Decellularized Extracellular Matrices and Cardiac Differentiation: Study on Human Amniotic Fluid-Stem Cells

 , , , , and
, , , , and
Abstract
:1. Introduction
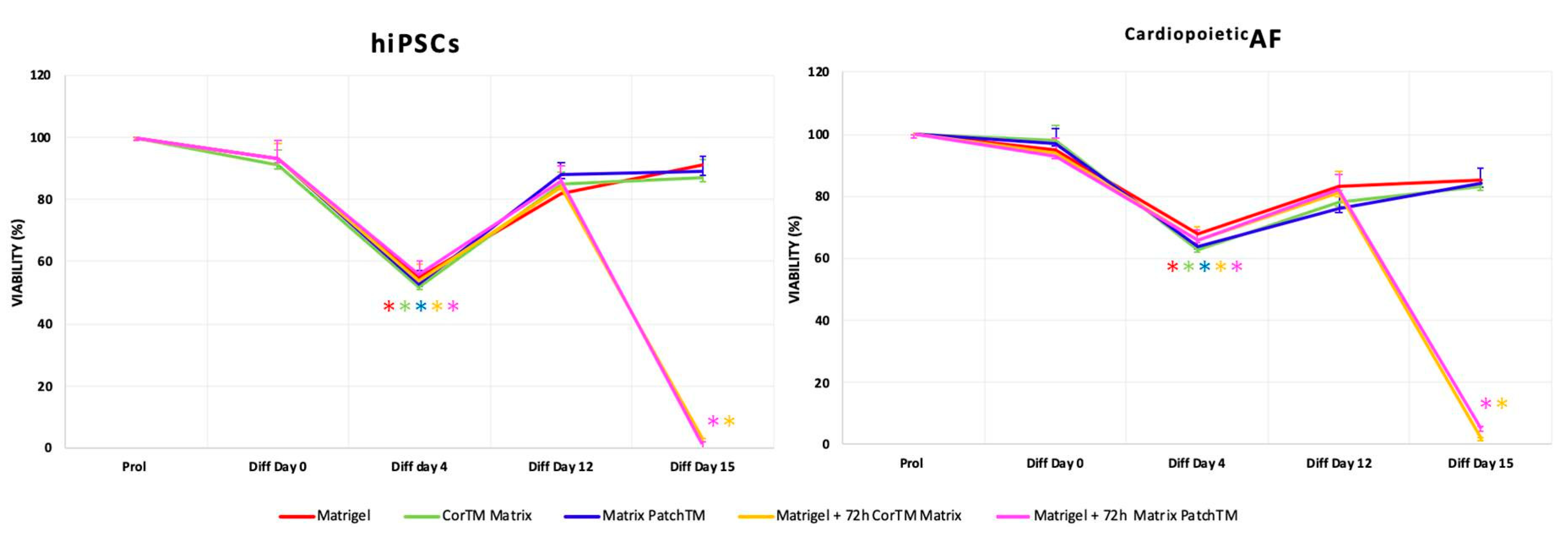
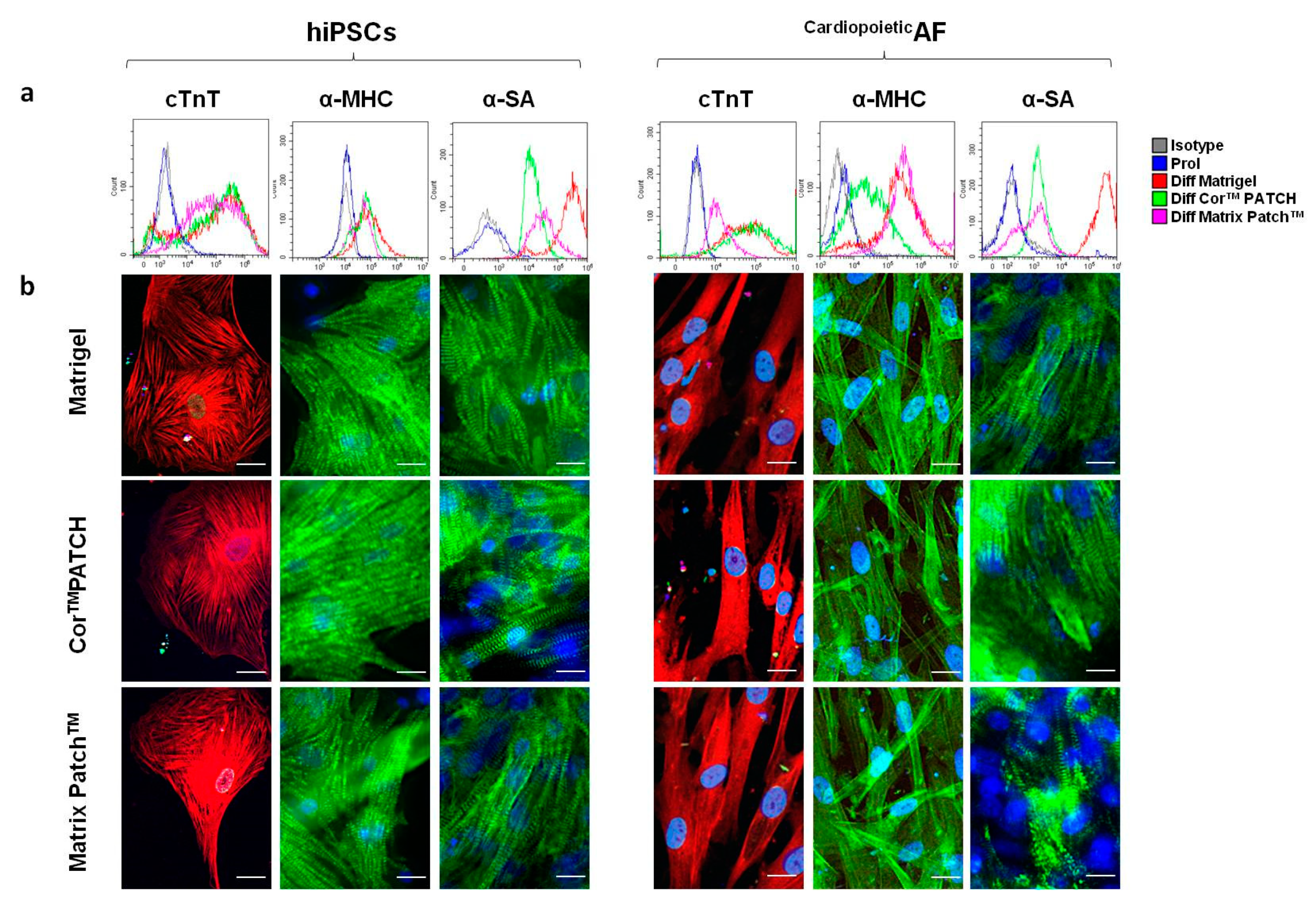
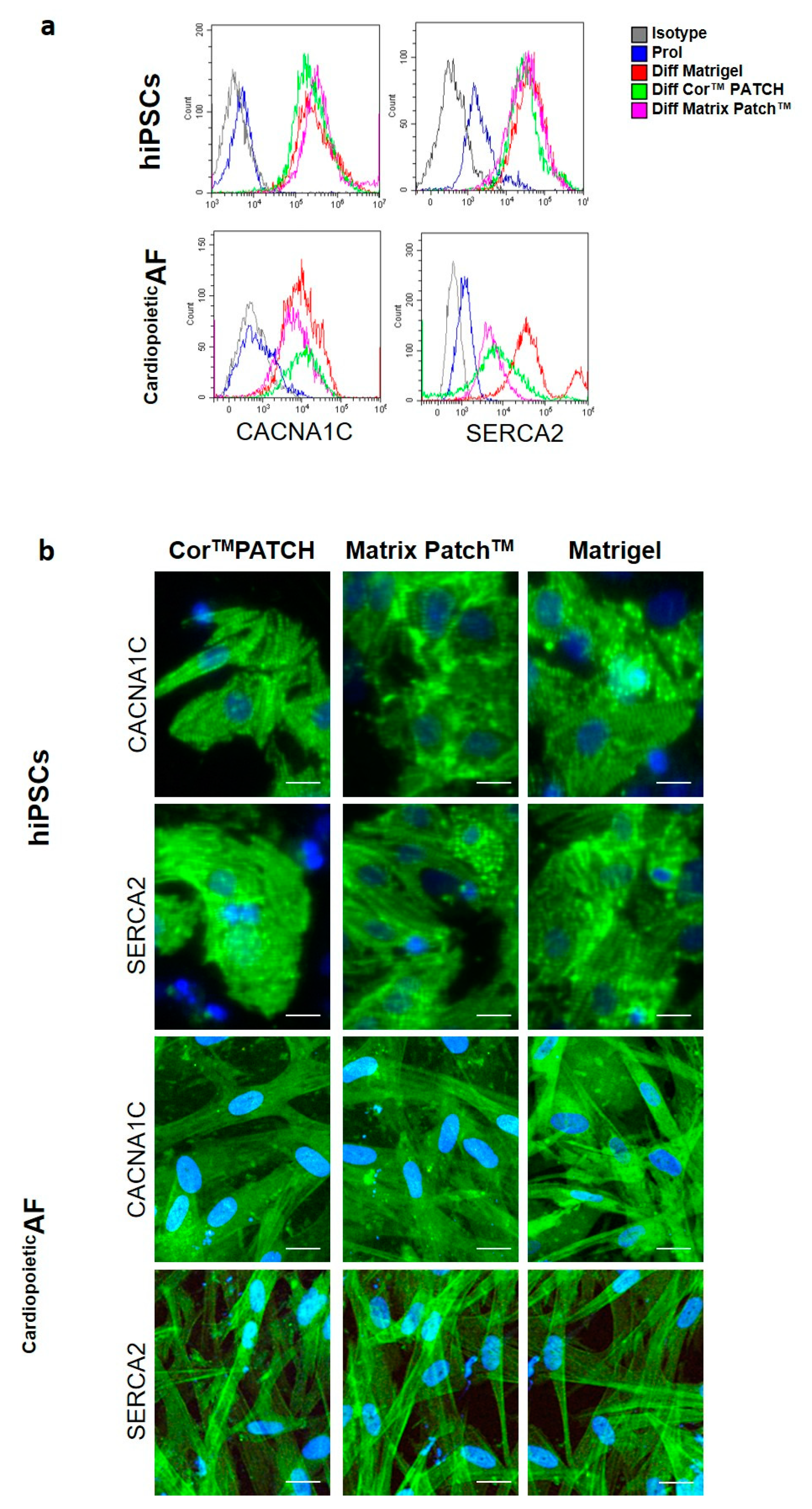
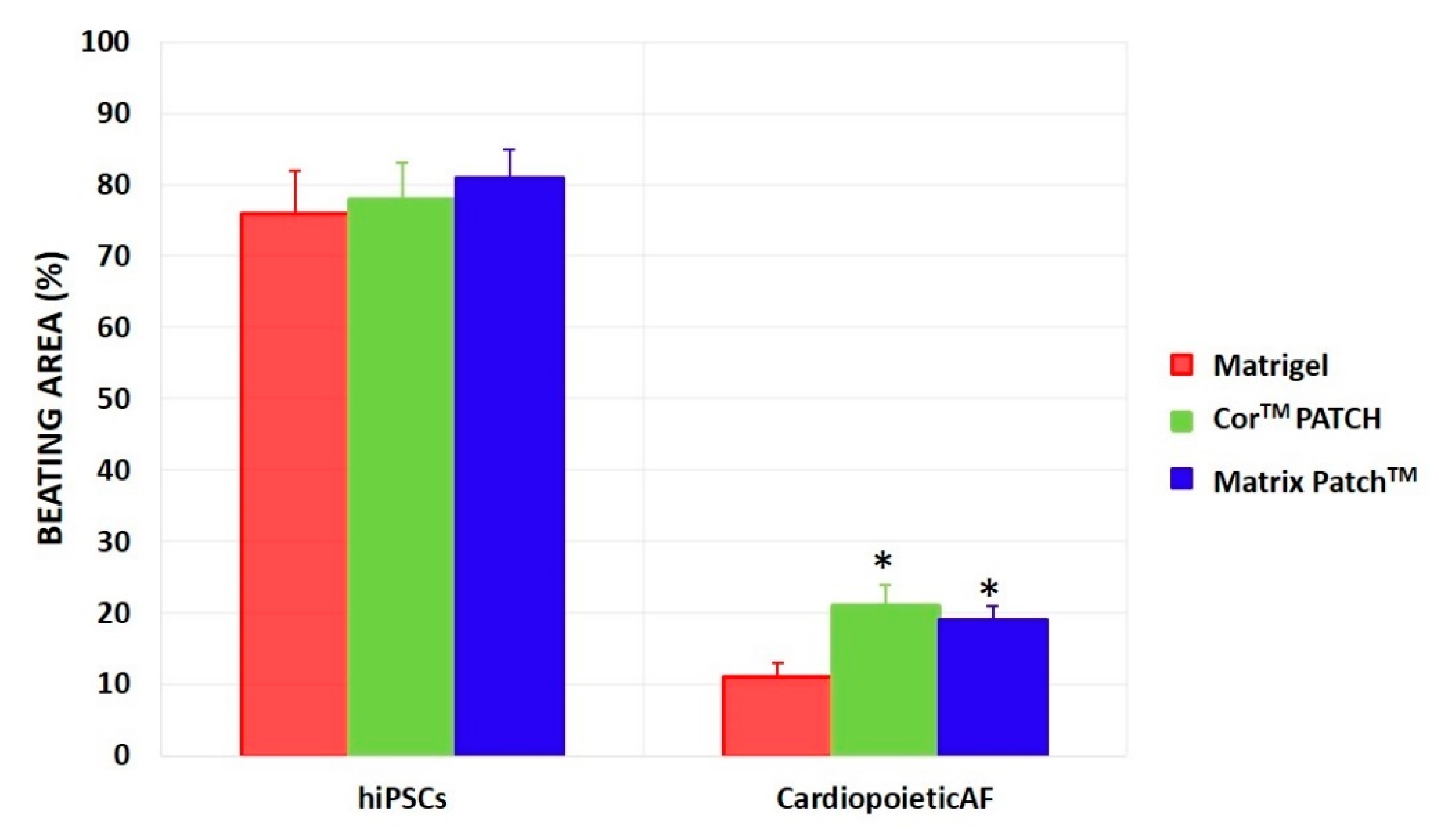
2. Results
- (1)
- (2)
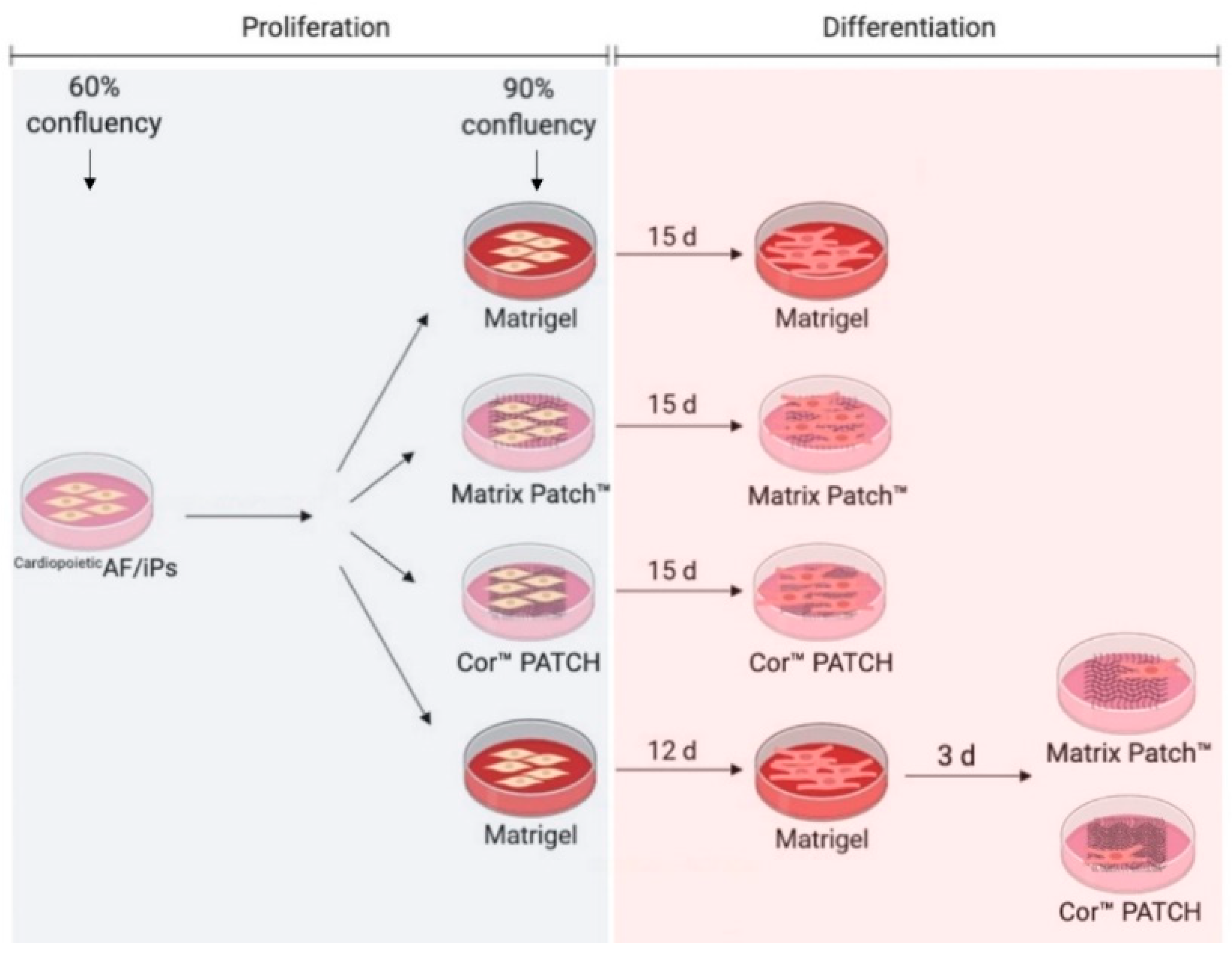
- The CardiopoieticAF and the hiPSCs were differentiated in a monolayer onto Matrigel® for 12 days and then dissociated and moved to the dECMs for 72 h before analysis.
3. Discussion
4. Materials and Methods
4.1. Isolation and Culture of CardiopoieticAF and hiPSC Cells
4.2. Culture in Monolayer and Cardiac Differentiation
4.3. MTT(3(4,5-Dimethylthiazol-2yl)-2,5-Diphenyl Tetrazolium Bromide) Assay
4.4. Flow Cytometry and Imaging Flow Cytometry
4.5. Immunofluorescence Staining and Microscopy
4.6. Statistical Analysis
Author Contributions
Funding
Conflicts of Interest
References
- Bleumink, G.; Knetsch, A.; Sturkenboom, M.; Straus, S.; Hofman, A.; Deckers, J.; Witteman, J.; Stricker, B. Quantifying the heart failure epidemic: Prevalence, incidence rate, lifetime risk and prognosis of heart failure: The Rotterdam Study. Eur. Heart J. 2004, 25, 1614–1619. [Google Scholar] [CrossRef] [Green Version]
- Di Baldassarre, A.; D’Amico, M.A.; Izzicupo, P.; Gaggi, G.; Guarnieri, S.; Mariggiò, M.A.; Antonucci, I.; Corneo, B.; Sirabella, D.; Stuppia, L.; et al. Cardiomyocytes Derived from Human CardiopoieticAmniotic Fluids. Sci. Rep. 2018, 8, 12028. [Google Scholar] [CrossRef] [PubMed]
- Iop, L.; Palmosi, T.; Dal Sasso, E.; Gerosa, G. Bioengineered tissue solutions for repair, correction and reconstruction in cardiovascular surgery. J. Thorac. Dis. 2018, 10, S2390–S2411. [Google Scholar] [CrossRef] [PubMed]
- Ponikowski, P.; Voors, A.A.; Anker, S.D.; Bueno, H.; Cleland, J.G.F.; Coats, A.J.S.; Falk, V.; González-Juanatey, J.R.; Harjola, V.-P.; Jankowska, E.A.; et al. 2016 ESC Guidelines for the diagnosis and treatment of acute and chronic heart failure: The Task Force for the diagnosis and treatment of acute and chronic heart failure of the European Society of Cardiology (ESC). Developed with the spec. Eur. J. Heart Fail. 2016, 18, 891–975. [Google Scholar] [CrossRef] [PubMed]
