Recent Advances on the Model, Measurement Technique, and Application of Single Cell Mechanics


Abstract
:1. Introduction
2. Cell Mechanical Model
2.1. Continuous Cortical Membrane Model
2.1.1. Droplet Model Surrounded by Continuous Elastic Cortex
2.1.2. Solid Model
2.1.3. Damping Model of Power Series Structure
2.1.4. Two-Phase Model
2.2. Discrete Network Skeleton Model
2.2.1. Tension Integration Model
2.2.2. Porous Solid Model
2.2.3. Cable Net Model
2.2.4. Biochemical Mechanical Model
2.3. Cortical Skeleton Combined Model
2.3.1. Cortical Skeleton Combined Model Based on Finite Element Method
2.3.2. Cortical Skeleton Combined Model Based on Molecular Dynamics
3. Research Progress in Experimental Measurement Techniques of Cell Mechanics
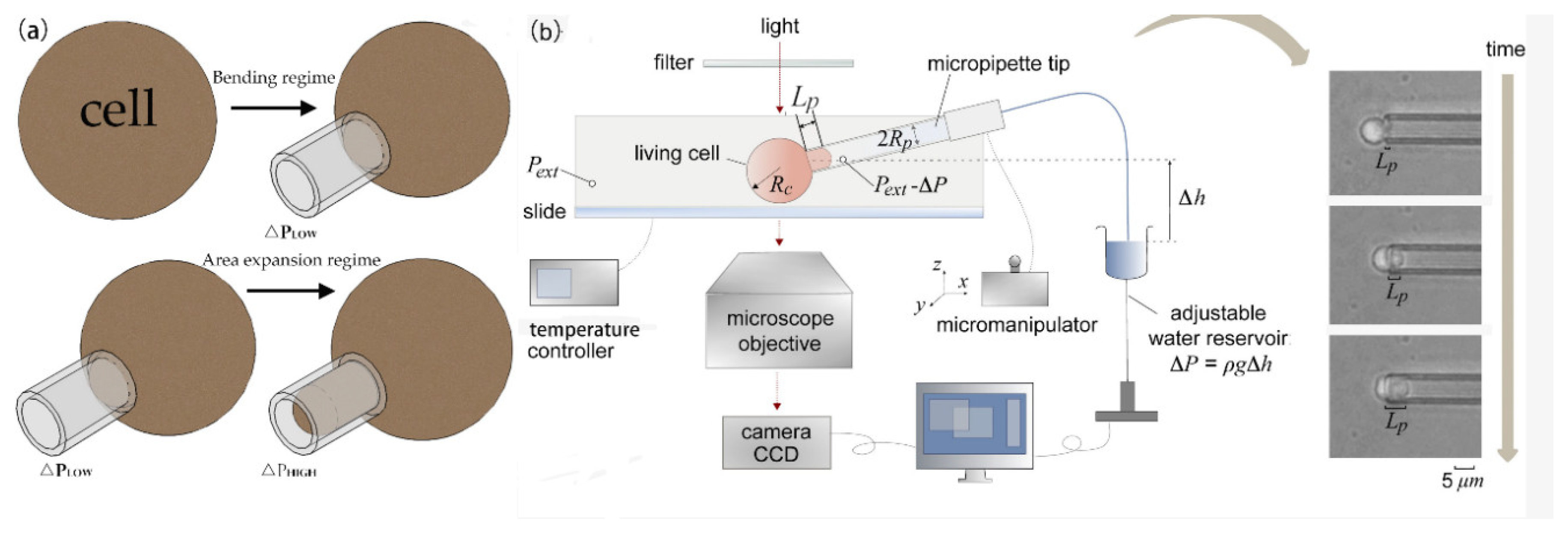
3.1. Micropipette Aspiration Technique
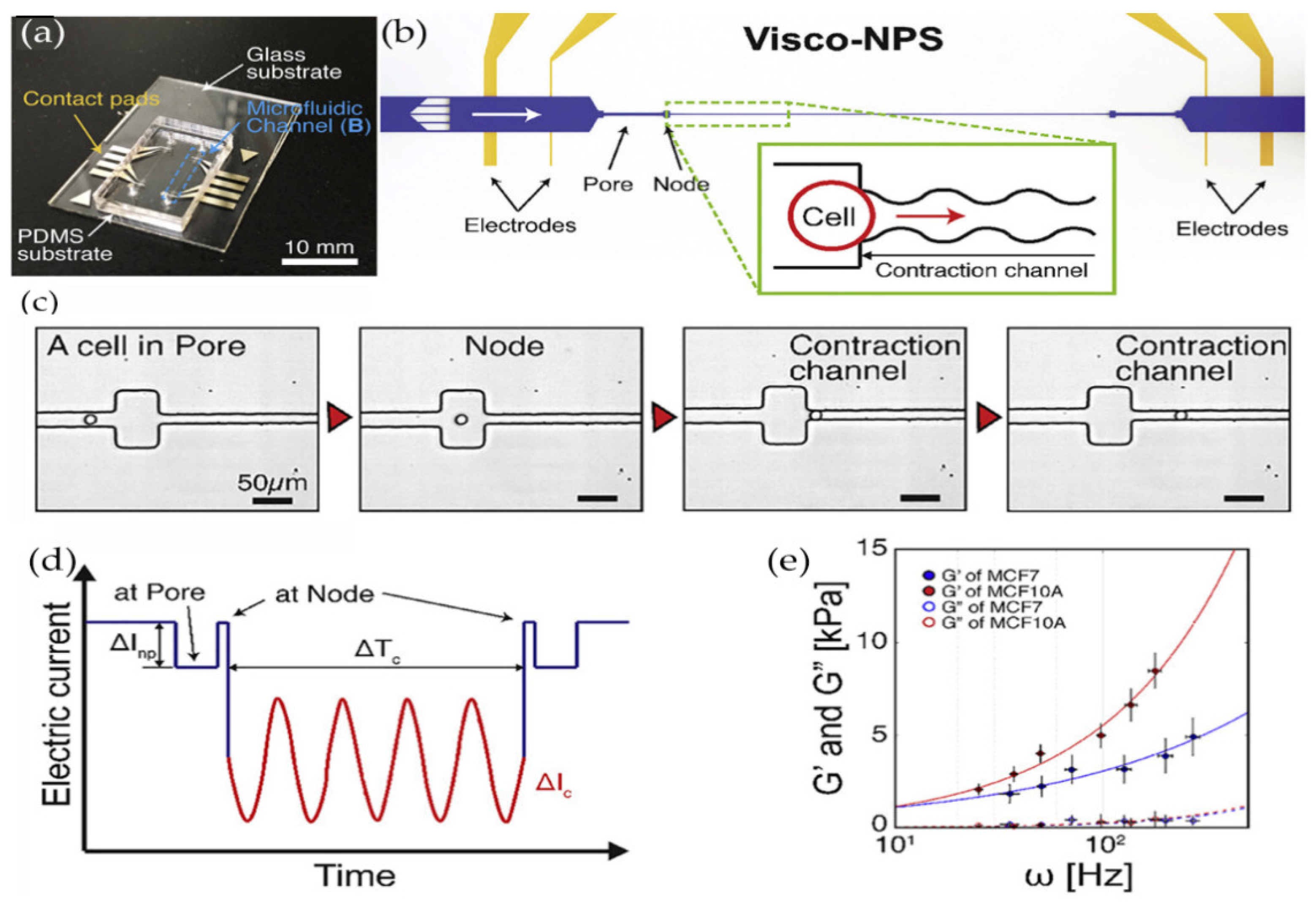
3.2. Microfluidic Technology
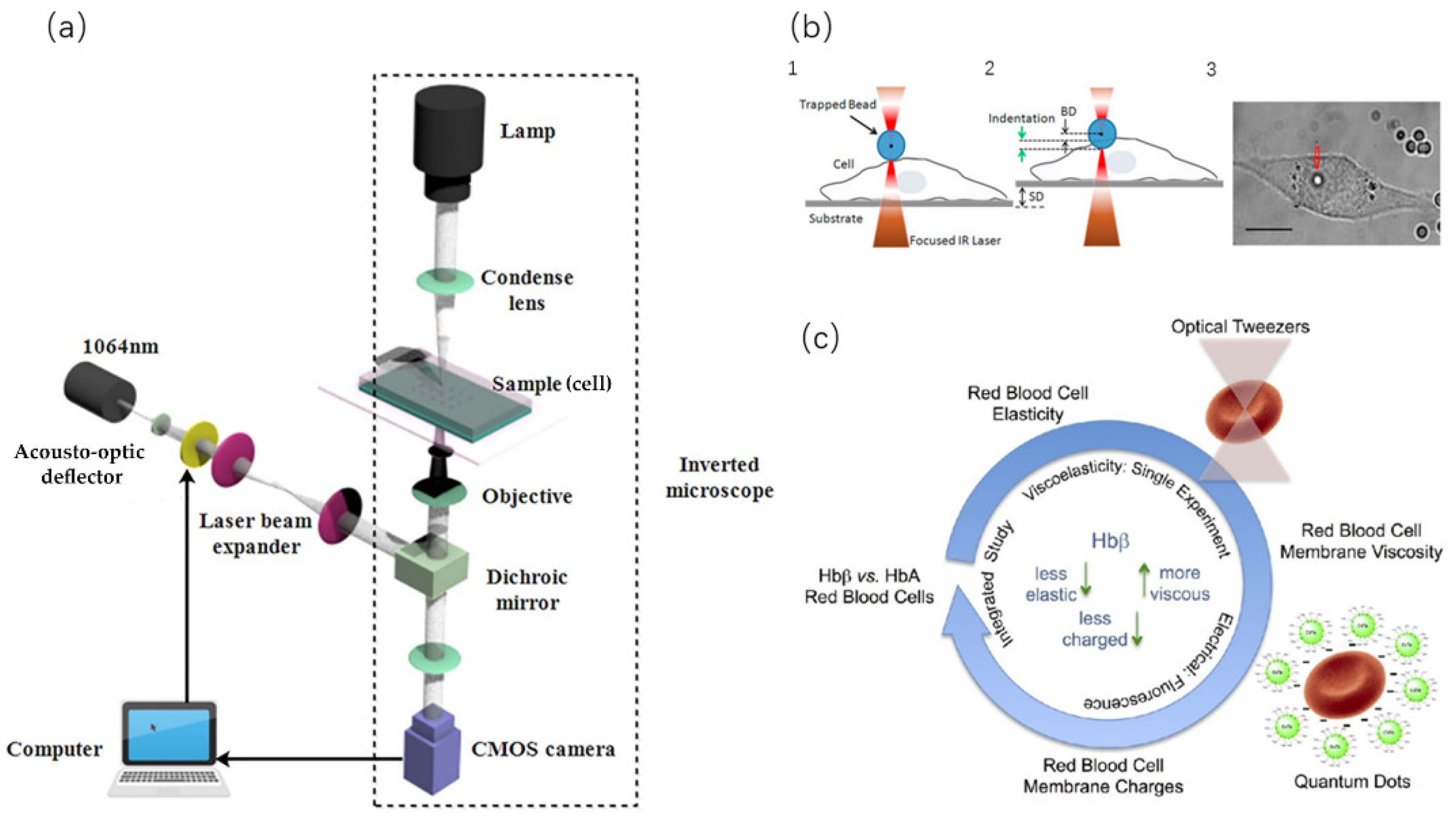
3.3. Optical Tweezer Technology
3.4. Atomic Force Microscope
4. Application Direction
4.1. Disease Diagnosis and Treatment
4.2. Regeneration and Repair of Biological Tissue
4.3. Single Cellular Micromanipulation
4.4. Other Applications
5. Concluding Remarks
Funding
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Biomechanical Terminology | Explanation | Common Formula or Law | Reference |
|---|---|---|---|
| Elasticity | Elasticity refers to the property that an object can recover its original size and shape after deformation. | △F = −k·△x, when the object deforms, the elasticity is directly proportional to the elongation. (Hooke’s law) | [14,19,39,40,43] |
| Viscosity | The resistance of fluid to deformation under shear stress is measured. | τ = µrefers to the velocity gradient along the Y direction, μ is the viscosity coefficient. | [14,54,55,56,58,61,65,69,82,83,109,110,123,143,151,155] |
| Viscoelasticity | The comprehensive properties of viscosity and elasticity of fluid | : total deformation; : ordinary deformation; : the delayed high elastic deformation;: viscous deformation. | [6,18,19,63,112,113,143] |
| Stress (σ) | Internal force per unit area | σ = P/A, ratio of load to section area. | [15,34] |
| Shear stress | The interaction force between two sides of any section (shear plane) | τ = load is parallel to the section. | [13,17,43] |
| Strain (ε) | The local relative deformation of the object under the action of external force and non-uniform temperature field. | , L is the original length and ΔL is the elongation. | [42] |
| Shear strain | The relative shape variable produced by the object during shearing | γ , is the skew angle; when the shear strain is infinitely small, γ = . | [44,85] |
| Elongational strain | The ratio of the change of the line length to the original line length. | The strain produced by an object in tension or compression. | [42,68,84,124,125,148] |
| Young’s modulus (E) | A physical quantity describing the ability of solid materials to resist deformation. | E = σ/ε, the ratio of stress to strain. | [7,17,18,38,39,43,48,69,70,105,106,107,123,125,126,133,134,139,140,141,142,143,149,152,155] |
| Shear modulus (G) | Shear stress characterizes the material’s ability to resist shear strain. | G = τ/γ, the ratio of shear stress to shear strain. | [48,104,110] |
| Poisson’s ratio() | The ratio of the absolute value of the transverse positive strain to the axial positive strain. | Load in elastic range:, is a constant, beyond the elastic range, increases with the increase of stress until 0.5; Relations between E, G, : | [15,16,55] |
| Dynamic viscoelasticity | The viscoelasticity of objects in vibration. | It describes the ratio of stress to strain of an object under dynamic load | [51,52,61] |
| Storage modulus (G’) | The measurement of energy storage in the process of strain cycling and is usually expressed as the real part of the complex modulus. | Complex modulus: =+ j Storage modulus: | [88,151] |
| Loss modulus (G’’) | The degree of energy loss when the material deforms; usually expressed as the imaginary part of the complex modulus. | Loss modulus: | [88,151] |
| Loss tangent(tan) | Reflect the ratio of viscosity and elasticity of material | Loss tangent: tan = | [88,151] |
| Tool | Measuring Force Range | Object Size |
|---|---|---|
| Micropipette aspiration | 0.1– nN | 1– µm |
| Microfluidic technology | 1– pN | 1– µm |
| Optical tweezers technology | 0.01– pN | 10– nm |
| Atomic force microscope | 10– pN | 1– nm |
| Magnetic twisting | 1–100 pN | 0.1–100 µm |
| Microarray method | 1–100 nN | 1– µm |
References
- Lodish, H.; Berk, A.; Kaiser, C.A. Molecular Cell Biology; W.H. Freeman and Company: New York, NY, USA, 2004. [Google Scholar]
- Butler D., L.; Goldstein, S.A.; Guilak, F. Functional Tissue Engineering: The Role of Biomechanics. J. Biomech. Eng. 2000, 122, 570–575. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- SëMa, K.; Hammad, N.C. Mechanical model of geometric cell and topological algorithm for cell dynamics from single-Cell to formation of monolayered tissues with pattern. PLoS ONE 2015, 10, e0126484. [Google Scholar]
- Sun, B. Advances in Cell Mechanics; Li, S., Sun, B., Eds.; Higher Education Press: Beijing, China, 2011. [Google Scholar]
- Lee, G.Y.H.; Lim, C.T. Biomechanics approaches to studying human diseases. Trends Biotechnol. 2007, 25, 111–118. [Google Scholar] [CrossRef] [PubMed]
- Tomaiuolo, G. Biomechanical properties of red blood cells in health and disease towards microfluidics. Biomicrofluidics 2014, 8, 89–110. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Offroy. M.; Razafitianamaharavo. A.; Beaussart. A. Fast automated processing of AFM PeakForce curves to evaluate spatially resolved Young modulus and stiffness of turgescent cells. RSC Adv. 2020, 10, 19258–19275. [Google Scholar] [CrossRef]
