Role of Plasma Gelsolin Protein in the Final Stage of Erythropoiesis and in Correction of Erythroid Dysplasia In Vitro


Abstract
:1. Introduction
2. Results
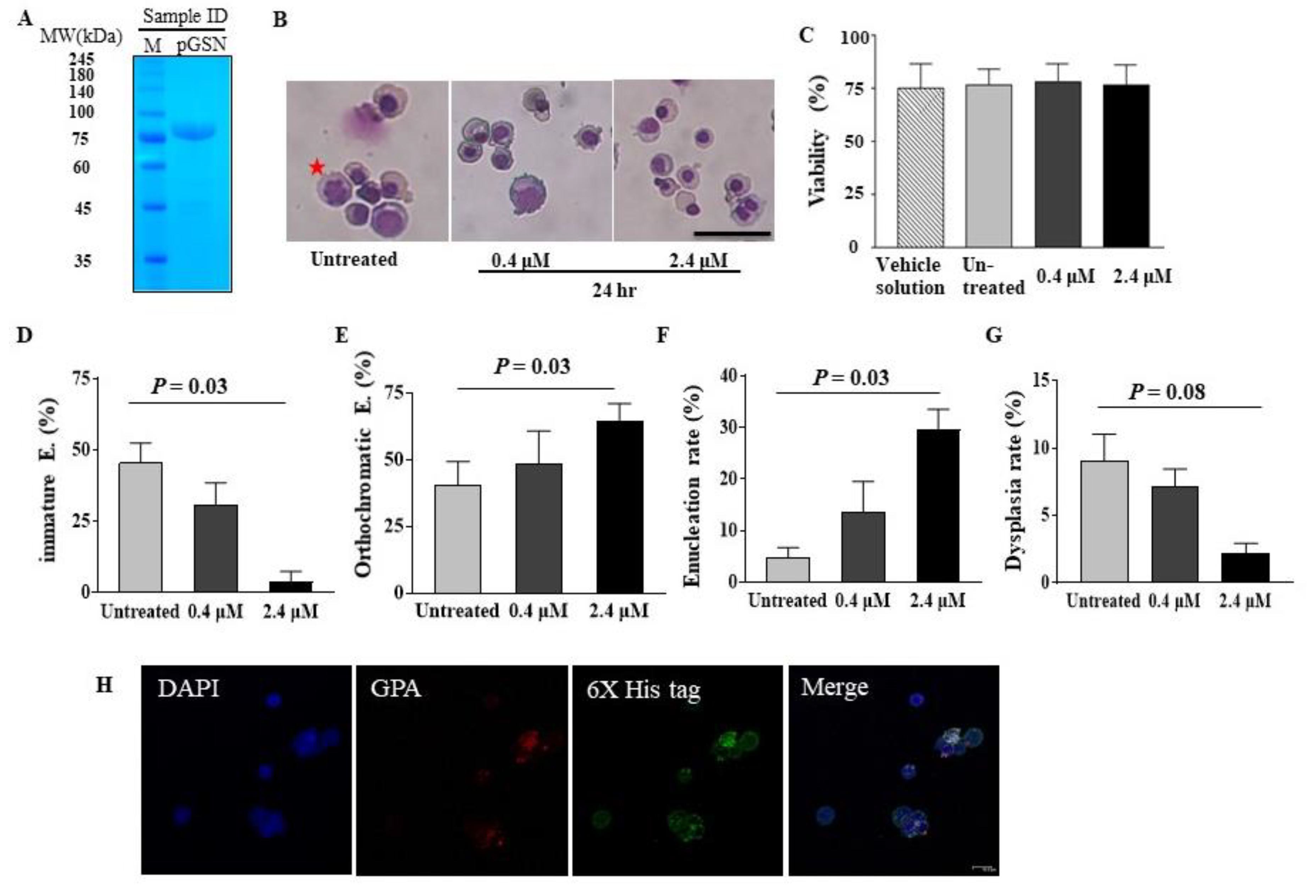
2.1. pGSN Accelerates In Vitro Terminal Erythropoiesis and Corrects Erythroid Dysplasia
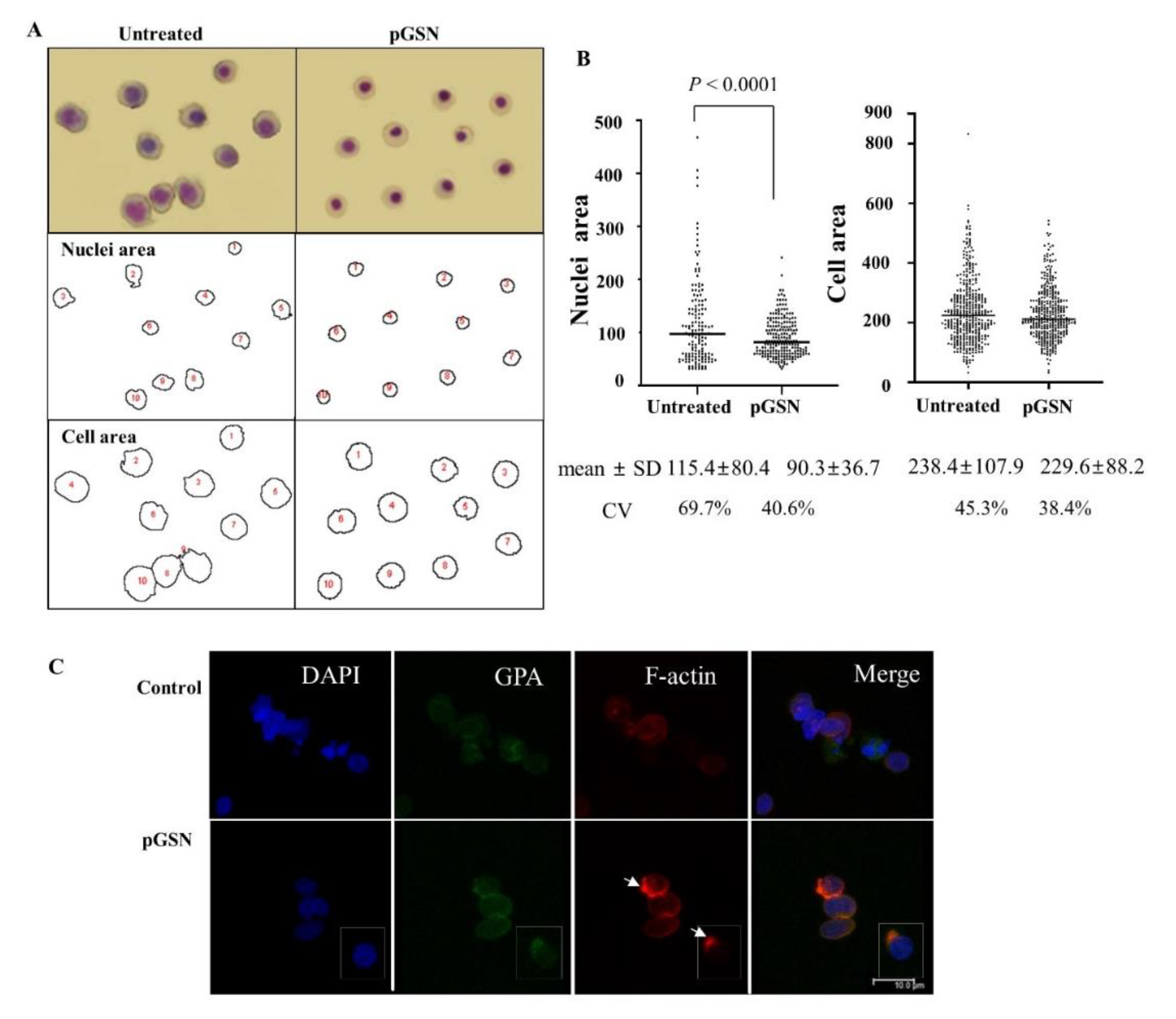
2.2. Effects of pGSN on Cell Size, Nuclear Size, Actin Skeleton, and Dysplasia of Late Stage Erythroblasts
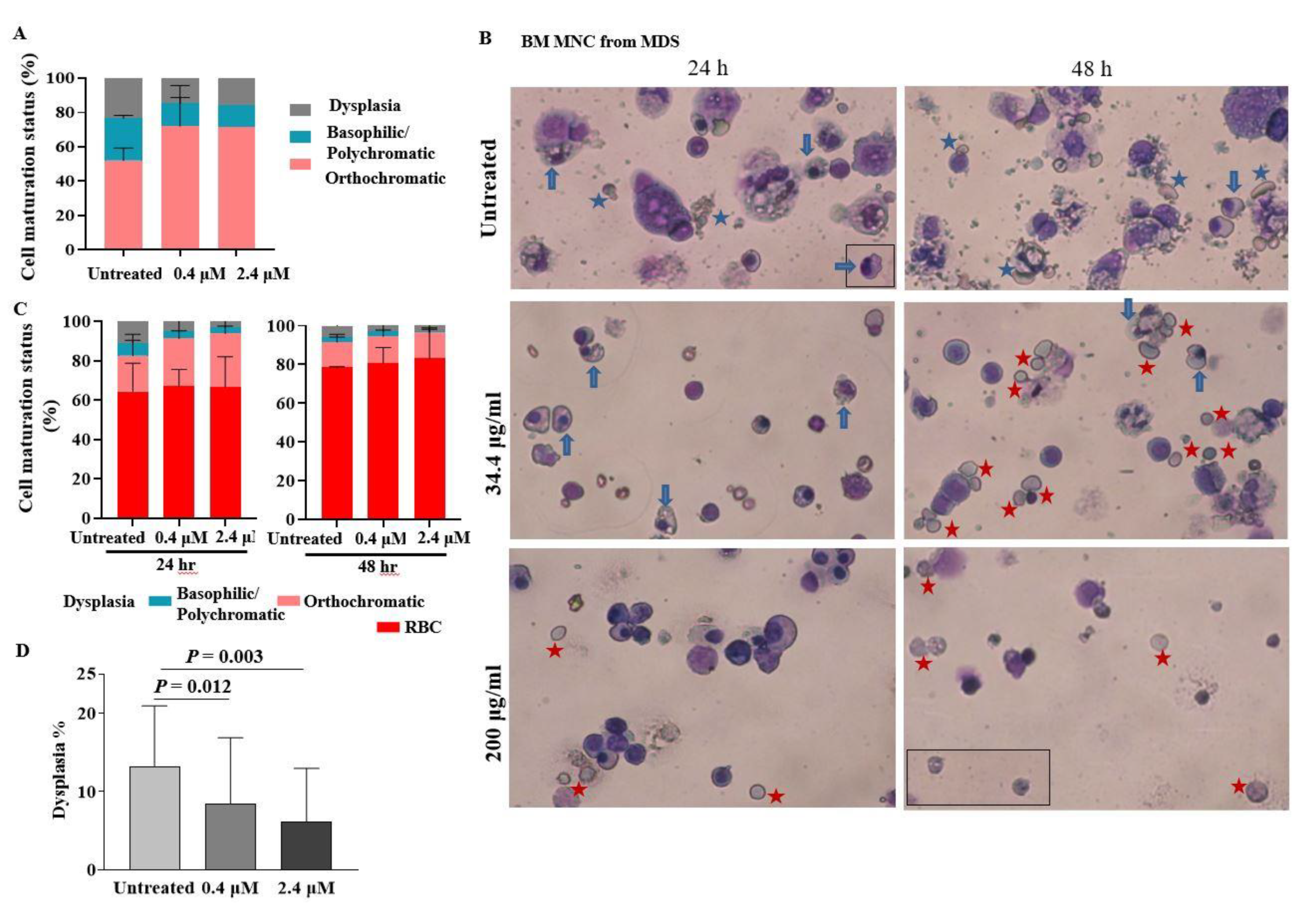
2.3. pGSN Enhances In Vitro Terminal Maturation of Erythroblasts from MDS Patients
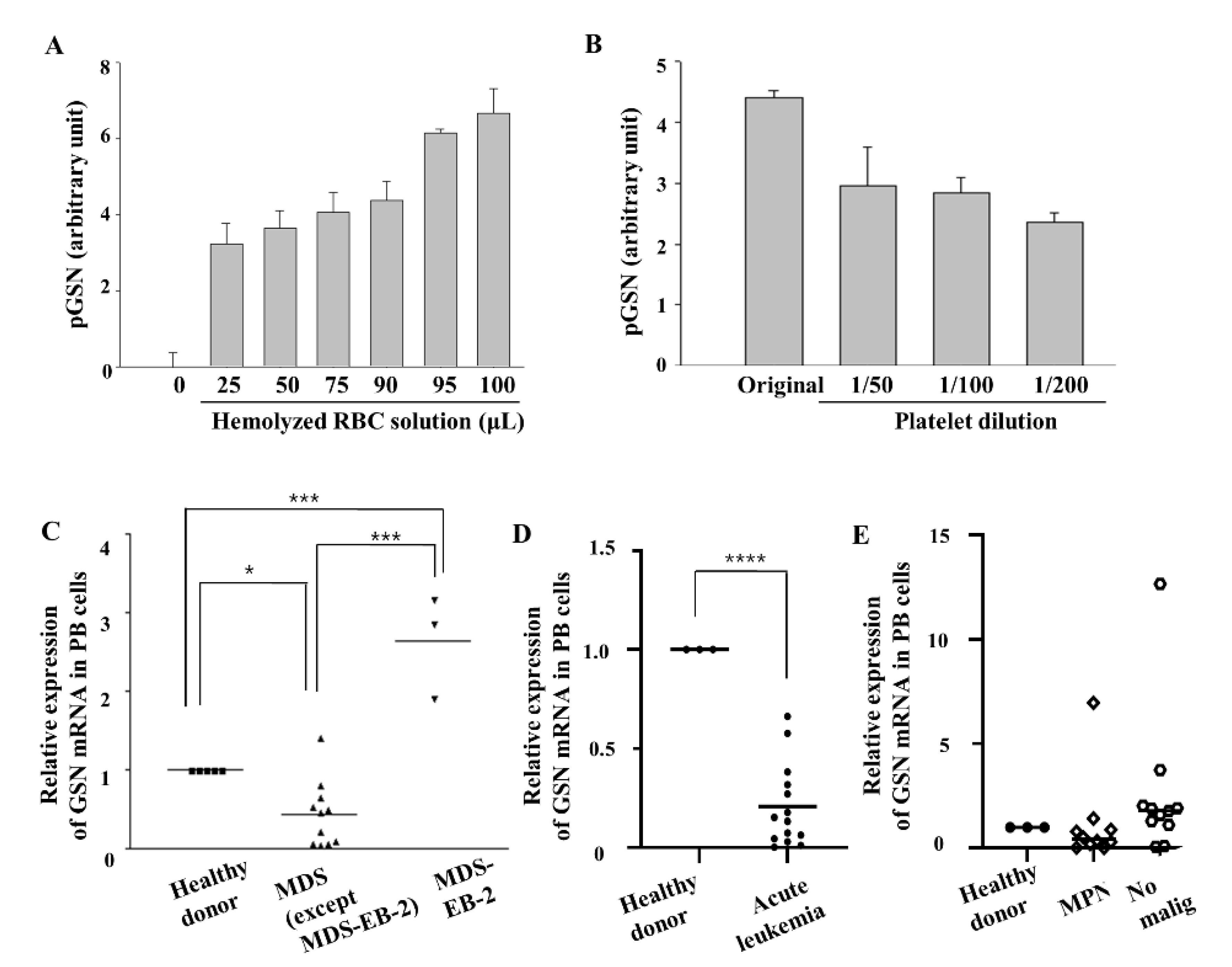
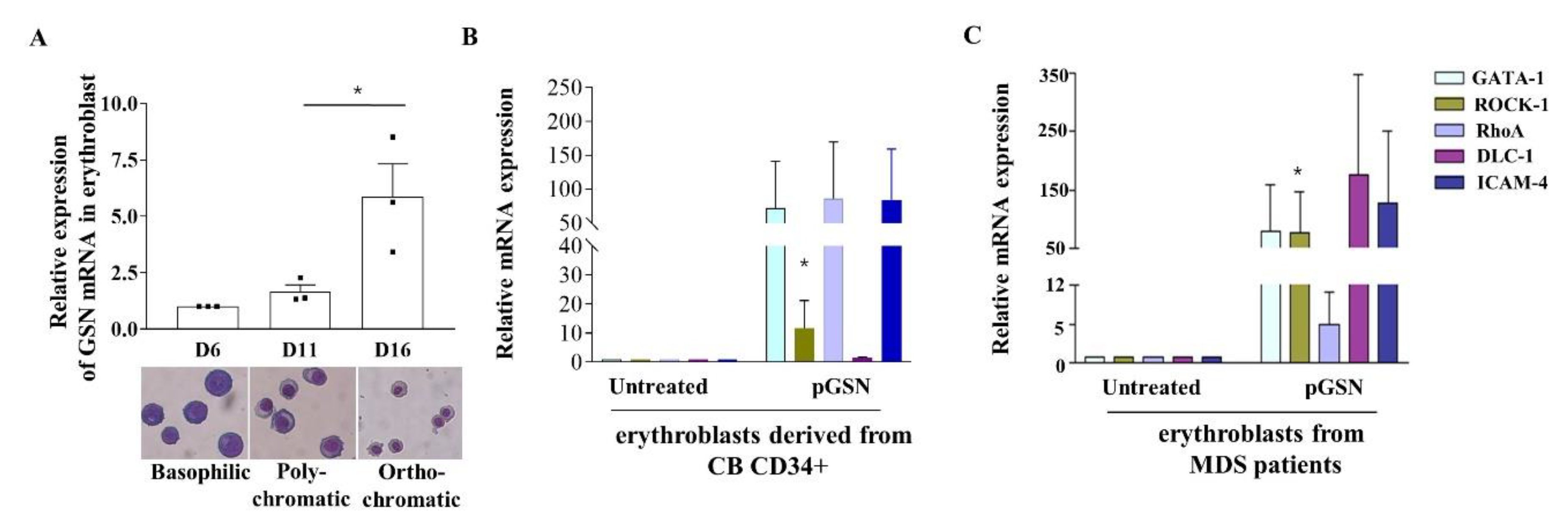
2.4. GSN mRNA Levels Are Low in PB of Patients with MDS
2.5. pGSN Enhances In Vitro Terminal Erythropoiesis of Erythroblasts in MDS Patients
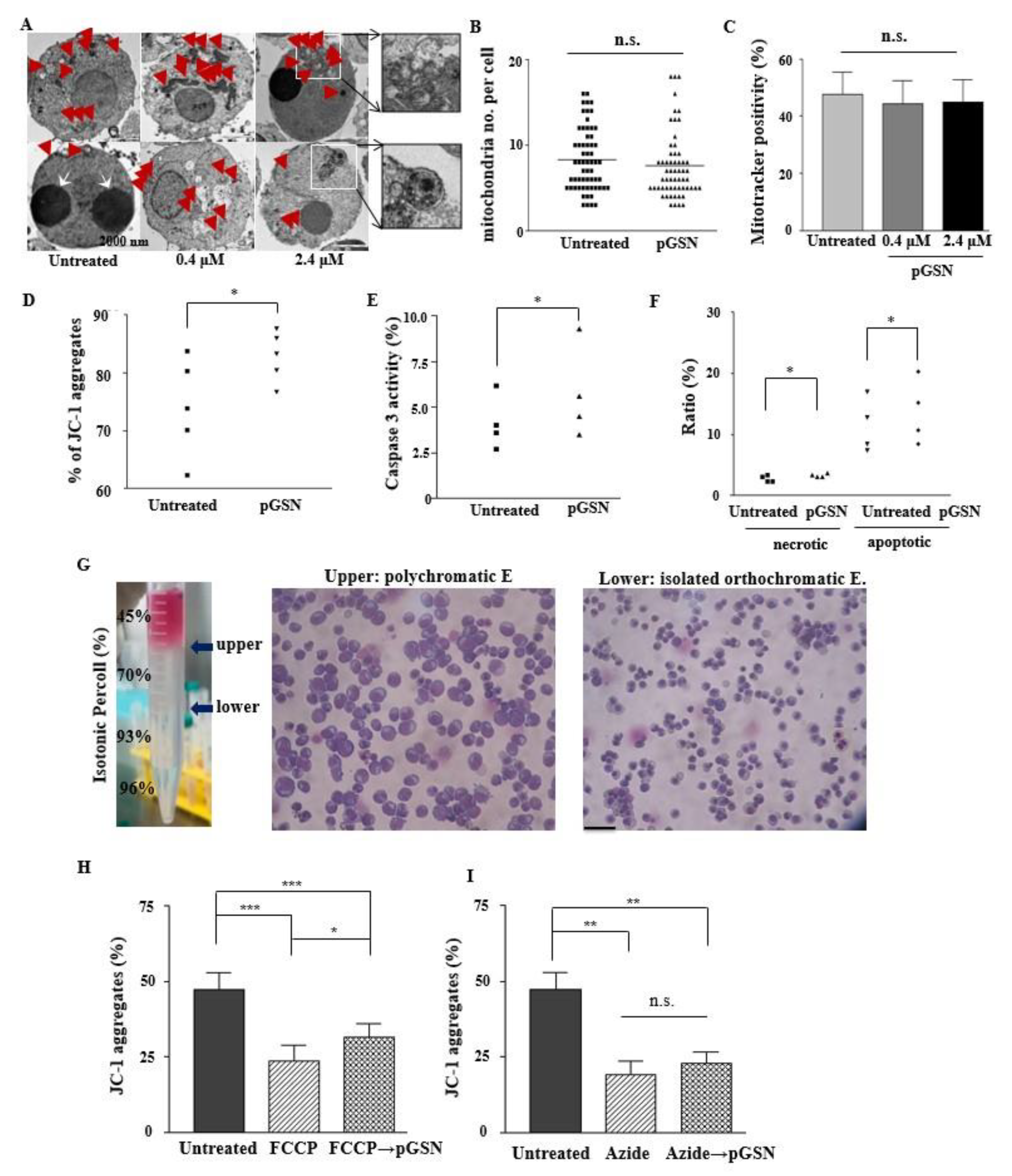
2.6. pGSN Enhances Mitochondrial Polarization in Normal Late Erythroblasts
2.7. pGSN Preserves and Restores MMP
3. Discussion
4. Methods
4.1. Cell Culture and BM Collection
4.2. Preparation and Purification of pGSN
4.3. pGSN Treatment in Culture
4.4. Isolation of Orthochromatic Erythroblasts Using Isotonic Percoll Gradient Methods
4.5. Morphological Analysis
4.6. Transmission Electron Microscopy
4.7. Immunofluorescence Staining
4.8. Measurement of Gelsolin Protein Levels Affected by RBC Hemolysis and Platelet Contamination
4.9. Enzyme-Linked Immunosorbent Assay (ELISA)
4.10. Real-Time Quantitative Polymerase Chain Reaction (RT-qPCR)
4.11. HPLC ESI-LC/MS
4.12. Mitochondrial Transmembrane Potential Inhibition
