1. Introduction
Both cutaneous (non-burn) and burn injuries immediately trigger the local wound healing process. This involves the four complex and overlapping phases of haemostasis, inflammation, proliferation and remodelling [
1,
2]. In contrast to simple cutaneous wounds, major burn injury simultaneously also triggers a systemic pathophysiological stress response and hypermetabolism [
3]. The systemic hypermetabolic response refers to a significant increase in the resting energy expenditure, which is largely driven by elevated levels of circulating catecholamines, glucocorticoids and pro-inflammatory cytokines following burn injury [
4]. A greater than 20% total body surface area (TBSA) full-thickness burn injury stimulates hypermetabolism in humans [
3]. Furthermore, hypermetabolic burn patients also suffer from endocrine dysfunction, immune compromise, insulin resistance and whole-body catabolism [
5,
6]. These clinical features are associated with delayed recovery, prolonged hospital admission and increased morbidity and mortality. Currently available treatments to ameliorate hypermetabolism in major burn patients include local measures to reduce systemic cytokine stimulus (early excision, closure of the wound) as well as nutritional support and pharmacologic modalities such as androgens (testosterone, oxandrolone) and other anabolic hormones (insulin, growth hormone etc.) [
4].
In male mammals, the major circulating androgen is testosterone, which is secreted by testicular Leydig cells. After testicular secretion, a small proportion of circulating testosterone (5~10%) is converted to the more potent androgen, dihydrotestosterone (DHT) by 5α-reductase enzymes [
7,
8]. In previous studies, castration of male mice to reduce circulating androgens, accelerates cutaneous wound healing through the attenuation of the local inflammatory response and increased extracellular matrix (ECM) deposition [
9,
10,
11]. The castration effect on wound healing was counteracted by the restoring androgens in orchiectomised mice [
9,
10,
11]. Delayed wound healing was also reported after chemical blockade of testosterone conversion to DHT or in genetic androgen receptor (AR) knockout mice [
9,
11,
12,
13]. In these studies, androgens play an inhibitory role in cutaneous wound healing through the increased production of IL-6 and TNF and reduced wound re-epithelialization and collagen deposition [
9,
11,
12,
13].
Systemic inflammation is crucial in wound healing by supplying the growth factors, cytokines and chemokines needed to recruit immune cells mainly neutrophil, monocytes and macrophages to the site of burn injury. In cutaneous wounds, the systemic and local inflammatory phase lasts for 5–7 days. However, a major burn may display chronic persistent inflammation with a marked release of abundant pro-inflammatory mediators that linked to inflammatory response syndrome, sepsis, and multi-organ failure in patients.
In contrast to the known inhibitory effects of androgens by prolonging inflammation in cutaneous wound healing, clinical studies have reported that testosterone and the synthetic androgen, oxandrolone can enhance recovery from a burn injury. Oxandrolone-treated burn patients maintain body mass better and have improved body composition and hepatic protein synthesis during the acute post-burn phase [
14,
15,
16,
17,
18]. Although there are reports of improved patient outcomes mainly in treated children (with low circulating androgens) after major burns, the role of androgens in major burn wound healing processes is still unknown.
DHT is the most potent natural androgen, with higher affinity and slower dissociation from AR binding leading to greater molar potency in the transactivation of the AR [
8]. Most importantly, DHT lacks the hepatotoxicity of 17α–alkylated synthetic androgens like oxandrolone, highlighting its overlooked potential as an important therapeutic option for male burn patients. Therefore, we hypothesise that pure androgen, DHT, can promote major burn wound healing via altering post-burn inflammation response. This study aims to investigate DHT treatment as a new therapeutic approach in treating major burn injury.