- Lu, Z.; Zhang, X.; Leung, C. Robotic ICSI (Intracytoplasmic sperm injection). IEEE Trans. Biomed. Eng. 2011, 58, 2102–2108. [Google Scholar]
- Bo, L.; Waugh, R.E. Determination of bilayer membrane bending stiffness by tether formation from giant, thin-walled vesicles. Biophys. J. 1989, 55, 509–517. [Google Scholar] [CrossRef] [Green Version]
- Los, D.A.; Murata NLos, D.A.; Murata, N. Membrane fluidity and its roles in the perception of environmental signals. Biochim. Biophys. Acta 2004, 1666, 142–157. [Google Scholar] [CrossRef] [Green Version]
- Shepherd, V.A. The cytomatrix as a cooperative system of macromolecular and water networks. Curr. Top. Dev. Biol. 2006, 75, 171–223. [Google Scholar]
- Fang, S.; Dan, L.; Ting, Y.U. Vinblastine differs from Taxol as it inhibits the malignant phenotypes of NSCLC cells by increasing the phosphorylation of Op18/stathmin. Oncol. Rep. 2017, 37, 2481–2489. [Google Scholar]
- Tartibi, M.; Komvopoulos, K. Characterization of the Mechanical Behavior of Cell Components with an Atomic Force Microscope. In Proceedings of the Stle/asme International Joint Tribology Conference, San Francisco, CA, USA, 17–20 October 2010; pp. 53–55. [Google Scholar]
- Evans, E.; Yeung, A. Apparent viscosity and cortical tension of blood granulocytes determined by micropipet aspiration. Biophys. J. 1989, 56, 151–160. [Google Scholar] [CrossRef] [Green Version]
- Jingjie, H.; Yuxiao, Z.; Obayemi, J.D. An Investigation of the Viscoelastic Properties and the Actin Cytoskeletal Structure of Triple egative Breast Cancer Cells. J. Mech. Behav. Biomed. Mater. 2018, 86, 1–13. [Google Scholar]
- Feric, M.; Broedersz, C.P.; Brangwynne, C.P. Soft viscoelastic properties of nuclear actin age oocytes due to gravitational creep. Sci. Rep. 2015, 5, 16607. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guilak, F.; Tedrow, J.R.; Burgkart, R. Viscoelastic Properties of the Cell Nucleus. Biochem. Biophys. Res. Commun. 2000, 269, 781–786. [Google Scholar] [CrossRef] [PubMed]
- Janmey, P.; Fletcher, D.; Reinhart-King, C. Stiffness sensing by cells. Physiol. Rev. 2020, 100, 695–724. [Google Scholar] [CrossRef] [PubMed]
- Mijailovich, S.M.; Kojic, M.; Zivkovic, M. A finite element model of cell deformation during magnetic bead twisting. J. Appl. Physiol. 2002, 93, 1429–1436. [Google Scholar] [CrossRef] [Green Version]
- Schmid-Schunbein, G.W.; Sung, K.L.; Tozeren, H. Passive mechanical properties of human leukocytes. Biophys. J. 1981, 36, 243–256. [Google Scholar] [CrossRef] [Green Version]
- Mehrayin, M.; Farmanzad, F.; Mozafari, M. A critical stress model for cell motility. Theor. Biol. Med. Model. 2012, 9, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Darling, E.M.; Topel, M.; Zauscher, S. Viscoelastic properties of human mesenchymally-derived stem cells and primary osteoblasts, chondrocytes, and adipocytes. J. Biomech. 2008, 41, 454–464. [Google Scholar] [CrossRef] [Green Version]
- Alcaraz, J.; Buscemi, L.; Grabulosa, M. Microrheology of Human Lung Epithelial Cells Measured by Atomic Force Microscopy. Biophys. J. 2003, 84, 2071–2079. [Google Scholar] [CrossRef] [Green Version]
- Marina PD, M.; Mireia, G.; Jordi, A. Measurement of cell microrheology by magnetic twisting cytometry with frequency domain demodulation. J. Appl. Physiol. 2001, 91, 1152–1159. [Google Scholar]
- Butler, J.P.; Fabry, B.; Glogauer, M. Scaling the Microrheology of Living Cells. Phys. Rev. Lett. 2001, 87, 148102. [Google Scholar]
- Fabry, B.; Maksym, G.N.; Butler, J.P. Time scale and other invariants of integrative mechanical behavior in living cells. Phys. Rev. E 2003, 68, 041914. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shin, D.; Athanasiou, K. Cytoindentation for obtaining cell biomechanical properties. J. Orthop. Res. 1999, 17, 880–890. [Google Scholar] [CrossRef] [PubMed]
- Ingber, D.E. Cellular tensegrity: Defining new rules of biological design that govern the cytoskeleton. J. Cell Sci. 1993, 104, 613–627. [Google Scholar] [PubMed]
- Ingber, D. Mechanobiology and diseases of mechanotransduction. Ann. Med. 2003, 35, 564–577. [Google Scholar] [CrossRef] [PubMed]
- Sultan, C.; Dimitrije, S.I.; Donald, E. A computational tensegrity model predicts dynamic rheological behaviors in living cells. Ann. Biomed. Eng. 2004, 32, 520–530. [Google Scholar] [CrossRef]
- Wang, L.; Chen, W. Modelling Cell origami via a tensegrity model of the cytoskeleton in adherent cells. Appl. Bionics Biomech. 2019, 2019, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Alippi, A.; Bettucci, A.; Biagioni, A. Non linear behaviour of cell tensegrity models. AIP Conf. Proc. 2012, 1433, 329–332. [Google Scholar]
- Ingber, D.E.; Wang, N.; Stamenovi, D. Tensegrity, cellular biophysics, and the mechanics of living systems. Rep. Prog. Phys. 2014, 77, 046603. [Google Scholar] [CrossRef] [Green Version]
- Bennett, A.W. Origin of species by means of natural selection or the preservation of favoured races in the struggle for life. Am. Anthropol. 1979, 61, 176–177. [Google Scholar] [CrossRef] [Green Version]
- Satcher, R.L.; Dewey, C.F. Theoretical estimates of mechanical properties of the endothelial cell cytoskeleton. Biophys. J. 1996, 71, 109–118. [Google Scholar] [CrossRef] [Green Version]
- Stamenovi, D.; Ingber, D.E. Models of cytoskeletal mechanics of adherent cells. Biomech. Model. Mechanobiol. 2002, 1, 95–108. [Google Scholar] [CrossRef] [PubMed]
- Deshpande, V.S.; McMeeking, R.M.; Evans, A.G. A bio-chemo-mechanical model for cell contractility. Proc. Natl. Acad. Sci. USA 2006, 103, 14015–14020. [Google Scholar] [CrossRef] [Green Version]