4.13. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Lapillonne, H.; Kobari, L.; Mazurier, C.; Tropel, P.; Giarratana, M.C.; Zanella-Cleon, I.; Kiger, L.; Wattenhofer-Donze, M.; Puccio, H.; Hebert, N.; et al. Red blood cell generation from human induced pluripotent stem cells: Perspectives for transfusion medicine. Haematologica 2010, 95, 1651–1659. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dorn, I.; Lazar-Karsten, P.; Boie, S.; Ribbat, J.; Hartwig, D.; Driller, B.; Kirchner, H.; Schlenke, P. In vitro proliferation and differentiation of human CD34+ cells from peripheral blood into mature red blood cells with two different cell culture systems. Transfusion 2008, 48, 1122–1132. [Google Scholar] [CrossRef] [PubMed]
- Khodabux, C.M.; van Hensbergen, Y.; Slot, M.C.; Bakker-Verweij, M.; Giordano, P.C.; Brand, A. Exploring the use of expanded erythroid cells for autologous transfusion for anemia of prematurity. Transfusion 2013, 53, 3230–3239. [Google Scholar] [CrossRef] [PubMed]
- Trakarnsanga, K.; Wilson, M.C.; Griffiths, R.E.; Toye, A.M.; Carpenter, L.; Heesom, K.J.; Parsons, S.F.; Anstee, D.J.; Frayne, J. Qualitative and quantitative comparison of the proteome of erythroid cells differentiated from human iPSCs and adult erythroid cells by multiplex TMT labelling and nanoLC-MS/MS. PLoS ONE 2014, 9, e100874. [Google Scholar] [CrossRef]
- Koury, S.T.; Koury, M.J.; Bondurant, M.C. Cytoskeletal distribution and function during the maturation and enucleation of mammalian erythroblasts. J. Cell Biol. 1989, 109, 3005–3013. [Google Scholar] [CrossRef] [Green Version]
- Houwerzijl, E.J.; Pol, H.W.; Blom, N.R.; van der Want, J.J.; de Wolf, J.T.; Vellenga, E. Erythroid precursors from patients with low-risk myelodysplasia demonstrate ultrastructural features of enhanced autophagy of mitochondria. Leukemia 2009, 23, 886–891. [Google Scholar] [CrossRef]