3. Discussion
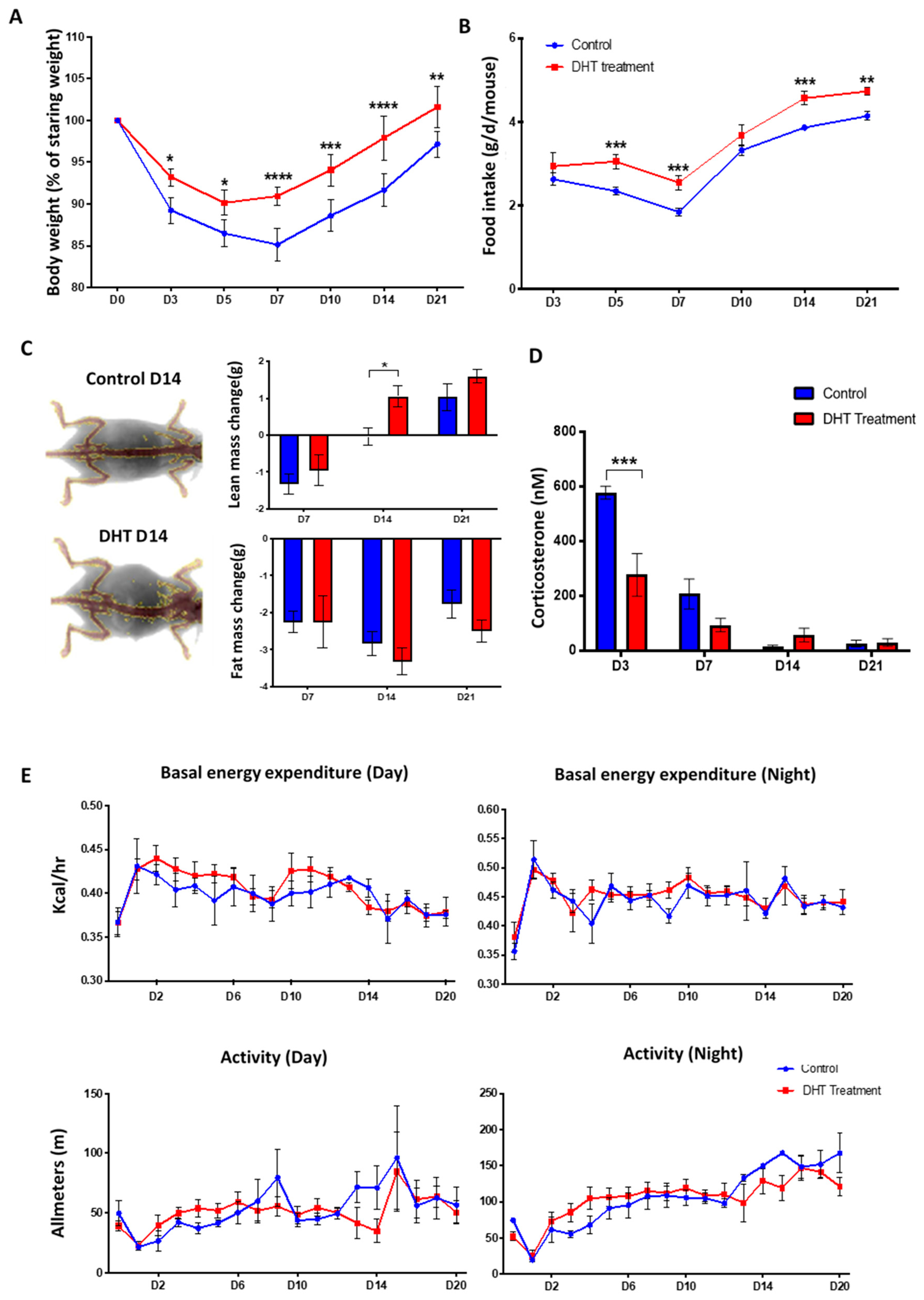
In this study, we examined the role of a pure (non-aromatisable) androgen, DHT, in the local wound healing process and the systemic hypermetabolic response following major burn injury in mice. Our results showed that DHT has a positive impact on both local wound healing and metabolic catabolic responses, which differ from those reported after cutaneous injury.
DHT-treated mice showed a faster healing rate, particularly in the early stages (day 3–14) of the healing process. DHT contributed to these effects by influencing local inflammation, cell proliferation of keratinocytes and ECM production. DHT-treated mice also exhibited a favourable local inflammatory response in the wound area demonstrated by an early increase of pro-inflammation cytokines IL-6 and TNF post-burn on day 7 followed by a rapid reduction on day 14 and 21, which counteracts the elevated level of anti-inflammation cytokine TGFβ-1 (
Figure 1C). Our findings are in contrast with previous studies that show that androgens inhibit cutaneous wound healing by enhancing TNF production and prolonging inflammation following non-penetrating skin injury [
13]. Moreover, burn injury wounds in the DHT treatment group exhibited more connective tissues and collagen production over the remodelling phase. This also contrasts with previous findings that androgens suppress collagen production but contribute to collagen degradation in cutaneous non-burn wound healing [
9,
12,
13]. The different effects of DHT on local wound healing of non-burn cutaneous and major burn injury may be explained by the effect of DHT on systemic responses following burn injury. Major burn injury simultaneously induces a systemic hypermetabolic response in humans [
4] and mice [
19], resulting in a pronounced and prolonged increase in basal energy expenditure (BEE), a greater stress response with higher levels of corticosterone (in mice) and cortisol (in humans) as well as inflammatory cytokines all culminating in profound catabolic consequences [
19].