- Di Baldassarre, A.; Cimetta, E.; Bollini, S.; Gaggi, G.; Ghinassi, B. Human-Induced Pluripotent Stem Cell Technology and Cardiomyocyte Generation: Progress and Clinical Applications. Cells 2018, 7, 48. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Filardi, T.; Ghinassi, B.; Di Baldassarre, A.; Tanzilli, G.; Morano, S.; Lenzi, A.; Basili, S.; Crescioli, C. Cardiomyopathy Associated with Diabetes: The Central Role of the Cardiomyocyte. Int. J. Mol. Sci. 2019, 20, 3299. [Google Scholar] [CrossRef] [Green Version]
- Lian, X.; Zhang, J.; Azarin, S.M.; Zhu, K.; Hazeltine, L.B.; Bao, X.; Hsiao, C.; Kamp, T.J.; Palecek, S.P. Directed cardiomyocyte differentiation from human pluripotent stem cells by modulating Wnt/β-catenin signaling under fully defined conditions. Nat. Protoc. 2013, 8, 162–175. [Google Scholar] [CrossRef] [Green Version]
- Burridge, P.W.; Keller, G.; Gold, J.D.; Wu, J.C. Production of De Novo Cardiomyocytes: Human Pluripotent Stem Cell Differentiation and Direct Reprogramming. Cell Stem Cell 2012, 10, 16–28. [Google Scholar] [CrossRef] [Green Version]
- Antonucci, I.; Di Pietro, R.; Alfonsi, M.; Centurione, M.A.; Centurione, L.; Sancilio, S.; Pelagatti, F.; D’amico, M.A.; Di Baldassarre, A.; Piattelli, A.; et al. Human Second Trimester Amniotic Fluid Cells are Able to Create Embryoid Body-Like Structures in Vitro and to Show Typical Expression Profiles of Embryonic and Primordial Germ Cells. Cell Transplant. 2014, 23, 1501–1515. [Google Scholar] [CrossRef]
- Gaggi, G.; Di Credico, A.; Izzicupo, P.; Antonucci, I.; Crescioli, C.; Di Giacomo, V.; Di Ruscio, A.; Amabile, G.; Alviano, F.; Di Baldassarre, A.; et al. Epigenetic Features of Human Perinatal Stem Cells Redefine Their Stemness Potential. Cells 2020, 9, 1304. [Google Scholar] [CrossRef]
- Conboy, I.; Freimer, J.; Weisenstein, L.; Liu, Y.; Mehdipour, M.; Gathwala, R. 6.13 Tissue Engineering of Muscle Tissue ☆. In Comprehensive Biomaterials II; Elsevier: Amsterdam, The Netherlands, 2017; pp. 216–235. ISBN 978-0-08-100692-4. [Google Scholar]
- Kc, P.; Hong, Y.; Zhang, G. Cardiac tissue-derived extracellular matrix scaffolds for myocardial repair: Advantages and challenges. Regen. Biomater. 2019, 6, 185–199. [Google Scholar] [CrossRef] [PubMed]
- Rajabi, S.; Aghdami, N.; Varzideh, F.; Parchehbaf-Kashani, M.; Nobakht Lahrood, F. Decellularized muscle-derived hydrogels support in vitro cardiac microtissue fabrication. J. Biomed. Mater. Res. B Appl. Biomater. 2020. [Google Scholar] [CrossRef] [PubMed]