- Greenberg, P.L.; Young, N.S.; Gattermann, N. Myelodysplastic syndromes. Hematol. Am. Soc. Hematol. Educ. Program. 2002, 136–161. [Google Scholar] [CrossRef] [Green Version]
- Burtnick, L.D.; Koepf, E.K.; Grimes, J.; Jones, E.Y.; Stuart, D.I.; McLaughlin, P.J.; Robinson, R.C. The crystal structure of plasma gelsolin: Implications for actin severing, capping, and nucleation. Cell 1997, 90, 661–670. [Google Scholar] [CrossRef] [Green Version]
- Spinardi, L.; Witke, W. Gelsolin and diseases. In Calcium Signalling and Disease; Springer: Berlin/Heidelberg, Germany, 2007; pp. 55–69. [Google Scholar]
- Kwiatkowski, D.J.; Stossel, T.P.; Orkin, S.H.; Mole, J.E.; Coltens, H.R.; Yin, H.L. Plasma and cytoplasmic gelsolins are encoded by a single gene and contain a duplicated actin-binding domain. Nature 1986, 323, 455. [Google Scholar] [CrossRef]
- Cooper, J.A.; Bryan, J.; Schwab, B.; Frieden, C.; Loftus, D.J.; Elson, E.L. Microinjection of gelsolin into living cells. J. Cell Biol. 1987, 104, 491–501. [Google Scholar] [CrossRef] [Green Version]
- Jagadish, T.; Pottiez, G.; Fox, H.S.; Ciborowski, P. Plasma gelsolin accumulates in macrophage nodules in brains of simian immunodeficiency virus infected rhesus macaques. J. Neurovirol. 2012, 18, 113–119. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bucki, R.; Byfield, F.J.; Kulakowska, A.; McCormick, M.E.; Drozdowski, W.; Namiot, Z.; Hartung, T.; Janmey, P.A. Extracellular gelsolin binds lipoteichoic acid and modulates cellular response to proinflammatory bacterial wall components. J. Immunol. 2008, 181, 4936–4944. [Google Scholar] [CrossRef] [Green Version]
