The Synthetic Cannabinoids THJ-2201 and 5F-PB22 Enhance In Vitro CB1 Receptor-Mediated Neuronal Differentiation at Biologically Relevant Concentrations


 , ,
, ,  and
and 
Abstract
:1. Introduction
2. Results
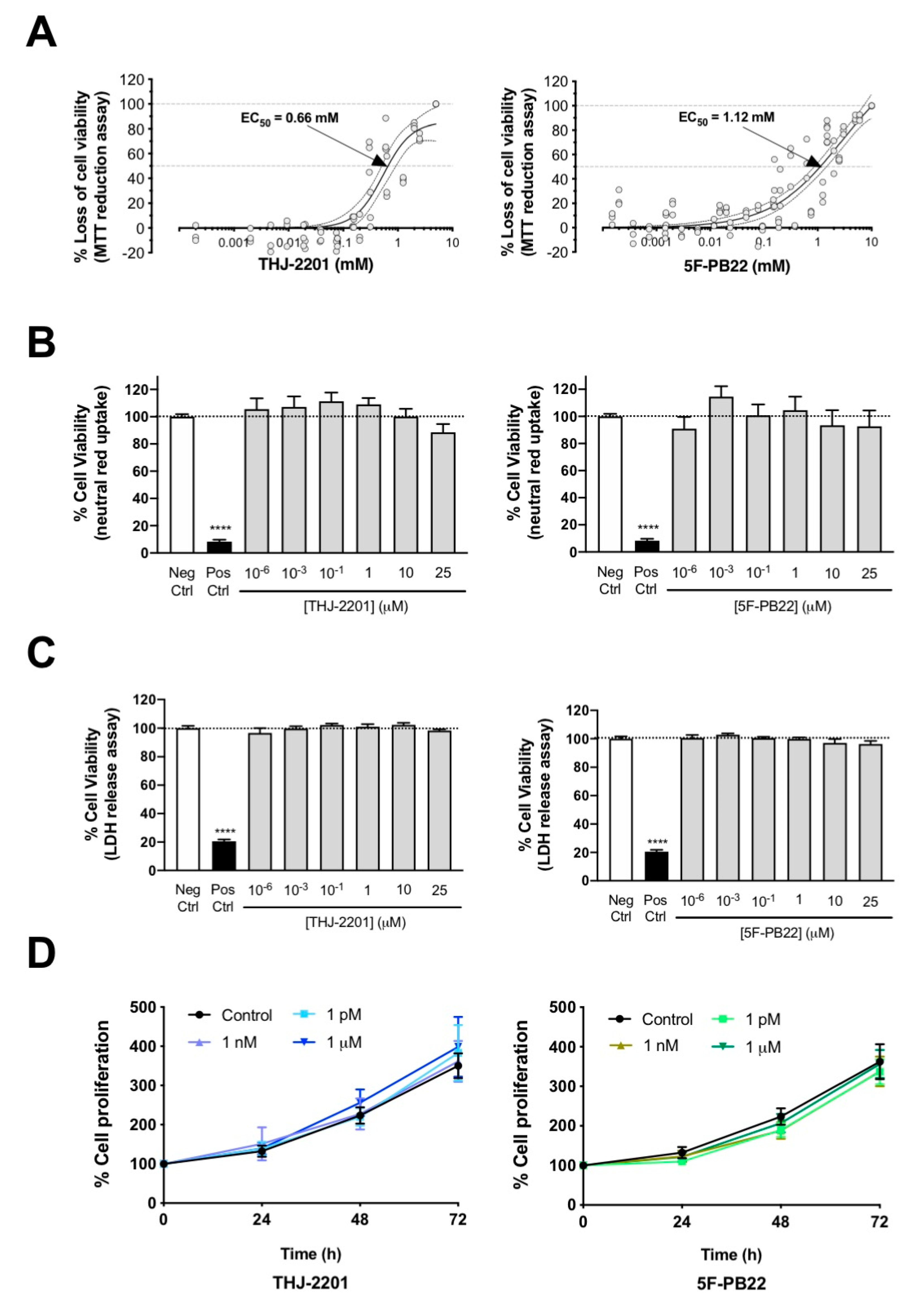
2.1. Cell Viability and Proliferation Were Not Affected by 5F-PB22 or THJ-2201 at Biologically Relevant Concentrations
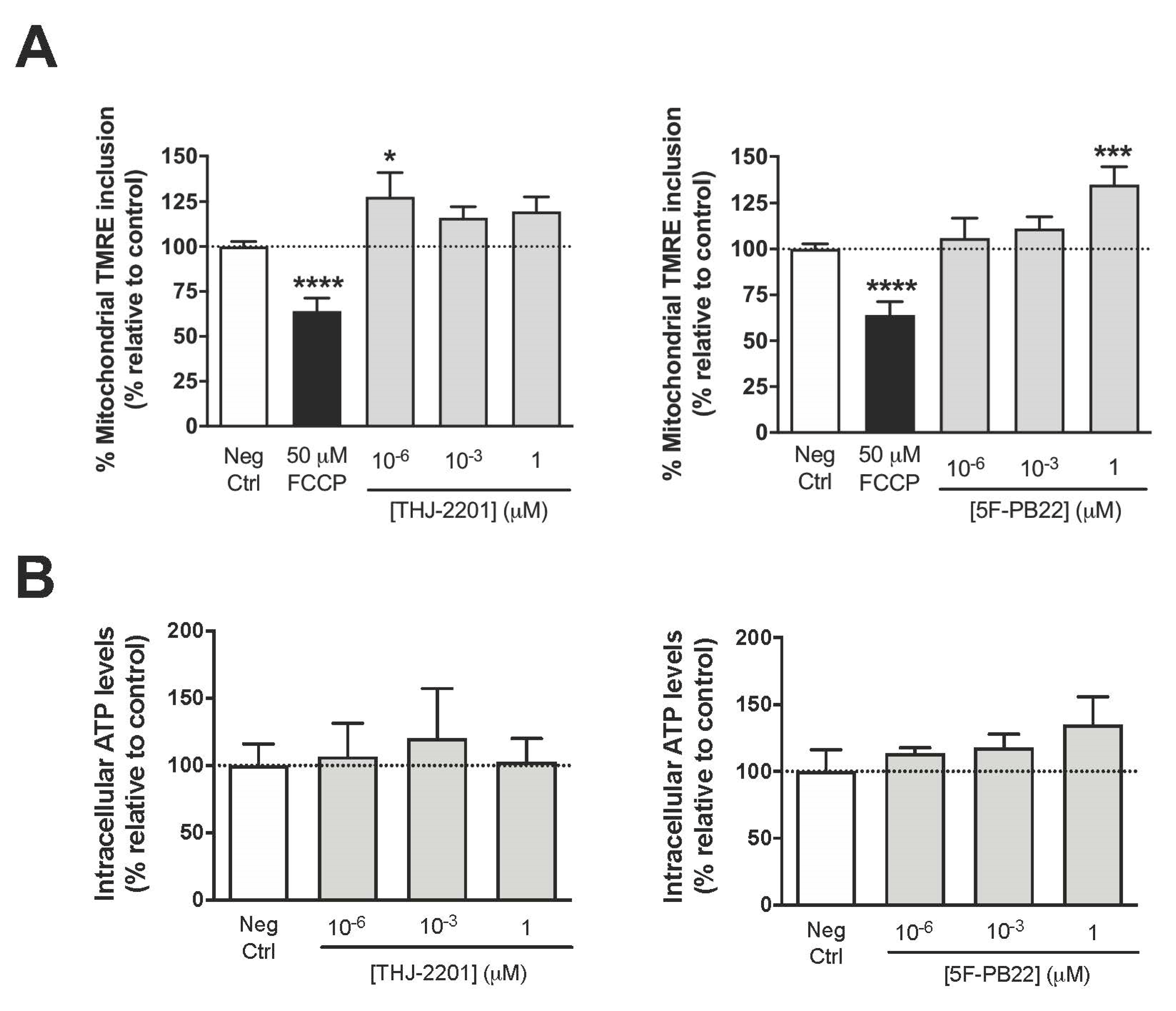
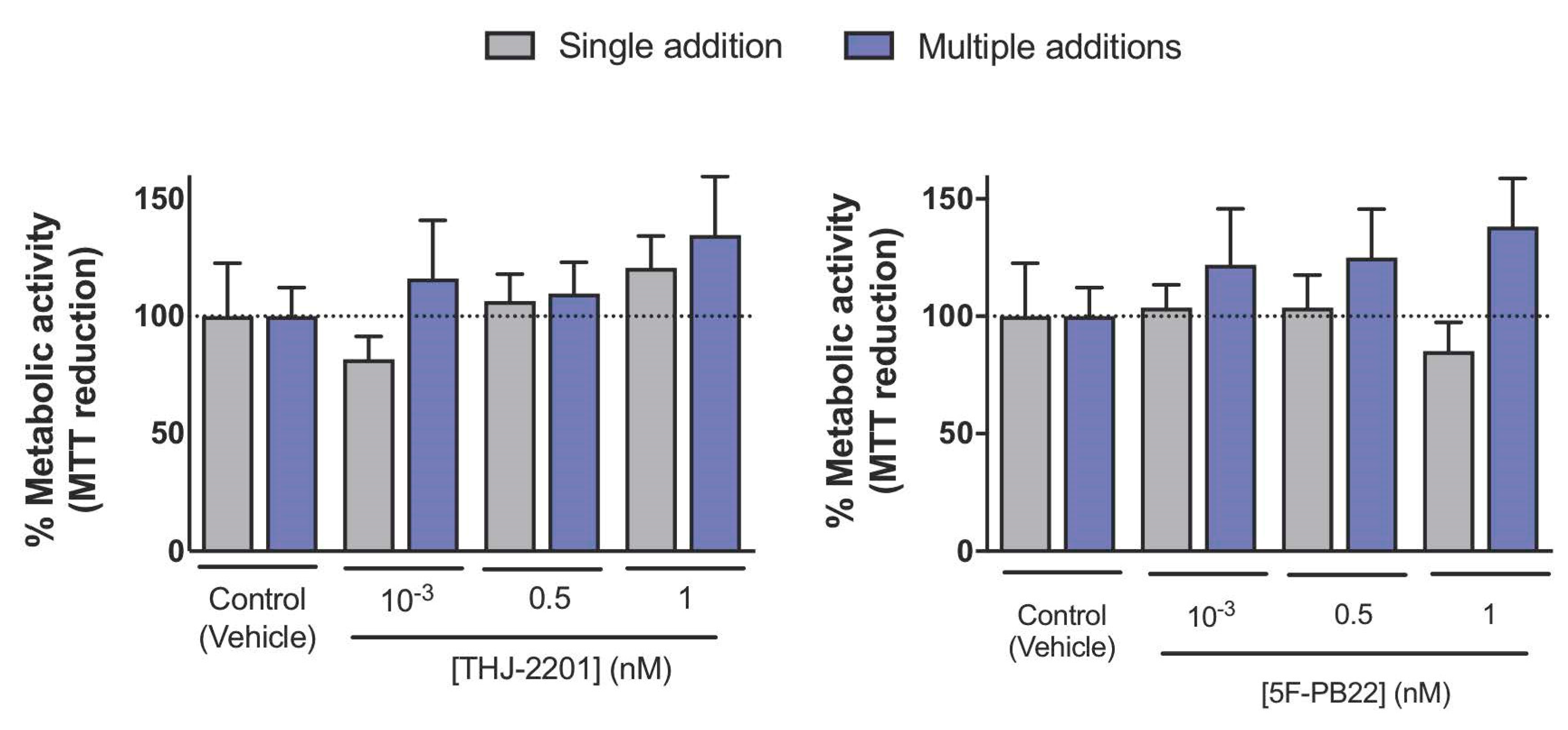
2.2. Mitochondrial Function Was Altered after Exposure to THJ-2201 and 5F-PB22
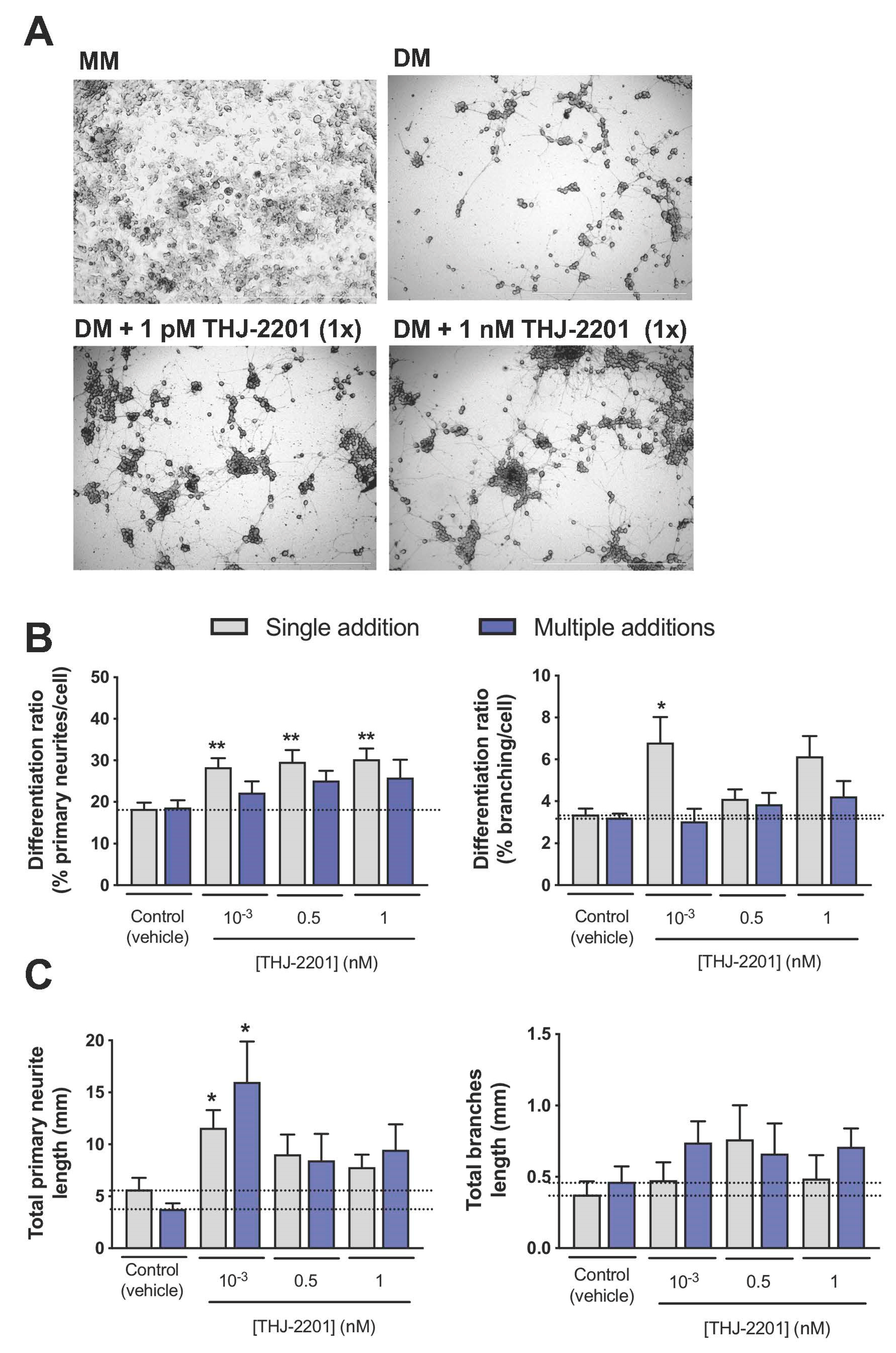
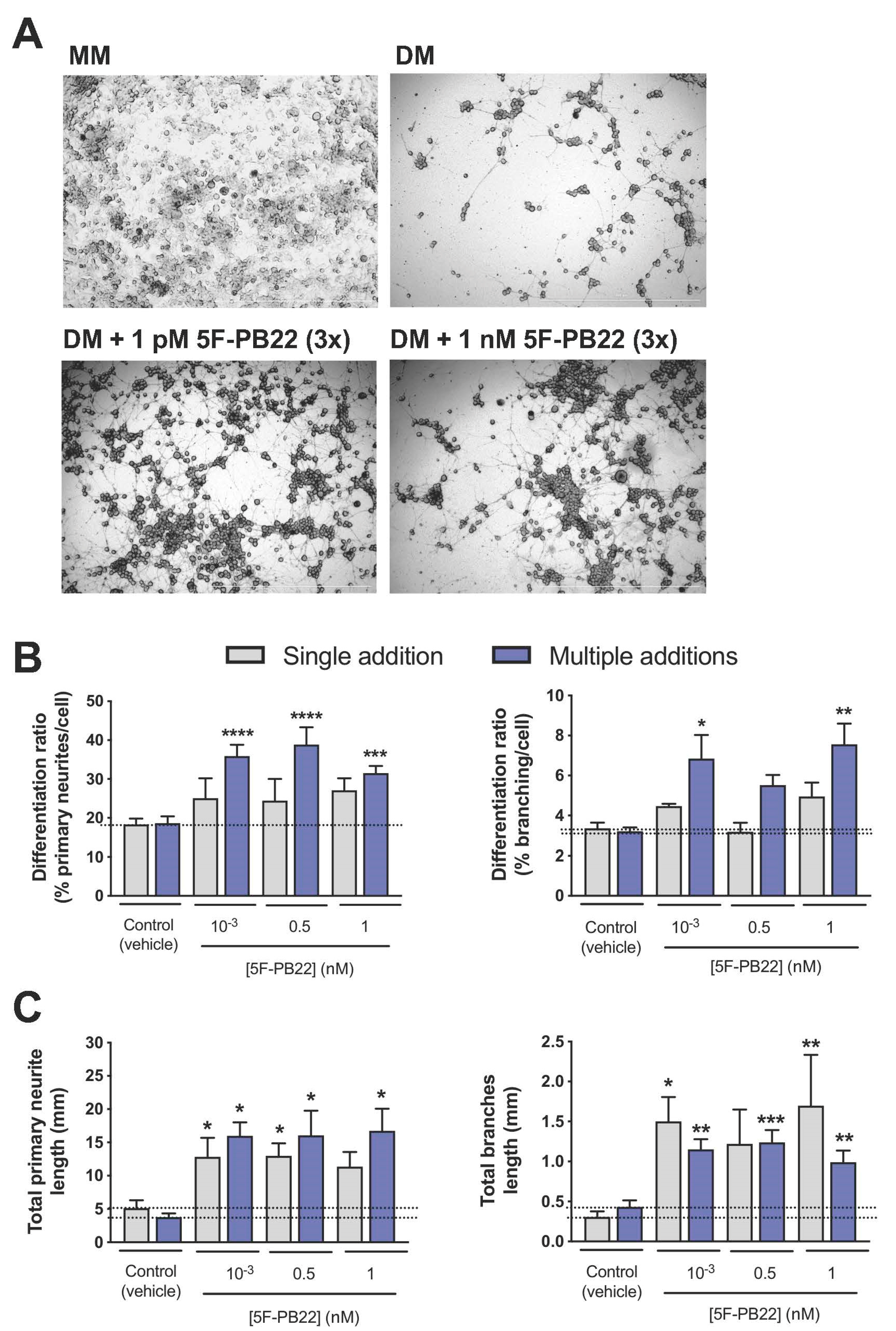
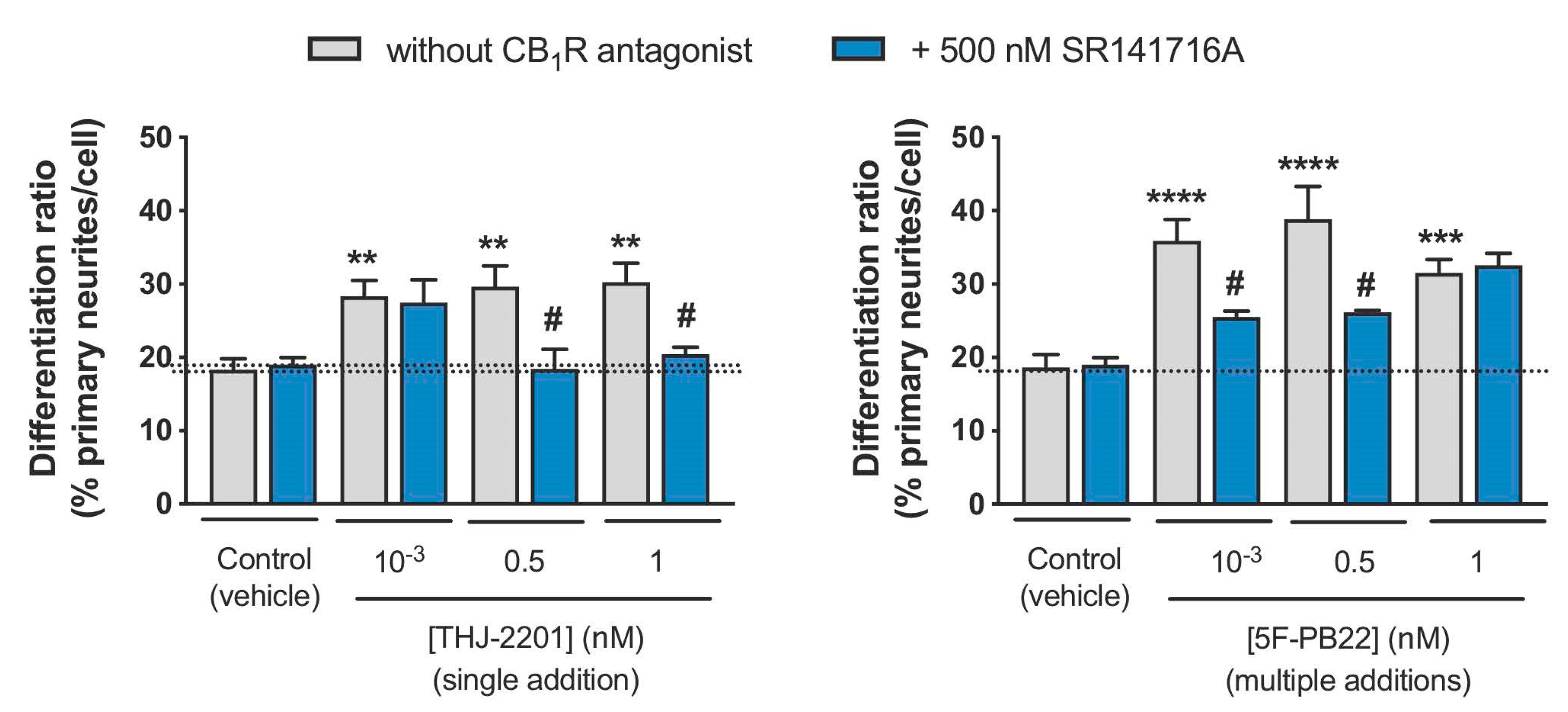
2.3. Both THJ-2201 and 5F-PB22 Promoted Neuronal Differentiation
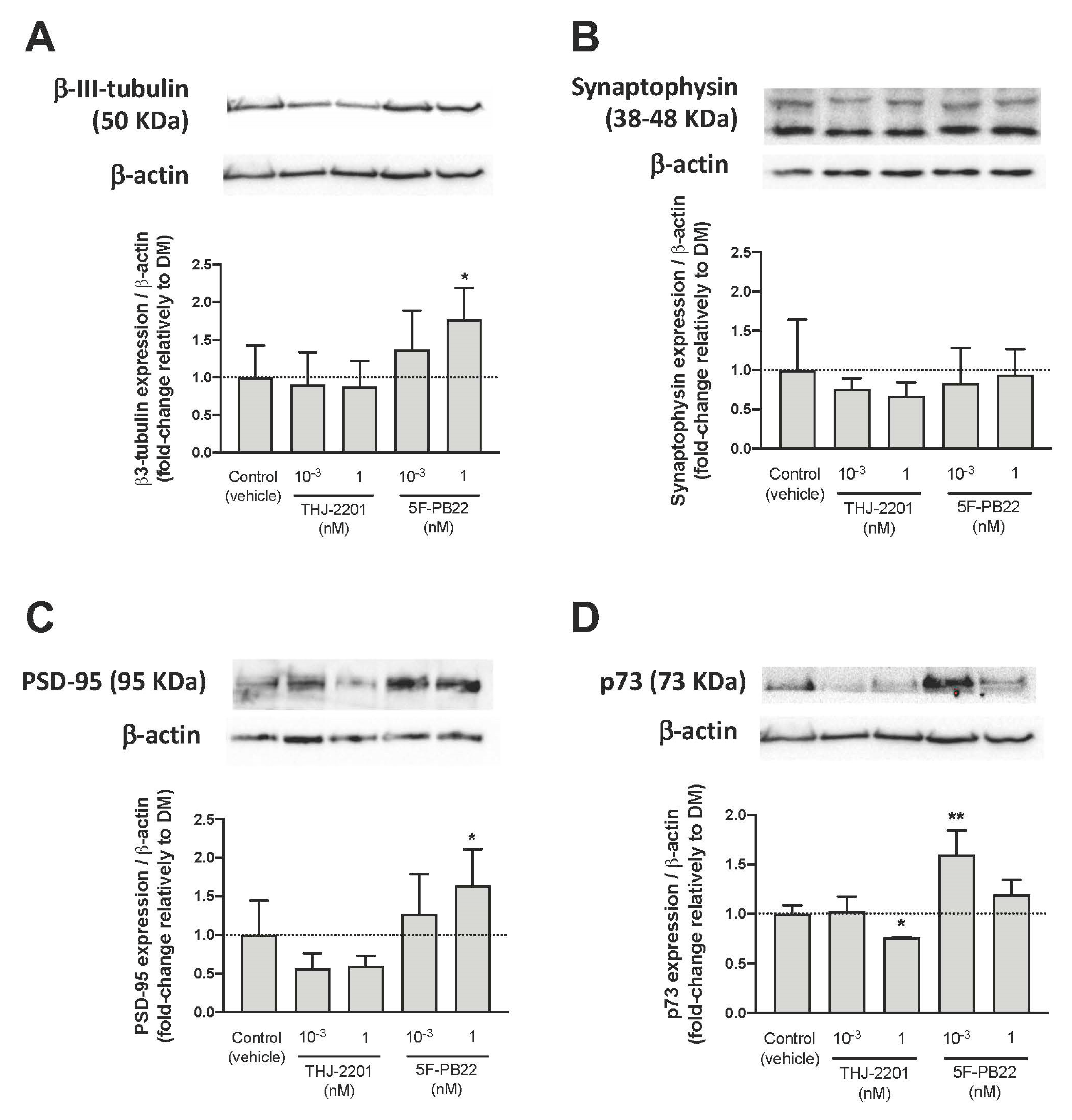
2.4. Expression of Neuronal Markers
3. Discussion
4. Materials and Methods
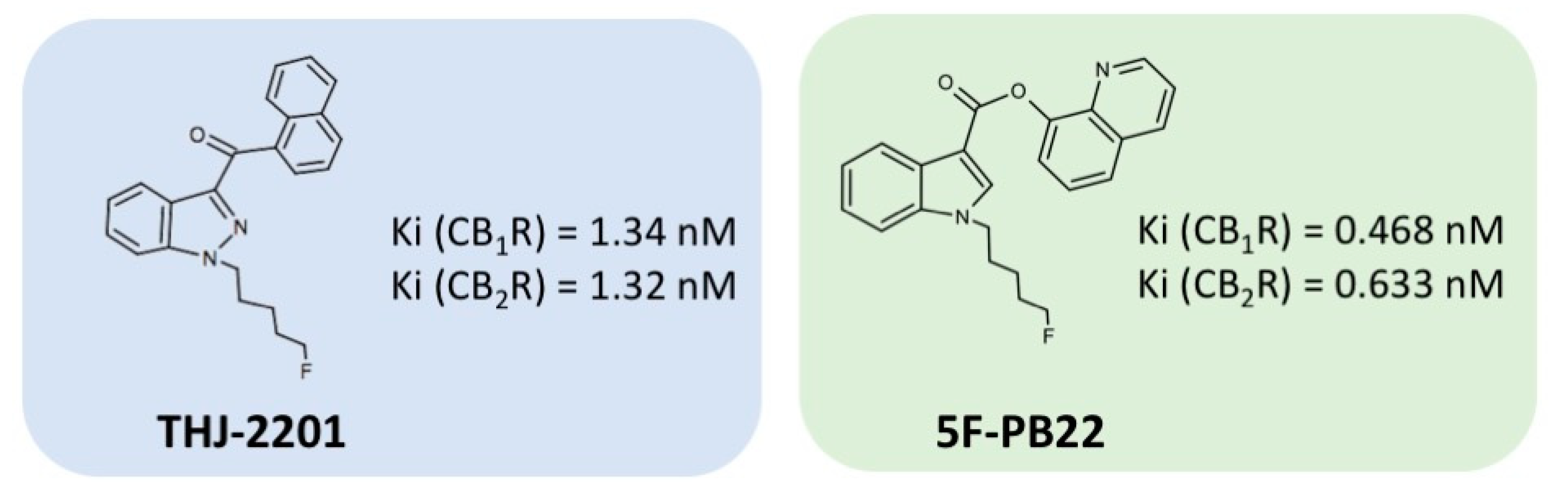
4.1. Chemicals
4.2. Cell Culture
4.3. Cell Viability
4.3.1. MTT Reduction Assay
4.3.2. Neutral Red Inclusion
4.3.3. Lactate Dehydrogenase (LDH) Release
4.4. Sulforhodamine B (SRB) Assay
4.5. Mitochondrial Function
4.6. Intracellular ATP Levels
4.7. Neuronal Differentiation
4.8. Total Protein Extraction
4.9. Western-Blot Analysis
4.10. Statistical Analysis
Author Contributions
Funding
Conflicts of Interest
References
- European Monitoring Center for Drugs and Drug Addiction (EMCDDA). European Drug Report 2019: Trends and Developments; Publications Office of the European Union: Luxembourg, 2019. [Google Scholar]
- Cristino, L.; Bisogno, T.; Di Marzo, V. Cannabinoids and the expanded endocannabinoid system in neurological disorders. Nat. Rev. Neurol. 2020, 16, 9–29. [Google Scholar] [CrossRef]
- Hall, W.; Stjepanović, D.; Caulkins, J.; Lynskey, M.; Leung, J.; Campbell, G.; Degenhardt, L. Public health implications of legalising the production and sale of cannabis for medicinal and recreational use. Lancet 2019, 394, 1580–1590. [Google Scholar] [CrossRef]
- Frau, R.; Miczán, V.; Traccis, F.; Aroni, S.; Pongor, C.I.; Saba, P.; Serra, V.; Sagheddu, C.; Fanni, S.; Congiu, M.; et al. Prenatal THC exposure produces a hyperdopaminergic phenotype rescued by pregnenolone. Nat. Neurosci. 2019, 22, 1975–1985. [Google Scholar] [CrossRef] [Green Version]
