A New Era of Antibiotics: The Clinical Potential of Antimicrobial Peptides

 ,
,  ,
,  , ,
, ,
Abstract
:1. Introduction
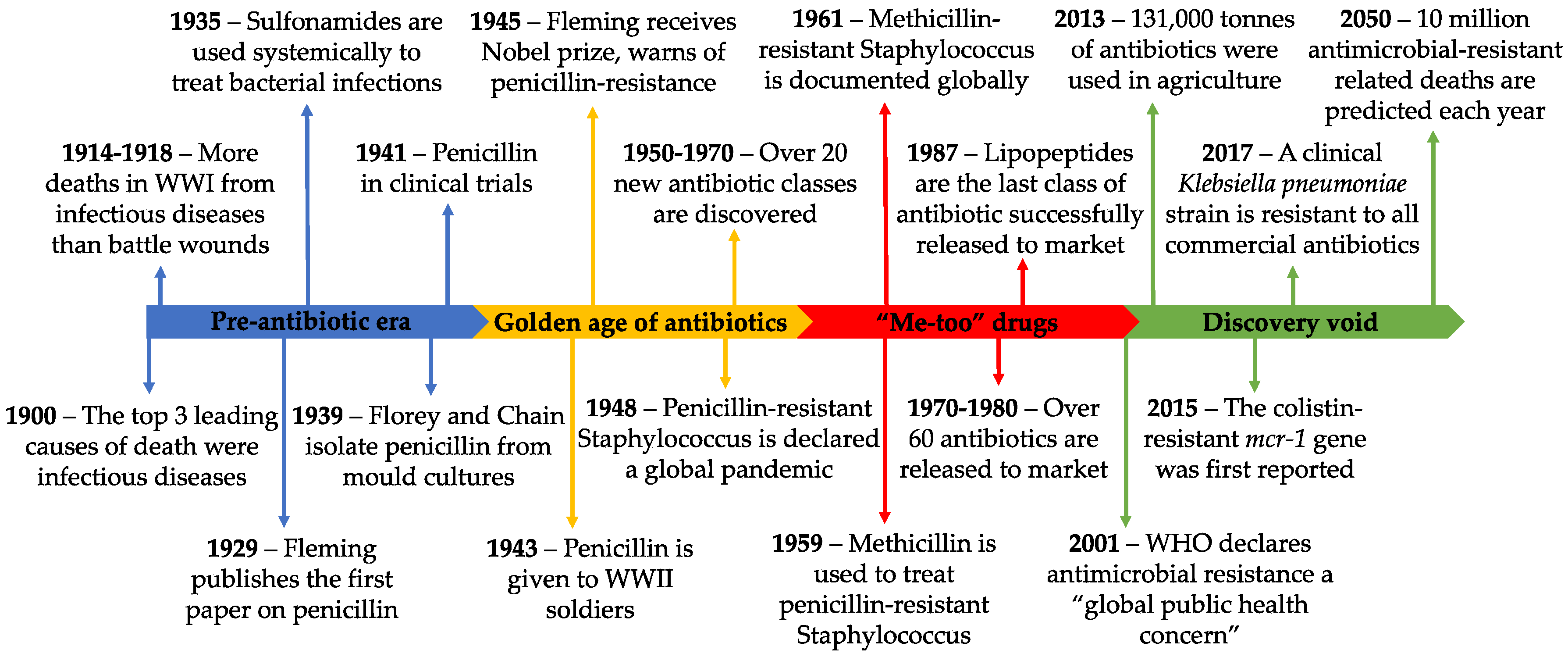
2. History of Antibiotics and Resistance
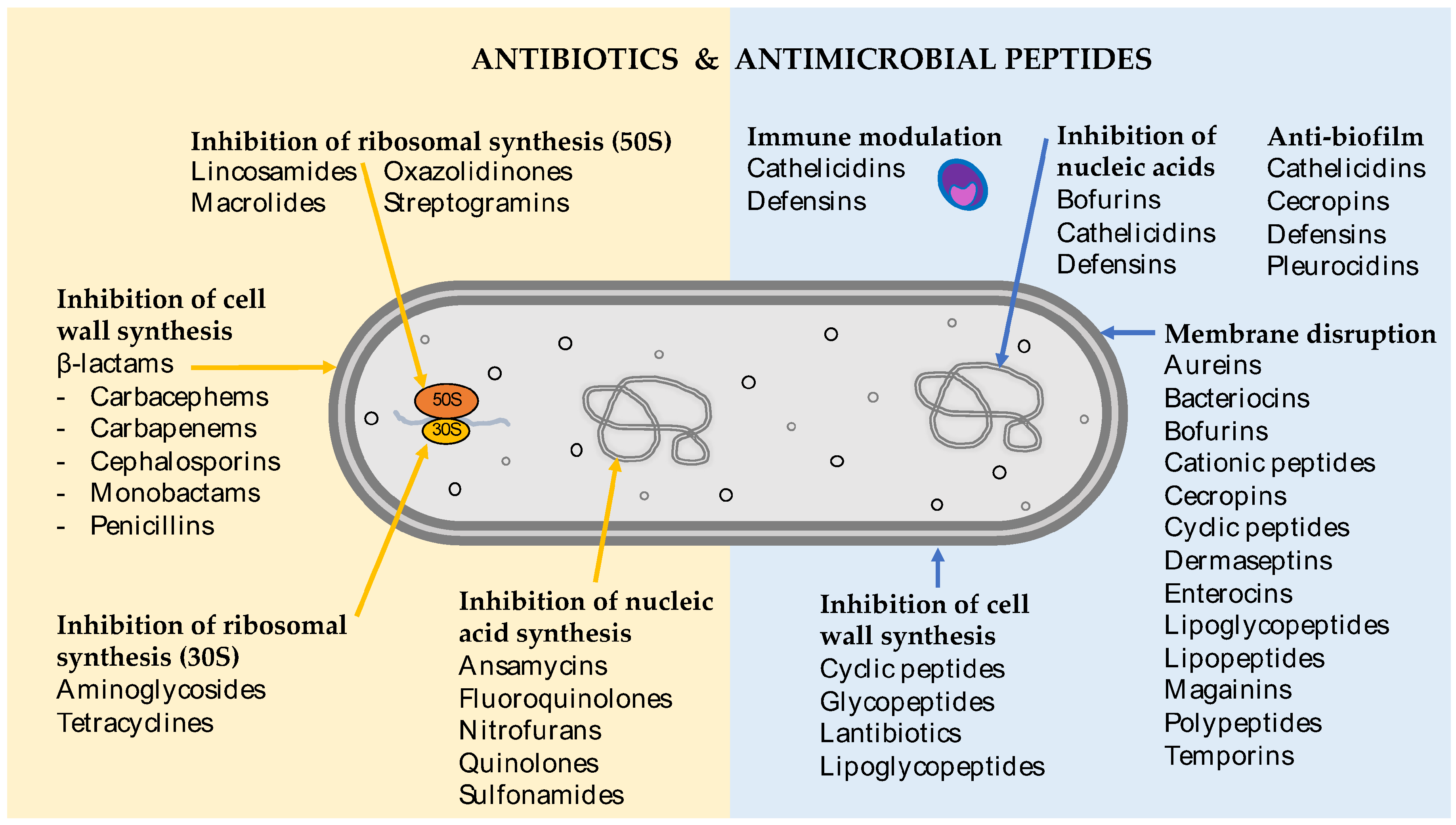
3. Antimicrobial Peptides in Nature
4. Therapeutic Potential of Antimicrobial Peptides
5. Commercially Available Peptide-Based Antibiotics

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Active Ingredient | Origin | Target Organism | Class | Mechanism of Action | Indication | Dosage | Route of Administration | Ref |
|---|---|---|---|---|---|---|---|---|
| Bacitracin | Bacteria (Bacillus subtilis) | Gram-positive bacteria | Cyclic peptide | Inhibits cell wall synthesis | Skin infections | 500 units/g 500 units/g 5000 units/vial | Topical Ophthalmic Intramuscular | [74] |
| Dalbavancin | Teicoplanin derivative | Gram-positive bacteria | Lipoglycopeptide | Inhibits cell wall synthesis | Skin infections | 1000 mg/vial | Intravenous | [75,76] |
| Daptomycin | Bacteria (Streptomyces roseosporus) | Gram-positive bacteria | Lipopeptide | Membrane lysis | Skin infections | 500 mg/vial | Intravenous | [77] |
| Colistin | Bacteria (Bacillus polymyxa) | Gram-negative bacteria | Cyclic peptide | Membrane lysis | Multi drug-resistant gram-negative infections | 150 mg/vial | Intravenous | [78,79] |
| Gramicidin D | Bacteria (Bacillus brevis) | Gram-positive bacteria, some gram-negative bacteria | Mix of three polypeptides | Membrane poration/lysis | Skin and eye infection | 0.25 mg/mL 0.025 mg/mL | Topical Ophthalmic | [80] |
| Oritavancin | Vancomycin derivative | Gram-positive bacteria | Lipoglycopeptide | Membrane lysis and inhibits cell wall synthesis | Skin infections | 800 mg/vial | Intravenous | [81] |
| Polymyxin B | Bacteria (Bacillus polymyxa) | Gram-negative bacteria | Polypeptide | Membrane lysis | Urinary tract and bloodstream infections | 10,000 units/g 10,000 units/g 500,000 units/vial | Ophthalmic Topical Intravenous | [82] |
| Teicoplanin | Bacteria (Actinoplanes teichomyceticus) | Gram-positive bacteria | Glycopeptide | Inhibits cell wall synthesis | Serious gram-positive infections | 400 mg/vial 400 mg/vial | Intramuscular Intravenous | [83] |
| Telavancin | Vancomycin derivative | Gram-positive bacteria | Lipoglycopeptide | Membrane lysis and inhibits cell wall synthesis | Skin infections | 750 mg/vial | Intravenous | [84] |
| Vancomycin | Bacteria (Amycolatopsis orientalis) | Gram-positive bacteria | Glycopeptide | Inhibits cell wall synthesis | Serious gram-positive infections | 250 mg 10 g/vial | Oral Intravenous | [85] |
6. Antimicrobial Peptides in Clinical Trials
| Name | Origin | Target Organism | Class | Mechanism of Action | Indication | Dosage | Route of Administration | Ref |
|---|---|---|---|---|---|---|---|---|
| Phase 3 | ||||||||
| Dusquetide (IMX942, SGX942) | Rational drug design | p62 protein (sequestosome-1) | Synthetic peptide | Immune modulation | Oral complications of chemotherapy | 1.5 mg/mL | Intravenous | NCT03237325 |
| Iseganan (IB-367) | Protegrin analogue | Broad spectrum antibacterial | Synthetic peptide | Bacterial membrane disruption | Oral complications of radiation therapy, Ventilator-associated pneumonia | Undefined 9 mg | Oral rinse Inhalation | NCT00022373 NCT00118781 |
| Mel4 | Melimine analogue | Broad spectrum antibacterial | Synthetic chimeric peptide | Bacterial membrane disruption | Keratitis | Undefined | Ocular | ACTRN1261500072556 |
| Murepavadin (POL7080) | Protegrin-1 synthetic mimic | Pseudomonas | Synthetic peptide | Bacterial membrane disruption via LptD binding | Ventilator-associated pneumonia | Undefined | Intravenous | NCT03409679 |
| Omiganan (MX-226) | Indolicidin analogue | Broad spectrum antifungal, antibacterial | Synthetic peptide | Bacterial membrane disruption | Severe papulopustular rosacea | Undefined | Topical | NCT02576847 |
| p2TA (AB103) | Rational drug design | CD28 receptor on T-helper 1 lymphocytes | Synthetic peptide | Immune modulation | Necrotizing soft tissue infections | 0.5 mg/kg | Intravenous | NCT02469857 |
| Pexiganan (MSI-78) | Magainin analogue | Broad spectrum antibacterial | Synthetic peptide | Bacterial membrane disruption | Diabetic foot ulcers | 0.8% w/w | Topical | NCT01590758 |
| Surotomycin (CB-183,315) | Daptomycin analogue | Gram-positive bacteria | Synthetic cyclic lipopeptide | Bacterial membrane disruption | Clostridium difficile infection | 250 mg | Oral | NCT01597505 |
| Talactoferrin (TLF, rhLF) | Lactoferrin analogue | Gastrointestinal epithelium | Synthetic glycoprotein | Immune modulation | Severe sepsis | 100 mg/mL | Oral solution | NCT01273779 |
| Phase 2 | ||||||||
| Brilacidin (PMX-30063) | Defensin mimetic | Broad spectrum antibacterial | Arylamide foldamer | Bacterial membrane disruption | Oral complications of radiation therapy | 3 mg/mL | Oral rinse | NCT02324335 |
| C16G2 | Novispirin analogue | Streptococcus mutans | Synthetic peptide | Strain selective membrane disruption, intracellular targets | Dental caries | 13.6 mg 9.2 mg | Oral varnish Oral strip | NCT03196219 |
| DPK 060 (GKH17-WWW) | Human protein kininogen derivative | Broad spectrum antibacterial | Synthetic peptide | Bacterial membrane disruption | Acute otitis externa | 2% w/w | Auricular | NCT01447017 |
| EA-230 | Human chorionic gonadotrophin hormone derivative | Proinflammatory immune pathway | Synthetic linear tetrapeptide | Immune modulation | Systemic inflammatory response syndrome | 90 mg/kg | Intravenous | NCT03145220 |
