Aldo-Keto Reductase 1C1 (AKR1C1) as the First Mutated Gene in a Family with Nonsyndromic Primary Lipedema

 ,
,  ,
,  , , , , ,
, , , , ,
Abstract
:1. Introduction
2. Results
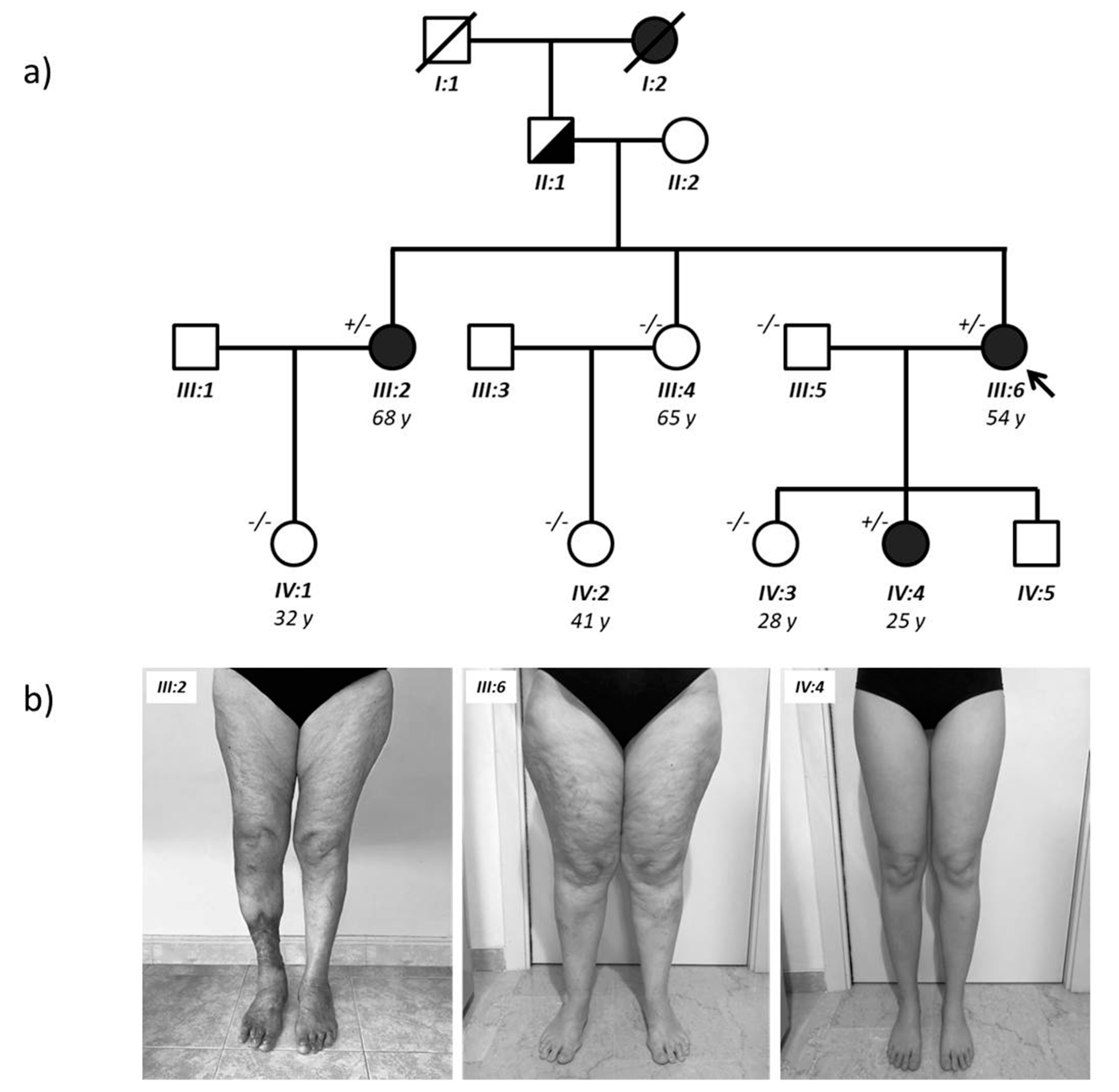
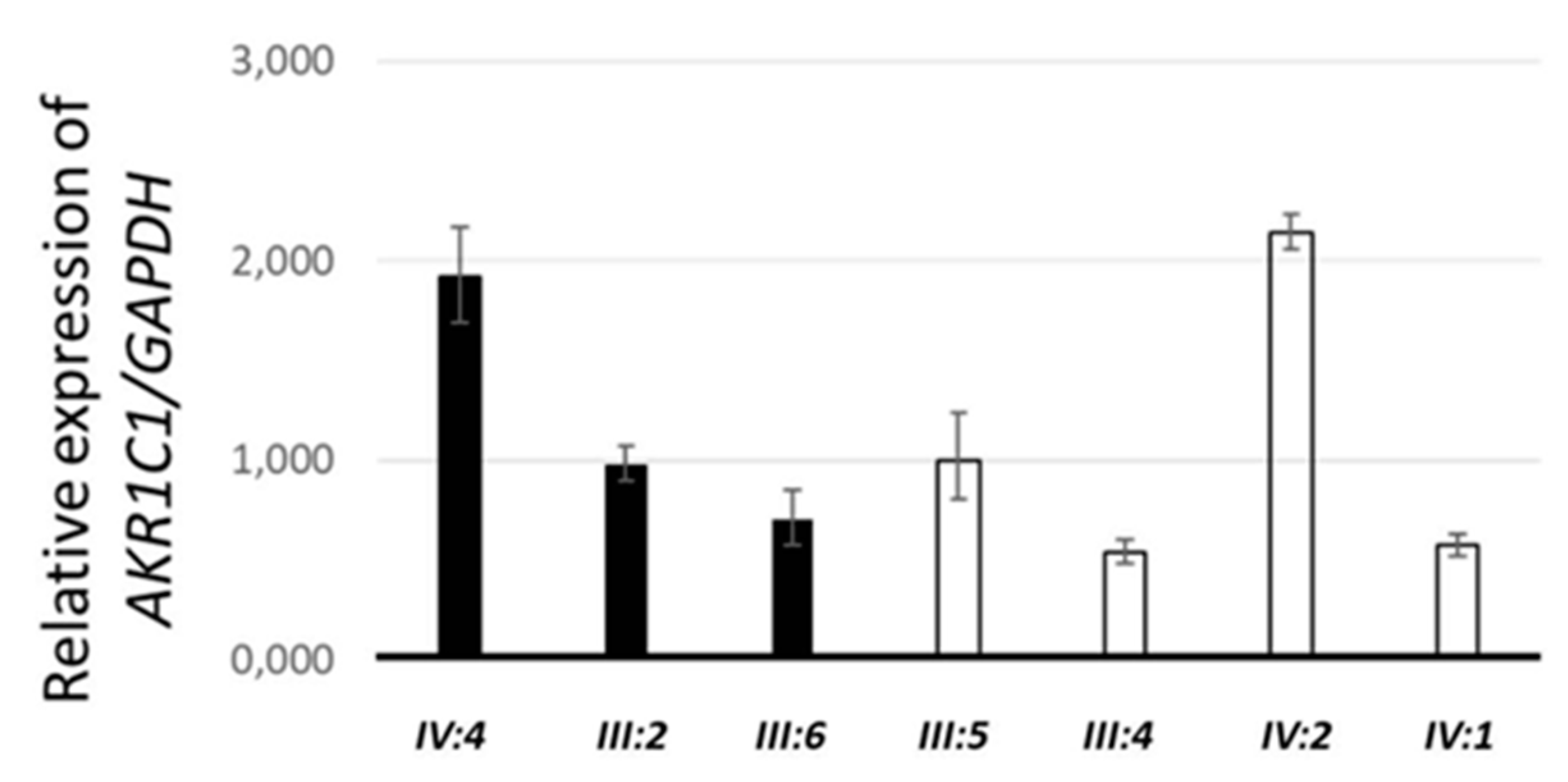
2.1. Identification of a Missense AKR1C1 Variant in Lipedema Patients
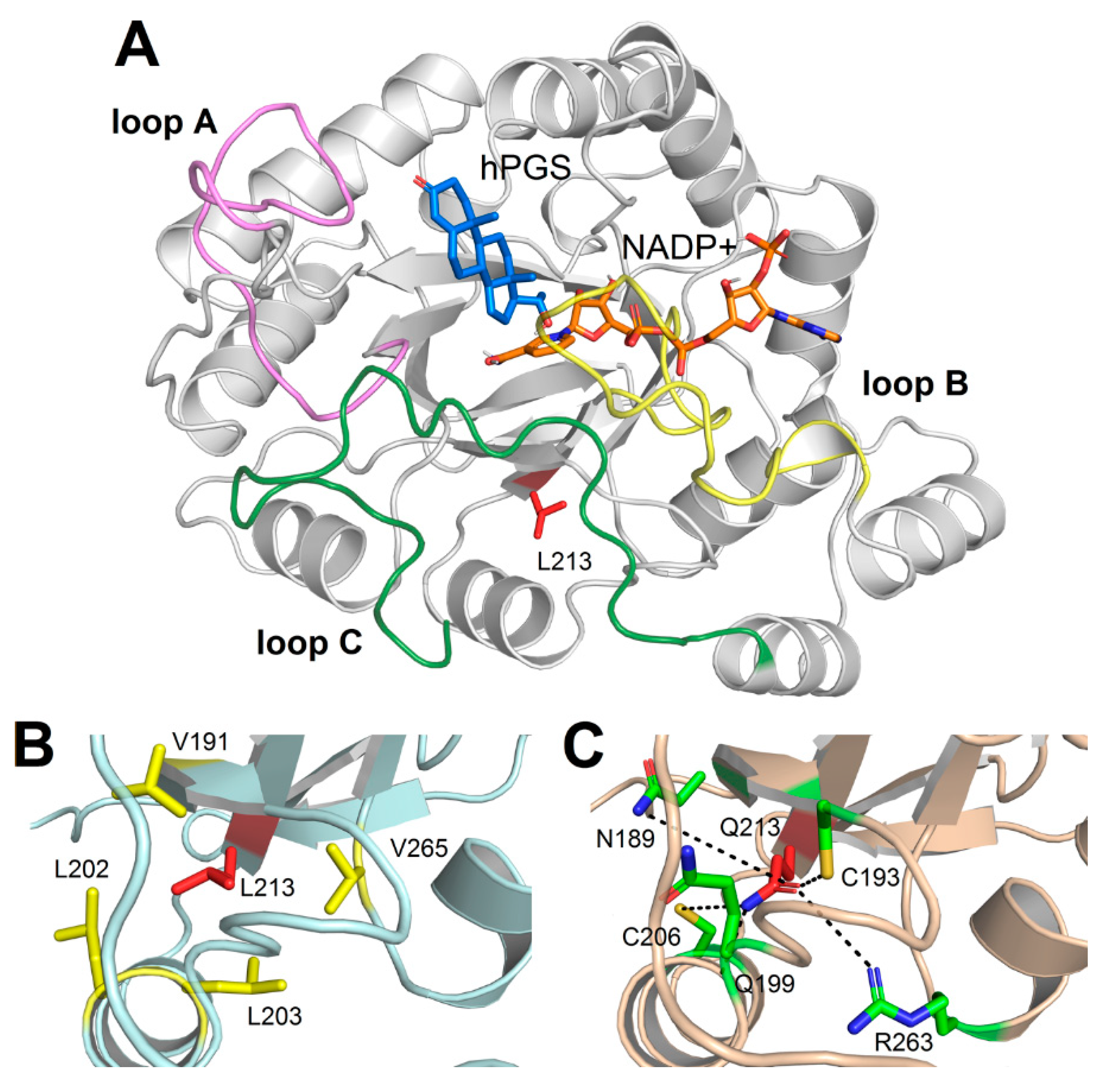
2.2. Structural Analysis and Molecular Dynamics Simulations
2.3. QSAR Models Predict a Partial Loss of Function for The L12Q AK1RC1 Variant
3. Discussion
4. Materials and Methods
4.1. Ethical Compliance
4.2. Subjects
4.3. Whole Exome Sequencing
4.4. Sanger Sequencing Analysis
4.5. Quantitative Real-Time Polymerase Chain Reaction
4.6. Molecular Modeling And Molecular Dynamics (MD) Simulations
4.7. QSAR Models and Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Bauer, A.T.; von Lukowicz, D.; Lossagk, K.; Aitzetmueller, M.; Moog, P.; Cerny, M.; Erne, H.; Schmauss, D.; Duscher, D.; Machens, H.G. New Insights on Lipedema: The Enigmatic Disease of the Peripheral Fat. Plast. Reconstr. Surg. 2019, 144, 1475–1484. [Google Scholar] [CrossRef] [PubMed]
- Forner-Cordero, I.; Szolnoky, G.; Forner-Cordero, A.; Kemeny, L. Lipedema: An overview of its clinical manifestations, diagnosis and treatment of the disproportional fatty deposition syndrome - systematic review. Clin. Obes. 2012, 2, 86–95. [Google Scholar] [CrossRef] [PubMed]
- Buso, G.; Depairon, M.; Tomson, D.; Raffoul, W.; Vettor, R.; Mazzolai, L. Lipedema: A Call to Action! Obes. (Silver Spring) 2019, 27, 1567–1576. [Google Scholar] [CrossRef] [PubMed]
- Herbst, K.L. Rare adipose disorders (RADs) masquerading as obesity. Acta. Pharmacol. Sin. 2012, 33, 155–172. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paolacci, S.; Precone, V.; Acquaviva, F.; Chiurazzi, P.; Fulcheri, E.; Pinelli, M.; Buffelli, F.; Michelini, S.; Herbst, K.L.; Unfer, V.; et al. Genetics of lipedema: New perspectives on genetic research and molecular diagnoses. Eur. Rev. Med. Pharmacol. Sci. 2019, 23, 5581–5594. [Google Scholar] [CrossRef] [PubMed]
