Molecular Profile of Barrett’s Esophagus and Gastroesophageal Reflux Disease in the Development of Translational Physiological and Pharmacological Studies


 , , , , ,
, , , , ,  and
and
Abstract
:1. Introduction
2. Results
2.1. Analysis of BE Expression Profile for Selected Genes in Human Biopsies Based on GSE Datasets
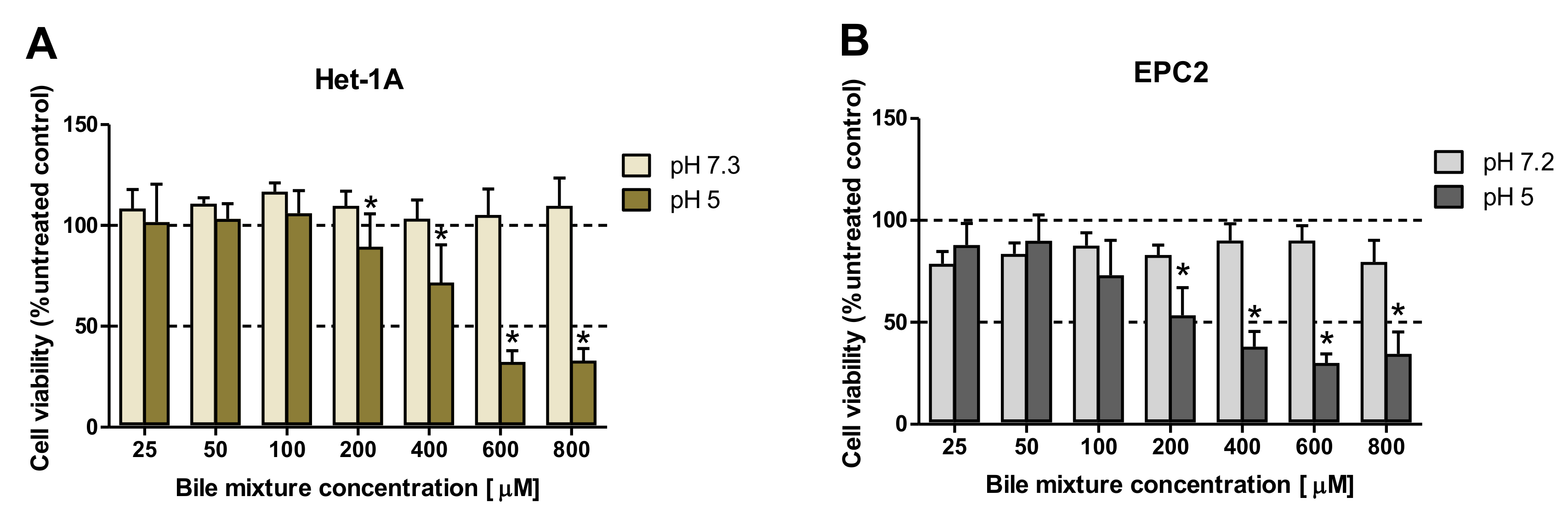
2.2. The Effect of Exposition to Various Bile Mixture (BM) Concentrations and pH Values on the Viability of Esophageal Epithelial Cell Lines
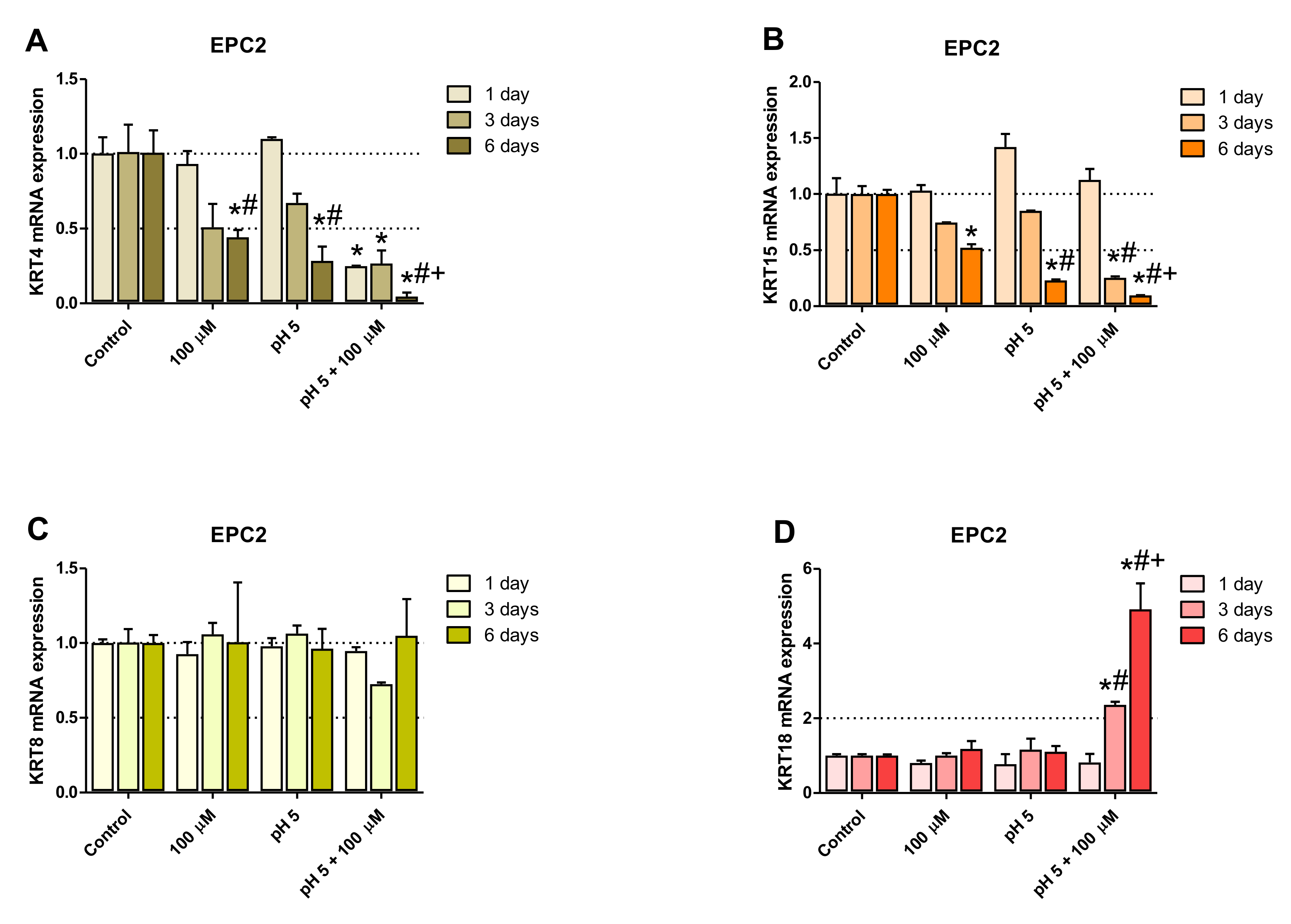