In severely burned male patients, testicular steroid production is substantially decreased. The reduction in circulating testosterone level contributes greatly to the loss of lean body mass observed in burned patients [
20,
21,
22]. Analogous findings were observed in control burn-injured mice in the current study (
Figure 2A). DHT-treated mice maintained a more stable body weight following injury, with increased lean body mass compared to mice in the control group. The faster recovery of body weight was associated with increased food intake (
Figure 2B). This finding is supported by clinical data that the administration of synthetic 17-α alkylated androgen, oxandrolone, can improve gross parameter of lean body mass, total body mass, bone mineral composition, strength and shorten the length of hospital stay for major burn injury child patents [
4,
15,
16,
18]. Moreover, serum level of corticosterone on days 3 and 7 post-injury was low in the DHT group (
Figure 2D), indicating that DHT can minimise stress response, perhaps contributing to better food intake and body weight recovery as acute stress response has been reported to suppress appetite and food intake [
23,
24]. However, DHT treatment had no effect on basal energy expenditure or movement in burn-injured mice, although DHT is an anabolic hormone that can increase protein synthesis and ameliorate catabolism post burn injury. Similar findings were reported before in clinical studies in which oxandrolone showed no effect on resting expenditure rate or basal metabolic rate in children with burn injuries [
18].
In this study, we have demonstrated an overall increase in inflammatory cytokines and chemokines post-burn injury (
Figure 3) as previously reported clinically [
25,
26,
27,
28]. Significantly increased pro-inflammation cytokines IL-1α, IL-6 and TNF on day 3 in DHT mice (
Figure 1C and
Figure 3) suggested that DHT treatment may play a role in accelerating the turnover of early inflammatory response. While the elevated serum level of the macrophage recruiting cytokines such as IL-12(p40), chemokine RANTES at a later stage of wound healing (
Figure 3) indicating early resolution of inflammation which correlates to the increased cell proliferation and collagen deposition observed.
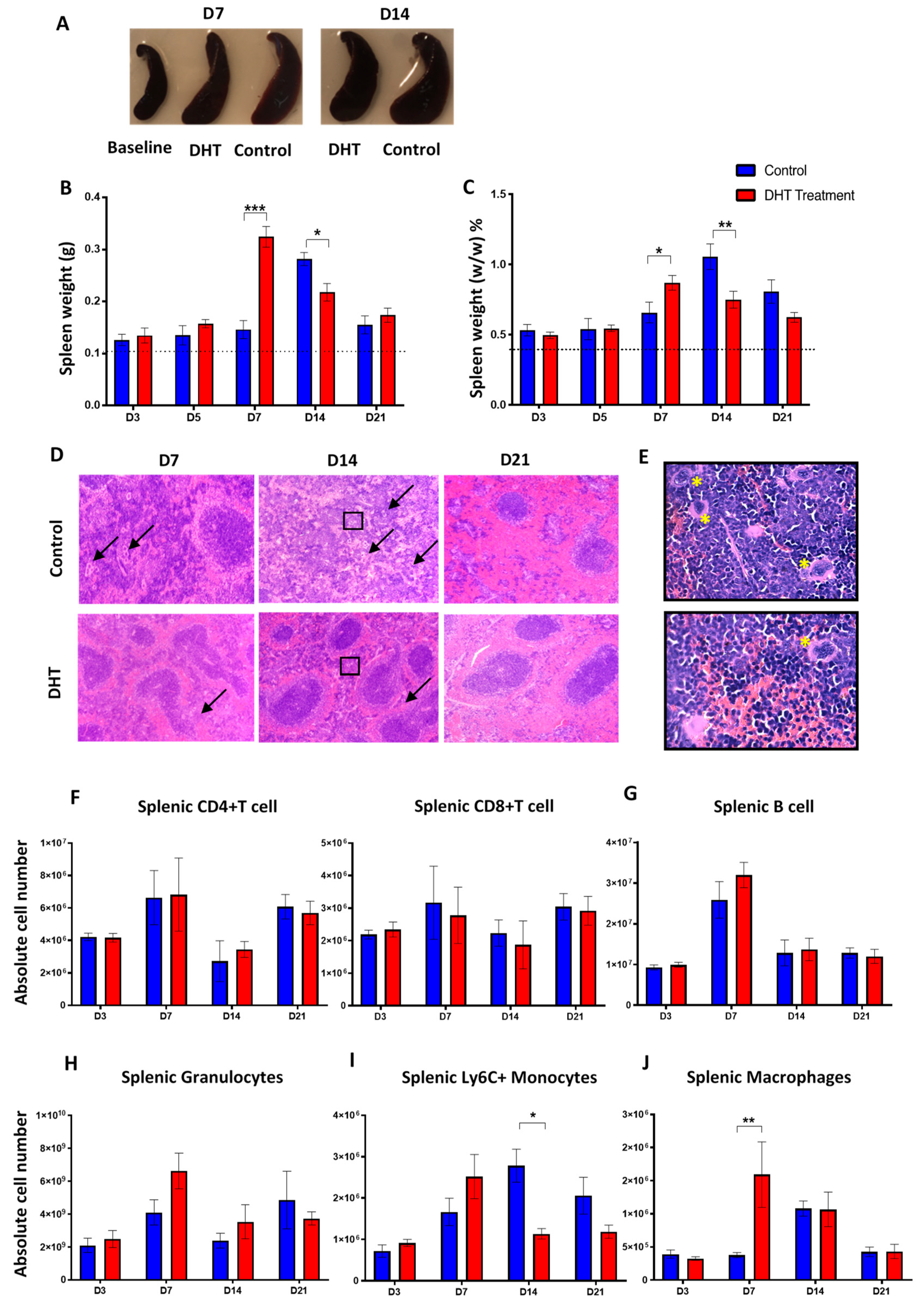
Splenomegaly after major burn injury in both experimental groups was observed, confirming a burn injury-induced systemic inflammation change (
Figure 4A). Enumeration of splenic immune populations demonstrated an increase of granulocytes, monocytes and macrophages in DHT-treated mice 7 days earlier than control animals (
Figure 4H–J). These findings suggested that DHT enhanced systemic inflammation post major injury, resulting in a quick resolution of inflammation. Unlike cutaneous wound healing, during which androgens prolong inflammation phase [