- Kwiatkowski, D.J. Functions of gelsolin: Motility, signaling, apoptosis, cancer. Curr. Opin. Cell Biol. 1999, 11, 103–108. [Google Scholar] [CrossRef]
- Hinssen, H.; Vandekerckhove, J.; Lazarides, E. Gelsolin is expressed in early erythroid progenitor cells and negatively regulated during erythropoiesis. J. Cell Biol. 1987, 105, 1425–1433. [Google Scholar] [CrossRef]
- Han, S.Y.; Lee, E.M.; Choi, H.S.; Chun, B.H.; Baek, E.J. The effects of plasma gelsolin on human erythroblast maturation for erythrocyte production. Stem Cell Res. 2018, 29, 64–75. [Google Scholar] [CrossRef] [PubMed]
- Keerthivasan, G.; Wickrema, A.; Crispino, J.D. Erythroblast enucleation. Stem Cell Int. 2011, 2011, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Li, J. Myelodysplastic syndrome hematopoietic stem cell. Int. J. Cancer 2013, 133, 525–533. [Google Scholar] [CrossRef]
- Sundaravel, S.; Duggan, R.; Bhagat, T.; Ebenezer, D.L.; Liu, H.; Yu, Y.; Bartenstein, M.; Unnikrishnan, M.; Karmakar, S.; Liu, T.C.; et al. Reduced DOCK4 expression leads to erythroid dysplasia in myelodysplastic syndromes. Proc. Natl. Acad. Sci. USA 2015, 112, E6359–E6368. [Google Scholar] [CrossRef] [Green Version]
- Chiang, C.L.; Chen, S.S.; Lee, S.J.; Tsao, K.C.; Chu, P.L.; Wen, C.H.; Hwang, S.M.; Yao, C.L.; Lee, H. Lysophosphatidic acid induces erythropoiesis through activating lysophosphatidic acid receptor 3. Stem Cells 2011, 29, 1763–1773. [Google Scholar] [CrossRef]
- Lin, K.H.; Ho, Y.H.; Chiang, J.C.; Li, M.W.; Lin, S.H.; Chen, W.M.; Chiang, C.L.; Lin, Y.N.; Yang, Y.J.; Chen, C.N.; et al. Pharmacological activation of lysophosphatidic acid receptors regulates erythropoiesis. Sci. Rep. 2016, 6, 27050. [Google Scholar] [CrossRef]