- Truong, D.; Bahls, C.R.; Nebe, B. Simulation of actin distribution of osteoblasts on titanium pillar arrays using a bio-chemo-mechanical model. Int. J. Numer. Methods Biomed. Eng. 2018, 34, e3097. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mcgarry, J.G.; Prendergast, P.J. A three-dimensional finite element model of an adherent eukaryotic cell. Eur. Cells Mater. 2004, 7, 27–34. [Google Scholar] [CrossRef]
- Xue, F.; Lennon, A.B.; Mckayed, K.K. Effect of membrane stiffness and cytoskeletal element density on mechanical stimuli within cells: An analysis of the consequences of ageing in cells. Comput. Methods Biomech. Biomed. Eng. 2015, 18, 468–476. [Google Scholar] [CrossRef] [Green Version]
- Vassaux, M.; Milan, J.L. Stem cell mechanical behaviour modelling: Substrate’s curvature influence during adhesion. Biomech. Modeling Mechanobiol. 2017, 16, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Deepak, B.Y.; Takeo, M.; Kazuaki, N. A finite element bendo-tensegrity model of eukaryotic cell. J. Biomech. Eng. 2018, 140, 101001–101010. [Google Scholar]
- Barreto, S.; Clausen, C.H.; Perrault, C.M. A multi-structural single cell model of force-induced interactions of cytoskeletal components. Biomaterials 2013, 34, 6119–6126. [Google Scholar] [CrossRef] [Green Version]
- Barreto, S.; Perrault, C.M. D Structural finite element analysis to explain cell mechanics variability. J. Mech. Behav. Biomed. Mater. 2014, 38, 219–231. [Google Scholar] [PubMed]
- Kirill, L.; Yasaman, N.; Menglin, S. Probing eukaryotic cell mechanics via mesoscopic simulations. PLoS Comput. Biol. 2017, 13, 1–22. [Google Scholar]
- Warren, P.B. Dissipative particle dynamics. Curr. Opin. Colloid Interface Sci. 1998, 3, 620–624. [Google Scholar]
- Liu, F.; Wu, D.; Wu, X.; Ken, C. Analyses of the cell mechanical damage during microinjection. Soft Matter. 2015, 11, 1434–1442. [Google Scholar]
- Vliet KJ, V.; Bao, G.; Suresh, S. The biomechanics toolbox: Experimental approaches for living cells and biomolecules. Acta Mater. 2003, 51, 5881–5905. [Google Scholar] [CrossRef]
- Lim, C.T.; Zhou, E.H.; Li, A. Experimental techniques for single cell and single molecule biomechanics. Mater. Sci. Eng. C 2006, 26, 1278–1288. [Google Scholar]
- Bausch, A.R.; Moller, W.; Sackmann, E. Measurement of local viscoelasticity and forces in living cells by magnetic tweezers. Biophys. J. 1999, 76, 573–579. [Google Scholar]
- Brown, T.D. Techniques for Mechanical Stimulation of Cells in vitro: A Review. J. Biomech. 2000, 33, 3–14. [Google Scholar] [CrossRef]
- Rand, R.P.; Burton, A.C. Mechanical properties of the red cell membrane. Biophys. J. 1964, 4, 115–135. [Google Scholar] [CrossRef]
- Cheng, Q.H.; Liu, P.; Gao, H.J. A computational modeling for micropipette-manipulated cell detachment from a substrate mediated by receptor-ligand binding. J. Mech. Phys. Solids 2009, 57, 205–220. [Google Scholar]
- Shao, J.Y.; Xu, J. A modified micropipette aspiration technique and its application to tether formation from human neutrophils. J. Biomech. Eng. 2002, 124, 388–396. [Google Scholar] [CrossRef] [PubMed]
- Gao, Z.; Wang, S.; Zhu, H. Using selected uniform cells in round shape with a micropipette to measure cell adhesion strength on silk fibroin-based materials. Mater. Sci. Eng. C 2008, 28, 1227–1235. [Google Scholar] [CrossRef]
- Ruogang, Z.; Krista, L.S.; Simmons, C.A. Measurement of layer-specific mechanical properties in multilayered biomaterials by micropipette aspiration. Acta Biomater. 2011, 7, 1220–1227. [Google Scholar]
- Ding, Y.; Wang, G.; Feng, X. Micropipette aspiration method for characterizing biological materials with surface energy. J. Biomech. 2018, 80, 32–36. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guevorkian, K.; Maître, J.L. Micropipette aspiration: A unique tool for exploring cell and tissue mechanics in vivo. Methods Cell Biol. 2017, 139, 187–201. [Google Scholar] [PubMed]
- Trickey, W.R.; Baaijens FP, T.; Laursen, T.A. Determination of the poisson’s ratio of the cell: Recovery properties of chondrocytes after release from complete micropipette aspiration. J. Biomech. 2006, 39, 78–87. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Li, Y.; Zhang, Q. Mechanical properties of chondrocytes estimated from different models of micropipette aspiration. Biophys. J. 2019, 116, 2181–2194. [Google Scholar] [CrossRef]
- Sliogeryte, K.; Thorpe, S.D.; Wang, Z. Differential effects of LifeAct-GFP and actin-GFP on cell mechanics assessed using micropipette aspiration. J. Biomech. 2016, 49, 310–317. [Google Scholar] [CrossRef] [Green Version]
- Hogan, B.; Babataheri, A.; Hwang, Y. Characterizing cell adhesion by using micropipette aspiration. Biophys. J. 2015, 109, 209–219. [Google Scholar] [CrossRef] [Green Version]
- Guilak, F.; Mow, V.C. The mechanical environment of the chondrocyte: A biphasic finite element model of cell-matrix interactions in articular cartilage. J. Biomech. 2001, 33, 1663–1673. [Google Scholar] [CrossRef]
- Vaziri, A.; Mofrad MR, K. Mechanics and deformation of the nucleus in micropipette aspiration experiment. J. Biomech. 2007, 40, 2053–2062. [Google Scholar] [CrossRef] [PubMed]
- González-Bermúdez, B.; Guinea, G.V.; Plaza, G.R. Advances in micropipette aspiration: Applications in Cell Biomechanics, Models, and Extended Studies. Biophys. J. 2019, 116, 587–594. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gerecsei, T.; Erdődi, I.; Peter, B. Adhesion force measurements on functionalized microbeads: An in-depth comparison of computer controlled micropipette and fluidic force microscopy. J. Colloid Interface Sci. 2019, 555, 245–253. [Google Scholar] [CrossRef]