9,
11,
13,
29], our study reported after DHT treatment, the majority of inflammation markers and immune cell numbers dropped to normal level after 14–21 days post major burn injury.
In major burn wound healing, monocytes circulate through the blood-stream and extravasate into wound site during inflammation phase. In addition to the bone marrow, splenic monocytes produced within the subcapsular red pulp can also be recruited to wound site [
30,
31]. This recruitment of pro-inflammatory monocyte is predominantly dependent on MCP-1 and its receptor CCR2 [
31]. A recent study found that circulating monocytes expressing a high level of Ly6C
+ infiltrate into the skin can differentiate into wound repair macrophages expressing F4/80 antigen [
30,
31]. These macrophages contribute to phagocytosing dead tissues, stimulating angiogenesis, promoting granulation tissue formation, secreting extracellular matrix and re-epithelization the wound [
30,
31,
32]. In our study, an elevated circulating MCP-1 concentration was observed in DHT-treated mice on day 3 (
Figure 3), resulting more Ly6C
+ monocytes being mobilised into the circulation and recruited to the spleen (
Figure 4H). This finding correlates with the increased in macrophages in the wound area of the DHT-treated mice (
Figure 5) that facilitate wound healing by enhancing cell proliferation, re-epithelialization and collagen deposition as suggested by previous studies [
33,
34].
Taken together, our findings from the current study shows that burn injuries induced hypermetabolic-catabolic and stress response and resulted in the production of pro-inflammation cytokines including IL-1α, IL-6 and TNF. Thereby DHT acting via AR on neutrophil, T cell, B cell, monocytes and macrophages are able to recruit other immune cells into the circulation via elevated inflammation factors. Circulating immune cells then infiltrate and recruit to the spleen. The increase in MCP-1 level at the wound area and systemically recruits more monocytes to the blood circulation and the wound site. These monocytes are then differentiated to macrophages that are involved in removing bacteria, preventing infection and contributing in collagen disposition once the inflammation is resolved with the increase of anti-inflammation cytokine such as TGFβ-1 (
Figure 6). We show here that DHT treatment induced acceleration of the inflammatory turnover both locally and systemically as the key in promoting major burn wound healing. DHT, a potent natural pure androgen, showed no adverse effects in mice over 21 days of treatment, highlighting its potential as a novel therapeutic approach for treating a major injury.