2.3. Optimization of the Experimental Procedure Duration
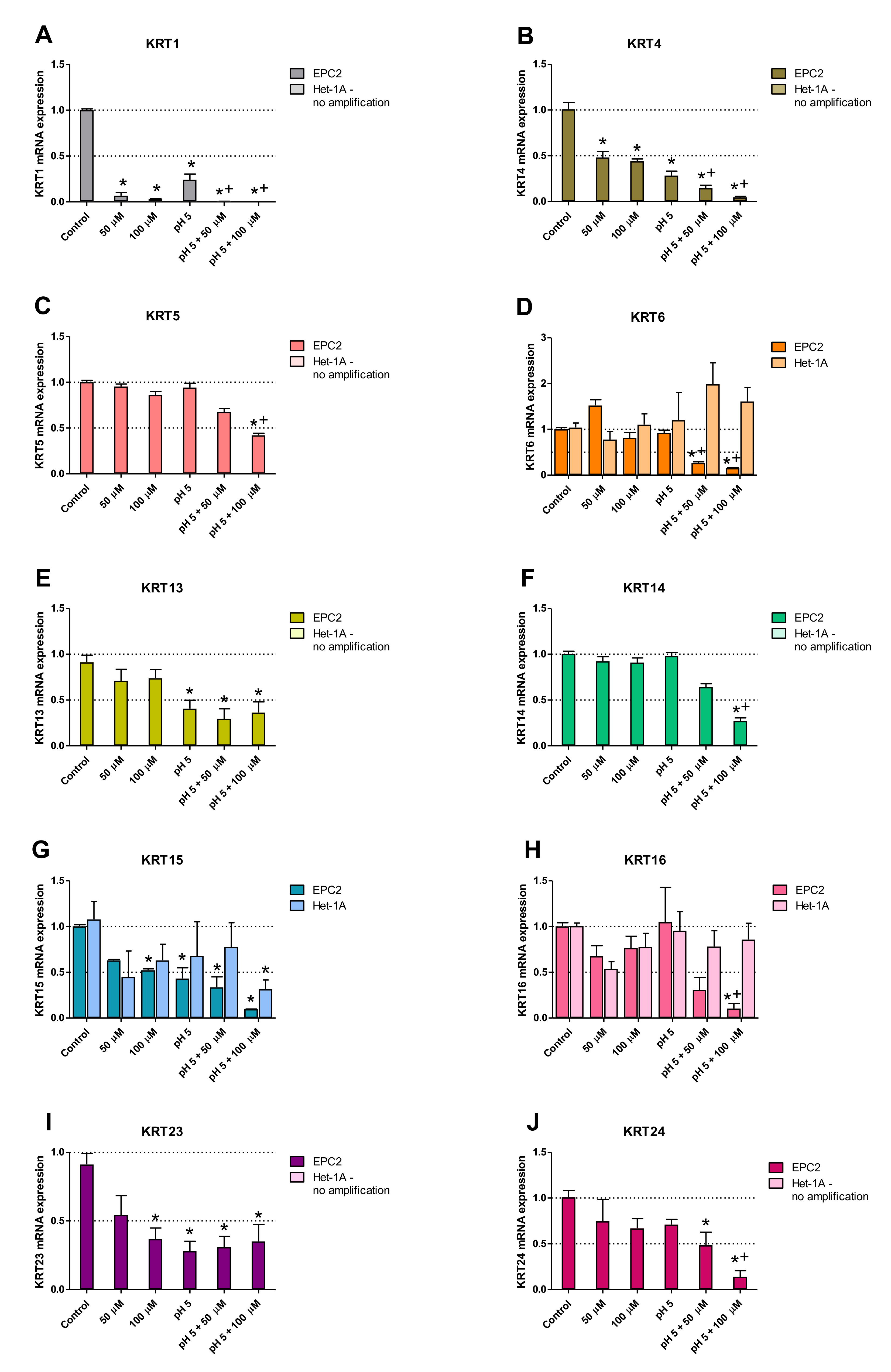
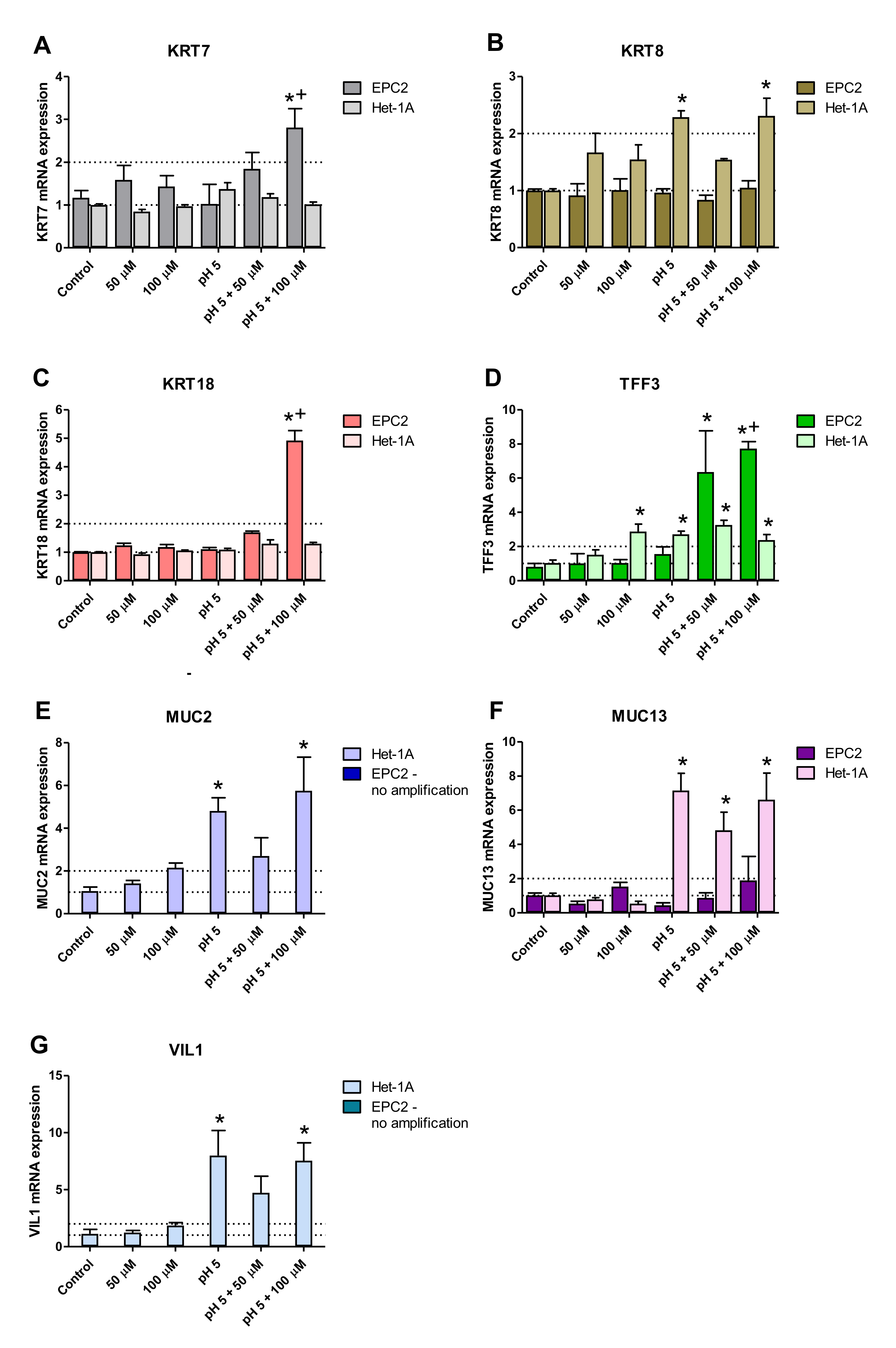
2.4. Squamous and Columnar Epithelium-Specific Genes Expression in In Vitro Model
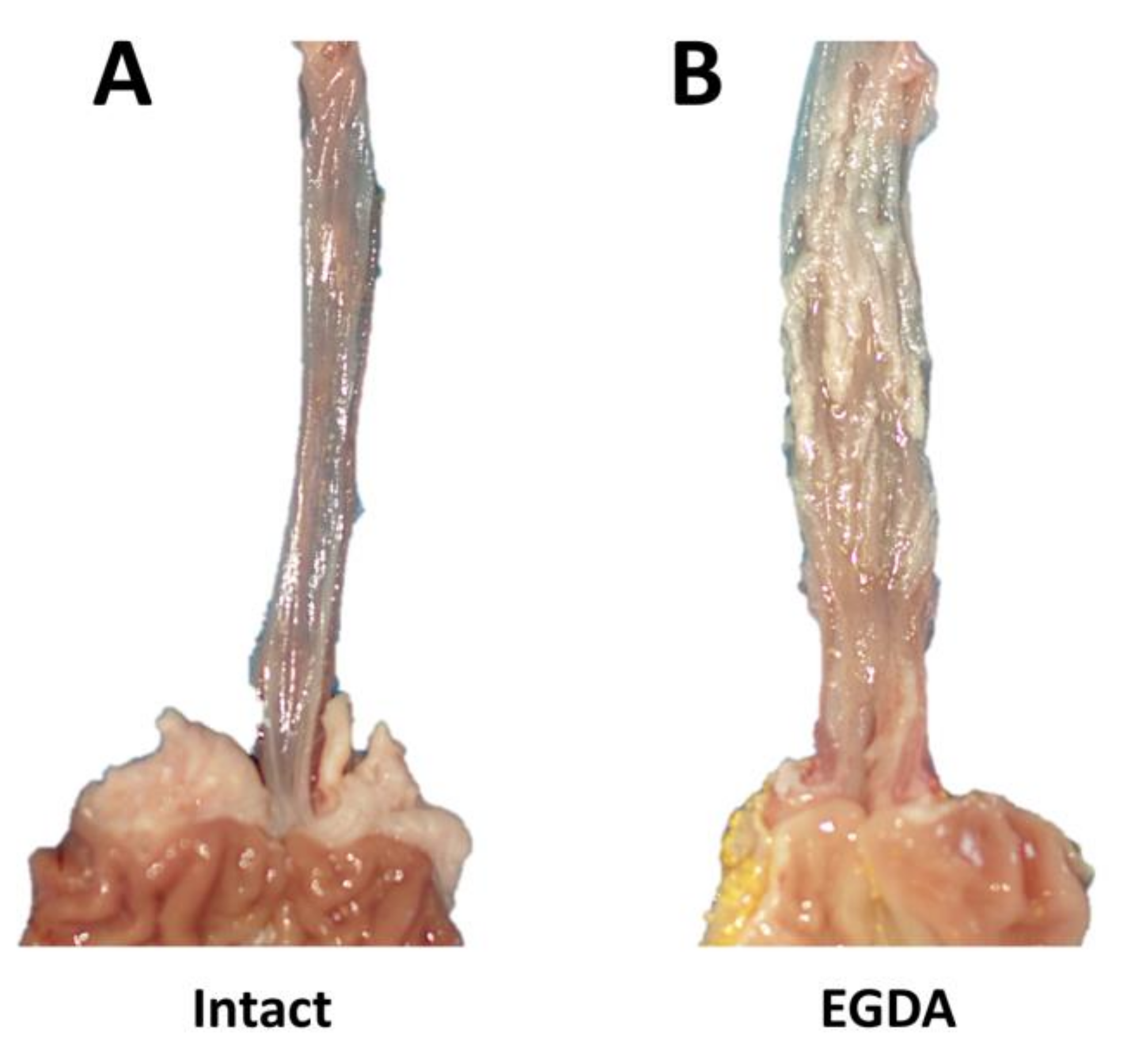
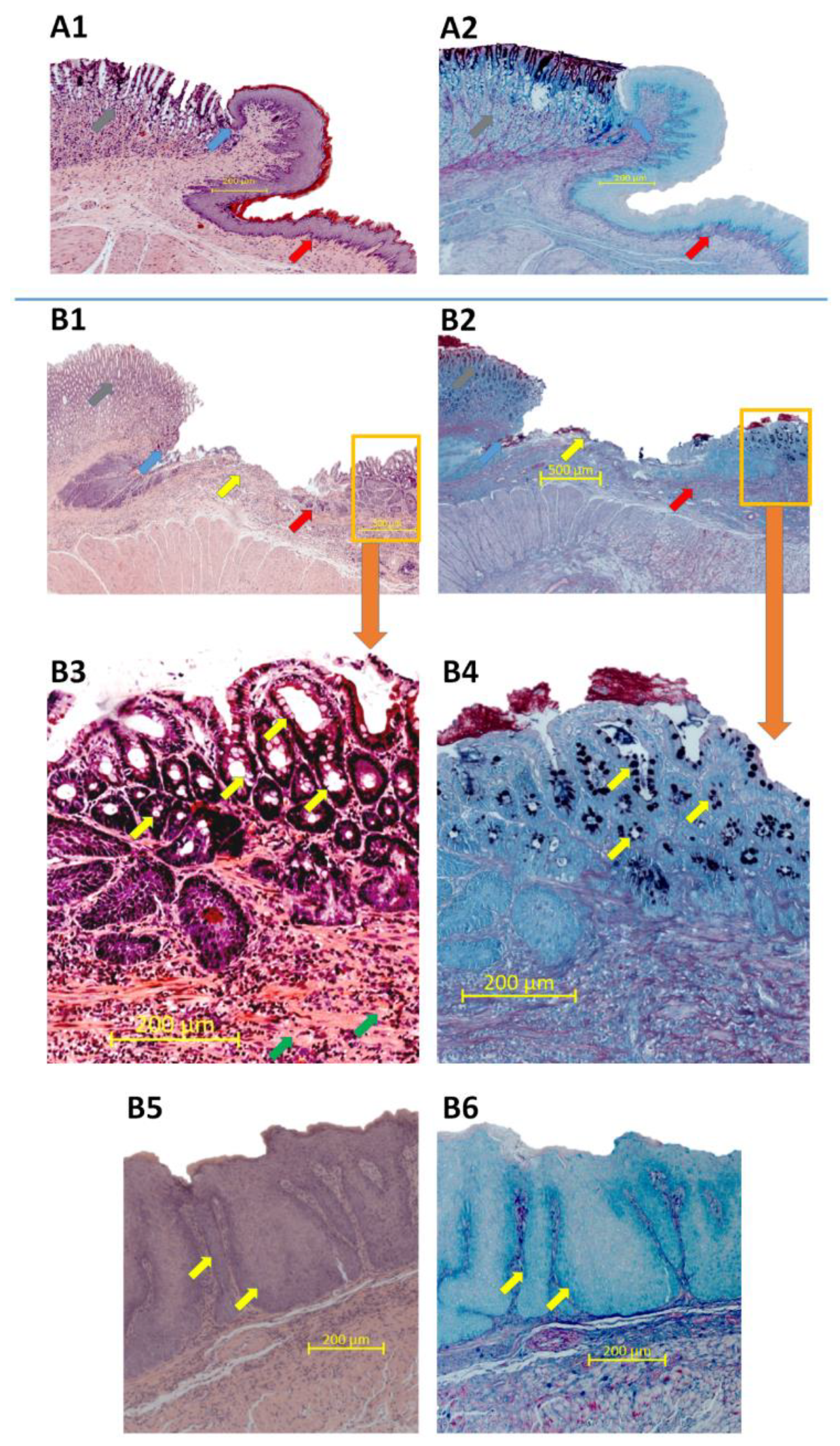
2.5. Morphology of Esophageal Mucosa, Gastroesophageal Junction (GEJ), and Gastric Cardia in Rats with Esophagogastroduodenal Anastomosis (EGDA)
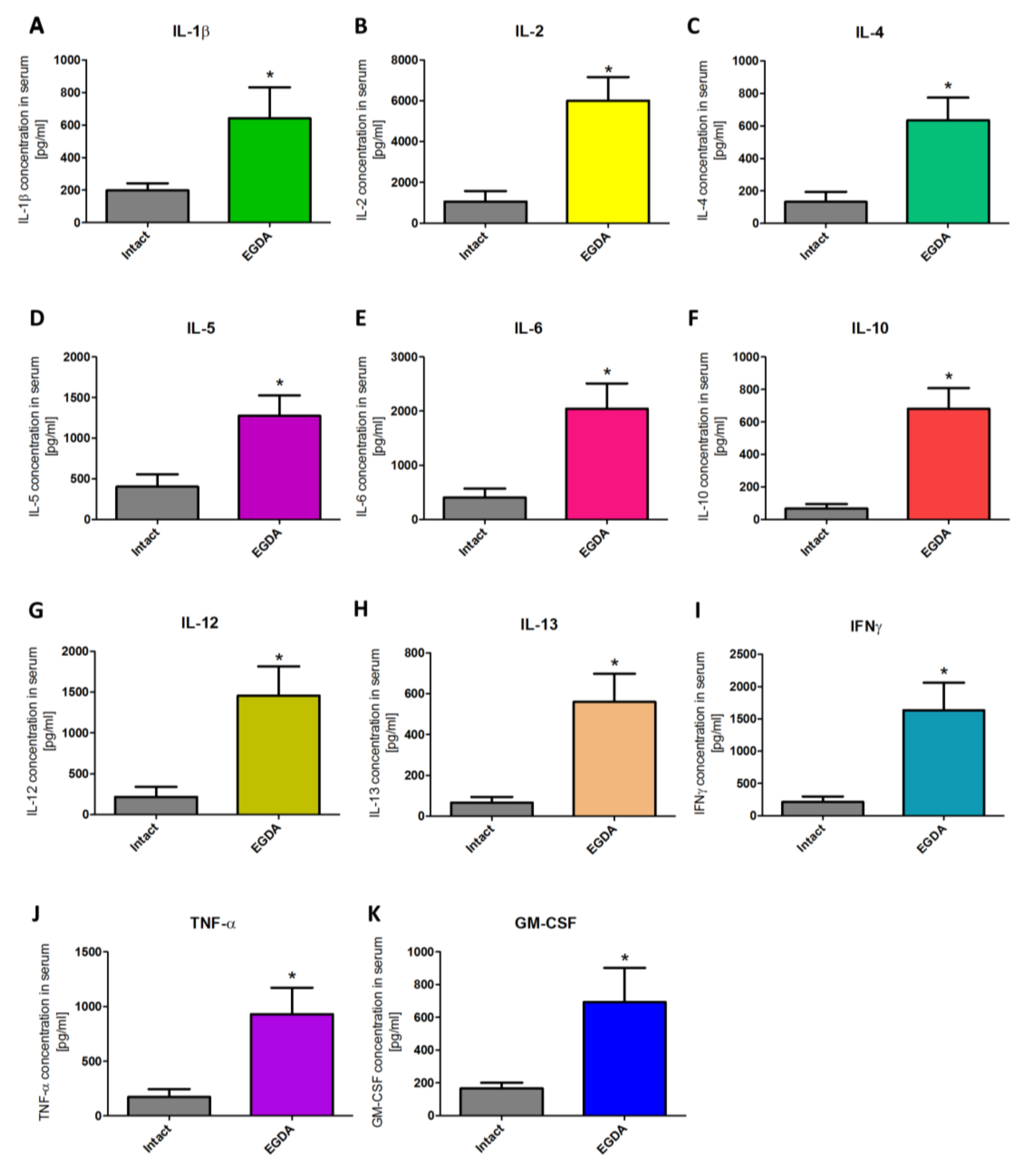
2.6. Alterations in Serum Content of Pro- and Anti-Inflammatory Cytokines in Rats with EGDA