- Hirano, Y.; Kowata, K.; Kodama, M. Development of a scanning electrochemical microscopy-based micropipette and its application to analysis of topographic change of single-cell. Bioelectrochemistry 2013, 92, 1–5. [Google Scholar] [CrossRef] [PubMed]
- Gangotra, A.; Willmott, R.G. Mechanical properties of bovine erythrocytes derived from ion current measurements using micropipettes. Bioelectrochemistry 2019, 128, 204–210. [Google Scholar] [CrossRef] [PubMed]
- Daza, R.B. Comparison of cell mechanical measurements provided by Atomic Force Microscopy (AFM) and Micropipette Aspiration (MPA). J. Mech. Behav. Biomed. Mater. 2019, 95, 103–115. [Google Scholar] [CrossRef]
- Mohammadkarim, A.; Mokhtari-Dizaji, M.; Kazemian, A. The mechanical characteristics of human endothelial cells in response to single ionizing radiation doses by using micropipette aspiration technique. Mol. Cell. Biomech. 2019, 16, 275–287. [Google Scholar] [CrossRef]
- Liu, W.M.; Li, L.I.; Ren, L. Diversification of microfluidic chip for applications in cell-based bioanalysis. Chin. J. Anal. Chem. 2012, 40, 24–31. [Google Scholar] [CrossRef]
- Alam, M.K.; Koomson, E.; Zou, H. Recent advances in microfluidic technology for manipulation and analysis of biological cells (2007–2017). Anal. Chim. Acta 2018, 1044, 29–65. [Google Scholar] [CrossRef]
- Chen, H.; Yu, Z.; Bai, S. Microfluidic models of physiological or pathological flow shear stress for cell biology, disease modeling and drug development. Trac Trends Anal. Chem. 2019, 117, 186–199. [Google Scholar] [CrossRef]
- Kelsey, M.; Stroka, M.K.G. Vascular endothelial cell mechanosensing: New insights gained from biomimetic microfluidic models. Semin. Cell Dev. Biol. 2017, 71, 106–117. [Google Scholar]
- Whitesides, G.M. The origins and future of microfluidics. Nature 2006, 442, 368–373. [Google Scholar] [CrossRef] [PubMed]
- El, J.A.; Sorger, P.K.; Jensen, K.F. Cells on chips. Nature 2006, 442, 403–411. [Google Scholar]
- Huang, J.; Lin, F.; Xiong, C. Mechanical characterization of single cells based on microfluidic techniques. Trac Trends Anal. Chem. 2019, 117, 47–57. [Google Scholar] [CrossRef]
- Wang, S.; Sohrabi, S.; Xu, J. Geometry design of herringbone structures for cancer cell capture in a microfluidic device. Microfluid. Nanofluid. 2016, 20, 148. [Google Scholar] [CrossRef]
- Weihua Fan Xi Chen Yuqing, G.e. Single-cell impedance analysis of osteogenic differentiation by droplet-based microfluidics. Biosens. Bioelectron. 2019, 145, 111730. [Google Scholar]
- Jhaveri, D. Study of cell mechanics and surface rheology of cancer cells using high throughput microfluidic device–eScholarship. Master’s Thesis, University of California, Irvine, CA, USA, 2015; p. 1591961. [Google Scholar]
- Ye, T.; Shi, H.; Phan-Thien, N. Numerical design of a microfluidic chip for probing mechanical properties of cells. J. Biomech. 2019, 84, 103–112. [Google Scholar] [CrossRef]
- Alapan, Y.; Matsuyama, Y.; Little, J.A. Dynamic deformability of sickle red blood cells in microphysiological flow. Technology 2016, 04, 71–79. [Google Scholar] [CrossRef] [Green Version]
- Guillou, L.; Joanna, B.; Dahl, J.M.; Lin, G. Measuring cell viscoelastic properties using a microfluidic extensional flow device. Biophys. J. 2016, 111, 2039–2050. [Google Scholar] [CrossRef] [Green Version]
- Solis-Tinoco, V.; Marquez, S.; Quesada-Lopez, T.; Villarroya, F. Building of a flexible microfluidic plasmo-nanomechanical biosensor for live cell analysis. Sens. Actuators B Chem. 2019, 291, 48–57. [Google Scholar] [CrossRef] [Green Version]
- Qin-Qin, J.; Guan-Sheng, D.; Van Uden, M.J. Microfluidic cytometer based on dual photodiode detection for cell size and deformability analysis. Talanta 2013, 111, 178–182. [Google Scholar]
- Spuul, P.; Chi, P.E.; Billottet, C. Microfluidic devices for the study of actin cytoskeleton in constricted environments: Evidence for podosome formation in endothelial cells exposed to a confined slit. Methods 2016, 94, 65–74. [Google Scholar] [CrossRef] [PubMed]
- Soffe, R.; Baratchi, S.; Nasabi, M. Lateral trapezoid microfluidic platform for investigating mechanotransduction of cells to spatial shear stress gradients. Sens. Actuators B Chem. 2017, 251, 963–975. [Google Scholar] [CrossRef]
- Kim, J.; Li, B.; Scheideler, O.J. Visco-node-pore Sensing: A microfluidic rheology platform to characterize viscoelastic properties of epithelial cells. Iscience 2019, 13, 214–228. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Badman, R.P.; Ye, F.; Wang, D.M. Towards biological applications of nanophotonic tweezers. Curr. Opin. Chem. Biol. 2019, 53, 158–166. [Google Scholar] [CrossRef] [PubMed]
- Kamal, R.; Dhakal, V.L. Chapter One–Optical tweezers: Fundamentals and some biophysical applications. In Progress in Optics; Taco, D.V., Ed.; Elsevier: Cambridge, MA, USA, 2018; Volume 1, pp. 1–31. [Google Scholar]
- Shabestari, M.H.; Meijering, A.E.C. Chapter Four–Recent advances in biological single-molecule applications of optical tweezers and fluorescence microscopy. In Methods in Enzymology; Spies, M., Chemla, Y.R., Eds.; Academic Press: Cambridge, MA, USA, 2017; Volume 582, pp. 85–119. [Google Scholar]
- Timo, A.; Du Preez-Wilkinson, N.N. Optical tweezers: Theory and modelling. J. Quant. Spectrosc. Radiat. Transf. 2014, 146, 59–80. [Google Scholar]
- Ashkin, A. Acceleration and trapping of particles by radiation pressure. Phys. Rev. Lett. 1970, 24, 156–159. [Google Scholar] [CrossRef] [Green Version]
- Ashkin, A.; Dziedzic, J. Optical trapping and manipulation of viruses and bacteria. Science 1987, 235, 1517–1520. [Google Scholar] [CrossRef]
- Li, X.; Cheah, C.C.; Hu, S.; Sun, D. Dynamic trapping and manipulation of biological cells with optical tweezers. Automatica 2013, 49, 1614–1625. [Google Scholar] [CrossRef]