This research is comprehensive in providing evidence on androgen treatments in major burn wound healing. However, a few limitations of these studies need to be acknowledged. Firstly, burn wound healing studies were conducted only in male mice, and differences in the response to the androgen treatments may exist in female mice. Secondly, due to differences between mouse and human metabolism, wound healing mechanism and skin structure, translational research on the effect of systemic DHT administration needs to be conducted in the clinical settings of a major burn injury. The findings will be valuable to further evaluate its therapeutic potential.
4. Materials and Methods
4.1. Major Burn Injury Model and Wound Healing Experiment
Male BALB/c mice (Animal Resource Centre, Murdoch, WA, Australia) (n = 120) were housed with free access to water, in the specific-pathogen-free Translational Research Facility (TRF) at the ANZAC Research Institute. The environment was closely controlled at 24–26 °C and 44–46% humidity under a 12:12 h light–dark cycle with lights on at 6 am. All protocols were approved by the Sydney Local Health District Animal Welfare Committee (Protocol No. 2018/020) under the Australian National Health and Medical Research Council Guidelines for animal experimentation.
At 12 weeks of age, mice weighted ~22–27 g were anaesthetised with 3% isoflurane and the dorsum of the mouse was shaved. In the treatment group, mice were subdermally implanted with 1 cm silastic tube filled with ~10mg crystalline DHT [
35] in the neck region, while mice in the control group were also implanted with an empty silastic tube.
At the same day of DHT implantation, mice were then subjected to a major burn injury as previously described [
19]. Briefly, a 4 cm
2 contact burn wound (representing ~10% TBSA) was generated using a hot brass rod. The wound site was debrided 48 h later, removing the damaged skin and to avoid infection, prior to wound dressing for 10 days. Wound tissues were harvested at day 3, 5, 7, 14, 21 post debridement (day 0). Wound samples were bisected for histology and immediately snap-frozen in liquid nitrogen for molecular analysis. Androgen sensitive organs (kidney, testes, seminal vesicle), liver, brown adipose tissue, and white adipose tissue were also collected for further analysis. The spleen was weighed and harvested for analysis by flow cytometry. Wound healing rate was quantified using the Visitrack digital instrument (Smith and Nephew).
4.2. Body Composition and Food Intake
Bodyweight and food intake were measured before and after burn injury at days 3, 5, 7, 10, 14, 21. Body compositions were measured using dual-energy X-ray absorptiometry (DEXA, Lunar Piximus, GE Lunar Corp, WI, USA) following the manufacturer’s manual.
4.3. Metabolism
Gas exchange, energy expenditure and movement of mice (n = 4 per group) were individually continuously measured following major burn injury in Promethion metabolic cage systems for 21 days (Sable Systems International, North Las Vegas, NV, USA). Energy expenditure was calculated using the Weir equation: Kcal/h = 60 × (0.003941 × VO2 + 0.001106 × VCO2). Results were analyzed by R and R studio software.
4.4. Histology, Immunocytochemistry, and Image Analysis
Wound tissues were embedded in paraffin. Multiple 5 µm sections were stained with hematoxylin and eosin (H&E) for general histological analysis; Masson’s Trichrome for collagen deposition; proliferating cell nuclear antigen (PCNA) staining for cell proliferating. The total number of positive stained cells/30 cells in 3 random chosen wound areas was counted and examined by two individual researchers. Multiple sections were stained with anti-mouse F4/80–Alexa Fluor (AF) 647 (1:100, BM8; BD Biosciences, NSW, Australia) for detecting macrophage at wound site.
4.5. Real-Time Quantitative Polymerase Chain Reaction
Total RNA from normal skin and wound tissues were extracted with TRI Reagent (Sigma Aldrich) according to the manufacturer’s instructions. Polymerase chain reaction (PCR) primers for IL-6, TNF-α, TGF-β1, Col3α, Col1α1, (
Supplementary Materials Table S1) were designed using Primer-BLAST Software. RT-qPCR was performed using Ssoadvanced Universal SYBR Green Supermix (Bio-Rad, Hercules, CA, USA). The CFX Manager Software was used to obtain cycle threshold (Ct) values for each sample, which were expressed as a fold change relative to expression of 18s RNA housekeeping gene.