- Child, A.H.; Gordon, K.D.; Sharpe, P.; Brice, G.; Ostergaard, P.; Jeffery, S.; Mortimer, P.S. Lipedema: An inherited condition. Am. J. Med. Genet. A 2010, 152A, 970–976. [Google Scholar] [CrossRef]
- Bano, G.; Mansour, S.; Brice, G.; Ostergaard, P.; Mortimer, P.S.; Jeffery, S.; Nussey, S. Pit-1 mutation and lipoedema in a family. Exp. Clin. Endocrinol. Diabetes 2010, 118, 377–380. [Google Scholar] [CrossRef]
- Van Pelt, R.E.; Gozansky, W.S.; Hickner, R.C.; Schwartz, R.S.; Kohrt, W.M. Acute modulation of adipose tissue lipolysis by intravenous estrogens. Obes. (Silver Spring) 2006, 14, 2163–2172. [Google Scholar] [CrossRef] [Green Version]
- Gavin, K.M.; Cooper, E.E.; Hickner, R.C. Estrogen receptor protein content is different in abdominal than gluteal subcutaneous adipose tissue of overweight-to-obese premenopausal women. Metabolism 2013, 62, 1180–1188. [Google Scholar] [CrossRef]
- O’Brien, S.N.; Welter, B.H.; Mantzke, K.A.; Price, T.M. Identification of progesterone receptor in human subcutaneous adipose tissue. J. Clin. Endocrinol. Metab. 1998, 83, 509–513. [Google Scholar] [CrossRef] [Green Version]
- Xu, Y.; Lopez, M. Central regulation of energy metabolism by estrogens. Mol. Metab. 2018, 15, 104–115. [Google Scholar] [CrossRef] [PubMed]
- Gray, J.M.; Wade, G.N. Food intake, body weight, and adiposity in female rats: Actions and interactions of progestins and antiestrogens. Am. J. Physiol. 1981, 240, E474–E481. [Google Scholar] [CrossRef] [PubMed]
- Wade, G.N. Some effects of ovarian hormones on food intake and body weight in female rats. J. Comp. Physiol. Psychol. 1975, 88, 183–193. [Google Scholar] [CrossRef] [PubMed]
- Kessler, R.C.; Chiu, W.T.; Demler, O.; Merikangas, K.R.; Walters, E.E. Prevalence, severity, and comorbidity of 12-month DSM-IV disorders in the National Comorbidity Survey Replication. Arch. Gen. Psychiatry 2005, 62, 617–627. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Da Pozzo, E.; Costa, B.; Martini, C. Translocator protein (TSPO) and neurosteroids: Implications in psychiatric disorders. Curr. Mol. Med. 2012, 12, 426–442. [Google Scholar] [CrossRef]