- Young-Wolff, K.C.; Sarovar, V.; Tucker, L.-Y.; Avalos, L.A.; Alexeeff, S.; Conway, A.; Armstrong, M.A.; Weisner, C.; Campbell, C.I.; Goler, N. Trends in marijuana use among pregnant women with and without nausea and vomiting in pregnancy, 2009–2016. Drug Alcohol. Depend. 2019, 196, 66–70. [Google Scholar] [CrossRef]
- Day, N.L.; Goldschmidt, L.; Day, R.; Larkby, C.; Richardson, G.A. Prenatal marijuana exposure, age of marijuana initiation, and the development of psychotic symptoms in young adults. Psychol. Med. 2015, 45, 1779–1787. [Google Scholar] [CrossRef]
- El Marroun, H.; Brown, Q.L.; Lund, I.O.; Coleman-Cowger, V.H.; Loree, A.M.; Chawla, D.; Washio, Y. An epidemiological, developmental and clinical overview of cannabis use during pregnancy. Prev. Med. 2018, 116, 1–5. [Google Scholar] [CrossRef]
- Pertwee, R.G.; Howlett, A.C.; Abood, M.E.; Alexander, S.P.H.; Di Marzo, V.; Elphick, M.R.; Greasley, P.J.; Hansen, H.S.; Kunos, G.; Mackie, K.; et al. International Union of Basic and Clinical Pharmacology. LXXIX. Cannabinoid receptors and their ligands: Beyond CB1 and CB2. Pharmacol. Rev. 2010, 62, 588–631. [Google Scholar] [CrossRef] [Green Version]
- Alexandre, J.; Carmo, H.; Carvalho, F.; Silva, J.P. Synthetic cannabinoids and their impact on neurodevelopmental processes. Addict. Biol. 2019, e12824. [Google Scholar] [CrossRef]
- Kim, D.; Thayer, S.A. Cannabinoids inhibit the formation of new synapses between hippocampal neurons in culture. J. Neurosci. 2001, 21, RC146. [Google Scholar] [CrossRef] [PubMed]
- Mereu, G.; Fa, M.; Ferraro, L.; Cagiano, R.; Antonelli, T.; Tattoli, M.; Ghiglieri, V.; Tanganelli, S.; Gessa, G.L.; Cuomo, V. Prenatal exposure to a cannabinoid agonist produces memory deficits linked to dysfunction in hippocampal long-term potentiation and glutamate release. Proc. Natl. Acad. Sci. USA 2003, 100, 4915–4920. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Psychoyos, D.; Hungund, B.; Cooper, T.; Finnell, R.H. A cannabinoid analogue of Δ9-tetrahydrocannabinol disrupts neural development in chick. Birth Defects Res. B Dev. Rep. Toxicol. 2008, 83, 477–488. [Google Scholar] [CrossRef] [PubMed]