- Masumoto, H.; Ikuno, T.; Takeda, M.; Fukushima, H.; Marui, A.; Katayama, S.; Shimizu, T.; Ikeda, T.; Okano, T.; Sakata, R.; et al. Human iPS cell-engineered cardiac tissue sheets with cardiomyocytes and vascular cells for cardiac regeneration. Sci. Rep. 2015, 4, 6716. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Perea-Gil, I.; Gálvez-Montón, C.; Prat-Vidal, C.; Jorba, I.; Segú-Vergés, C.; Roura, S.; Soler-Botija, C.; Iborra-Egea, O.; Revuelta-López, E.; Fernández, M.A.; et al. Head-to-head comparison of two engineered cardiac grafts for myocardial repair: From scaffold characterization to pre-clinical testing. Sci. Rep. 2018, 8, 6708. [Google Scholar] [CrossRef] [Green Version]
- Hamdi, H.; Planat-Benard, V.; Bel, A.; Neamatalla, H.; Saccenti, L.; Calderon, D.; Bellamy, V.; Bon, M.; Perrier, M.-C.; Mandet, C.; et al. Long-Term Functional Benefits of Epicardial Patches as Cell Carriers. Cell Transplant. 2014, 23, 87–96. [Google Scholar] [CrossRef]
- Di Mauro, M.; Gallina, S.; D’Amico, M.A.; Izzicupo, P.; Lanuti, P.; Bascelli, A.; Di Fonso, A.; Bartoloni, G.; Calafiore, A.M.; Di Baldassarre, A. Functional mitral regurgitation. Int. J. Cardiol. 2013, 163, 242–248. [Google Scholar] [CrossRef]
- Parveen, S.; Singh, S.P.; Panicker, M.M.; Gupta, P.K. Amniotic membrane as novel scaffold for human iPSC-derived cardiomyogenesis. Vitro Cell. Dev. Biol. Anim. 2019, 55, 272–284. [Google Scholar] [CrossRef]
- Liguori, G.R.; Liguori, T.T.A.; de Moraes, S.R.; Sinkunas, V.; Terlizzi, V.; van Dongen, J.A.; Sharma, P.K.; Moreira, L.F.P.; Harmsen, M.C. Molecular and Biomechanical Clues From Cardiac Tissue Decellularized Extracellular Matrix Drive Stromal Cell Plasticity. Front. Bioeng. Biotechnol. 2020, 8, 520. [Google Scholar] [CrossRef]
- Woo, J.S.; Fishbein, M.C.; Reemtsen, B. Histologic examination of decellularized porcine intestinal submucosa extracellular matrix (CorMatrix) in pediatric congenital heart surgery. Cardiovasc. Pathol. 2016, 25, 12–17. [Google Scholar] [CrossRef]
- Zaidi, A.H.; Nathan, M.; Emani, S.; Baird, C.; del Nido, P.J.; Gauvreau, K.; Harris, M.; Sanders, S.P.; Padera, R.F. Preliminary experience with porcine intestinal submucosa (CorMatrix) for valve reconstruction in congenital heart disease: Histologic evaluation of explanted valves. J. Thorac. Cardiovasc. Surg. 2014, 148, 2216–2225.e1. [Google Scholar] [CrossRef] [Green Version]