| Exeporfinium chloride (XF-73) | Dicationic porphyrin derivative | Broad spectrum antibacterial | Synthetic porphyrin | Bacterial membrane disruption | Staphylococcal infections | 0.2% w/w | Nasal gel | NCT03915470 |
| LL-37 | Human cathelicidin LL-37 | Antibacterial, antibiofilm | Cathelicidin | Bacterial membrane disruption, immune modulation | Diabetic foot ulcer | 0.5 mg/mL | Topical | NCT04098562 |
| Lytixar (LTX-109) | Host defense peptide derivative | Broad spectrum antibacterial | Synthetic Oligopeptide | Bacterial membrane disruption | Gram-positive skin infections | 5% w/w | Topical | NCT01223222 |
| Melimine | Melittin / protamine splice | Broad spectrum antibacterial | Synthetic chimeric peptide | Bacterial membrane disruption | Keratitis | Undefined | Ocular | ACTRN12613000369729 |
| Novexatin (NP213) | Rational drug design | Antifungal | Synthetic, cyclic peptide | Bacterial membrane disruption | Fungal nail infection | 10% w/w | Topical | NCT02933879 |
| OP-145 | Human cathelicidin LL-37 derivative | Broad spectrum antibacterial | Synthetic peptide | Bacterial membrane disruption | Chronic suppurative otitis media | 0.5 mg/mL | Auricular | ISRCTN12149720 |
| Opebacan (rBPI21, neuprex) | Human recombinant endotoxin-binding protein | Lipopolysaccharides/endotoxins of gram-negative bacteria | Synthetic peptide | Bacterial membrane disruption | Graft versus host disease | 4 mg/kg | Intravenous | NCT00454155 |
| PAC113 (Nal-P-113) | Histatin analogue | Broad spectrum antibacterial | Synthetic peptide | Bacterial membrane disruption, immune modulation, anti-biofilm | Oral candidiasis | 0.15% | Oral rinse | NCT00659971 |
| XOMA-629 (XMP-629) | Human recombinant endotoxin-binding protein | Lipopolysaccharides/endotoxins of gram-negative bacteria | Synthetic peptide | Bacterial membrane disruption | Bacterial skin infections | 1% w/w | Topical | [96] |
| Phase 1 | ||||||||
| hLF1-11 | Lactoferrin derivative | Broad spectrum antibacterial, antifungal | Synthetic peptide | Chelating agent, immune modulation | Staphylococcal bacteremia | 0.5 mg | Intravenous | NCT00509847 |
| WLBU2 (PLG0206) | Rationally designed | Broad spectrum antibacterial, antiviral | Synthetic peptide | Bacterial membrane disruption | Microbial infections | 3 mg/kg | Intravenous | ACTRN12618001920280 |
| Preclinical | ||||||||
| Bac8c | Bactenecin derivative | Broad spectrum antibacterial | Synthetic peptide | Bacterial membrane disruption | Dental carries | MIC E. coli 6 μg/mL | Oral spray | [97,98,99] |
| Bacteriocin OR-7 | Lactobacillus salivarius | Gram negative bacteria, campylobacter jejuni | Bacteriocin | Bacterial membrane disruption | Bacterial infections | MIC C. coli 0.5 μg/mL | Undefined | [100,101] |
| Buforin II | Bufo bufo gargarizans | Broad spectrum antibacterial | Bofurin I | Nucleic acid binding | A. baumannii infections E. coli infections | 1 mg/kg 0.05 mg/mL | Intravenous Oral | [102,103,104,105] |
| CA(1-7)M(2-9) | Cecropin A/melittin splice | Broad spectrum antibacterial | Synthetic chimeric peptide | Bacterial membrane disruption | Bacterial infections | MIC A. baumannii 2 µg/mL | Undefined | [106,107] |
| Colicin E1 | Escherichia coli H22 | Antibacterial | Bacteriocin | Bacterial membrane disruption | Bacterial infections | MIC E. coli 1 µg/mL | Undefined | [108,109] |
| Demegel (D2A21) | Cecropin analogue | Antifungal, antibacterial | Synthetic peptide | Bacterial, fungal membrane disruption | Burn wounds | 1.5% w/w | Topical | [110,111] |
| ETD151 | Heliomycin analogue | Antifungal | Synthetic peptide | Fungal membrane disruption | Fungal infections | Undefined | Intravenous | [112] |
| HB-107 | Cecropin B | Wound healing | Cecropin B fragment | Undefined, nonbacteriostatic | Wound infections | 100 µg/mL | Topical | [113,114] |
| HB-50 | Cecropin analogue | Broad spectrum antibacterial | Synthetic peptide | Bacterial membrane disruption | Wound infections | 1% | Topical | [110,115] |
| HB1345 | Rational design | Broad spectrum antibacterial, anti-inflammatory | Synthetic lipohexapeptide | Bacterial membrane disruption | Skin infections, acne | MIC P. acnes 1 µg/mL | Topical | [116,117] |
| IDR-1002 | Bactenecin derivative | Staphylococcus aureus | Synthetic peptide | Immune modulation | P. aeruginosa lung infections | 50 µM | Intratracheal | [118,119] |
| Lactocin 160 | Lactobacillus rhamnosus | Antibacterial, Gardnerella vaginalis | Bacteriocin | Bacterial membrane disruption | Bacterial vaginosis | 10 mg/mL | Intravaginal | [120] |
| Nisin A | Lactococcus lactis | Antibacterial, spermicidal contraceptive | Type A lantibiotic, bacteriocin | Bacterial membrane disruption | Bacterial infections | 8 µg/mL | Undefined | [121,122] |
| Novarifyn (NP432) | Rationally designed | Broad spectrum antibacterial | Synthetic peptide | Bacterial membrane disruption | Bacterial infections | Undefined | Topical | [123,124] |
| Pediocin PA-1 | Pediococcus acidilactici UL5 | Antibacterial | Bacteriocin | Bacterial membrane disruption | Bacterial infections | 1.8 nM | Undefined | [125,126] |
| Planosporicin | Planomonospora | Gram-positive bacteria | Lantibiotic | Inhibition of cell wall biosynthesis | Methicillin-resistant S. aureus infections | MIC S. aureus 2 µg/mL | Undefined | [127,128] |
| Ruminococcin C | Ruminococcus gnavus | Anti-clostridial | Bacteriocin | Bacterial membrane disruption | Gastrointestinal infections | 1.2 µM | Oral | [129,130,131] |