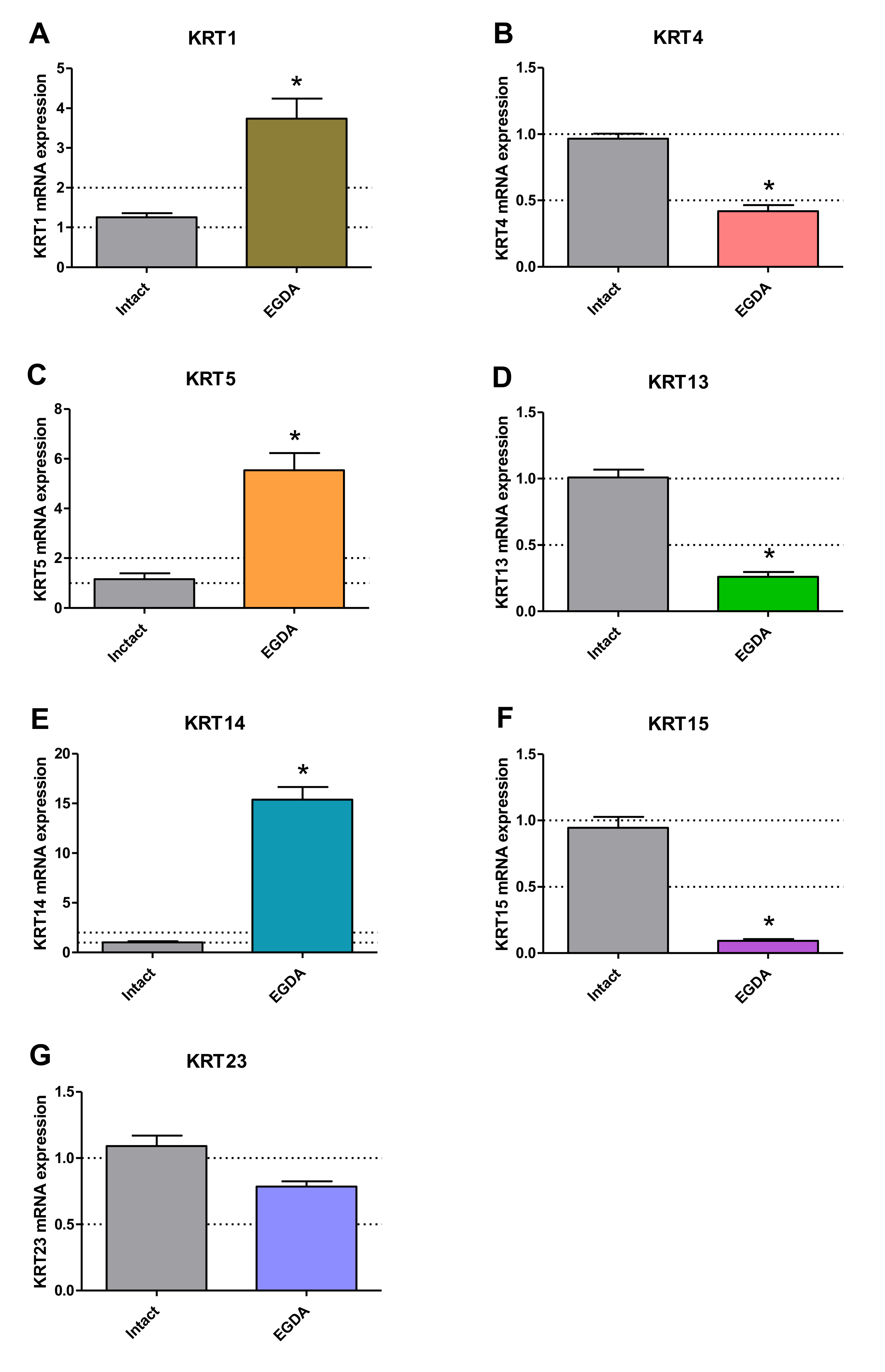
2.7. Squamous and Columnar Epithelium-Specific mRNA Expression in Esophageal Mucosa of Rats with EGDA
3. Discussion
4. Material and Methods
4.1. Analysis of BE Expression Profile for Selected Genes in Human Biopsies Based on GSE Datasets
4.2. Cell Cultures
4.3. Acid/Bile Mixture (BM) Treatment
4.4. Cell-Viability Assays
4.5. Analysis of mRNA Expression by Real-Time Polymerase Chain Reaction (PCR)
4.6. Animal Model of BE
- hyperplasia of squamous epithelium,
- fibrosis of lamina propria,
- esophagitis: 1—thickening of squamous epithelium with basal cell layer occupying up to 30% of its height; elongation of connective tissue papillae, 2—regeneration layer occupying 50% of the epithelium thickness; hyperemia and scanty inflammatory infiltrate are present in connective tissue papillae, 3—expansion of the regeneration zone to 75% of the epithelial height; moderate inflammatory infiltrate in connective tissue papillae, 4—ulceration or massive inflammatory infiltrate,
- Barrett’s-like lesion with the presence of goblet cells.