- Stelly, M.; Stelly, T.C. Histology of CorMatrix Bioscaffold 5 Years after Pericardial Closure. Ann. Thorac. Surg. 2013, 96, e127–e129. [Google Scholar] [CrossRef] [PubMed]
- Di Mauro, M.; Ghinassi, B.; Di Baldassarre, A. Commentary: I fix what’s broken - inclusive the heart. J. Thorac. Cardiovasc. Surg. 2020, S0022522320317669. [Google Scholar] [CrossRef]
- Dohmen, P.M.; da Costa, F.; Lopes, S.V.; Vilani, R.; Bloch, O.; Konertz, W. Successful implantation of a decellularized equine pericardial patch into the systemic circulation. Med. Sci. Monit. Basic Res. 2014, 20, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Nelson, D.M.; Ma, Z.; Fujimoto, K.L.; Hashizume, R.; Wagner, W.R. Intra-myocardial biomaterial injection therapy in the treatment of heart failure: Materials, outcomes and challenges. Acta Biomater. 2011, 7, 1–15. [Google Scholar] [CrossRef] [Green Version]
- van der Velden, J.; Stienen, G.J.M. Cardiac Disorders and Pathophysiology of Sarcomeric Proteins. Physiol. Rev. 2019, 99, 381–426. [Google Scholar] [CrossRef]
- Levenberg, S.; Huang, N.F.; Lavik, E.; Rogers, A.B.; Itskovitz-Eldor, J.; Langer, R. Differentiation of human embryonic stem cells on three-dimensional polymer scaffolds. Proc. Natl. Acad. Sci. USA 2003, 100, 12741–12746. [Google Scholar] [CrossRef] [Green Version]
- Calafiore, A.M.; Lorusso, R.; Kheirallah, H.; Alsaied, M.M.; Alfonso, J.J.; Di Baldassarre, A.; Gallina, S.; Gaudino, M.; Di Mauro, M. Late tricuspid regurgitation and right ventricular remodeling after tricuspid annuloplasty. J. Card. Surg. 2020, 35, 1891–1900. [Google Scholar] [CrossRef]
- Lian, X.; Hsiao, C.; Wilson, G.; Zhu, K.; Hazeltine, L.B.; Azarin, S.M.; Raval, K.K.; Zhang, J.; Kamp, T.J.; Palecek, S.P. Cozzarelli Prize Winner: Robust cardiomyocyte differentiation from human pluripotent stem cells via temporal modulation of canonical Wnt signaling. Proc. Natl. Acad. Sci. USA 2012, 109, E1848–E1857. [Google Scholar] [CrossRef] [Green Version]
- Burridge, P.W.; Matsa, E.; Shukla, P.; Lin, Z.C.; Churko, J.M.; Ebert, A.D.; Lan, F.; Diecke, S.; Huber, B.; Mordwinkin, N.M.; et al. Chemically defined generation of human cardiomyocytes. Nat. Methods 2014, 11, 855–860. [Google Scholar] [CrossRef] [Green Version]
