—Role for glycosylation. Jt. Bone Spine 2019, 86, 562–567. [Google Scholar] [CrossRef]
- Sakaguchi, W.; To, M.; Yamamoto, Y.; Inaba, K.; Yakeishi, M.; Saruta, J.; Fuchida, S.; Hamada, N.; Tsukinoki, K. Detection of anti-citrullinated protein antibody (ACPA) in saliva for rheumatoid arthritis using DBA mice infected with Porphyromonas gingivalis. Arch. Oral Biol. 2019, 108, 104510. [Google Scholar] [CrossRef]
- Tarjanyi, O.; Boldizsar, F.; Nemeth, P.; Mikecz, K.; Glant, T.T. Age-related changes in arthritis susceptibility and severity in a murine model of rheumatoid arthritis. Immun. Ageing 2009, 6, 8. [Google Scholar] [CrossRef] [Green Version]
- Minta, A.; Kao, J.P.; Tsien, R.Y. Fluorescent indicators for cytosolic calcium based on rhodamine and fluorescein chromophores. J. Biol. Chem. 1989, 264, 8171–8178. [Google Scholar]
- Boldizsár, F.; Berki, T.; Miseta, A.; Németh, P. Effect of hyperglycemia on the basal cytosolic free calcium level, calcium signal and tyrosine-phosphorylation in human T-cells. Immunol. Lett. 2002, 82, 159–164. [Google Scholar] [CrossRef]




© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Khanfar, E.; Olasz, K.; Gábris, F.; Gajdócsi, E.; Botz, B.; Kiss, T.; Kugyelka, R.; Berki, T.; Balogh, P.; Boldizsár, F. Ameliorated Autoimmune Arthritis and Impaired B Cell Receptor-Mediated Ca2+ Influx in Nkx2-3 Knock-out Mice. Int. J. Mol. Sci. 2020, 21, 6162. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21176162
Khanfar E, Olasz K, Gábris F, Gajdócsi E, Botz B, Kiss T, Kugyelka R, Berki T, Balogh P, Boldizsár F. Ameliorated Autoimmune Arthritis and Impaired B Cell Receptor-Mediated Ca2+ Influx in Nkx2-3 Knock-out Mice. International Journal of Molecular Sciences. 2020; 21(17):6162. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21176162
Chicago/Turabian StyleKhanfar, Esam, Katalin Olasz, Fanni Gábris, Erzsébet Gajdócsi, Bálint Botz, Tamás Kiss, Réka Kugyelka, Tímea Berki, Péter Balogh, and Ferenc Boldizsár. 2020. "Ameliorated Autoimmune Arthritis and Impaired B Cell Receptor-Mediated Ca2+ Influx in Nkx2-3 Knock-out Mice" International Journal of Molecular Sciences 21, no. 17: 6162. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21176162