| SB006 (M6) | Rational design | Gram-negative bacteria | Synthetic peptide | Bacterial membrane disruption | Bacterial infections | 4 µg/mL | Undefined | [132] |
| Syphaxin (SPX1-22) | Leptodactylus syphax | Broad spectrum antibacterial | Ocellatin-S1 | Bacterial membrane disruption | Bacterial infections | MIC S. aureus 64 µg/mL | Undefined | [133] |
| Temporin10a | Rana ornativentris | Gram-positive bacteria | Temporin | Bacterial membrane disruption | Bacterial infections | MIC S. aureus 2 µM | Undefined | [134] |
7. Future Perspectives
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| mcr | Mobilized colistin-resistance |
| MIC | Minimum inhibitory concentration |
| WHO | World Health Organization |
References
- O’Niel, J. Tackling Drug-Resistant Infections Globally: Final Report and Recommendations; Review on Antimicrobial Resistance; HM Government and Welcome Trust: London, UK, 2016. [Google Scholar]
- Alanis, A.J. Resistance to antibiotics: Are we in the post-antibiotic era? Arch. Med. Res. 2005, 36, 697–705. [Google Scholar] [CrossRef]
- Hlth, N.C.E.; Stat, N.C.H. Control of infectious diseases, 1900–1999. JAMA J. Am. Med. Assoc. 1999, 282, 1029–1032, reprinted in MMWR 1999, 48, 621–629. [Google Scholar]
- Cossart, Y.E. The rise and fall of infectious diseases: Australian perspectives, 1914–2014. Med. J. Aust. 2014, 201, S11–S14. [Google Scholar] [CrossRef]
- Fleming, A. On the Antibacterial Action of Cultures of a Penicillium, with Special Reference to Their Use in the Isolation of B. Influenzae. Br. J. Exp. Pathol. 1929, 10, 226–236. [Google Scholar] [CrossRef]
- Fleming, A. Pencillin. Nobel Lecture. 11 December 1945. Available online: https://www.nobelprize.org/prizes/medicine/1945/fleming/lecture (accessed on 8 September 2020).
- Gharaibeh, M.H.; Shatnawi, S.Q. An overview of colistin resistance, mobilized colistin resistance genes dissemination, global responses, and the alternatives to colistin: A review. Vet. World 2019, 12, 1735–1746. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martin, M.J.; Thottathil, S.E.; Newman, T.B. Antibiotics Overuse in Animal Agriculture: A Call to Action for Health Care Providers. Am. J. Public Health 2015, 105, 2409–2410. [Google Scholar] [CrossRef]
- Auta, A.; Hadi, M.A.; Oga, E.; Adewuyi, E.O.; Abdu-Aguye, S.N.; Adeloye, D.; Strickland-Hodge, B.; Morgan, D.J. Global access to antibiotics without prescription in community pharmacies: A systematic review and meta-analysis. J. Infect. 2019, 78, 8–18. [Google Scholar] [CrossRef] [PubMed]
- Milani, R.V.; Wilt, J.K.; Entwisle, J.; Hand, J.; Cazabon, P.; Bohan, J.G. Reducing inappropriate outpatient antibiotic prescribing: Normative comparison using unblinded provider reports. BMJ Open Qual. 2019, 8, e000351. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ierano, C.; Thursky, K.; Marshall, C.; Koning, S.; James, R.; Johnson, S.; Imam, N.; Worth, L.J.; Peel, T. Appropriateness of Surgical Antimicrobial Prophylaxis Practices in Australia. JAMA Netw. Open 2019, 2, e1915003. [Google Scholar] [CrossRef] [Green Version]
- Landstedt, K.; Sharma, A.; Johansson, F.; Stalsby Lundborg, C.; Sharma, M. Antibiotic prescriptions for inpatients having non-bacterial diagnosis at medicine departments of two private sector hospitals in Madhya Pradesh, India: A cross-sectional study. BMJ Open 2017, 7, e012974. [Google Scholar] [CrossRef] [Green Version]
- Shiley, K.T.; Lautenbach, E.; Lee, I. The use of antimicrobial agents after diagnosis of viral respiratory tract infections in hospitalized adults: Antibiotics or anxiolytics? Infect. Control Hosp. Epidemiol. 2010, 31, 1177–1183. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dinh, A.; Duran, C.; Davido, B.; Bouchand, F.; Deconinck, L.; Matt, M.; Senard, O.; Guyot, C.; Levasseur, A.S.; Attal, J.; et al. Impact of an antimicrobial stewardship programme to optimize antimicrobial use for outpatients at an emergency department. J. Hosp. Infect. 2017, 97, 288–293. [Google Scholar] [CrossRef] [PubMed]
- Munita, J.M.; Arias, C.A. Mechanisms of Antibiotic Resistance. Microbiol. Spectr. 2016, 4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thomas, C.M.; Nielsen, K.M. Mechanisms of, and barriers to, horizontal gene transfer between bacteria. Nat. Rev. Microbiol. 2005, 3, 711–721. [Google Scholar] [CrossRef]
- Lawrence, J.G.; Ochman, H. Amelioration of bacterial genomes: Rates of change and exchange. J. Mol. Evol. 1997, 44, 383–397. [Google Scholar] [CrossRef]
- Madsen, J.S.; Burmolle, M.; Hansen, L.H.; Sorensen, S.J. The interconnection between biofilm formation and horizontal gene transfer. FEMS Immunol. Med. Microbiol. 2012, 65, 183–195. [Google Scholar] [CrossRef] [Green Version]
- Long, K.S.; Vester, B. Resistance to linezolid caused by modifications at its binding site on the ribosome. Antimicrob. Agents Chemother. 2012, 56, 603–612. [Google Scholar] [CrossRef] [Green Version]
- Egorov, A.M.; Ulyashova, M.M.; Rubtsova, M.Y. Bacterial Enzymes and Antibiotic Resistance. Acta Nat. 2018, 10, 33–48. [Google Scholar] [CrossRef] [Green Version]
- Charles, P.G.; Grayson, M.L. The dearth of new antibiotic development: Why we should be worried and what we can do about it. Med. J. Aust. 2004, 181, 549–553. [Google Scholar] [CrossRef]
- Frieden, T. Antibiotic Resistance Threats in the United States; Centers for Disease Control and Prevention (CDC): Atlanta, GA, USA, 2019.