- Quast, C.; Reif, A.; Bruckl, T.; Pfister, H.; Weber, H.; Mattheisen, M.; Cichon, S.; Lang, T.; Hamm, A.; Fehm, L.; et al. Gender-specific association of variants in the AKR1C1 gene with dimensional anxiety in patients with panic disorder: Additional evidence for the importance of neurosteroids in anxiety? Depress. Anxiety 2014, 31, 843–850. [Google Scholar] [CrossRef]
- Joksimovic, S.L.; Covey, D.F.; Jevtovic-Todorovic, V.; Todorovic, S.M. Neurosteroids in Pain Management: A New Perspective on an Old Player. Front. Pharm. 2018, 9, 1127. [Google Scholar] [CrossRef] [Green Version]
- Coronel, M.F.; Labombarda, F.; Roig, P.; Villar, M.J.; De Nicola, A.F.; Gonzalez, S.L. Progesterone prevents nerve injury-induced allodynia and spinal NMDA receptor upregulation in rats. Pain Med. 2011, 12, 1249–1261. [Google Scholar] [CrossRef] [Green Version]
- Kwon, J.M.; Goate, A.M. The candidate gene approach. Alcohol Res. Health 2000, 24, 164–168. [Google Scholar]
- Kim, H.J.; Park, J.H.; Lee, S.; Son, H.Y.; Hwang, J.; Chae, J.; Yun, J.M.; Kwon, H.; Kim, J.I.; Cho, B. A Common Variant of NGEF Is Associated with Abdominal Visceral Fat in Korean Men. PLoS ONE 2015, 10, e0137564. [Google Scholar] [CrossRef] [Green Version]
- Norris, J.M.; Langefeld, C.D.; Talbert, M.E.; Wing, M.R.; Haritunians, T.; Fingerlin, T.E.; Hanley, A.J.; Ziegler, J.T.; Taylor, K.D.; Haffner, S.M.; et al. Genome-wide association study and follow-up analysis of adiposity traits in Hispanic Americans: The IRAS Family Study. Obes. (Silver Spring) 2009, 17, 1932–1941. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.; Kent, J.W., Jr.; Olivier, M.; Ali, O.; Cerjak, D.; Broeckel, U.; Abdou, R.M.; Dyer, T.D.; Comuzzie, A.; Curran, J.E.; et al. A comprehensive analysis of adiponectin QTLs using SNP association, SNP cis-effects on peripheral blood gene expression and gene expression correlation identified novel metabolic syndrome (MetS) genes with potential role in carcinogenesis and systemic inflammation. BMC Med. Genom. 2013, 6, 14. [Google Scholar] [CrossRef] [Green Version]