- Jiang, W.; Zhang, Y.; Xiao, L.; Van Cleemput, J.; Ji, S.P.; Bai, G.; Zhang, X. Cannabinoids promote embryonic and adult hippocampus neurogenesis and produce anxiolytic- and antidepressant-like effects. J. Clin. Investig. 2005, 115, 3104–3116. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Palazuelos, J.; Ortega, Z.; Díaz-Alonso, J.; Guzmán, M.; Galve-Roperh, I. CB2 cnnabinoid receptors promote neural progenitor cell proliferation via mTORC1 signaling. J. Biol. Chem 2012, 287, 1198–1209. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Campanha, H.M.; Carvalho, F.; Schlosser, P.M. Active and peripheral anionic sites of acetylcholinesterase have differential modulation effects on cell proliferation, adhesion and neuritogenesis in the NG108-15 cell line. Toxicol. Lett. 2014, 230, 122–131. [Google Scholar] [CrossRef]
- Tojima, T.; Yamane, Y.; Takahashi, M.; Ito, E. Acquisition of neuronal proteins during differentiation of NG108-15 cells. Neurosci. Res. 2000, 37, 153–161. [Google Scholar] [CrossRef]
- Chemin, J.; Nargeot, J.; Lory, P. Neuronal T-type α1H calcium channels induce neuritogenesis and expression of high-voltage-Activated calcium channels in the NG108–15 cell line. J. Neurosci. 2002, 22, 6856–6862. [Google Scholar] [CrossRef]
- Stella, N. Cannabinoid and cannabinoid-like receptors in microglia, astrocytes and astrocytomas. Glia 2010, 58, 1017–1030. [Google Scholar] [CrossRef] [Green Version]
- Ernst, C. Proliferation and differentiation deficits are a major convergence point for neurodevelopmental disorders. Trends Neurosci. 2016, 39, 290–299. [Google Scholar] [CrossRef]
- Wyatt, C.N.; Buckler, K.J. The effect of mitochondrial inhibitors on membrane currents in isolated neonatal rat carotid body type I cells. J. Physiol. 2004, 556, 175–191. [Google Scholar] [CrossRef]
- Bagkos, G.; Koufopoulos, K.; Piperi, C. A new model for mitochondrial membrane potential production and storage. Med. Hypotheses 2014, 83, 175–181. [Google Scholar] [CrossRef]
- Harris, J.J.; Jolivet, R.; Attwell, D. Synaptic energy use and supply. Neuron 2012, 75, 762–777. [Google Scholar] [CrossRef] [Green Version]
- Magistretti, P.J.; Allaman, I. A cellular perspective on brain energy metabolism and functional imaging. Neuron 2015, 86, 883–901. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cao, S.; Du, J.; Lv, Y.; Lin, H.; Mao, Z.; Xu, M.; Liu, M.; Liu, Y. PAX3 inhibits beta-Tubulin-III expression and neuronal differentiation of neural stem cell. Biochem. Biophys. Res. Commun. 2017, 485, 307–311. [Google Scholar] [CrossRef] [PubMed]