- Li, Y.; Guo, Z.; Qu, S. Living cell manipulation in a microfluidic device by femtosecond optical tweezers. Opt. Lasers Eng. 2014, 55, 150–154. [Google Scholar] [CrossRef]
- Li, X.; Yang, H.; Wang, J.; Sun, D. Design of a robust unified controller for cell manipulation with a robot-aided optical tweezers system. Automatica 2015, 55, 279–286. [Google Scholar] [CrossRef]
- Li, X.; Yang, H.; Huang, H.; Sun, D. A switching controller for high speed cell transportation by using a robot-aided optical tweezers system. Automatica 2018, 89, 308–315. [Google Scholar] [CrossRef]
- Cao, B.; Kelbauskas, L.; Chan, S. Rotation of single live mammalian cells using dynamic holographic optical tweezers. Opt. Lasers Eng. 2017, 92, 70–75. [Google Scholar]
- Zhang, Y.; Li, Y.; Zhang, Y.; Hu, C. HACF-based optical tweezers available for living cells manipulating and sterile transporting. Opt. Commun. 2018, 427, 563–566. [Google Scholar] [CrossRef]
- Isaac, C.D.L.; Armstrong, D.J.; Alexander, B. Orientation of swimming cells with annular beam optical tweezers. Opt. Commun. 2020, 459, 124864. [Google Scholar]
- Seeger, S.; Monajembashi, S.; Hutter, K.J. Application of laser optical tweezers in immunology and molecular genetics. Cytometry 1991, 12, 497–504. [Google Scholar] [CrossRef]
- Polimeno, P.; Magazzù, A.; Iatì, M.A. Optical tweezers and their applications. J. Quant. Spectrosc. Radiat. Transf. 2018, 218, 131–150. [Google Scholar] [CrossRef] [Green Version]
- Henon, S.; Lenormand, G.; Richert, A. A new determination of the shear modulus of the human erythrocyte membrane using optical tweezers. Biophys. J. 1999, 76, 1145–1151. [Google Scholar] [CrossRef] [Green Version]
- Muhammad, S.Y.; Ndoye, F.; Coceano, G. Substrate-dependent cell elasticity measured by optical tweezers indentation. Opt. Lasers Eng. 2016, 76, 27–33. [Google Scholar]
- Dagro, A.; Rajbhandari, L.; Orrego, S. Quantifying the local mechanical properties of cells in a fibrous three-dimensional microenvironment. Biophys. J. 2019, 117, 817–828. [Google Scholar] [CrossRef]
- Muhammad, S.Y.; Ndoye, F.; Coceano, G.; Bonin, S. Investigating the effect of cell substrate on cancer cell stiffness by optical tweezers. Opt. Lasers Eng. 2017, 60, 266–269. [Google Scholar]
- Lee, K.; Kinnunen, M.; Danilina, A.V.; Ustinov, V.D. Priezzhev. Characterization at the individual cell level and in whole blood samples of shear stress preventing red blood cells aggregation. J. Biomech. 2016, 49, 1021–1026. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.; Sun, D.; Huang, W. Mechanical force characterization in manipulating live cells with optical tweezers. J. Biomech. 2011, 44, 741–746. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Zhang, F.; Zhu, L. Mechanical properties of RBCs under oxidative stress measured by optical tweezers. Opt. Commun. 2019, 442, 56–59. [Google Scholar] [CrossRef]
- Crick, A.J.; Theron, M.; Tiffert, T. Quantitation of malaria parasite-erythrocyte cell-cell interactions using optical tweezers. Biophys. J. 2014, 107, 846–853. [Google Scholar] [CrossRef] [Green Version]
- Lima, C.N.; Moura, D.S.; Silva, Y.S.S. Evaluating viscoelastic properties and membrane electrical charges of red blood cells with optical tweezers and cationic quantum dots–applications to β-thalassemia intermedia hemoglobinopathy. Colloids Surf. B Biointerfaces 2020, 186, 110671. [Google Scholar] [CrossRef]
- Yu, L.; Sheng, Y. Effect of the object 3D shape on the viscoelastic testing in optical tweezers. Opt. Express 2015, 23, 6020–6028. [Google Scholar] [CrossRef]
- Tan, Y.; Sun, D.; Huang, W. Mechanical characterization of human red blood cells by robotic manipulation with optical tweezers. In Proceedings of the IEEE Press International Conference on Robotics & Biomimetics, Guilin, China, 19–23 December 2009. [Google Scholar]
- Tan, Y.; Sun, D.; Wang, J. Mechanical characterization of human red blood cells under different osmotic conditions by robotic manipulation with optical tweezers. IEEE Trans. Biomed. Eng. 2010, 57, 1816–1825. [Google Scholar] [CrossRef]
- Bareil, P.B.; Sheng, Y.; Chen, Y.Q. Calculation of spherical red blood cell deformation in a dual-beam optical stretcher. Opt. Express 2007, 15, 16029–16034. [Google Scholar] [CrossRef]
- Bui, A.A.M.; Stilgoe, B.A. Theory and practice of simulation of optical tweezers. J. Quant. Spectrosc. Radiat. Transf. 2017, 195, 66–75. [Google Scholar] [CrossRef] [Green Version]
- Johnson, D.; Oatley-Radcliffe, D.L.; Hilal, N. Atomic Force Microscopy. In Membrane Characterization; Hilal, N., Fauzi, A.I., Matsuura, T., Oatley-Radcliffe, D., Eds.; Wiley: New York, NY, USA, 2017; Volume 7, pp. 115–144. [Google Scholar]
- Chang, Y.-R.; Raghunathan, V.K.; P. Garland, S. Automated AFM force curve analysis for determining elastic modulus of biomaterials and biological samples. J. Mech. Behav. Biomed. Mater. 2014, 37, 209–218. [Google Scholar] [CrossRef] [Green Version]
- Guo, Q.; Xia, Y.; Sandig, M. Characterization of cell elasticity correlated with cell morphology by atomic force microscope. J. Biomech. 2012, 45, 304–309. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Chen, Y.; Chen, Y. An AFM-based pit-measuring method for indirect measurements of cell-surface membrane vesicles. Biochem. Biophys. Res. Commun. 2014, 446, 375–379. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Tian, L.; Wang, Y. Determination of viscohyperelastic properties of tubule epithelial cells by an approach combined with AFM nanoindentation and finite element analysis. Micron 2020, 129, 102779. [Google Scholar] [CrossRef] [PubMed]
- Codan, B.; Martinelli, V.; Mestroni, L.; Sbaizero, O. Atomic force microscopy of 3T3 and SW-13 cell lines: An investigation of cell elasticity changes due to fixation. Mater. Sci. Eng. C 2013, 33, 3303–3308. [Google Scholar] [CrossRef]