- Gourlay, C.W.; Ayscough, K.R. The actin cytoskeleton: A key regulator of apoptosis and ageing? Nat. Rev. Mol. Cell Biol. 2005, 6, 583–589. [Google Scholar] [CrossRef] [PubMed]
- Pellagatti, A.; Cazzola, M.; Giagounidis, A.; Perry, J.; Malcovati, L.; Della Porta, M.G.; Jädersten, M.; Killick, S.; Verma, A.; Norbury, C.J.; et al. Deregulated gene expression pathways in myelodysplastic syndrome hematopoietic stem cells. Leukemia 2010, 24, 756–764. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pellagatti, A.; Benner, A.; Mills, K.I.; Cazzola, M.; Giagounidis, A.; Perry, J.; Malcovati, L.; Della Porta, M.G.; Jadersten, M.; Verma, A.; et al. Identification of gene expression-based prognostic markers in the hematopoietic stem cells of patients with myelodysplastic syndromes. J. Clin. Oncol. 2013, 31, 3557–3564. [Google Scholar] [CrossRef] [PubMed]
- de Thonel, A.; Vandekerckhove, J.; Lanneau, D.; Selvakumar, S.; Courtois, G.; Hazoume, A.; Brunet, M.; Maurel, S.; Hammann, A.; Ribeil, J.A.; et al. HSP27 controls GATA-1 protein level during erythroid cell differentiation. Blood 2010, 116, 85–96. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gabet, A.S.; Coulon, S.; Fricot, A.; Vandekerckhove, J.; Chang, Y.; Ribeil, J.A.; Lordier, L.; Zermati, Y.; Asnafi, V.; Belaid, Z.; et al. Caspase-activated ROCK-1 allows erythroblast terminal maturation independently of cytokine-induced Rho signaling. Cell Death Differ. 2011, 18, 678–689. [Google Scholar] [CrossRef] [PubMed]
- Konstantinidis, D.G.; Giger, K.M.; Risinger, M.; Pushkaran, S.; Zhou, P.; Dexheimer, P.; Yerneni, S.; Andreassen, P.; Klingmuller, U.; Palis, J.; et al. Cytokinesis failure in RhoA-deficient mouse erythroblasts involves actomyosin and midbody dysregulation and triggers p53 activation. Blood 2015, 126, 1473–1482. [Google Scholar] [CrossRef] [Green Version]