- Shields, R.K.; Chen, L.; Cheng, S.; Chavda, K.D.; Press, E.G.; Snyder, A.; Pandey, R.; Doi, Y.; Kreiswirth, B.N.; Nguyen, M.H.; et al. Emergence of Ceftazidime-Avibactam Resistance Due to Plasmid-Borne blaKPC-3 Mutations during Treatment of Carbapenem-Resistant Klebsiella pneumoniae Infections. Antimicrob. Agents Chemother. 2017, 61. [Google Scholar] [CrossRef] [Green Version]
- Jenssen, H.; Hamill, P.; Hancock, R.E. Peptide antimicrobial agents. Clin. Microbiol. Rev. 2006, 19, 491–511. [Google Scholar] [CrossRef] [Green Version]
- Lei, J.; Sun, L.; Huang, S.; Zhu, C.; Li, P.; He, J.; Mackey, V.; Coy, D.H.; He, Q. The antimicrobial peptides and their potential clinical applications. Am. J. Transl. Res. 2019, 11, 3919–3931. [Google Scholar] [PubMed]
- Wang, G. PDB ID: 2K6O. Human LL-37 Structure. 2008. Available online: https://www.wwpdb.org/pdb?id=pdb_00002k6o (accessed on 23 September 2020). [CrossRef]
- Gesell, J.J.; Zasloff, M.; Opella, S.J. PDB ID: 2MAG. NMR Structure of Magainin 2 in DPC Micelles, 10 Structures. 1997. Available online: https://www.rcsb.org/structure/2MAG (accessed on 23 September 2020). [CrossRef]
- Craik, D.J.; Trabi, M.; Schirra, H.J. PDB ID: 1HVZ. RTD-1, a Cyclic Antimicrobial Defensin from Rhesus Macaque Leukocytes. 2001. Available online: https://www.rcsb.org/structure/1HVZ (accessed on 23 September 2020). [CrossRef]
- Eisenberg, D.; Gribskov, M.; Terwilliger, T.C. PDB ID: 2MLT. Melittin. 1990. Available online: https://www.wwpdb.org/pdb?id=pdb_00002mlt (accessed on 23 September 2020). [CrossRef]
- Richter, V.; Lay, F.T.; Hulett, M.D.; Kvansakul, M. PDB ID: 4UJ0. Crystal Structure of the Tomato Defensin TPP3. 2015. Available online: http://www.rcsb.org/pdb/explore/litView.do?structureId=4UJ0 (accessed on 23 September 2020). [CrossRef]
- Sanchez-Barrena, M.J.; Martinez-Ripoll, M.; Galvez, A.; Valdivia, E.; Maqueda, M.; Cruz, V.; Albert, A. PDB ID: 1O83. Crystal Structure of Bacteriocin AS-48 at Ph 7.5, Phosphate Bound Crystal form I. 2002. Available online: http://www.rcsb.org/pdb/explore/litView.do?structureId=1O83 (accessed on 23 September 2020). [CrossRef]
- Diamond, G.; Beckloff, N.; Weinberg, A.; Kisich, K.O. The roles of antimicrobial peptides in innate host defense. Curr. Pharm. Des. 2009, 15, 2377–2392. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, L.J.; Gallo, R.L. Antimicrobial peptides. Curr. Biol. 2016, 26, R14–R19. [Google Scholar] [CrossRef] [PubMed]
- Lai, Y.; Gallo, R.L. AMPed up immunity: How antimicrobial peptides have multiple roles in immune defense. Trends Immunol. 2009, 30, 131–141. [Google Scholar] [CrossRef] [Green Version]
- Lemaitre, B.; Nicolas, E.; Michaut, L.; Reichhart, J.M.; Hoffmann, J.A. The dorsoventral regulatory gene cassette spatzle/Toll/cactus controls the potent antifungal response in Drosophila adults. Cell 1996, 86, 973–983. [Google Scholar] [CrossRef] [Green Version]
- Van‘t Hof, W.; Veerman, E.C.; Helmerhorst, E.J.; Amerongen, A.V. Antimicrobial peptides: Properties and applicability. Biol. Chem. 2001, 382, 597–619. [Google Scholar] [CrossRef]
- Pushpanathan, M.; Gunasekaran, P.; Rajendhran, J. Antimicrobial peptides: Versatile biological properties. Int. J. Pept. 2013, 2013, 675391. [Google Scholar] [CrossRef] [Green Version]
- Kumar, P.; Kizhakkedathu, J.N.; Straus, S.K. Antimicrobial Peptides: Diversity, Mechanism of Action and Strategies to Improve the Activity and Biocompatibility in Vivo. Biomolecules 2018, 8, 4. [Google Scholar] [CrossRef] [Green Version]
- Seyfi, R.; Kahaki, F.A.; Ebrahimi, T.; Montazersaheb, S.; Eyvazi, S.; Babaeipour, V.; Tarhriz, V. Antimicrobial Peptides (AMPs): Roles, Functions and Mechanism of Action. Int. J. Pept. Res. Ther. 2020, 26, 1451–1463. [Google Scholar] [CrossRef]
- Al-Rayahi, I.A.; Sanyi, R.H. The overlapping roles of antimicrobial peptides and complement in recruitment and activation of tumor-associated inflammatory cells. Front. Immunol. 2015, 6, 2. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zasloff, M. Antimicrobial peptides of multicellular organisms. Nature 2002, 415, 389–395. [Google Scholar] [CrossRef] [PubMed]
- Bucki, R.; Leszczynska, K.; Namiot, A.; Sokolowski, W. Cathelicidin LL-37: A multitask antimicrobial peptide. Arch. Immunol. Ther. Exp. 2010, 58, 15–25. [Google Scholar] [CrossRef]
- Agier, J.; Efenberger, M.; Brzezinska-Blaszczyk, E. Cathelicidin impact on inflammatory cells. Cent. Eur. J. Immunol. 2015, 40, 225–235. [Google Scholar] [CrossRef]
- Overhage, J.; Campisano, A.; Bains, M.; Torfs, E.C.; Rehm, B.H.; Hancock, R.E. Human host defense peptide LL-37 prevents bacterial biofilm formation. Infect. Immun. 2008, 76, 4176–4182. [Google Scholar] [CrossRef] [Green Version]
- Dean, S.N.; Bishop, B.M.; van Hoek, M.L. Susceptibility of Pseudomonas aeruginosa Biofilm to Alpha-Helical Peptides: D-enantiomer of LL-37. Front. Microbiol. 2011, 2, 128. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hancock, R.E. Cationic peptides: Effectors in innate immunity and novel antimicrobials. Lancet Infect. Dis. 2001, 1, 156–164. [Google Scholar] [CrossRef]
- Radek, K.; Gallo, R. Antimicrobial peptides: Natural effectors of the innate immune system. Semin. Immunopathol. 2007, 29, 27–43. [Google Scholar] [CrossRef]
- Marxer, M.; Vollenweider, V.; Schmid-Hempel, P. Insect antimicrobial peptides act synergistically to inhibit a trypanosome parasite. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2016, 371. [Google Scholar] [CrossRef]
- Franzenburg, S.; Walter, J.; Kunzel, S.; Wang, J.; Baines, J.F.; Bosch, T.C.; Fraune, S. Distinct antimicrobial peptide expression determines host species-specific bacterial associations. Proc. Natl. Acad. Sci. USA 2013, 110, E3730–E3738. [Google Scholar] [CrossRef] [Green Version]