- Yoshida, Y.; Yamanaka, S. Induced Pluripotent Stem Cells 10 Years Later: For Cardiac Applications. Circ. Res. 2017, 120, 1958–1968. [Google Scholar] [CrossRef]
- Kanda, Y.; Yamazaki, D.; Osada, T.; Yoshinaga, T.; Sawada, K. Development of torsadogenic risk assessment using human induced pluripotent stem cell-derived cardiomyocytes: Japan iPS Cardiac Safety Assessment (JiCSA) update. J. Pharmacol. Sci. 2018, 138, 233–239. [Google Scholar] [CrossRef]
- Izzicupo, P.; Ghinassi, B.; D’Amico, M.A.; Di Blasio, A.; Gesi, M.; Napolitano, G.; Gallina, S.; Di Baldassarre, A. Effects of ACE I/D Polymorphism and Aerobic Training on the Immune–Endocrine Network and Cardiovascular Parameters of Postmenopausal Women. J. Clin. Endocrinol. Metab. 2013, 98, 4187–4194. [Google Scholar] [CrossRef] [Green Version]
- Guan, X.; Delo, D.M.; Atala, A.; Soker, S. In vitro cardiomyogenic potential of human amniotic fluid stem cells. J. Tissue Eng. Regen. Med. 2011, 5, 220–228. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cananzi, M.; De Coppi, P. CD117 + amniotic fluid stem cells: State of the art and future perspectives. Organogenesis 2012, 8, 77–88. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Laflamme, M.A.; Chen, K.Y.; Naumova, A.V.; Muskheli, V.; Fugate, J.A.; Dupras, S.K.; Reinecke, H.; Xu, C.; Hassanipour, M.; Police, S.; et al. Cardiomyocytes derived from human embryonic stem cells in pro-survival factors enhance function of infarcted rat hearts. Nat. Biotechnol. 2007, 25, 1015–1024. [Google Scholar] [CrossRef] [PubMed]
- Albini, A. Extracellular Matrix Invasion in Metastases and Angiogenesis: Commentary on the Matrigel “Chemoinvasion Assay”. Cancer Res. 2016, 76, 4595–4597. [Google Scholar] [CrossRef] [Green Version]
- Arumugam, R.; Srinadhu, E.S.; Subramanian, B.; Nallani, S. β-PVDF based electrospun nanofibers—A promising material for developing cardiac patches. Med. Hypotheses 2019, 122, 31–34. [Google Scholar] [CrossRef]
- Rajabi, S.; Pahlavan, S.; Ashtiani, M.K.; Ansari, H.; Abbasalizadeh, S.; Sayahpour, F.A.; Varzideh, F.; Kostin, S.; Aghdami, N.; Braun, T.; et al. Human embryonic stem cell-derived cardiovascular progenitor cells efficiently colonize in bFGF-tethered natural matrix to construct contracting humanized rat hearts. Biomaterials 2018, 154, 99–112. [Google Scholar] [CrossRef]