4.7. Determination of Serum Content of Pro- and Anti-Inflammatory Factors by Luminex Microbeads Fluorescent Assays
4.8. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Bus, P.; Siersema, P.D.; van Baal, J.W. Cell culture models for studying the development of Barrett’s esophagus: A systematic review. Cell. Oncol. 2012, 35, 149–161. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kosoff, R.E.; Gardiner, K.L.; Merlo, L.M.; Pavlov, K.; Rustgi, A.K.; Maley, C.C. Development and characterization of an organotypic model of Barrett’s esophagus. J. Cell. Physiol. 2012, 227, 2654–2659. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Naini, B.V.; Souza, R.F.; Odze, R.D. Barrett’s esophagus: A comprehensive and contemporary review for pathologists. Am. J. Surg. Pathol. 2016, 40, e45–e66. [Google Scholar] [CrossRef] [PubMed]
- Su, Z.; Gay, L.J.; Strange, A.; Palles, C.; Band, G.; Whiteman, D.C.; Lescai, F.; Langford, C.; Nanji, M.; Edkins, S.; et al. Common variants at the MHC locus and at chromosome 16q24.1 predispose to Barrett’s esophagus. Nat. Genet. 2012, 44, 1131–1136. [Google Scholar] [CrossRef]
- Jankowski, J.A.; De Caestecker, J.; Love, S.B.; Reilly, G.; Watson, P.; Sanders, S.; Ang, Y.; Morris, D.; Bhandari, P.; Brooks, C.; et al. Esomeprazole and aspirin in Barrett’s oesophagus (AspECT): A randomised factorial trial. Lancet 2018, 392, 400–408. [Google Scholar] [CrossRef] [Green Version]
- Savarino, E.; Zentilin, P.; Frazzoni, M.; Cuoco, D.L.; Pohl, D.; Dulbecco, P.; Marabotto, E.; Sammito, G.; Gemignani, L.; Tutuian, R.; et al. Characteristics of gastro-esophageal reflux episodes in Barrett’s esophagus, erosive esophagitis and healthy volunteers. Neurogastroenterol. Motil. Off. J. Eur. Gastrointest. Motil. Soc. 2010, 22, 1061-e280. [Google Scholar] [CrossRef]
- Tolone, S.; Limongelli, P.; Romano, M.; Federico, A.; Docimo, G.; Ruggiero, R.; Brusciano, L.; Del Genio, G.; Docimo, L. The patterns of reflux can affect regression of non-dysplastic and low-grade dysplastic Barrett’s esophagus after medical and surgical treatment: A prospective case-control study. Surg. Endosc. 2015, 29, 648–657. [Google Scholar] [CrossRef]
- Stairs, D.B.; Nakagawa, H.; Klein-Szanto, A.; Mitchell, S.D.; Silberg, D.G.; Tobias, J.W.; Lynch, J.P.; Rustgi, A.K. Cdx1 and c-Myc foster the initiation of transdifferentiation of the normal esophageal squamous epithelium toward Barrett’s esophagus. PLoS ONE 2008, 3, e3534. [Google Scholar] [CrossRef] [Green Version]
- di Pietro, M.; Lao-Sirieix, P.; Boyle, S.; Cassidy, A.; Castillo, D.; Saadi, A.; Eskeland, R.; Fitzgerald, R.C. Evidence for a functional role of epigenetically regulated midcluster HOXB genes in the development of Barrett esophagus. Proc. Natl. Acad. Sci. USA 2012, 109, 9077–9082. [Google Scholar] [CrossRef] [Green Version]
- Van de Bovenkamp, J.H.; Korteland-Van Male, A.M.; Warson, C.; Büller, H.A.; Einerhand, A.W.; Ectors, N.L.; Dekker, J. Gastric-Type mucin and TFF-peptide expression in Barrett’s oesophagus is disturbed during increased expression of MUC2. Histopathology 2003, 42, 555–565. [Google Scholar] [CrossRef]
- Poehlmann, A.; Kuester, D.; Malfertheiner, P.; Guenther, T.; Roessner, A. Inflammation and Barrett’s carcinogenesis. Pathol. Res. Pract. 2012, 208, 269–280. [Google Scholar] [CrossRef] [PubMed]
- Squier, C.A.; Kremer, M.J. Biology of oral mucosa and esophagus. J. Natl. Cancer Inst. Monogr. 2001, 2001, 7–15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Flejou, J.F. Barrett’s oesophagus: From metaplasia to dysplasia and cancer. Gut 2005, 54 (Suppl. S1), i6–i12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rees, J.R.; Lao-Sirieix, P.; Wong, A.; Fitzgerald, R.C. Treatment for Barrett’s oesophagus. Cochrane Database Syst. Rev. 2010, CD004060. [Google Scholar] [CrossRef]
- Majka, J.; Rembiasz, K.; Migaczewski, M.; Budzynski, A.; Ptak-Belowska, A.; Pabianczyk, R.; Urbanczyk, K.; Zub-Pokrowiecka, A.; Matlok, M.; Brzozowski, T. Cyclooxygenase-2 (Cox-2) is the key event in pathophysiology of Barrett’s esophagus. lesson from experimental animal model and human subjects. J. Physiol. Pharmacol. 2010, 61, 409–418. [Google Scholar]
- Lagorce, C.; Paraf, F.; Vidaud, D.; Couvelard, A.; Wendum, D.; Martin, A.; Fléjou, J.F. Cyclooxygenase-2 is expressed frequently and early in Barrett’s oesophagus and associated adenocarcinoma. Histopathology 2003, 42, 457–465. [Google Scholar] [CrossRef]
- Menke, V.; Pot, R.G.; Moons, L.M.; van Zoest, K.P.; Hansen, B.; van Dekken, H.; Siersema, P.D.; Kusters, J.G.; Kuipers, E.J. Functional single-nucleotide polymorphism of epidermal growth factor is associated with the development of Barrett’s esophagus and esophageal adenocarcinoma. J. Hum. Genet. 2012, 57, 26–32. [Google Scholar] [CrossRef]
- Jankowski, J.; Coghill, G.; Tregaskis, B.; Hopwood, D.; Wormsley, K.G. Epidermal growth factor in the oesophagus. Gut 1992, 33, 1448–1453. [Google Scholar] [CrossRef]