- Kwon, S.E.; Chapman, E.R. Synaptophysin regulates the kinetics of synaptic vesicle endocytosis in central neurons. Neuron 2011, 70, 847–854. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- El-Husseini, A.E.-D.; Schnell, E.; Chetkovich, D.M.; Nicoll, R.A.; Bredt, D.S. PSD-95 Involvement in Maturation of Excitatory Synapses. Science 2000, 290, 1364–1368. [Google Scholar]
- De Laurenzi, V.; Raschellá, G.; Barcaroli, D.; Annicchiarico-Petruzzelli, M.; Ranalli, M.; Catani, M.V.; Tanno, B.; Costanzo, A.; Levrero, M.; Melino, G. Induction of neuronal differentiation by p73 in a neuroblastoma cell line. J. Biol. Chem. 2000, 275, 15226–15231. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Velletri, T.; Romeo, F.; Tucci, P.; Peschiaroli, A.; Annicchiarico-Petruzzelli, M.; Niklison-Chirou, M.V.; Amelio, I.; Knight, R.A.; Mak, T.W.; Melino, G.; et al. GLS2 is transcriptionally regulated by p73 and contributes to neuronal differentiation. Cell Cycle 2013, 12, 3564–3573. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- European Monitoring Center for Drugs and Drug Addiction (EMCDDA). Fentanils and Synthetic Cannabinoids: Driving Greater Complexity into the Drug Situation. An Update from the EU Early Warning System; Publications Office of the European Union: Luxembourg, 2018. [Google Scholar]
- Silva, J.P.; Araújo, A.M.; Carmo, H.; Guedes de Pinho, P.; Carvalho, F. Synthetic cannabinoids JWH-122 and THJ-2201 disrupt endocannabinoid-regulated mitochondrial function and activate apoptotic pathways as a primary mechanism of in vitro nephrotoxicity at in vivo relevant concentrations. Toxicol. Sci. 2019, 169, 422–435. [Google Scholar] [CrossRef]
- Djeungoue-Petga, M.A.; Hebert-Chatelain, E. Linking mitochondria and synaptic transmission: The CB1 receptor. Bioessays 2017, 39. [Google Scholar] [CrossRef]
- Khacho, M.; Harris, R.; Slack, R.S. Mitochondria as central regulators of neural stem cell fate and cognitive function. Nat. Rev. Neurosci. 2019, 20, 34–48. [Google Scholar] [CrossRef]
- Compagnucci, C.; Di Siena, S.; Bustamante, M.B.; Di Giacomo, D.; Di Tommaso, M.; Maccarrone, M.; Grimaldi, P.; Sette, C. Type-1 (CB1) cannabinoid receptor promotes neuronal differentiation and maturation of neural stem cells. PLoS ONE 2013, 8, e54271. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jordan, J.D.; He, J.C.; Eungdamrong, N.J.; Gomes, I.; Ali, W.; Nguyen, T.; Bivona, T.G.; Philips, M.R.; Devi, L.A.; Iyengar, R. Cannabinoid Receptor-induced Neurite outgrowth is mediated by Rap1 activation through Gαo/i-triggered proteasomal degradation of Rap1GAPII. J. Biol. 2005, 280, 11413–11421. [Google Scholar]