- Kalfa, T.A.; Zheng, Y. Rho GTPases in erythroid maturation. Curr. Opin. Hematol. 2014, 21, 165–171. [Google Scholar] [CrossRef] [Green Version]
- Hall, A. Rho GTPases and the actin cytoskeleton. Science 1998, 279, 509–514. [Google Scholar] [CrossRef] [Green Version]
- Liu, T.; Hannafon, B.; Gill, L.; Kelly, W.; Benbrook, D. Flex-Hets differentially induce apoptosis in cancer over normal cells by directly targeting mitochondria. Mol. Cancer Ther. 2007, 6, 1814–1822. [Google Scholar] [CrossRef] [Green Version]
- Scarlett, J.L.; Sheard, P.W.; Hughes, G.; Ledgerwood, E.C.; Ku, H.H.; Murphy, M.P. Changes in mitochondrial membrane potential during staurosporine-induced apoptosis in Jurkat cells. FEBS Lett. 2000, 475, 267–272. [Google Scholar] [CrossRef] [Green Version]
- Zermati, Y.; Garrido, C.; Amsellem, S.; Fishelson, S.; Bouscary, D.; Valensi, F.; Varet, B.; Solary, E.; Hermine, O. Caspase activation is required for terminal erythroid differentiation. J. Exp. Med. 2001, 193, 247–254. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liang, R.; Ghaffari, S. Mitochondria are implicated in the regulation of terminal erythropoiesis. Exp. Hematol. 2017, 53, S83. [Google Scholar] [CrossRef]
- Liang, R.; Menon, V.; Lin, M.; Hartmann, B.; Nowak, R.; Fowler, V.; Girnun, G.; Ghaffari, S. Mitochondrial Regulation is Essential for Erythroid Nuclear Removal. Exp. Hematol. 2018, 64, S47. [Google Scholar] [CrossRef]
- Yin, H.L.; Stossel, T.P. Control of cytoplasmic actin gel-sol transformation by gelsolin, a calcium-dependent regulatory protein. Nature 1979, 281, 583–586. [Google Scholar] [CrossRef]