- Majka, J.; Wierdak, M.; Szlachcic, A.; Magierowski, M.; Targosz, A.; Urbanczyk, K.; Krzysiek-Maczka, G.; Ptak-Belowska, A.; Bakalarz, D.; Magierowska, K.; et al. Interaction of epidermal growth factor with COX-2 products and peroxisome proliferator-activated receptor gamma system in experimental rat Barrett’s esophagus. Am. J. Physiol. Gastrointest. Liver Physiol. 2020, 318, G375–G389. [Google Scholar] [CrossRef]
- Vona-Davis, L.; Frankenberry, K.; Cunningham, C.; Riggs, D.R.; Jackson, B.J.; Szwerc, M.F.; McFadden, D.W. MAPK and PI3K inhibition reduces proliferation of Barrett’s adenocarcinoma in vitro. J. Surg. Res. 2005, 127, 53–58. [Google Scholar] [CrossRef]
- Sarosi, G.A., Jr.; Jaiswal, K.; Herndon, E.; Lopez-Guzman, C.; Spechler, S.J.; Souza, R.F. Acid increases MAPK-mediated proliferation in Barrett’s esophageal adenocarcinoma cells via intracellular acidification through a Cl-/HCO3- exchanger. Am. J. Physiol. Gastrointest. Liver Physiol. 2005, 289, G991–G997. [Google Scholar] [CrossRef] [Green Version]
- Han, Y.M.; Park, J.M.; Kangwan, N.; Jeong, M.; Lee, S.; Cho, J.Y.; Ko, W.J.; Hahm, K.B. Role of proton pump inhibitors in preventing hypergastrinemia-associated carcinogenesis and in antagonizing the trophic effect of gastrin. J. Physiol. Pharmacol. 2015, 66, 159–167. [Google Scholar] [PubMed]
- Ahrens, T.D.; Lutz, L.; Lassmann, S.; Werner, M. Turning skyscrapers into town houses: Insights into Barrett’s esophagus. Pathobiol. J. Immunopathol. Mol. Cell. Biol. 2017, 84, 87–98. [Google Scholar] [CrossRef] [PubMed]
- Kimchi, E.T.; Posner, M.C.; Park, J.O.; Darga, T.E.; Kocherginsky, M.; Karrison, T.; Hart, J.; Smith, K.D.; Mezhir, J.J.; Weichselbaum, R.R.; et al. Progression of Barrett’s metaplasia to adenocarcinoma is associated with the suppression of the transcriptional programs of epidermal differentiation. Cancer Res. 2005, 65, 3146–3154. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Buskens, C.J.; Hulscher, J.B.; van Gulik, T.M.; Ten Kate, F.J.; van Lanschot, J.J. Histopathologic evaluation of an animal model for Barrett’s esophagus and adenocarcinoma of the distal esophagus. J. Surg. Res. 2006, 135, 337–344. [Google Scholar] [CrossRef] [PubMed]
- Quante, M.; Abrams, J.A.; Lee, Y.; Wang, T.C. Barrett esophagus: What a mouse model can teach us about human disease. Cell Cycle 2012, 11, 4328–4338. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sayin, S.I.; Baumeister, T.; Wang, T.C.; Quante, M. Origins of metaplasia in the esophagus: Is this a GE junction stem cell disease? Dig. Dis. Sci. 2018, 63, 2013–2021. [Google Scholar] [CrossRef] [PubMed]
- Pavlov, K.; Maley, C.C. New models of neoplastic progression in Barrett’s oesophagus. Biochem. Soc. Trans. 2010, 38, 331–336. [Google Scholar] [CrossRef] [Green Version]
- Attwood, S.E.; Harrison, L.A.; Preston, S.L.; Jankowski, J.A. Esophageal adenocarcinoma in “mice and men”: Back to basics! Am. J. Gastroenterol. 2008, 103, 2367–2372. [Google Scholar] [CrossRef]
- Nishijima, K.; Miwa, K.; Miyashita, T.; Kinami, S.; Ninomiya, I.; Fushida, S.; Fujimura, T.; Hattori, T. Impact of the biliary diversion procedure on carcinogenesis in Barrett’s esophagus surgically induced by duodenoesophageal reflux in rats. Ann. Surg. 2004, 240, 57–67. [Google Scholar] [CrossRef]
- Wang, R.H. From reflux esophagitis to Barrett’s esophagus and esophageal adenocarcinoma. World J. Gastroenterol. 2015, 21, 5210–5219. [Google Scholar] [CrossRef] [PubMed]
- Whelan, K.A.; Muir, A.B.; Nakagawa, H. Esophageal 3D culture systems as modeling tools in esophageal epithelial pathobiology and personalized medicine. Cell. Mol. Gastroenterol. Hepatol. 2018, 5, 461–478. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kohata, Y.; Fujiwara, Y.; Machida, H.; Okazaki, H.; Yamagami, H.; Tanigawa, T.; Watanabe, K.; Watanabe, T.; Tominaga, K.; Wei, M.; et al. Role of Th-2 cytokines in the development of Barrett’s esophagus in rats. J. Gastroenterol. 2011, 46, 883–893. [Google Scholar] [CrossRef] [PubMed]
- Kapoor, H.; Lohani, K.R.; Lee, T.H.; Agrawal, D.K.; Mittal, S.K. Animal models of Barrett’s esophagus and esophageal adenocarcinoma-past, present, and future. Clin. Transl. Sci. 2015, 8, 841–847. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nakagawa, H.; Whelan, K.; Lynch, J.P. Mechanisms of Barrett’s oesophagus: Intestinal differentiation, stem cells, and tissue models. Best Pract. Res. Clin. Gastroenterol. 2015, 29, 3–16. [Google Scholar] [CrossRef] [Green Version]
- Underwood, T.J.; Derouet, M.F.; White, M.J.; Noble, F.; Moutasim, K.A.; Smith, E.; Drew, P.A.; Thomas, G.J.; Primrose, J.N.; Blaydes, J.P. A comparison of primary oesophageal squamous epithelial cells with HET-1A in organotypic culture. Biol. Cell 2010, 102, 635–644. [Google Scholar] [CrossRef]
- Sun, T.T.; Shih, C.; Green, H. Keratin cytoskeletons in epithelial cells of internal organs. Proc. Natl. Acad. Sci. USA 1979, 76, 2813–2817. [Google Scholar] [CrossRef] [Green Version]
- Mari, L.; Milano, F.; Parikh, K.; Straub, D.; Everts, V.; Hoeben, K.K.; Fockens, P.; Buttar, N.S.; Krishnadath, K.K. A pSMAD/CDX2 complex is essential for the intestinalization of epithelial metaplasia. Cell Rep. 2014, 7, 1197–1210. [Google Scholar] [CrossRef] [Green Version]