- Basavarajappa, B.S.; Subbanna, S. Potential Mechanisms Underlying the Deleterious Effects of Synthetic Cannabinoids Found in Spice/K2 Products. Brain Sci. 2019, 9, 14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fay, J.F.; Farrens, D.L. Structural dynamics and energetics underlying allosteric inactivation of the cannabinoid receptor CB1. Proc. Natl. Acad. Sci. USA 2015, 112, 8469–8474. [Google Scholar] [CrossRef] [Green Version]
- Bambico, F.R.; Katz, N.; Debonnel, G.; Gobbi, G. Cannabinoids elicit antidepressant-like behavior and activate serotonergic neurons through the medial prefrontal cortex. J. Neurosci. 2007, 27, 11700–11711. [Google Scholar] [CrossRef]
- Laprairie, R.B.; Bagher, A.M.; Kelly, M.E.M.; Denovan-Wright, E.M. Biased Type 1 cannabinoid receptor signaling influences neuronal viability in a cell culture model of Huntington disease. Mol. Pharmacol. 2016, 89, 364–375. [Google Scholar] [CrossRef] [Green Version]
- Zheng, S.; Gray, E.E.; Chawla, G.; Porse, B.T.; O’Dell, T.J.; Black, D.L. PSD-95 is post-transcriptionally repressed during early neural development by PTBP1 and PTBP2. Nat. Neurosci. 2012, 15, 381. [Google Scholar] [CrossRef] [Green Version]
- Katsetos, C.D.; Legido, A.; Perentes, E.; Mörk, S.J. Class III β-tubulin isotype: A key cytoskeletal protein at the crossroads of developmental neurobiology and tumor neuropathology. J. Child. Neurol. 2003, 18, 851–866. [Google Scholar] [CrossRef]
- Liu, C.; Zhong, Y.; Apostolou, A.; Fang, S. Neural differentiation of human embryonic stem cells as an in vitro tool for the study of the expression patterns of the neuronal cytoskeleton during neurogenesis. Biochem. Biophys. Res. Commun. 2013, 439, 154–159. [Google Scholar] [CrossRef]
- Fan, W.-j.; Li, X.; Yao, H.-l.; Deng, J.-x.; Liu, H.-l.; Cui, Z.-j.; Wang, Q.; Wu, P.; Deng, J.-b. Neural differentiation and synaptogenesis in retinal development. Neural. Regen Res. 2016, 11, 312–318. [Google Scholar]
- Saillour, Y.; Broix, L.; Bruel-Jungerman, E.; Lebrun, N.; Muraca, G.; Rucci, J.; Poirier, K.; Belvindrah, R.; Francis, F.; Chelly, J. Beta tubulin isoforms are not interchangeable for rescuing impaired radial migration due to Tubb3 knockdown. Hum. Mol. Gen. 2014, 23, 1516–1526. [Google Scholar] [CrossRef] [PubMed]