- Park, H.W.; Dahlin, A.; Tse, S.; Duan, Q.L.; Schuemann, B.; Martinez, F.D.; Peters, S.P.; Szefler, S.J.; Lima, J.J.; Kubo, M.; et al. Genetic predictors associated with improvement of asthma symptoms in response to inhaled corticosteroids. J. Allergy Clin. Immunol. 2014, 133, 664–669.e5. [Google Scholar] [CrossRef] [Green Version]
- Rizner, T.L.; Penning, T.M. Role of aldo-keto reductase family 1 (AKR1) enzymes in human steroid metabolism. Steroids 2014, 79, 49–63. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Couture, J.F.; Legrand, P.; Cantin, L.; Luu-The, V.; Labrie, F.; Breton, R. Human 20alpha-hydroxysteroid dehydrogenase: Crystallographic and site-directed mutagenesis studies lead to the identification of an alternative binding site for C21-steroids. J. Mol. Biol. 2003, 331, 593–604. [Google Scholar] [CrossRef]
- Dell’orco, D.; De Benedetti, P.G. Quantitative structure-activity relationship analysis of canonical inhibitors of serine proteases. J. Comput. Aided. Mol. Des. 2008, 22, 469–478. [Google Scholar] [CrossRef]
- Penning, T.M. Molecular endocrinology of hydroxysteroid dehydrogenases. Endocr. Rev. 1997, 18, 281–305. [Google Scholar] [CrossRef] [Green Version]
- Blouin, K.; Richard, C.; Brochu, G.; Hould, F.S.; Lebel, S.; Marceau, S.; Biron, S.; Luu-The, V.; Tchernof, A. Androgen inactivation and steroid-converting enzyme expression in abdominal adipose tissue in men. J. Endocrinol. 2006, 191, 637–649. [Google Scholar] [CrossRef] [Green Version]
- Brozic, P.; Cesar, J.; Kovac, A.; Davies, M.; Johnson, A.P.; Fishwick, C.W.; Lanisnik Rizner, T.; Gobec, S. Derivatives of pyrimidine, phthalimide and anthranilic acid as inhibitors of human hydroxysteroid dehydrogenase AKR1C1. Chem. Biol. Interact. 2009, 178, 158–164. [Google Scholar] [CrossRef]