- Kaplan, C.W.; Sim, J.H.; Shah, K.R.; Kolesnikova-Kaplan, A.; Shi, W.; Eckert, R. Selective membrane disruption: Mode of action of C16G2, a specifically targeted antimicrobial peptide. Antimicrob. Agents Chemother. 2011, 55, 3446–3452. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gaspar, D.; Veiga, A.S.; Castanho, M.A. From antimicrobial to anticancer peptides. A review. Front. Microbiol. 2013, 4, 294. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Andrea Giuliani, G.P. Silvia Fabiole Nicoletto. Antimicrobial peptides: An overview of a promising class of therapeutics. Cent. Eur. J. Biol. 2007, 2, 1–33. [Google Scholar] [CrossRef]
- Hancock, R.E.; Chapple, D.S. Peptide antibiotics. Antimicrob. Agents Chemother. 1999, 43, 1317–1323. [Google Scholar] [CrossRef] [Green Version]
- Marr, A.K.; Gooderham, W.J.; Hancock, R.E. Antibacterial peptides for therapeutic use: Obstacles and realistic outlook. Curr. Opin. Pharmacol. 2006, 6, 468–472. [Google Scholar] [CrossRef]
- Carrick, S.; Parker, S.; Thornton, C.E.; Ghersi, D.; Simes, J.; Wilcken, N. Single agent versus combination chemotherapy for metastatic breast cancer. Cochrane Database Syst. Rev. 2009. [Google Scholar] [CrossRef]
- Doern, C.D. When does 2 plus 2 equal 5? A review of antimicrobial synergy testing. J. Clin. Microbiol. 2014, 52, 4124–4128. [Google Scholar] [CrossRef] [Green Version]
- Wu, X.; Li, Z.; Li, X.; Tian, Y.; Fan, Y.; Yu, C.; Zhou, B.; Liu, Y.; Xiang, R.; Yang, L. Synergistic effects of antimicrobial peptide DP7 combined with antibiotics against multidrug-resistant bacteria. Drug Des. Dev. Ther. 2017, 11, 939–946. [Google Scholar] [CrossRef] [Green Version]
- Kampshoff, F.; Willcox, M.D.P.; Dutta, D. A Pilot Study of the Synergy between Two Antimicrobial Peptides and Two Common Antibiotics. Antibiotics 2019, 8, 60. [Google Scholar] [CrossRef] [Green Version]
- Pizzolato-Cezar, L.R.; Okuda-Shinagawa, N.M.; Machini, M.T. Combinatory Therapy Antimicrobial Peptide-Antibiotic to Minimize the Ongoing Rise of Resistance. Front. Microbiol. 2019, 10, 1703. [Google Scholar] [CrossRef] [Green Version]
- Ruden, S.; Rieder, A.; Chis Ster, I.; Schwartz, T.; Mikut, R.; Hilpert, K. Synergy Pattern of Short Cationic Antimicrobial Peptides against Multidrug-Resistant Pseudomonas aeruginosa. Front. Microbiol. 2019, 10, 2740. [Google Scholar] [CrossRef] [Green Version]
- Pasupuleti, M.; Schmidtchen, A.; Malmsten, M. Antimicrobial peptides: Key components of the innate immune system. Crit. Rev. Biotechnol. 2012, 32, 143–171. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mahlapuu, M.; Hakansson, J.; Ringstad, L.; Bjorn, C. Antimicrobial Peptides: An Emerging Category of Therapeutic Agents. Front. Cell. Infect. Microbiol. 2016, 6, 194. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gai, Z.; Samodelov, S.L.; Kullak-Ublick, G.A.; Visentin, M. Molecular Mechanisms of Colistin-Induced Nephrotoxicity. Molecules 2019, 24, 653. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, F.F.; Brimble, M.A. Using chemical synthesis to optimise antimicrobial peptides in the fight against antimicrobial resistance. Pure Appl. Chem. 2019, 91, 181–198. [Google Scholar] [CrossRef]
- Merrifield, R.B.; Merrifield, E.L.; Juvvadi, P.; Andreu, D.; Boman, H.G. Design and Synthesis of an Antimicrobial Peptides. Ciba Found. Symp. 1994, 186, 5–20. [Google Scholar]
- Chongsiriwatana, N.P.; Patch, J.A.; Czyzewski, A.M.; Dohm, M.T.; Ivankin, A.; Gidalevitz, D.; Zuckermann, R.N.; Barron, A.E. Peptoids that mimic the structure, function, and mechanism of helical antimicrobial peptides. Proc. Natl. Acad. Sci. USA 2008, 105, 2794–2799. [Google Scholar] [CrossRef] [Green Version]
- Thaker, H.D.; Som, A.; Ayaz, F.; Lui, D.; Pan, W.; Scott, R.W.; Anguita, J.; Tew, G.N. Synthetic mimics of antimicrobial peptides with immunomodulatory responses. J. Am. Chem. Soc. 2012, 134, 11088–11091. [Google Scholar] [CrossRef] [Green Version]
- Ebenhan, T.; Gheysens, O.; Kruger, H.G.; Zeevaart, J.R.; Sathekge, M.M. Antimicrobial peptides: Their role as infection-selective tracers for molecular imaging. Biomed. Res. Int. 2014, 2014, 867381. [Google Scholar] [CrossRef]
- Gang, D.; Kim, D.W.; Park, H.S. Cyclic Peptides: Promising Scaffolds for Biopharmaceuticals. Genes 2018, 9, 557. [Google Scholar] [CrossRef] [Green Version]
- Zeng, D.; Debabov, D.; Hartsell, T.L.; Cano, R.J.; Adams, S.; Schuyler, J.A.; McMillan, R.; Pace, J.L. Approved Glycopeptide Antibacterial Drugs: Mechanism of Action and Resistance. Cold Spring Harb. Perspect. Med. 2016, 6, a026989. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fair, R.J.; Tor, Y. Antibiotics and bacterial resistance in the 21st century. Perspect. Med. Chem. 2014, 6, 25–64. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Oliveira, D.M.P.; Forde, B.M.; Kidd, T.J.; Harris, P.N.A.; Schembri, M.A.; Beatson, S.A.; Paterson, D.L.; Walker, M.J. Antimicrobial Resistance in ESKAPE Pathogens. Clin. Microbiol. Rev. 2020, 33. [Google Scholar] [CrossRef] [PubMed]
- Al Jalali, V.; Zeitlinger, M. Clinical Pharmacokinetics and Pharmacodynamics of Telavancin Compared with the Other Glycopeptides. Clin. Pharmacokinet. 2018, 57, 797–816. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nguyen, R.; Khanna, N.R.; Safadi, A.O.; Sun, Y. Bacitracin Topical. In StatPearls; StatPearls Publishing: Treasure Island, FL, USA, 2020. Available online: https://0-www-ncbi-nlm-nih-gov.brum.beds.ac.uk/books/NBK536993/ (accessed on 23 September 2020).