- Ly, C.; Greb, A.C.; Cameron, L.P.; Wong, J.M.; Barragan, E.V.; Wilson, P.C.; Burbach, K.F.; Soltanzadeh Zarandi, S.; Sood, A.; Paddy, M.R.; et al. Psychedelics promote structural and functional neural plasticity. Cell Rep. 2018, 23, 3170–3182. [Google Scholar] [CrossRef] [PubMed]
- Hill, A.S.; Sahay, A.; Hen, R. Increasing adult hippocampal neurogenesis is sufficient to reduce anxiety and depression-like behaviors. Neuropsychopharmacology 2015, 40, 2368–2378. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, X.; Zhao, X. Epigenetic regulation of mammalian stem cells. Stem Cells Dev. 2008, 17, 1043–1052. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.P.; Fu, R.H.; Yu, H.H.; Li, K.W.; Tsai, C.H.; Shyu, W.C.; Lin, S.Z. MicroRNAs regulation modulated self-renewal and lineage differentiation of stem cells. Cell Transplant. 2009, 18, 1039–1045. [Google Scholar] [CrossRef] [PubMed]
- Gilbert, M.T.; Sulik, K.K.; Fish, E.W.; Baker, L.K.; Dehart, D.B.; Parnell, S.E. Dose-dependent teratogenicity of the synthetic cannabinoid CP-55,940 in mice. Neurotoxicol. Teratol. 2016, 58, 15–22. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ferreira, F.F.; Ribeiro, F.F.; Rodrigues, R.S.; Sebastião, A.M.; Xapelli, S. Brain-derived neurotrophic factor (BDNF) role in cannabinoid-mediated neurogenesis. Front. Cell Neurosci. 2018, 12, 441. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jacobsson, S.O.P.; Wallin, T.; Fowler, C.J. Inhibition of rat C6 glioma cell proliferation by endogenous and synthetic cannabinoids. Relative involvement of cannabinoid and vanilloid receptors. J. Pharmacol. Exp. Therap. 2001, 299, 951–959. [Google Scholar]
- Ibsen, M.S.; Connor, M.; Glass, M. Cannabinoid CB(1) and CB(2) receptor signaling and bias. Cannabis Cannabinoid Res. 2017, 2, 48–60. [Google Scholar] [CrossRef] [Green Version]
- Atwood, B.K.; Mackie, K. CB2: A cannabinoid receptor with an identity crisis. Br. J. Pharmacol. 2010, 160, 467–479. [Google Scholar] [CrossRef] [Green Version]