- Pattar, S.S.; Fatehi Hassanabad, A.; Fedak, P.W.M. Acellular Extracellular Matrix Bioscaffolds for Cardiac Repair and Regeneration. Front. Cell Dev. Biol. 2019, 7, 63. [Google Scholar] [CrossRef]
- Pfannkuche, K.; Neuss, S.; Pillekamp, F.; Frenzel, L.P.; Attia, W.; Hannes, T.; Salber, J.; Hoss, M.; Zenke, M.; Fleischmann, B.K.; et al. Fibroblasts Facilitate the Engraftment of Embryonic Stem Cell-Derived Cardiomyocytes on Three-Dimensional Collagen Matrices and Aggregation in Hanging Drops. Stem Cells Dev. 2010, 19, 1589–1599. [Google Scholar] [CrossRef] [Green Version]
- van Deel, E.D.; Najafi, A.; Fontoura, D.; Valent, E.; Goebel, M.; Kardux, K.; Falcão-Pires, I.; van der Velden, J. In vitro model to study the effects of matrix stiffening on Ca 2+ handling and myofilament function in isolated adult rat cardiomyocytes: Matrix stiffening alters cardiomyocyte function. J. Physiol. 2017, 595, 4597–4610. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, Y.-K.; Ng, K.-M.; Lai, W.-H.; Chan, Y.-C.; Lau, Y.-M.; Lian, Q.; Tse, H.-F.; Siu, C.-W. Calcium Homeostasis in Human Induced Pluripotent Stem Cell-Derived Cardiomyocytes. Stem Cell Rev. Rep. 2011, 7, 976–986. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, G.; Herron, T.J.; Di Bernardo, J.; Walker, K.A.; O’Shea, K.S.; Kunisaki, S.M. Human Cardiomyocytes Prior to Birth by Integration-Free Reprogramming of Amniotic Fluid Cells: Amniotic Cardiomyocytes. STEM CELLS Transl. Med. 2016, 5, 1595–1606. [Google Scholar] [CrossRef]
- Moschidou, D.; Mukherjee, S.; Blundell, M.P.; Jones, G.N.; Atala, A.J.; Thrasher, A.J.; Fisk, N.M.; De Coppi, P.; Guillot, P.V. Human Mid-Trimester Amniotic Fluid Stem Cells Cultured Under Embryonic Stem Cell Conditions with Valproic Acid Acquire Pluripotent Characteristics. Stem Cells Dev. 2013, 22, 444–458. [Google Scholar] [CrossRef]
- Yang, X.; Pabon, L.; Murry, C.E. Engineering Adolescence: Maturation of Human Pluripotent Stem Cell–Derived Cardiomyocytes. Circ. Res. 2014, 114, 511–523. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Izzicupo, P.; Di Valerio, V.; D’Amico, M.A.; Di Mauro, M.; Pennelli, A.; Falone, S.; Alberti, G.; Amicarelli, F.; Miscia, S.; Gallina, S.; et al. Nad(P)H Oxidase and Pro-Inflammatory Response during Maximal Exercise: Role of C242T Polymorphism of the P22PHOX Subunit. Int. J. Immunopathol. Pharmacol. 2010, 23, 203–211. [Google Scholar] [CrossRef]