- Chen, A.Y.; Zervos, M.J.; Vazquez, J.A. Dalbavancin: A novel antimicrobial. Int. J. Clin. Pract. 2007, 61, 853–863. [Google Scholar] [CrossRef] [Green Version]
- Malabarba, A.; Goldstein, B.P. Origin, structure, and activity in vitro and in vivo of dalbavancin. J. Antimicrob. Chemother. 2005, 55 (Suppl. S2), ii15–ii20. [Google Scholar] [CrossRef]
- Tedesco, K.L.; Rybak, M.J. Daptomycin. Pharmacotherapy 2004, 24, 41–57. [Google Scholar] [CrossRef]
- Glascott, E.L. Polymyxin B or polymyxin E: Does it really matter? J. Pharm. Pract. Res. 2018, 48, 492–494. [Google Scholar] [CrossRef] [Green Version]
- Li, J.; Nation, R.L.; Turnidge, J.D.; Milne, R.W.; Coulthard, K.; Rayner, C.R.; Paterson, D.L. Colistin: The re-emerging antibiotic for multidrug-resistant Gram-negative bacterial infections. Lancet Infect. Dis. 2006, 6, 589–601. [Google Scholar] [CrossRef]
- Burkhart, B.M.; Gassman, R.M.; Langs, D.A.; Pangborn, W.A.; Duax, W.L.; Pletnev, V. Gramicidin D conformation, dynamics and membrane ion transport. Biopolymers 1999, 51, 129–144. [Google Scholar] [CrossRef]
- Bouza, E.; Burillo, A. Oritavancin: A novel lipoglycopeptide active against Gram-positive pathogens including multiresistant strains. Int. J. Antimicrob. Agents 2010, 36, 401–407. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zavascki, A.P.; Goldani, L.Z.; Li, J.; Nation, R.L. Polymyxin B for the treatment of multidrug-resistant pathogens: A critical review. J. Antimicrob. Chemother. 2007, 60, 1206–1215. [Google Scholar] [CrossRef] [Green Version]
- Wilson, A.P. Clinical pharmacokinetics of teicoplanin. Clin. Pharmacokinet. 2000, 39, 167–183. [Google Scholar] [CrossRef] [PubMed]
- Saravolatz, L.D.; Stein, G.E.; Johnson, L.B. Telavancin: A novel lipoglycopeptide. Clin. Infect. Dis. 2009, 49, 1908–1914. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Patel, S.; Preuss, C.V.; Bernice, F. Vancomycin. In StatPearls; StatPearls Publishing: Treasure Island, FL, USA, 2020. Available online: https://0-www-ncbi-nlm-nih-gov.brum.beds.ac.uk/books/NBK459263/ (accessed on 23 September 2020).
- Lewies, A.; Du Plessis, L.H.; Wentzel, J.F. Antimicrobial Peptides: The Achilles’ Heel of Antibiotic Resistance? Probiotics Antimicrob. Proteins 2019, 11, 370–381. [Google Scholar] [CrossRef] [PubMed]
- Higgins, D.L.; Chang, R.; Debabov, D.V.; Leung, J.; Wu, T.; Krause, K.M.; Sandvik, E.; Hubbard, J.M.; Kaniga, K.; Schmidt, D.E., Jr.; et al. Telavancin, a multifunctional lipoglycopeptide, disrupts both cell wall synthesis and cell membrane integrity in methicillin-resistant Staphylococcus aureus. Antimicrob. Agents Chemother. 2005, 49, 1127–1134. [Google Scholar] [CrossRef] [Green Version]
- Lunde, C.S.; Hartouni, S.R.; Janc, J.W.; Mammen, M.; Humphrey, P.P.; Benton, B.M. Telavancin disrupts the functional integrity of the bacterial membrane through targeted interaction with the cell wall precursor lipid II. Antimicrob. Agents Chemother. 2009, 53, 3375–3383. [Google Scholar] [CrossRef] [Green Version]
- Das, B.; Sarkar, C.; Das, D.; Gupta, A.; Kalra, A.; Sahni, S. Telavancin: A novel semisynthetic lipoglycopeptide agent to counter the challenge of resistant Gram-positive pathogens. Ther. Adv. Infect. Dis. 2017, 4, 49–73. [Google Scholar] [CrossRef] [Green Version]
- Bahar, A.A.; Ren, D. Antimicrobial peptides. Pharmaceuticals 2013, 6, 1543–1575. [Google Scholar] [CrossRef] [Green Version]
- Pfalzgraff, A.; Brandenburg, K.; Weindl, G. Antimicrobial Peptides and Their Therapeutic Potential for Bacterial Skin Infections and Wounds. Front. Pharmacol. 2018, 9, 281. [Google Scholar] [CrossRef]
- Dutta, D.; Vijay, A.K.; Kumar, N.; Willcox, M.D. Melimine-Coated Antimicrobial Contact Lenses Reduce Microbial Keratitis in an Animal Model. Investig. Ophthalmol. Vis. Sci. 2016, 57, 5616–5624. [Google Scholar] [CrossRef] [Green Version]
- Yasir, M.; Dutta, D.; Willcox, M.D.P. Comparative mode of action of the antimicrobial peptide melimine and its derivative Mel4 against Pseudomonas aeruginosa. Sci. Rep. 2019, 9, 7063. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dutta, D.; Ozkan, J.; Willcox, M.D. Biocompatibility of antimicrobial melimine lenses: Rabbit and human studies. Optom. Vis. Sci. 2014, 91, 570–581. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yasir, M.; Dutta, D.; Hossain, K.R.; Chen, R.; Ho, K.K.K.; Kuppusamy, R.; Clarke, R.J.; Kumar, N.; Willcox, M.D.P. Mechanism of Action of Surface Immobilized Antimicrobial Peptides against Pseudomonas aeruginosa. Front. Microbiol. 2019, 10, 3053. [Google Scholar] [CrossRef] [PubMed]
- Jenssen, H.; Hancock, R.E. Therapeutic potential of HDPs as immunomodulatory agents. Methods Mol. Biol. 2010, 618, 329–347. [Google Scholar] [CrossRef]
- Hilpert, K.; Volkmer-Engert, R.; Walter, T.; Hancock, R.E. High-throughput generation of small antibacterial peptides with improved activity. Nat. Biotechnol. 2005, 23, 1008–1012. [Google Scholar] [CrossRef]
- Spindler, E.C.; Hale, J.D.; Giddings, T.H., Jr.; Hancock, R.E.; Gill, R.T. Deciphering the mode of action of the synthetic antimicrobial peptide Bac8c. Antimicrob. Agents Chemother. 2011, 55, 1706–1716. [Google Scholar] [CrossRef] [Green Version]
- Xiong, K.; Chen, X.; Hu, H.; Hou, H.; Gao, P.; Zou, L. Antimicrobial Effect of a Peptide Containing Novel Oral Spray on Streptococcus mutans. Biomed. Res. Int. 2020, 2020, 6853652. [Google Scholar] [CrossRef] [Green Version]
- Stern, N.J.; Svetoch, E.A.; Eruslanov, B.V.; Perelygin, V.V.; Mitsevich, E.V.; Mitsevich, I.P.; Pokhilenko, V.D.; Levchuk, V.P.; Svetoch, O.E.; Seal, B.S. Isolation of a Lactobacillus salivarius strain and purification of its bacteriocin, which is inhibitory to Campylobacter jejuni in the chicken gastrointestinal system. Antimicrob. Agents Chemother. 2006, 50, 3111–3116. [Google Scholar] [CrossRef] [Green Version]
- Hoang, K.V.; Stern, N.J.; Saxton, A.M.; Xu, F.; Zeng, X.; Lin, J. Prevalence, development, and molecular mechanisms of bacteriocin resistance in Campylobacter. Appl. Environ. Microbiol. 2011, 77, 2309–2316. [Google Scholar] [CrossRef] [Green Version]
- Giacometti, A.; Cirioni, O.; Del Prete, M.S.; Paggi, A.M.; D’Errico, M.M.; Scalise, G. Combination studies between polycationic peptides and clinically used antibiotics against Gram-positive and Gram-negative bacteria. Peptides 2000, 21, 1155–1160. [Google Scholar] [CrossRef]