- Uchiyama, N.; Kikura-Hanajiri, R.; Goda, Y. Identification of a novel cannabimimetic phenylacetylindole, cannabipiperidiethanone, as a designer drug in a herbal product and its affinity for cannabinoid CB1 and CB2 receptors. Chem. Pharm. Bull. 2011, 59, 1203–1205. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hess, C.; Schoeder, C.T.; Pillaiyar, T.; Madea, B.; Müller, C.E. Pharmacological evaluation of synthetic cannabinoids identified as constituents of spice. Forensic Toxicol. 2016, 34, 329–343. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Whitemarsh, R.C.M.; Pier, C.L.; Tepp, W.H.; Pellett, S.; Johnson, E.A. Model for studying Clostridium botulinum neurotoxin using differentiated motor neuron-like NG108-15 cells. Biochem. Biophys. Res. Commun. 2012, 427, 426–430. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chang, C.-Y.; Ma, K.-H.; Wang, J.-K.; Tung, Y.-L.; Chueh, S.-H. Inhibition of protein kinase C promotes differentiation of neuroblastoma × glioma NG108-15 hybrid cells. Genes Dev. 2011, 34, 1074–1084. [Google Scholar] [CrossRef]
- Dias da Silva, D.; Ferreira, B.; Roque Bravo, R.; Rebelo, R.; Duarte de Almeida, T.; Valente, M.J.; Silva, J.P.; Carvalho, F.; Bastos, M.L.; Carmo, H. The new psychoactive substance 3-methylmethcathinone (3-MMC or metaphedrone) induces oxidative stress, apoptosis, and autophagy in primary rat hepatocytes at human-relevant concentrations. Arch. Toxicol. 2019, 93, 2617–2634. [Google Scholar] [CrossRef]
- Repetto, G.; del Peso, A.; Zurita, J.L. Neutral red uptake assay for the estimation of cell viability/cytotoxicity. Nat. Protoc. 2008, 3, 1125. [Google Scholar] [CrossRef]
- Arbo, M.D.; Silva, R.; Barbosa, D.J.; da Silva, D.D.; Silva, S.P.; Teixeira, J.P.; Bastos, M.L.; Carmo, H. In vitro neurotoxicity evaluation of piperazine designer drugs in differentiated human neuroblastoma SH-SY5Y cells. J. Appl. Toxicol. 2016, 36, 121–130. [Google Scholar] [CrossRef]
- Silva, J.P.; Carmo, H.; Carvalho, F. The synthetic cannabinoid XLR-11 induces in vitro nephrotoxicity by impairment of endocannabinoid-mediated regulation of mitochondrial function homeostasis and triggering of apoptosis. Toxicol. Lett. 2018, 287, 59–69. [Google Scholar] [CrossRef]
- Silva, J.P.; Sardao, V.A.; Coutinho, O.P.; Oliveira, P.J. Nitrogen compounds prevent H9c2 myoblast oxidative stress-induced mitochondrial dysfunction and cell death. Cardiovasc. Toxicol. 2010, 10, 51–65. [Google Scholar] [CrossRef] [Green Version]
- Papazisis, K.T.; Geromichalos, G.D.; Dimitriadis, K.A.; Kortsaris, A.H. Optimization of the sulforhodamine B colorimetric assay. J. Immunol. Methods 1997, 208, 151–158. [Google Scholar] [CrossRef]