4.6. Obtaining Single-Cell Suspensions from Mouse Tissues
Peripheral blood was obtained from mice by tail bleeding. Single-cell suspensions were made from mouse spleen by mechanical disruption through 70-μm cell strainers (Edwards, NSW, Australia) in RPMI 1640 media supplemented with 2% heat-inactivated fetal bovine serum (Thermo Fisher, WI, USA; 2% FCS/RPMI 1640). Red blood cells were removed from spleens with 1 × RC lysis buffer (eBioscience, CA, USA). To enumerate, cells were counted in a fixed volume (10–50 μL) using the ACCURI C6 flow cytometer (BD Biosciences) for spleen samples or XN-450 automated haematology analyzer (Sysmex, NSW, Australia) for peripheral blood.
4.7. Flow Cytometry
Single-cell suspensions were stained in an FACS buffer (0.1% BSA and 2 mM EDTA in PBS (Sigma Aldrich)) using optimised concentrations of Fc Block (2.4G2; BD Biosciences) and then combinations of the following fluorochrome-conjugated anti-mouse monoclonal antibodies (mAbs) for peripheral blood samples: IA-IE (M5/114-15.2) from BD Biosciences, Ly6C (HK1.4) from eBioscience and F4/80 (BM8) from BioLegend (NSW, Australia); for spleen samples: CD11c (N418), CD4 (GK1.5), B220 (RA3-6B2), CD8a (53-6.7), F4/80 (BM8), IA-IE (M5/114-15.2), Ly6C (HK1.4). To assist in identifying rare myeloid populations, cells were also stained with a lineage (Lin) mixture containing biotinylated mAbs: CD3e (145-2C11), CD19 (1D3) from BD Biosciences, and Ly6G (1A8) and NK1.1 (PK136) from BioLegend. Lin mAbs were detected using streptavidin–Brilliant Violet 421 (BD Biosciences) (
Supplementary Materials Figure S1). Data were collected on the ACCURI C6 flow cytometer (BD Biosciences) and analyzed using FlowJo software (Tree Star, version 9).
4.8. Cytokines Expression
The Bio-Plex ProTM Mouse Cytokine 23-plex Assay (Bio-Rad, Hercules, CA, USA) was used to profile the expression of 23 inflammatory mediators interleukin IL-1a, IL-1β, IL-2, IL-3, IL-4, IL-5, IL-6, IL-9, IL-10, IL-12p40, IL-12p70, IL-13, IL-17α, granulocyte colony-stimulating factor (G-CSF), granulocyte-macrophage colony-stimulating factor (GM-CSF), interferon-gamma (IFN-λ), monocyte chemoattractant protein-1 (MCP-1), macrophage inflammatory protein-1 alpha and beta (MIP-1α, MIP-1β) tumour necrosis factor-alpha (TNF-α), Chemokine ligand 5 (RANTES), eotaxin and chemokine ligand 1 (KC). Burn-injured and control mice had serum samples collected at day 3, 7 and 14 and measurements were performed according to the manufacturer’s instructions and analyzed using the Bio-Plex Manager Software (Bio-Rad, Hercules, CA, USA).
4.9. Corticosterone Assay
Blood sampling via tail snips was continuously collected in mice before burn injury and in the morning of day 3, 7, 14 and 21 post-debridement. All blood samples were centrifuged at 2000× g for 5 min at room temperature and stored at −80 °C until further analysis. Plasma corticosterone concentrations were measured using the DetectX® Corticosterone Enzyme Immunoassay Kit (Arbor Assays, Ann Arbor, MI, USA).
4.10. Statistical Analysis
Repeated measurement analysis of variance (ANOVA) was used for all continuous variables including, wound healing rate, body weight changes and food intakes to correct bias caused by multiple observation from each mouse. The data are presented as mean ± SEM and significant differences were determined by two-way analysis of variance (ANOVA) and pair-T test with p ≤ 0.05 accepted for statistical significance. n = 6 per group per time point * p < 0.05, ** p < 0.01, *** p < 0.001.
This work was performed at the ANZAC Research Institute, Concord Hospital.

 ,
, 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}