- Park, C.B.; Kim, H.S.; Kim, S.C. Mechanism of action of the antimicrobial peptide buforin II: Buforin II kills microorganisms by penetrating the cell membrane and inhibiting cellular functions. Biochem. Biophys. Res. Commun. 1998, 244, 253–257. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cirioni, O.; Silvestri, C.; Ghiselli, R.; Orlando, F.; Riva, A.; Gabrielli, E.; Mocchegiani, F.; Cianforlini, N.; Trombettoni, M.M.; Saba, V.; et al. Therapeutic efficacy of buforin II and rifampin in a rat model of Acinetobacter baumannii sepsis. Crit. Care Med. 2009, 37, 1403–1407. [Google Scholar] [CrossRef] [PubMed]
- Tang, Z.R.; Deng, H.; Zhang, X.L.; Zen, Y.; Xiao, D.F.; Sun, W.Z.; Zhang, Z. Effects of orally administering the antimicrobial peptide buforin II on small intestinal mucosal membrane integrity, the expression of tight junction proteins and protective factors in weaned piglets challenged by enterotoxigenic Escherichia coli. Anim. Feed Sci. Technol. 2013, 186, 177–185. [Google Scholar] [CrossRef]
- Sang, Y.; Blecha, F. Antimicrobial peptides and bacteriocins: Alternatives to traditional antibiotics. Anim. Health Res. Rev. 2008, 9, 227–235. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rodriguez-Hernandez, M.J.; Saugar, J.; Docobo-Perez, F.; de la Torre, B.G.; Pachon-Ibanez, M.E.; Garcia-Curiel, A.; Fernandez-Cuenca, F.; Andreu, D.; Rivas, L.; Pachon, J. Studies on the antimicrobial activity of cecropin A-melittin hybrid peptides in colistin-resistant clinical isolates of Acinetobacter baumannii. J. Antimicrob. Chemother. 2006, 58, 95–100. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cursino, L.; Smajs, D.; Smarda, J.; Nardi, R.M.; Nicoli, J.R.; Chartone-Souza, E.; Nascimento, A.M. Exoproducts of the Escherichia coli strain H22 inhibiting some enteric pathogens both in vitro and in vivo. J. Appl. Microbiol. 2006, 100, 821–829. [Google Scholar] [CrossRef]
- Budic, M.; Rijavec, M.; Petkovsek, Z.; Zgur-Bertok, D. Escherichia coli bacteriocins: Antimicrobial efficacy and prevalence among isolates from patients with bacteraemia. PLoS ONE 2011, 6, e28769. [Google Scholar] [CrossRef]
- Zhang, L.; Falla, T.J. Antimicrobial peptides: Therapeutic potential. Expert Opin. Pharmacother. 2006, 7, 653–663. [Google Scholar] [CrossRef]
- Chalekson, C.P.; Neumeister, M.W.; Jaynes, J. Improvement in burn wound infection and survival with antimicrobial peptide D2A21 (Demegel). Plast. Reconstr. Surg. 2002, 109, 1338–1343. [Google Scholar] [CrossRef]
- Andres, E.; Dimarcq, J.L. Cationic antimicrobial peptides: Update of clinical development. J. Intern. Med. 2004, 255, 519–520. [Google Scholar] [CrossRef] [PubMed]
- Steinstraesser, L.; Hirsch, T.; Schulte, M.; Kueckelhaus, M.; Jacobsen, F.; Mersch, E.A.; Stricker, I.; Afacan, N.; Jenssen, H.; Hancock, R.E.; et al. Innate defense regulator peptide 1018 in wound healing and wound infection. PLoS ONE 2012, 7, e39373. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, P.H.; Rudisill, J.A.; Lin, K.H.; Zhang, L.; Harris, S.M.; Falla, T.J.; Gallo, R.L. HB-107, a nonbacteriostatic fragment of the antimicrobial peptide cecropin B, accelerates murine wound repair. Wound Repair Regen. 2004, 12, 351–358. [Google Scholar] [CrossRef] [PubMed]
- Harris, S.M.; Zhang, L.J.; Parente, J.; Rodeheaver, G.T.; Falla, T.J. HB-50: A pre-clinical study of a prophylactic for wound infection. In Understanding Biology Using Peptides; Springer: New York, NY, USA, 2006; pp. 285–286. [Google Scholar] [CrossRef]
- Javia, A.; Amrutiya, J.; Lalani, R.; Patel, V.; Bhatt, P.; Misra, A. Antimicrobial peptide delivery: An emerging therapeutic for the treatment of burn and wounds. Ther. Deliv. 2018, 9, 375–386. [Google Scholar] [CrossRef]
- Sierra, J.M.; Fuste, E.; Rabanal, F.; Vinuesa, T.; Vinas, M. An overview of antimicrobial peptides and the latest advances in their development. Expert Opin. Biol. Ther. 2017, 17, 663–676. [Google Scholar] [CrossRef]
- Nijnik, A.; Madera, L.; Ma, S.; Waldbrook, M.; Elliott, M.R.; Easton, D.M.; Mayer, M.L.; Mullaly, S.C.; Kindrachuk, J.; Jenssen, H.; et al. Synthetic cationic peptide IDR-1002 provides protection against bacterial infections through chemokine induction and enhanced leukocyte recruitment. J. Immunol. 2010, 184, 2539–2550. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wuerth, K.C.; Falsafi, R.; Hancock, R.E.W. Synthetic host defense peptide IDR-1002 reduces inflammation in Pseudomonas aeruginosa lung infection. PLoS ONE 2017, 12, e0187565. [Google Scholar] [CrossRef] [Green Version]
- Dover, S.E.; Aroutcheva, A.A.; Faro, S.; Chikindas, M.L. Safety study of an antimicrobial peptide lactocin 160, produced by the vaginal Lactobacillus rhamnosus. Infect. Dis. Obstet. Gynecol. 2007, 2007, 78248. [Google Scholar] [CrossRef] [Green Version]
- Reddy, K.V.; Aranha, C.; Gupta, S.M.; Yedery, R.D. Evaluation of antimicrobial peptide nisin as a safe vaginal contraceptive agent in rabbits: In vitro and in vivo studies. Reproduction 2004, 128, 117–126. [Google Scholar] [CrossRef]
- Field, D.; R, O.C.; Cotter, P.D.; Ross, R.P.; Hill, C. In Vitro Activities of Nisin and Nisin Derivatives Alone and in Combination with Antibiotics against Staphylococcus Biofilms. Front. Microbiol. 2016, 7, 508. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boto, A.; Perez de la Lastra, J.M.; Gonzalez, C.C. The Road from Host-Defense Peptides to a New Generation of Antimicrobial Drugs. Molecules 2018, 23, 311. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kawano, Y.; Jordan, O.; Hanawa, T.; Borchard, G.; Patrulea, V. Are Antimicrobial Peptide Dendrimers an Escape from ESKAPE? Adv. Wound Care 2020. [Google Scholar] [CrossRef] [PubMed]
- Dabour, N.; Zihler, A.; Kheadr, E.; Lacroix, C.; Fliss, I. In vivo study on the effectiveness of pediocin PA-1 and Pediococcus acidilactici UL5 at inhibiting Listeria monocytogenes. Int. J. Food Microbiol. 2009, 133, 225–233. [Google Scholar] [CrossRef] [PubMed]
- Oppegard, C.; Fimland, G.; Anonsen, J.H.; Nissen-Meyer, J. The Pediocin PA-1 Accessory Protein Ensures Correct Disulfide Bond Formation in the Antimicrobial Peptide Pediocin PA-1. Biochemistry 2015, 54, 2967–2974. [Google Scholar] [CrossRef]
- Castiglione, F.; Cavaletti, L.; Losi, D.; Lazzarini, A.; Carrano, L.; Feroggio, M.; Ciciliato, I.; Corti, E.; Candiani, G.; Marinelli, F.; et al. A novel lantibiotic acting on bacterial cell wall synthesis produced by the uncommon actinomycete Planomonospora sp. Biochemistry 2007, 46, 5884–5895. [Google Scholar] [CrossRef] [PubMed]
- Hammami, R.; Fernandez, B.; Lacroix, C.; Fliss, I. Anti-infective properties of bacteriocins: An update. Cell. Mol. Life Sci. 2013, 70, 2947–2967. [Google Scholar] [CrossRef] [PubMed]