- Hirata, M.; Yamaoka, T. Effect of stem cell niche elasticity/ECM protein on the self-beating cardiomyocyte differentiation of induced pluripotent stem (iPS) cells at different stages. Acta Biomater. 2018, 65, 44–52. [Google Scholar] [CrossRef]
- Amabile, G.; Welner, R.S.; Nombela-Arrieta, C.; D’Alise, A.M.; Di Ruscio, A.; Ebralidze, A.K.; Kraytsberg, Y.; Ye, M.; Kocher, O.; Neuberg, D.S.; et al. In vivo generation of transplantable human hematopoietic cells from induced pluripotent stem cells. Blood 2013, 121, 1255–1264. [Google Scholar] [CrossRef]
- Prinz, R.D.; Willis, C.M.; van Kuppevelt, T.H.; Klüppel, M. Biphasic Role of Chondroitin Sulfate in Cardiac Differentiation of Embryonic Stem Cells through Inhibition of Wnt/β-Catenin Signaling. PLoS ONE 2014, 9, e92381. [Google Scholar] [CrossRef] [Green Version]
- D’amico, M.A.; Ghinassi, B.; Izzicupo, P.; Di Ruscio, A.; Di Baldassarre, A. IL-6 Activates PI3K and PKCζ Signaling and Determines Cardiac Differentiation in Rat Embryonic H9c2 Cells: IL-6 AND CARDIAC DIFFERENTIATION OF H9c2 CELLS. J. Cell. Physiol. 2016, 231, 576–586. [Google Scholar] [CrossRef]
- Rapino, M.; Di Valerio, V.; Zara, S.; Gallorini, M.; Marconi, G.D.; Sancilio, S.; Marsich, E.; Ghinassi, B.; di Giacomo, V.; Cataldi, A. Chitlac-coated Thermosets Enhance Osteogenesis and Angiogenesis in a Co-culture of Dental Pulp Stem Cells and Endothelial Cells. Nanomaterials 2019, 9, 928. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martelli, F.; Ghinassi, B.; Lorenzini, R.; Vannucchi, A.M.; Rana, R.A.; Nishikawa, M.; Partamian, S.; Migliaccio, G.; Migliaccio, A.R. Thrombopoietin Inhibits Murine Mast Cell Differentiation. Stem Cells 2008, 26, 912–919. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ghinassi, B.; Ferro, L.; Masiello, F.; Tirelli, V.; Sanchez, M.; Migliaccio, G.; Whitsett, C.; Kachala, S.; Riviere, I.; Sadelain, M.; et al. Recovery and Biodistribution of Ex Vivo Expanded Human Erythroblasts Injected into NOD/SCID/IL2R γ null mice. Stem Cells Int. 2011, 2011, 1–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Migliaccio, A.R.; Martelli, F.; Verrucci, M.; Sanchez, M.; Valeri, M.; Migliaccio, G.; Vannucchi, A.M.; Zingariello, M.; Di