- Codan, B.; G del Favero, V.; Martinelli, C.S.L. Exploring the elasticity and adhesion behavior of cardiac fibroblasts by atomic force microscopy indentation. Mater. Sci. Eng. C 2014, 40, 427–434. [Google Scholar] [CrossRef] [Green Version]
- Luo, Q.; Kuang, D.; Zhang, B. Cell stiffness determined by atomic force microscopy and its correlation with cell motility. Biochim. Biophys. Acta Gen. Subj. 2016, 1860, 1953–1960. [Google Scholar] [CrossRef]
- Hayashi, K.; Iwata, M. Stiffness of cancer cells measured with an AFM indentation method. J. Mech. Behav. Biomed. Mater. 2015, 49, 105–111. [Google Scholar] [CrossRef]
- Babahosseini, H.; Carmichael, B.; Strobl, J.S.; Mahmoodi, N.S. Sub-cellular force microscopy in single normal and cancer cells. Biochem. Biophys. Res. Commun. 2015, 463, 587–592. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Wang, Z.; Wang, X. AFM-based study of fullerenol (C60(OH)24)-induced changes of elasticity in living SMCC-7721 cells. J. Mech. Behav. Biomed. Mater. 2015, 45, 65–74. [Google Scholar] [CrossRef]
- Nguyen, N.; Shao, Y.; Wineman, A. Atomic force microscopy indentation and inverse analysis for non-linear viscoelastic identification of breast cancer cells. Math. Biosci. 2016, 277, 77–88. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.; Yamagishi, A.; Imaizumi, M. Quantitative measurements of intercellular adhesion between a macrophage and cancer cells using a cup-attached AFM chip. Colloids Surf. B Biointerfaces 2017, 155, 366–372. [Google Scholar] [CrossRef]
- Kulkarni, T.; Tam, A.; Mukhopadhyay, D. AFM study: Cell cycle and probe geometry influences nanomechanical characterization of Panc1 cells. Biochim. Biophys. Acta Gen. Subj. 2019, 1863, 802–812. [Google Scholar] [CrossRef] [PubMed]
- Svetlizky, D.; Levi, O.; Benhar, I.; Eliaz, N. Mechanical properties of Bio-Ferrography isolated cancerous cells studied by atomic force microscopy. J. Mech. Behav. Biomed. Mater. 2019, 91, 345–354. [Google Scholar] [CrossRef]
- Nikolaev, N.I.; Müller, T.; Williams, D.J. Changes in the stiffness of human mesenchymal stem cells with the progress of cell death as measured by atomic force microscopy. J. Biomech. 2014, 47, 625–630. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dulinska-Molak, I.; Chlanda, A.; Li, J. The influence of carbon-encapsulated iron nanoparticles on elastic modulus of living human mesenchymal stem cells examined by atomic force microscopy. Micron 2018, 108, 41–48. [Google Scholar] [CrossRef]
- Du, M.; Kavanagh, D.; Kalia, N.; Zhang, Z. Characterising the mechanical properties of haematopoietic and mesenchymal stem cells using micromanipulation and atomic force microscopy. Med. Eng. Phys. 2019, 73, 18–29. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Jin, Z.; Wang, X. AFM detection of biophysical characteristics of specific regulatory T Cells after allogeneic hematopoietic stem cell transplantation. BMC Biophys. 2014, 7, 1–5. [Google Scholar] [CrossRef] [Green Version]
- Yumiyama, S.; Kato, S.; Konishi, Y. Direct measurement of interaction forces between a yeast cell and a microbubble using atomic force microscopy. Colloids Surf. A Physicochem Eng. Asp. 2019, 583, 123963. [Google Scholar] [CrossRef]
- Shi, Y.; Cai, M.; Zhou, L.; Wang, H. Measurement of mechanical properties of naked cell membranes using atomic force microscope puncture test. Talanta 2020, 210, 120637. [Google Scholar] [CrossRef]
- Brückner Bastian, R.; Janshoff, A. Elastic properties of epithelial cells probed by atomic force microscopy. Biochim. Biophys. Acta 2015, 1853, 3075–3082. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zemła, J.; Danilkiewicz, J.; Orzechowska, B. Atomic force microscopy as a tool for assessing the cellular elasticity and adhesiveness to identify cancer cells and tissues. Semin. Cell Dev. Biol. 2017, 115–124. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Mollaeian, K.; Ren, J. Finite element modeling of living cells for AFM indentation-based biomechanical characterization. Micron 2019, 116, 108–115. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.S.; Lee, G.Y.H.; Ong, C.N. AFM indentation study of breast cancer cells. Biochem. Biophys. Res. Commun. 2008, 374, 609–613. [Google Scholar] [CrossRef]
- Sokolov, I.; Dokukin, E.M.; Guz, N.V. Method for quantitative measurements of the elastic modulus of biological cells in AFM indentation experiments. Methods 2013, 60, 202–213. [Google Scholar] [CrossRef]
- Song, J.; Meng, X.; Zhang, H.; Zhao, K.; Hu, Y.; Xie, H. Probing Multi-Dimens. Mech. Phenotyping Intracell. Struct. By Viscoelastic Spectroscopy. Acs Appl. Mater. Interfaces 2020, 12, 1913–1923. [Google Scholar] [CrossRef]
- Ryu, S.; Hashizume, Y.; Mishima, M. Measurement of cell adhesion force by vertical forcible detachment using an arrowhead nanoneedle and atomic force microscopy. Biochem. Biophys. Res. Commun. 2014, 451, 107–111. [Google Scholar] [CrossRef]
- Alcaraz, J.; Otero, J.; Jorba, I.; Navajas, D. Bidirectional mechanobiology between cells and their local extracellular matrix probed by atomic force microscopy. Semin. Cell Dev. Biol. 2018, 73, 71–81. [Google Scholar] [CrossRef]
- Li, Q.; Becker, T.; Sand, W. Quantification of cell-substratum interactions by atomic force microscopy. Colloids Surf. B Biointerfaces 2017, 159, 639–643. [Google Scholar] [CrossRef]
- Stewart, M.P.; Hodel, A.W.; Spielhofer, A.; Cattin, J.C.; Müller, D.J.; Helenius, J. Wedged AFM-cantilevers for parallel plate cell mechanics. Methods 2013, 60, 186–194. [Google Scholar] [CrossRef]