- Zhang, Y.; Nadeau, M.; Faucher, F.; Lescelleur, O.; Biron, S.; Daris, M.; Rheaume, C.; Luu-The, V.; Tchernof, A. Progesterone metabolism in adipose cells. Mol. Cell Endocrinol. 2009, 298, 76–83. [Google Scholar] [CrossRef]
- Lacasa, D.; Le Liepvre, X.; Ferre, P.; Dugail, I. Progesterone stimulates adipocyte determination and differentiation 1/sterol regulatory element-binding protein 1c gene expression. potential mechanism for the lipogenic effect of progesterone in adipose tissue. J. Biol. Chem. 2001, 276, 11512–11516. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, J.B.; Spiegelman, B.M. ADD1/SREBP1 promotes adipocyte differentiation and gene expression linked to fatty acid metabolism. Genes Dev. 1996, 10, 1096–1107. [Google Scholar] [CrossRef] [Green Version]
- Zhao, X.; Chen, S.; Tan, Z.; Wang, Y.; Zhang, F.; Yang, T.; Liu, Y.; Ao, H.; Xing, K.; Wang, C. Transcriptome Analysis of Landrace Pig Subcutaneous Preadipocytes during Adipogenic Differentiation. Genes (Basel) 2019, 10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dozier, B.L.; Watanabe, K.; Duffy, D.M. Two pathways for prostaglandin F2 alpha synthesis by the primate periovulatory follicle. Reprod. 2008, 136, 53–63. [Google Scholar] [CrossRef] [Green Version]
- Taketani, Y.; Yamagishi, R.; Fujishiro, T.; Igarashi, M.; Sakata, R.; Aihara, M. Activation of the prostanoid FP receptor inhibits adipogenesis leading to deepening of the upper eyelid sulcus in prostaglandin-associated periorbitopathy. Invest. Ophthalmol. Vis. Sci. 2014, 55, 1269–1276. [Google Scholar] [CrossRef] [Green Version]
- Lepak, N.M.; Serrero, G. Prostaglandin F2 alpha stimulates transforming growth factor-alpha expression in adipocyte precursors. Endocrinol. 1995, 136, 3222–3229. [Google Scholar] [CrossRef] [PubMed]
- Buck, D.W., II; Herbst, K.L. Lipedema: A Relatively Common Disease with Extremely Common Misconceptions. Plast. Reconstr. Surg. Glob. Open 2016, 4, e1043. [Google Scholar] [CrossRef]