Baldassarre, A.; Ghinassi, B.; et al. Gata1 expression driven by the alternative HS2 enhancer in the spleen rescues the hematopoietic failure induced by the hypomorphic Gata1low mutation. Blood 2009, 114, 2107–2120. [Google Scholar] [CrossRef] [PubMed] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| cTnT | α-MHC | α-SA | CACNA1C | SERCA2 | |
|---|---|---|---|---|---|
| hiPSCs | |||||
| Prol | 1.0 ± 0.5 | 1.2 ± 0.5 | 1.1 ± 1.1 | 6.5 ± 5.8 | 26.6 ± 9.5 |
| Diff Matrigel® | 83.5 ± 6.6 * | 83.4 ± 3.1 * | 95.3 ± 3.1 * | 79.9 ± 6.8 * | 93.4 ± 8.6 * |
| Diff CorTM PATCH | 88.1 ± 11.1 * | 69.2 ± 9.9 * | 88.5 ± 7.3 * | 76.2 ± 3.1 * | 83.6 ± 7.3 * |
| Diff Matrix PatchTM | 86.2 ± 5.6 * | 81.4 ± 7.5 * | 81.5 ± 6.4 *& | 75.9 ± 6.2 * | 86.1 ± 7.2 * |
| CardiopoieticAF | |||||
| Prol | 2.2 ± 2.1 | 1.1 ± 0.5 | 4.1 ± 2.1 | 12.5 ± 8.8 | 45.1 ± 12.5 |
| Diff Matrigel® | 85.5 ± 5.6 * | 88.4 ± 8.1 * | 89.3 ± 7.9 * | 88.9 ± 5.9 * | 83.5 ± 6.8 * |
| Diff CorTM PATCH | 78.6 ± 9.6 * | 79.5 ± 6.9 * | 78.5 ± 7.4 * | 96.2 ± 4.1 * | 84.5 ± 9.3 * |
| Diff Matrix PatchTM | 75.2 ± 6.6 * | 74.4 ± 8.8 * | 66.5 ± 9.4 *& | 92.4 ± 7.2 * | 83.5 ± 6.1 * |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gaggi, G.; Di Credico, A.; Izzicupo, P.; Sancilio, S.; Di Mauro, M.; Iannetti, G.; Dolci, S.; Amabile, G.; Di Baldassarre, A.; Ghinassi, B. Decellularized Extracellular Matrices and Cardiac Differentiation: Study on Human Amniotic Fluid-Stem Cells. Int. J. Mol. Sci. 2020, 21, 6317. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21176317
Gaggi G, Di Credico A, Izzicupo P, Sancilio S, Di Mauro M, Iannetti G, Dolci S, Amabile G, Di Baldassarre A, Ghinassi B. Decellularized Extracellular Matrices and Cardiac Differentiation: Study on Human Amniotic Fluid-Stem Cells. International Journal of Molecular Sciences. 2020; 21(17):6317. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21176317
Chicago/Turabian StyleGaggi, Giulia, Andrea Di Credico, Pascal Izzicupo, Silvia Sancilio, Michele Di Mauro, Giovanni Iannetti, Susanna Dolci, Giovanni Amabile, Angela Di Baldassarre, and Barbara Ghinassi. 2020. "Decellularized Extracellular Matrices and Cardiac Differentiation: Study on Human Amniotic Fluid-Stem Cells" International Journal of Molecular Sciences 21, no. 17: 6317. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21176317