- Nijenhuis, N.; Zhao, X.; Carisey, A. Combining AFM and acoustic probes to reveal changes in the elastic stiffness tensor of living cells. Biophys. J. 2014, 107, 1502–1512. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Andolfi, L.; Greco, S.L.M.; Tierno, D. Planar AFM macro-probes to study the biomechanical properties of large cells and 3D cell spheroids. Acta Biomater. 2019, 94, 505–513. [Google Scholar] [CrossRef]
- Rother, J.; Noding, H.; Mey, I. Atomic force microscopy-based microrheology reveals significant differences in the viscoelastic response between malign and benign cell lines. Open Biol. 2014, 4, 2046–2441. [Google Scholar] [CrossRef]
- Choi, H.; Choi, E.H.; Kim, K.S. Changes in the biomechanical properties of a single cell induced by nonthermal atmospheric pressure micro-dielectric barrier discharge plasma. Microsc. Res. Tech. 2017, 80, 1078–1084. [Google Scholar] [CrossRef] [PubMed]
- Friedrichs, J.; Legate, K.R.; Schubert, R. A practical guide to quantify cell adhesion using single-cell force spectroscopy. Methods 2013, 60, 169–178. [Google Scholar] [CrossRef] [PubMed]
- Hu, S. Mechanical anisotropy of adherent cells probed by a three-dimensional magnetic twisting device. Am. J. Physiol. Cell Physiol. 2004, 287, C1184. [Google Scholar] [CrossRef] [Green Version]
- Sniadecki, N.J.; Chen, C.S. Methods in Cell Biology: Cell Mechanics; Elsevier Inc.: Amsterdam, The Netherlands, 2007. [Google Scholar]
- Isabey, D.; Pelle, G.; Dias, S.A.; Bottier, M.; Nguyen, M.N.; Filoche, M.; Louis, B. Multiscale evaluation of cellular adhesion alteration and cytoskeleton remodeling by magnetic bead twisting. Biomech. Model. Mechanobiol. 2016, 15, 947–963. [Google Scholar] [CrossRef]
- Chen, L.; Kraus, M.V. Implementation and application of a novel 2D magnetic twisting cytometry based on multi-pole electromagnet. Rev. Sci. Instrum. 2016, 87, 0643011-8. [Google Scholar] [CrossRef] [Green Version]
- Beussman, K.M.; Rodriguez, M.L.; Leonard, A. Micropost arrays for measuring stem cell-derived cardiomyocyte contractility. Methods 2016, 2016, 43–50. [Google Scholar] [CrossRef] [Green Version]
- Majid, W.; Lim, C.T. Microfluidic Platforms for Human Disease Cell Mechanics Studies; Springer: Vienna, Italy, 2013; pp. 107–119. [Google Scholar]
- Makarova, N.; Kalaparthi, V.; Wang, A. Difference in biophysical properties of cancer-initiating cells in melanoma mutated zebrafish. J. Mech. Behav. Biomed. Mater. 2020, 107, 103746. [Google Scholar] [CrossRef]
- Ani, C.J.; Obayemi, J.D.; Uzonwanne, V.O. A shear assay study of single normal/breast cancer cell deformation and detachment from Poly-Di-Methyl-Siloxane (PDMS) surfaces. J. Mech. Behav. Biomed. Mater. 2018, 91, 76–90. [Google Scholar] [CrossRef] [PubMed]
- Lee, D.; Cha, C. Cell subtype-dependent formation of breast tumor spheroids and their variable responses to chemotherapeutics within microfluidics-generated 3D microgels with tunable mechanics. Mater. Sci. Eng. C 2020, 112, 110932. [Google Scholar] [CrossRef] [PubMed]
- Khan, Z.S. Probing the mechanical properties of brain cancer cells using a microfluidic cell squeezer device. Biomicrofluidics 2013, 7, 11086. [Google Scholar] [CrossRef] [Green Version]
- Pablo JG, D.; Armistead, F.J.; Peyman, S.A. Biochemical fingerprint of colorectal cancer cell lines using label-free live single-cell Raman spectroscopy. J. Raman Spectrosc. 2018, 49, 1323–1332. [Google Scholar] [CrossRef] [Green Version]
- Matsuura, K.; Hayashi, N.; Kuroda, Y. Improved development of mouse and human embryos using a tilting embryo culture system. Reprod. Biomed. Online 2010, 20, 358–364. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koike, T.; Matsuura, K. In-vitro culture with a tilting device in chemically defined media during meiotic maturation and early development improves the quality of blastocysts derived from in-vitro matured and fertilized porcine oocytes. J. Reprod. Dev. 2010, 15, 552–557. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chang, K.W.; Chang, P.Y.; Huang, H.Y. Womb-on-a-chip biomimetic system for improved embryo culture and development. Sens. Actuators B Chem. 2016, 226, 218–226. [Google Scholar] [CrossRef]
- Huang, H.Y.; Shen, H.H.; Tien, C.H. Digital microfluidic dynamic culture of mammalian embryos on an electrowetting on dielectric (EWOD) chip. PLoS ONE 2015, 10, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Ferreira, J.P.S.; Kuang, M. Altered mechanics of vaginal smooth muscle cells due to the lysyl oxidaselike1 knockout. Acta Biomater. 2020, 110, 175–187. [Google Scholar] [CrossRef]
- Venkataraman, L.; Lenis, A.T.; Couri, B.M. Induced regenerative elastic matrix repair in LOXL1 knockout mouse cell cultures: Towards Potential therapy for Pelvic Organ Prolapse. J. Tissue Sci. Eng. 2012, 3, 120. [Google Scholar]
- Allahverdian, S.; Chehroudi, A.C. Contribution of intimal smooth muscle cells to cholesterol accumulation and macrophage-like cells in human atherosclerosis. Circulation 2015, 31, e25. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sanyour, J.; Li, N. Statin-mediated cholesterol depletion exerts coordinated effects on the alterations in rat vascular smooth muscle cell biomechanics and migration. J Physiol. 2020, 598, 1505–1522. [Google Scholar] [CrossRef] [PubMed]
- Guo, Y.; Wang, Y.; Liu, Y. Effect of the same mechanical loading on osteogenesis and osteoclastogenesis in vitro. Chin. J. Traumatol. 2015, 18, 150–156. [Google Scholar] [CrossRef] [PubMed]
- Goggin, P.M.; Zygalakis, K.C.; Oreffo, R.O.C. High-resolution 3D imaging of osteocytes and computational modelling in mechanobiology: Insights on bone development, ageing, health and disease. Eur. Cells Mater. 2016, 31, 264–295. [Google Scholar] [CrossRef]
- Yang, X.; Sun, L.W. Finite element analysis of osteocytes mechanosensitivity under simulated microgravity. Microgravity Sci. Technol. 2018, 30, 469–481. [Google Scholar] [CrossRef]