- Ehrenberg, B.; Montana, V.; Wei, M.D.; Wuskell, J.P.; Loew, L.M. Membrane potential can be determined in individual cells from the nernstian distribution of cationic dyes. Biophys. J. 1988, 53, 785–794. [Google Scholar] [CrossRef] [Green Version]
- Allen, D.G.; Lannergren, J.; Westerblad, H. Intracellular ATP measured with luciferin/luciferase in isolated single mouse skeletal muscle fibres. Pflügers Arch. 2002, 443, 836–842. [Google Scholar] [CrossRef] [PubMed]
- Chao, C.C.; Chan, P.; Kuo, C.S.; Gong, C.L.; Cheng, T.H.; Liu, Z.M.; Shen, P.C.; Huang, C.C.; Leung, Y.M. Protection of differentiated neuronal NG108-15 cells from P2X7 receptor-mediated toxicity by taurine. Pharmacol. Rep. 2014, 66, 576–584. [Google Scholar] [CrossRef] [PubMed]
- Dias da Silva, D.; Silva, E.; Carmo, H. Cytotoxic effects of amphetamine mixtures in primary hepatocytes are severely aggravated under hyperthermic conditions. Toxicol. In Vitro 2013, 27, 1670–1678. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Drug | Parameters for the Logit Regression Model | EC50 (mM) | NOEC (μM) | LOEC (μM) | ||
|---|---|---|---|---|---|---|
| θ1 a | θ2 b | θmax c | ||||
| 5F-PB22 | −0.68 | 1.54 | 141.80 | 1.12 | 5.86 | 11.72 |
| THJ-2201 | 1.03 | 3.86 | 85.78 | 0.66 | 78.12 | 150 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Alexandre, J.; Malheiro, R.; Dias da Silva, D.; Carmo, H.; Carvalho, F.; Silva, J.P. The Synthetic Cannabinoids THJ-2201 and 5F-PB22 Enhance In Vitro CB1 Receptor-Mediated Neuronal Differentiation at Biologically Relevant Concentrations. Int. J. Mol. Sci. 2020, 21, 6277. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21176277
Alexandre J, Malheiro R, Dias da Silva D, Carmo H, Carvalho F, Silva JP. The Synthetic Cannabinoids THJ-2201 and 5F-PB22 Enhance In Vitro CB1 Receptor-Mediated Neuronal Differentiation at Biologically Relevant Concentrations. International Journal of Molecular Sciences. 2020; 21(17):6277. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21176277
Chicago/Turabian StyleAlexandre, João, Rui Malheiro, Diana Dias da Silva, Helena Carmo, Félix Carvalho, and João Pedro Silva. 2020. "The Synthetic Cannabinoids THJ-2201 and 5F-PB22 Enhance In Vitro CB1 Receptor-Mediated Neuronal Differentiation at Biologically Relevant Concentrations" International Journal of Molecular Sciences 21, no. 17: 6277. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21176277