- Rudomin, P.; Schmidt, R.F. Presynaptic inhibition in the vertebrate spinal cord revisited. Exp. Brain Res. 1999, 129, 1–37. [Google Scholar] [CrossRef] [PubMed]
- Marceddu, G.; Dallavilla, T.; Guerri, G.; Manara, E.; Chiurazzi, P.; Bertelli, M. PipeMAGI: An integrated and validated workflow for analysis of NGS data for clinical diagnostics. Eur. Rev. Med. Pharmacol. Sci. 2019, 23, 6753–6765. [Google Scholar] [CrossRef]
- Richards, S.; Aziz, N.; Bale, S.; Bick, D.; Das, S.; Gastier-Foster, J.; Grody, W.W.; Hegde, M.; Lyon, E.; Spector, E.; et al. Standards and guidelines for the interpretation of sequence variants: A joint consensus recommendation of the American College of Medical Genetics and Genomics and the Association for Molecular Pathology. Genet. Med. 2015, 17, 405–424. [Google Scholar] [CrossRef]
- Ji, Q.; Chang, L.; VanDenBerg, D.; Stanczyk, F.Z.; Stolz, A. Selective reduction of AKR1C2 in prostate cancer and its role in DHT metabolism. Prostate 2003, 54, 275–289. [Google Scholar] [CrossRef] [PubMed]
- Manara, E.; Baron, E.; Tregnago, C.; Aveic, S.; Bisio, V.; Bresolin, S.; Masetti, R.; Locatelli, F.; Basso, G.; Pigazzi, M. MLL-AF6 fusion oncogene sequesters AF6 into the nucleus to trigger RAS activation in myeloid leukemia. Blood 2014, 124, 263–272. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abraham, M.J.; Murtola, T.; Schulz, R.; Páll, S.; Smith, J.C.; Hess, B.; Lindahl, E. GROMACS: High performance molecular simulations through multi-level parallelism from laptops to supercomputers. SoftwareX 2015, 1, 19–25. [Google Scholar] [CrossRef] [Green Version]
- Huang, J.; Rauscher, S.; Nawrocki, G.; Ran, T.; Feig, M.; de Groot, B.L.; Grubmuller, H.; MacKerell, A.D., Jr. CHARMM36m: An improved force field for folded and intrinsically disordered proteins. Nat. Methods 2017, 14, 71–73. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jo, S.; Kim, T.; Vidyashankara, G.I.; Wonpil, I. CHARMM-GUI: A web-based graphical user interface for CHARMM. J. Comput. Chem. 2008, 29, 1859–1865. [Google Scholar] [CrossRef] [PubMed]