- Gu, Y.; Zhang, W.; Sun, Q. Microbead-guided reconstruction of the 3D osteocyte network during microfluidic perfusion culture. J. Mater. Chem. B 2015, 3, 3625–3633. [Google Scholar] [CrossRef] [Green Version]
- Bartolozzi, A.; Viti, F.; De Stefano, S. Development of label-free biophysical markers in osteogenic maturation. J. Mech. Behav. Biomed. Mater. 2020, 103, 103581. [Google Scholar] [CrossRef]
- Hsieh, W.T.; Liu, Y.S. Matrix dimensionality and stiffness cooperatively regulate osteogenesis of mesenchymal stromal cells. Acta Biomater. 2016, 32, 210–222. [Google Scholar] [CrossRef]
- Yen, M.H.; Cen, Y.H.; Liu, Y.S. Alteration of Young's modulus in mesenchymal stromal cells during osteogenesis measured by atomic force microscopy. Biochem. Biophys. Res. Commun. 2020, 526, 827–832. [Google Scholar] [CrossRef]
- Olivier-De La Cruz, J.; Nardone, G.; Vrbsky, J. Substrate mechanics controls adipogenesis through YAP phosphorylation by dictating cell spreading. Biomaterials 2019, 205, 64–80. [Google Scholar] [CrossRef]
- Tan, Y.; Sun, D.; Huang, W.; Cheng, S.H. Mechanical modeling of biological cells in microinjection. IEEE Trans. Nano Biosci. 2008, 7, 257–266. [Google Scholar] [CrossRef] [PubMed]
- Tan Sun, D.; Huang, W. Mechanical modeling characterization of biological cells using microrobotics cell injection test bed. Processings of the 2009 IEEE/RSJ International Conference on Intelligent Robots and Systems, St. Louis, MO, USA, 10–15 October 2009. [Google Scholar]
- Shen, T.; Shirinzadeh, B.; Zhong, Y. A hyperelastic model for mechanical responses of adherent cells in microinjection. In Proceedings of the IEEE International Conference on Manipulation, Shanghai, China, 8 February 2018. [Google Scholar]
- Liu, F.; Wu, D.; Chen, K. Mechanical behavior of cells in microinjection: A minimum potential energy study. J. Mech. Behav. Biomed. Mater. 2013, 24, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Yan, K.C.; Nair KSun, W. Three dimensional multi-scale modelling and analysis of cell damage in cell-encapsulated alginate constructs. J. Biomech. 2010, 43, 1031–1038. [Google Scholar]
- Kim, C.Y.; Kang, J.H.; Park, S.J. Microfluidic biomechanical device for compressive cell stimulation and lysis. Sens. Actuators B Chem. 2007, 128, 108–116. [Google Scholar] [CrossRef]
- Gayán Elisa Govers, S.; Aertsen, A. Impact of high hydrostatic pressure on bacterial proteostasis. Biophys. Chem. 2017, 231, 3–9. [Google Scholar] [CrossRef] [PubMed]
- Syed, Q.A.; Buffa, M.; Guamis, B. Factors Affecting bacterial inactivation during high hydrostatic pressure processing of foods: A Review. Crit. Rev. Food Sci. Nutr. 2016, 56, 474–483. [Google Scholar] [CrossRef]
- Cheng, B.; Wan Huang, G. Nanoscale integrin cluster dynamics controls cellular mechanosensing via FAKY397 phosphorylation. Sci. Adv. 2020, 6, eaax1909. [Google Scholar] [CrossRef] [Green Version]
- Kirby, T.J.; Lammerding, J. Emerging views of the nucleus as a cellular mechanosensor. Nat. Cell Biol. 2018, 20, 373–381. [Google Scholar] [CrossRef]








| Type | Description | Composition |
|---|---|---|
| membrane | Elastic semipermeable membrane composed of phospholipid. | Phospholipid bilayer, glycoprotein, glycolipid. |
| cytoplasm | A general term for all translucent, colloidal and granular materials outside the nuclear zone. | Golgi apparatus, mitochondria, endoplasmic reticulum, ribosome. |
| nucleus | The main site of storage, replication, and transcription of genetic information in cells. | Nuclear membrane, chromatin, nucleolus, nuclear matrix. |
| cytoskeleton | The network structure of protein fibers. | Microfilaments, microtubules, intermediate fibers. |
| Name | Type | Characteristic | Application |
|---|---|---|---|
| Continuous cortical membrane model | Droplet model | A fluid-like sphere surrounded by continuous cortex | Suspension cells |
| Solid model | Isotropic solid substances | Suspension and adherent cells | |
| Damping model | A frequency-domain model under dynamic forces | Adherent cells | |
| Two-phase model | The solid–liquid duality | Articular chondrocyte | |
| Discrete network skeleton model | Tension integration model | A space balance system | Adherent cells |
| Porous solid mode | A regular network model | Adherent cells | |
| Cable net model | An ideal articulated cable network | Red blood cells | |
| Biochemical mechanical model | Combination of biochemical process and mechanics | Adherent cells | |
| Cortical skeleton combined model | Based on finite element | From the point of view of finite element | Suspension and adherent cells |
| Based on molecular dynamic | From the point of view of molecules | Suspension and adherent cells |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Huang, H.; Dai, C.; Shen, H.; Gu, M.; Wang, Y.; Liu, J.; Chen, L.; Sun, L. Recent Advances on the Model, Measurement Technique, and Application of Single Cell Mechanics. Int. J. Mol. Sci. 2020, 21, 6248. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21176248
Huang H, Dai C, Shen H, Gu M, Wang Y, Liu J, Chen L, Sun L. Recent Advances on the Model, Measurement Technique, and Application of Single Cell Mechanics. International Journal of Molecular Sciences. 2020; 21(17):6248. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21176248
Chicago/Turabian StyleHuang, Haibo, Cihai Dai, Hao Shen, Mingwei Gu, Yangjun Wang, Jizhu Liu, Liguo Chen, and Lining Sun. 2020. "Recent Advances on the Model, Measurement Technique, and Application of Single Cell Mechanics" International Journal of Molecular Sciences 21, no. 17: 6248. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21176248