- Legrand, C.; Ferraz, C.; Clavel, M.C.; Rabie, A. Immunocytochemical localisation of gelsolin in oligodendroglia of the developing rabbit central nervous system. Brain Res. 1986, 395, 231–235. [Google Scholar] [CrossRef]
- Baek, E.J.; Kim, H.S.; Kim, S.; Jin, H.; Choi, T.Y.; Kim, H.O. In vitro clinical-grade generation of red blood cells from human umbilical cord blood CD34+ cells. Transfusion 2008, 48, 2235–2245. [Google Scholar] [CrossRef]
- Nowak, R.B.; Papoin, J.; Gokhin, D.S.; Casu, C.; Rivella, S.; Lipton, J.M.; Blanc, L.; Fowler, V.M. Tropomodulin 1 controls erythroblast enucleation via regulation of F-actin in the enucleosome. Blood 2017, 130, 1144–1155. [Google Scholar] [CrossRef]
- Leary, S.C.; Hill, B.C.; Lyons, C.N.; Carlson, C.G.; Michaud, D.; Kraft, C.S.; Ko, K.; Glerum, D.M.; Moyes, C.D. Chronic treatment with azide in situ leads to an irreversible loss of cytochrome c oxidase activity via holoenzyme dissociation. J. Biol. Chem. 2002, 277, 11321–11328. [Google Scholar] [CrossRef] [Green Version]
- Wyatt, C.N.; Buckler, K.J. The effect of mitochondrial inhibitors on membrane currents in isolated neonatal rat carotid body type I cells. J. Physiol. 2004, 556, 175–191. [Google Scholar] [CrossRef]
- Huff, J.D.; Keung, Y.K.; Thakuri, M.; Beaty, M.W.; Hurd, D.D.; Owen, J.; Molnar, I. Copper deficiency causes reversible myelodysplasia. Am. J. Hematol. 2007, 82, 625–630. [Google Scholar] [CrossRef]
- Gregg, X.T.; Reddy, V.; Prchal, J.T. Copper deficiency masquerading as myelodysplastic syndrome. Blood 2002, 100, 1493–1495. [Google Scholar] [CrossRef] [PubMed]