- Marino, V.; Sulmann, S.; Koch, K.W.; Dell’Orco, D. Structural effects of Mg(2)(+) on the regulatory states of three neuronal calcium sensors operating in vertebrate phototransduction. Biochim. Biophys. Acta 2015, 1853, 2055–2065. [Google Scholar] [CrossRef] [Green Version]
- Marino, V.; Dell’Orco, D. Allosteric communication pathways routed by Ca(2+)/Mg(2+) exchange in GCAP1 selectively switch target regulation modes. Sci. Rep. 2016, 6, 34277. [Google Scholar] [CrossRef]
- Tiberti, M.; Invernizzi, G.; Lambrughi, M.; Inbar, Y.; Schreiber, G.; Papaleo, E. PyInteraph: A framework for the analysis of interaction networks in structural ensembles of proteins. J. Chem. Inf. Model. 2014, 54, 1537–1551. [Google Scholar] [CrossRef]
- Marino, V.; Dell’Orco, D. Evolutionary-Conserved Allosteric Properties of Three Neuronal Calcium Sensor Proteins. Front. Mol. Neurosci. 2019, 12, 50. [Google Scholar] [CrossRef] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Variant | SAS NADP+ (nm2) | SAS hPGS (nm2) | IEP-hP (kJ/mol) | IEN-hP (kJ/mol) |
|---|---|---|---|---|
| WILD-TYPE | 1.10 ± 0.26 | 0.98 ± 0.43 | –122.91 ± 23.60 | –9.75 ± 7.74 |
| L123Q | 1.55 ± 0.35 | 1.46 ± 0.59 | –105.66 ± 23.88 | –5.22 ± 6.17 |
| Variant | IEP-hP (kJ/mol) | IEP-hP (kJ/mol) | Km (μM) | kcat (min−1) | kcat/Km (min−1 μM−1) |
|---|---|---|---|---|---|
| R304L | −119.28 ± 81.07 | −31.77 ± 8.89 | 283.3 ± 13.2 | 96.1 | 0.3 ↓ |
| E127D | −97.50 ± 63.49 | −14.29 ± 9.22 | 9.4 ± 2.5 | 4.4 | 0.4 ↓ |
| H222I | −111.04 ± 67.84 | −28.61 ± 8.76 | 28.6 ± 5.2 | 18.5 | 0.6 ↓ |
| H222S | −110.99 ± 68.25 | −8.57 ± 5.19 | 25.6 ± 5.7 | 39.6 | 1.5 ↓ |
| L213Q | −114.93 ± 24.89 | −8.90 ± 6.87 | 5.1 | 38.2 | 7.55 ↓ |
| T307V | −126.19 ± 70.32 | −33.38 ± 7.61 | 4.1 ± 0.3 | 41.3 | 10 ↓ |
| WILD-TYPE | −118.72 ± 13.23 | −5.69 ± 7.10 | 4.2 ± 0.8 | 65.1 | 15.2 |
| D309L | −126.98 ± 83.16 | −19.78 ± 10.08 | 4.6 ± 0.6 | 119.5 | 25.9 ↑ |
| Y305F | −120.12 ± 84.75 | −19.72 ± 6.08 | 5.6 ± 1.9 | 156.3 | 27.9 ↑ |