- Samarasena, J.B.; Ahluwalia, A.; Tarnawski, A.S.; Shinoura, S.; Choi, K.D.; Lee, J.G.; Chang, K.J. Expression of nerve growth factor, its trka receptor, and several neuropeptides in porcine esophagus. Implications for interactions between neural, vascular and epithelial components of the esophagus. J. Physiol. Pharmacol. 2015, 66, 415–420. [Google Scholar]
- Bus, P.; Siersema, P.D.; Verbeek, R.E.; van Baal, J.W.P.M. Upregulation of miRNA-143,-145,-192, and-194 in esophageal epithelial cells upon acidic bile salt stimulation. Dis. Esophagus 2014, 27, 591–600. [Google Scholar] [CrossRef]
- Nehra, D.; Howell, P.; Williams, C.P.; Pye, J.K.; Beynon, J. Toxic bile acids in gastro-oesophageal reflux disease: Influence of gastric acidity. Gut 1999, 44, 598–602. [Google Scholar] [CrossRef] [PubMed]
- Pera, M.; Brito, M.J.; Pera, M.; Poulsom, R.; Riera, E.; Grande, L.; Hanby, A.; Wright, N.A. Duodenal-Content reflux esophagitis induces the development of glandular metaplasia and adenosquamous carcinoma in rats. Carcinogenesis 2000, 21, 1587–1591. [Google Scholar] [CrossRef] [PubMed]
- Magierowska, K.; Korbut, E.; Hubalewska-Mazgaj, M.; Surmiak, M.; Chmura, A.; Bakalarz, D.; Buszewicz, G.; Wójcik, D.; Śliwowski, Z.; Ginter, G.; et al. Oxidative gastric mucosal damage induced by ischemia/reperfusion and the mechanisms of its prevention by carbon monoxide-releasing tricarbonyldichlororuthenium (II) dimer. Free Radic. Biol. Med. 2019, 145, 198–208. [Google Scholar] [CrossRef] [PubMed]













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Database No GSE13083 (n = 7): Barrett’s Metaplasia (7 Samples) vs. Normal Squamous Epithelium (7 Samples) | Database No GSE34619 (n = 18): Barrett’s Metaplasia (n =10) vs. Normal Squamous Epithelium (n = 8) | Database No GSE1420 (n = 16): Barrett’s Metaplasia (n = 8) vs. Normal Squamous Epithelium (n = 8) | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Gene Symbol | Predicted Type of Epithelium | Gene ID | logFC | p Value | Gene ID | logFC | p Value | Gene ID | logFC | p Value |
| KRT1 | squamous (esophageal) | 205900_at | −7.49126 | 1.11e−05 * | 7963491 | −3.3024845 | 1.10e−03 * | 205900_at | −2.217323 | 0.277797 |
| KRT4 | squamous (esophageal) | 213240_s_at | −4.6401743 | 5.80e−02 | 7963534 | −5.428193 | 2.16e−05 * | 214399_s_at | 0.9627934 | 0.291337 |
| KRT5 | squamous (esophageal) | 201820_at | −6.9473657 | 1.02e−02 * | 7963427 | −5.19733 | 1.27e−05 * | 201820_at | 0.2659335 | 0.939807 |
| KRT6A, 6B, 6C | squamous (esophageal) | 214580_x_at | −5.0172743 | 3.66e−02 * | 7963410 | −4.5273685 | 7.31e−05 * | 214580_x_at | 0.3930203 | 0.885997 |
| KRT10 | squamous (esophageal) | 207023_x_at | −1.9265443 | 4.80e−03 * | 8015104 | −1.9039765 | 1.43e−02 * | 207023_x_at | 0.0731168 | 0.925068 |
| KRT13 | squamous (esophageal) | 207935_s_at | −5.3344986 | 4.04e−02 * | 8015323 | −5.5419725 | 5.55e−05 * | 207935_s_at | 0.4701058 | 0.861817 |
| KRT14 | squamous (esophageal) | 209351_at | −4.0982471 | 1.17e−01 | 8015366 | −2.2296553 | 1.88e−03 * | 209351_at | 1.928972 | 0.499067 |
| KRT15 | squamous (esophageal) | 204734_at | −6.05938 | 2.09e−03 * | 8015337 | −4.686177 | 2.79e−10 * | 204734_at | −0.327417 | 0.941066 |
| KRT16 | squamous (esophageal) | 209800_at | −5.0731586 | 5.63e−03 * | 8015376 | −3.5205307 | 2.61e−05 * | 209800_at | 1.16582 | 0.682918 |
| KRT17 | squamous (esophageal) | 212236_x_at | −2.6745814 | 8.21e−02 | 8005449 | −1.7824035 | 1.42e−03 * | 205157_s_at | 2.6061311 | 0.297826 |
| KRT23 | squamous (esophageal) | 218963_s_at | −2.0889186 | 3.74e−02 * | 8015133 | −1.2800192 | 7.70e−02 | 218963_s_at | 0.0060589 | 0.997257 |
| KRT24 | squamous (esophageal) | 220267_at | −4.4806771 | 2.26e−03 * | 8015060 | −2.905637 | 7.73e−05 * | 220267_at | −0.676433 | 0.81305 |
| KRT7 | columnar (intestinal) | 209016_s_at | 2.2155471 | 4.07e−02 * | 7955613 | 1.9065172 | 5.46e−06 * | 209016_s_at | 1.6998427 | 0.204673 |
| KRT8 | columnar (intestinal) | 209008_x_at | 6.4172871 | 2.18e−09 * | 7963567 | 4.0091988 | 3.50e−11 * | 209008_x_at | 2.6806701 | 0.027973 * |
| KRT18 | columnar (intestinal) | 201596_x_at | 3.4490243 | 5.03e−06 * | 8154725 | 2.024285 | 2.16e−06 * | 201596_x_at | 1.9282774 | 0.082523 |
| KRT19 | columnar (intestinal) | 201650_at | 1.7536714 | 1.01e−03 * | 8015349 | 0.759105 | 3.08e−02 | 201650_at | 1.2141959 | 0.658343 |
| KRT20 | columnar (intestinal) | 213953_at | 8.5259443 | 3.44e−09 * | 8015124 | 4.374532 | 2.00e−04 * | 213953_at | 4.8072071 | 0.037302 * |
| TFF1 | columnar (intestinal) | 205009_at | 8.2017486 | 1.75e−05 * | 8070579 | 6.4595337 | 1.46e−12 * | 205009_at | 5.6843253 | 0.010407 * |
| TFF2 | columnar (intestinal) | 214476_at | 7.6665729 | 4.86e−05 * | 8070574 | 5.2354248 | 1.23e−08 * | 214476_at | 5.8714482 | 0.006909 * |
| TFF3 | columnar (intestinal) | 204623_at | 8.9897857 | 9.11e−08 * | 8070567 | 2.2346355 | 2.20e−04 * | 204623_at | 3.3738183 | 0.110978 |
| VIL1 | columnar (intestinal) | 209950_s_at | 5.8340643 | 8.63e−09 * | 8078665 | 2.2711873 | 1.88e−08 * | 209950_s_at | 2.5215122 | 0.025483 * |