- Crost, E.H.; Ajandouz, E.H.; Villard, C.; Geraert, P.A.; Puigserver, A.; Fons, M. Ruminococcin C, a new anti-Clostridium perfringens bacteriocin produced in the gut by the commensal bacterium Ruminococcus gnavus E1. Biochimie 2011, 93, 1487–1494. [Google Scholar] [CrossRef]
- Nagarajan, K.; Marimuthu, S.K.; Palanisamy, S.; Subbiah, L. Peptide Therapeutics Versus Superbugs: Highlight on Current Research and Advancements. Int. J. Pept. Res. Ther. 2018, 24, 19–33. [Google Scholar] [CrossRef]
- Roblin, C.; Chiumento, S.; Bornet, O.; Nouailler, M.; Muller, C.S.; Jeannot, K.; Basset, C.; Kieffer-Jaquinod, S.; Coute, Y.; Torelli, S.; et al. The unusual structure of Ruminococcin C1 antimicrobial peptide confers clinical properties. Proc. Natl. Acad. Sci. USA 2020, 117, 19168–19177. [Google Scholar] [CrossRef] [PubMed]
- Rossetto, G.; Bergese, P.; Colombi, P.; Depero, L.E.; Giuliani, A.; Nicoletto, S.F.; Pirri, G. Atomic force microscopy evaluation of the effects of a novel antimicrobial multimeric peptide on Pseudomonas aeruginosa. Nanomedicine 2007, 3, 198–207. [Google Scholar] [CrossRef]
- Dourado, F.S.; Leite, J.R.; Silva, L.P.; Melo, J.A.; Bloch, C., Jr.; Schwartz, E.F. Antimicrobial peptide from the skin secretion of the frog Leptodactylus syphax. Toxicon 2007, 50, 572–580. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.B.; Iwamuro, S.; Knoop, F.C.; Conlon, J.M. Antimicrobial peptides from the skin of the Japanese mountain brown frog, Rana ornativentris. J. Pept. Res. 2001, 58, 349–356. [Google Scholar] [CrossRef] [PubMed]
- Rautenbach, M.; Troskie, A.M.; Vosloo, J.A. Antifungal peptides: To be or not to be membrane active. Biochimie 2016, 130, 132–145. [Google Scholar] [CrossRef] [PubMed]
- Fernandez de Ullivarri, M.; Arbulu, S.; Garcia-Gutierrez, E.; Cotter, P.D. Antifungal Peptides as Therapeutic Agents. Front. Cell. Infect. Microbiol. 2020, 10, 105. [Google Scholar] [CrossRef]
- Sherwood, E.J.; Bibb, M.J. The antibiotic planosporicin coordinates its own production in the actinomycete Planomonospora alba. Proc. Natl. Acad. Sci. USA 2013, 110, E2500–E2509. [Google Scholar] [CrossRef] [Green Version]
- Kudrimoti, M.; Curtis, A.; Azawi, S.; Worden, F.; Katz, S.; Adkins, D.; Bonomi, M.; Elder, J.; Sonis, S.T.; Straube, R.; et al. Dusquetide: A novel innate defense regulator demonstrating a significant and consistent reduction in the duration of oral mucositis in preclinical data and a randomized, placebo-controlled phase 2a clinical study. J. Biotechnol. 2016, 239, 115–125. [Google Scholar] [CrossRef] [Green Version]
- Wang, H.Y.; Lin, L.; Tan, L.S.; Yu, H.Y.; Cheng, J.W.; Pan, Y.P. Molecular pathways underlying inhibitory effect of antimicrobial peptide Nal-P-113 on bacteria biofilms formation of Porphyromonas gingivalis W83 by DNA microarray. BMC Microbiol. 2017, 17, 37. [Google Scholar] [CrossRef] [Green Version]
- Brown, M.R.; Allison, D.G.; Gilbert, P. Resistance of bacterial biofilms to antibiotics: A growth-rate related effect? J. Antimicrob. Chemother. 1988, 22, 777–780. [Google Scholar] [CrossRef]
- Deva, A.K.; Adams, W.P.; Vickery, K. The Role of Bacterial Biofilms in Device-Associated Infection. Plast. Reconstr. Surg. 2013, 132, 1319–1328. [Google Scholar] [CrossRef] [Green Version]
- Yasir, M.; Willcox, M.D.P.; Dutta, D. Action of Antimicrobial Peptides against Bacterial Biofilms. Materials 2018, 11, 2468. [Google Scholar] [CrossRef] [Green Version]
- Fields, F.R.; Carothers, K.E.; Balsara, R.D.; Ploplis, V.A.; Castellino, F.J.; Lee, S.W. Rational design of syn-safencin, a novel linear antimicrobial peptide derived from the circular bacteriocin safencin AS-48. J. Antibiot. 2018, 71, 592–600. [Google Scholar] [CrossRef]
- Van der Worp, H.B.; Howells, D.W.; Sena, E.S.; Porritt, M.J.; Rewell, S.; O’Collins, V.; Macleod, M.R. Can animal models of disease reliably inform human studies? PLoS Med. 2010, 7, e1000245. [Google Scholar] [CrossRef] [Green Version]
- Kollef, M.; Pittet, D.; Sanchez Garcia, M.; Chastre, J.; Fagon, J.Y.; Bonten, M.; Hyzy, R.; Fleming, T.R.; Fuchs, H.; Bellm, L.; et al. A randomized double-blind trial of iseganan in prevention of ventilator-associated pneumonia. Am. J. Respir. Crit. Care Med. 2006, 173, 91–97. [Google Scholar] [CrossRef]
- Magana, M.; Pushpanathan, M.; Santos, A.L.; Leanse, L.; Fernandez, M.; Ioannidis, A.; Giulianotti, M.A.; Apidianakis, Y.; Bradfute, S.; Ferguson, A.L.; et al. The value of antimicrobial peptides in the age of resistance. Lancet Infect. Dis. 2020. [Google Scholar] [CrossRef]
- Greber, K.E.; Dawgul, M. Antimicrobial Peptides under Clinical Trials. Curr. Top. Med. Chem. 2017, 17, 620–628. [Google Scholar] [CrossRef] [PubMed]
- Divyashree, M.; Mani, M.K.; Reddy, D.; Kumavath, R.; Ghosh, P.; Azevedo, V.; Barh, D. Clinical Applications of Antimicrobial Peptides (AMPs): Where Do We Stand Now? Protein Pept. Lett. 2020, 27, 120–134. [Google Scholar] [CrossRef] [PubMed]
- Martin, L.; van Meegern, A.; Doemming, S.; Schuerholz, T. Antimicrobial Peptides in Human Sepsis. Front. Immunol. 2015, 6, 404. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gordon, Y.J.; Romanowski, E.G.; McDermott, A.M. A review of antimicrobial peptides and their therapeutic potential as anti-infective drugs. Curr. Eye Res. 2005, 30, 505–515. [Google Scholar] [CrossRef]
- Theuretzbacher, U.; Outterson, K.; Engel, A.; Karlen, A. The global preclinical antibacterial pipeline. Nat. Rev. Microbiol. 2020, 18, 275–285. [Google Scholar] [CrossRef] [Green Version]




© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Browne, K.; Chakraborty, S.; Chen, R.; Willcox, M.D.; Black, D.S.; Walsh, W.R.; Kumar, N. A New Era of Antibiotics: The Clinical Potential of Antimicrobial Peptides. Int. J. Mol. Sci. 2020, 21, 7047. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21197047
Browne K, Chakraborty S, Chen R, Willcox MD, Black DS, Walsh WR, Kumar N. A New Era of Antibiotics: The Clinical Potential of Antimicrobial Peptides. International Journal of Molecular Sciences. 2020; 21(19):7047. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21197047
Chicago/Turabian StyleBrowne, Katrina, Sudip Chakraborty, Renxun Chen, Mark DP Willcox, David StClair Black, William R Walsh, and Naresh Kumar. 2020. "A New Era of Antibiotics: The Clinical Potential of Antimicrobial Peptides" International Journal of Molecular Sciences 21, no. 19: 7047. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21197047