| Clinical Characteristics | III:2 Sister | III:6 Proband | IV:4 Daughter |
|---|---|---|---|
| Age | 68 | 54 | 25 |
| Age of onset | Puberty | Puberty | Puberty |
| Menarche | Regular | Regular | Regular |
| Height (m) | 1.68 | 1.65 | 1.75 |
| Weight (Kg) | 68 | 79 | 56 |
| BMI | 24.09 normal | 29.02 overweight | 18.29 normal |
| Lipedema stage Localization of fat depots | Stage 2, Type 2 thighs and buttocks | Stage 2, Type 2 thighs and buttocks | Stage 1, Type 2 thighs |
| Comorbidity (diabetes, hypertension, dyslipidemia) | Hypertension | None | none |
| Endocrine alteration (e.g., thyroid, insulin-resistance) | Thyroid disfunction | None | none |
| Pain in the morning (VAS scale 1–10) | 0 | 0 | 0 |
| Pain at night (VAS scale 1–10) | 0 | 0 | 0 |
| Anxiety/depression/fatigue | Anxiety | none | none |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Michelini, S.; Chiurazzi, P.; Marino, V.; Dell’Orco, D.; Manara, E.; Baglivo, M.; Fiorentino, A.; Maltese, P.E.; Pinelli, M.; Herbst, K.L.; et al. Aldo-Keto Reductase 1C1 (AKR1C1) as the First Mutated Gene in a Family with Nonsyndromic Primary Lipedema. Int. J. Mol. Sci. 2020, 21, 6264. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21176264
Michelini S, Chiurazzi P, Marino V, Dell’Orco D, Manara E, Baglivo M, Fiorentino A, Maltese PE, Pinelli M, Herbst KL, et al. Aldo-Keto Reductase 1C1 (AKR1C1) as the First Mutated Gene in a Family with Nonsyndromic Primary Lipedema. International Journal of Molecular Sciences. 2020; 21(17):6264. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21176264
Chicago/Turabian StyleMichelini, Sandro, Pietro Chiurazzi, Valerio Marino, Daniele Dell’Orco, Elena Manara, Mirko Baglivo, Alessandro Fiorentino, Paolo Enrico Maltese, Michele Pinelli, Karen Louise Herbst, and et al. 2020. "Aldo-Keto Reductase 1C1 (AKR1C1) as the First Mutated Gene in a Family with Nonsyndromic Primary Lipedema" International Journal of Molecular Sciences 21, no. 17: 6264. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21176264