| MUC1 | columnar (intestinal) | 213693_s_at | 1.5493643 | 1.21e−01 | 7920642 | 1.9449625 | 1.69e−05 * | 213693_s_at | 1.1435938 | 0.427828 |
| MUC2 | columnar (intestinal) | 204673_at | 6.7226214 | 3.16e−06 * | 7937560 | 2.1981597 | 2.77e−03 * | 204673_at | 2.9508572 | 0.271622 |
| MUC3A/B | columnar (intestinal) | 217117_x_at | 0.7757457 | 9.80e−02 | 8135015 | 4.3127432 | 6.68e−09 * | 217117_x_at | 0.9212935 | 0.225325 |
| MUC4 | columnar (intestinal) | 217109_at | −1.07354 | 1.13e−01 | 8092978 | 0.1271057 | 8.92e−01 | 204895_x_at | 1.903752 | 0.360547 |
| MUC5ac | columnar (intestinal) | 214385_s_at | 8.1766871 | 8.25e−07 * | not included in the database | 214385_s_at | 7.1466461 | 0.005318 * | ||
| MUC5B | columnar (intestinal) | 213432_at | 2.7593857 | 4.75e−02 * | 7937612 | 1.6075105 | 2.00e−03 * | 213432_at | 2.3107749 | 0.510519 |
| MUC6 | columnar (intestinal) | 214133_at | 3.1026371 | 7.45e−03 * | 7945595 | 5.9384813 | 1.46e−11 * | 214133_at | 3.4098322 | 0.017934 * |
| MUC12 | columnar (intestinal) | not included in the database | 8135033 | 2.2172345 | 1.33e−03 * | not included in the database | ||||
| MUC13 | columnar (intestinal) | 218687_s_at | 7.3541829 | 5.65e−10 * | 8090180 | 7.3474513 | 1.87e−11 * | 218687_s_at | 3.6273263 | 0.031468 * |
| MUC15 | columnar (intestinal) | not included in the database | 7947156 | −4.7653307 | 4.28e−10 * | not included in the database | ||||
| MUC17 | columnar (intestinal) | not included in the database | 8135048 | 5.953779 | 6.32e−08 * | not included in the database | ||||
| MUC21 | columnar (intestinal) | not included in the database | 8177931 | −5.732196 | 5.46e−07 * | not included in the database | ||||
| Macroscopic Lesion Score | Number of Animals with EGDA (%) (n = 10) |
|---|---|
| 2 | 1 (10%) |
| 3 | 7 (70%) |
| 4 | 2 (20%) |
| Assessed Microscopic Criteria | Number of Animals with EGDA with Presence of the Criteria (%) (n = 10) |
|---|---|
| hyperplasia of squamous epithelium | 10 (100%) |
| fibrosis of lamina propria | 10 (100%) |
| Barrett’s metaplasia | 6 (60%) |
| esophagitis with ulceration | 8 (80%) |
| Human | Rat | |
|---|---|---|
| Esophageal epithelium | non-keratinized | keratinized |
| Esophageal submucosal glands and papillae | Present | Absent |
| Stratum corneum | Absent | Present |
| Squamocolumnar transition at GEJ | yes | no |
| Natural reflux | yes | no |
| Natural BE to EAC progression | yes | no |
| Compartmentalized stomach (forestomach and distal stomach) | no | yes |
| BE progression time | 10 years | Around 2–3 months |
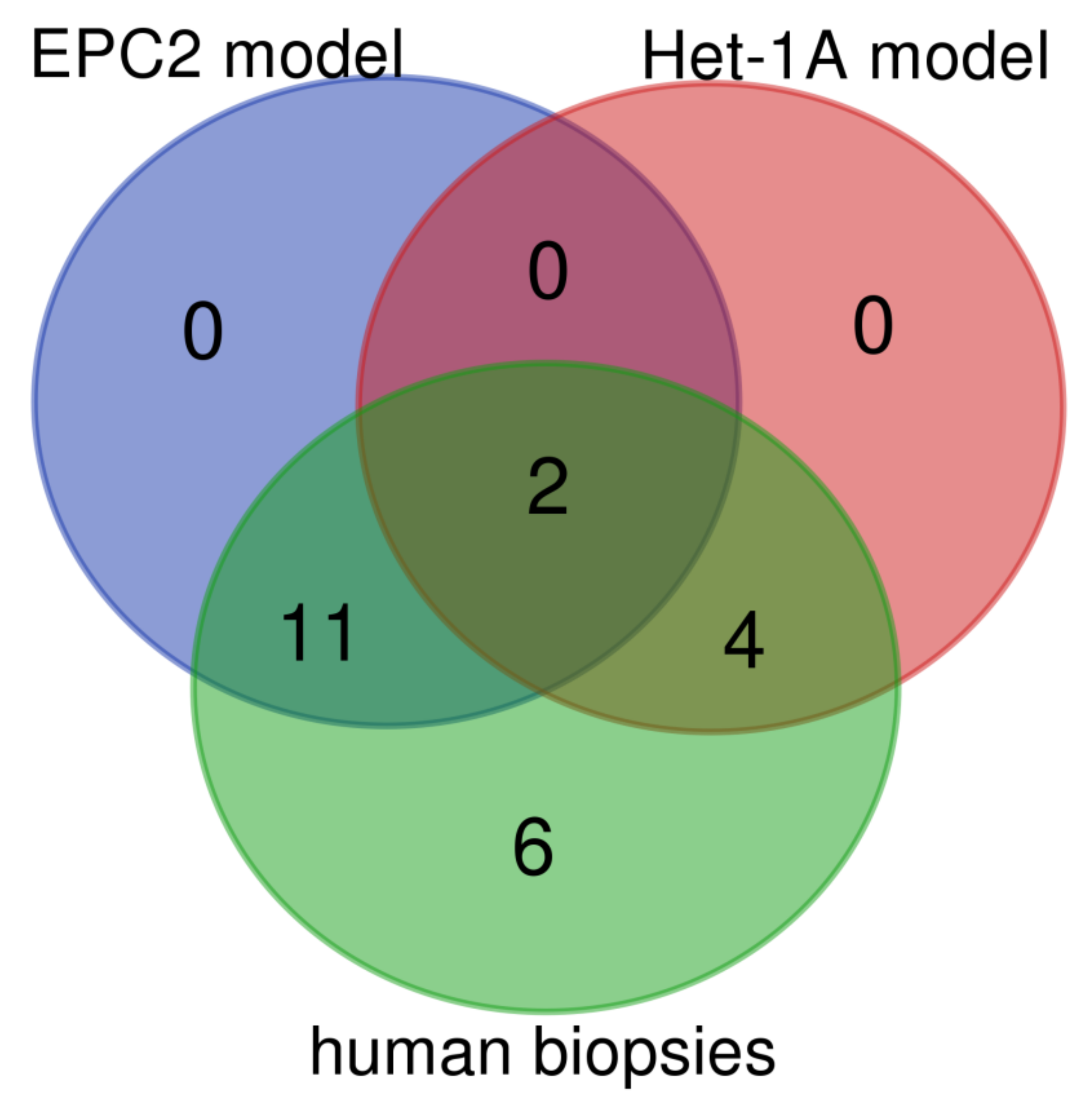
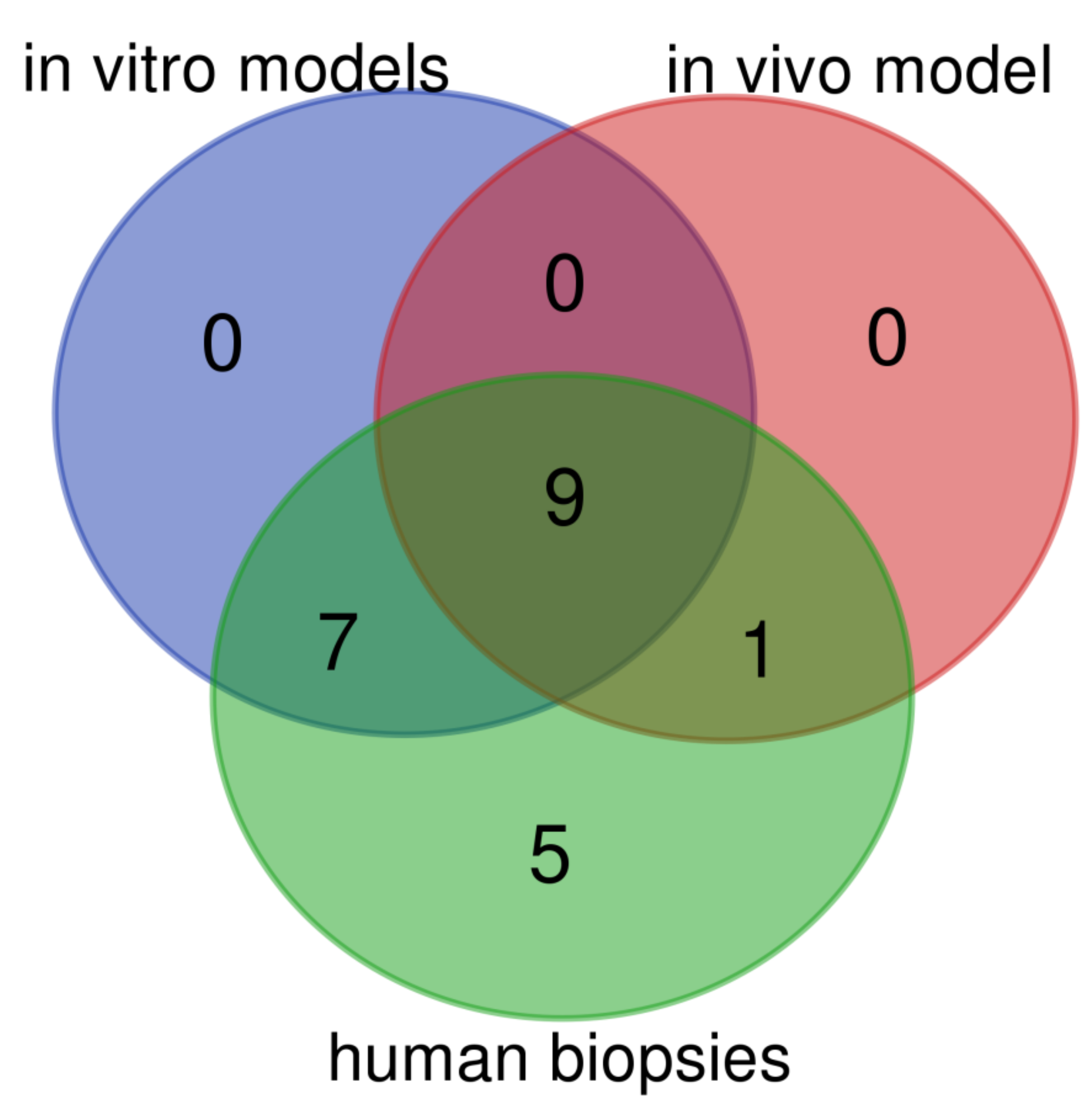
| Gene Symbol | Type of Epithelium | In Vitro Models | In Vivo Model | Human Biopsies | |
|---|---|---|---|---|---|
| Het-1A | EPC2 | ||||
| KRT1 | squamous | n.a. | ↓ | ↑ | ↓ |
| KRT4 | squamous | n.a. | ↓ | ↓ | ↓ |
| KRT5 | squamous | n.a. | ↓ | ↑ | ↓ |
| KRT6 | squamous | ↔ | ↓ | n.d. | ↓ |
| KRT13 | squamous | n.a. | ↓ | ↓ | ↓ |
| KRT14 | squamous | n.a. | ↓ | ↑ | ↓ |
| KRT15 | squamous | ↓ | ↓ | ↓ | ↓ |
| KRT16 | squamous | ↔ | ↓ | n.a. | ↓ |
| KRT23 | squamous | n.a. | ↓ | ↔ | ↓ |
| KRT24 | squamous | n.a | ↓ | n.a. | ↓ |
| KRT7 | columnar | ↔ | ↑ | ↑ | ↑ |
| KRT8 | columnar | ↑ | ↔ | ↑ | ↑ |
| KRT18 | columnar | ↔ | ↑ | ↑ | ↑ |
| KRT20 | columnar | n.a. | n.a. | ↑ | ↑ |
| TFF1 | columnar | n.a. | n.a. | ↓ | ↑ |
| TFF2 | columnar | n.a. | n.a. | n.a. | ↑ |
| TFF3 | columnar | ↑ | ↑ | ↑ | ↑ |
| VIL1 | columnar | ↑ | n.a. | ↔ | ↑ |
| MUC2 | columnar | ↑ | n.a. | ↑ | ↑ |
| MUC3 | columnar | n.a. | n.a. | n.a. | ↑ |
| MUC5B | columnar | n.a. | n.a. | n.a. | |
| MUC6 | columnar | n.a. | n.a. | n.a. | ↑ |
| MUC13 | columnar | ↑ | ↔ | ↑ | ↑ |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Korbut, E.; Janmaat, V.T.; Wierdak, M.; Hankus, J.; Wójcik, D.; Surmiak, M.; Magierowska, K.; Brzozowski, T.; Peppelenbosch, M.P.; Magierowski, M. Molecular Profile of Barrett’s Esophagus and Gastroesophageal Reflux Disease in the Development of Translational Physiological and Pharmacological Studies. Int. J. Mol. Sci. 2020, 21, 6436. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21176436
Korbut E, Janmaat VT, Wierdak M, Hankus J, Wójcik D, Surmiak M, Magierowska K, Brzozowski T, Peppelenbosch MP, Magierowski M. Molecular Profile of Barrett’s Esophagus and Gastroesophageal Reflux Disease in the Development of Translational Physiological and Pharmacological Studies. International Journal of Molecular Sciences. 2020; 21(17):6436. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21176436
Chicago/Turabian StyleKorbut, Edyta, Vincent T Janmaat, Mateusz Wierdak, Jerzy Hankus, Dagmara Wójcik, Marcin Surmiak, Katarzyna Magierowska, Tomasz Brzozowski, Maikel P Peppelenbosch, and Marcin Magierowski. 2020. "Molecular Profile of Barrett’s Esophagus and Gastroesophageal Reflux Disease in the Development of Translational Physiological and Pharmacological Studies" International Journal of Molecular Sciences 21, no. 17: 6436. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21176436