- Hellstrom-Lindberg, E.; Kanter-Lewensohn, L.; Ost, A. Morphological changes and apoptosis in bone marrow from patients with myelodysplastic syndromes treated with granulocyte-CSF and erythropoietin. Leuk. Res. 1997, 21, 415–425. [Google Scholar] [CrossRef]
- Parker, J.E.; Mufti, G.J.; Rasool, F.; Mijovic, A.; Devereux, S.; Pagliuca, A. The role of apoptosis, proliferation, and the Bcl-2-related proteins in the myelodysplastic syndromes and acute myeloid leukemia secondary to MDS. Blood 2000, 96, 3932–3938. [Google Scholar] [CrossRef] [PubMed]
- Tsujimoto, Y.; Shimizu, S. The voltage-dependent anion channel: An essential player in apoptosis. Biochimie 2002, 84, 187–193. [Google Scholar] [CrossRef]
- Li, G.H.; Arora, P.D.; Chen, Y.; McCulloch, C.A.; Liu, P. Multifunctional roles of gelsolin in health and diseases. Med. Res. Rev. 2012, 32, 999–1025. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Zhang, H.; Rodriguez, S.; Cao, L.; Parish, J.; Mumaw, C.; Zollman, A.; Kamoka, M.M.; Mu, J.; Chen, D.Z.; et al. Notch-dependent repression of miR-155 in the bone marrow niche regulates hematopoiesis in an NF-kappaB-dependent manner. Cell Stem Cell 2014, 15, 51–65. [Google Scholar] [CrossRef] [Green Version]
- Griffiths, E.A.; Gore, S.D. Epigenetic therapies in MDS and AML. Adv. Exp. Med. Biol. 2013, 754, 253–283. [Google Scholar]
- Wang, H.-C.; Minh, B.Q.; Susko, E.; Roger, A.J. Modeling site heterogeneity with posterior mean site frequency profiles accelerates accurate phylogenomic estimation. Syst. Biol. 2017, 67, 216–235. [Google Scholar] [CrossRef]
- Mielnicki, L.M.; Ying, A.M.; Head, K.L.; Asch, H.L.; Asch, B.B. Epigenetic regulation of gelsolin expression in human breast cancer cells. Exp. Cell Res. 1999, 249, 161–176. [Google Scholar] [CrossRef]
- Hoshikawa, Y.; Kwon, H.J.; Yoshida, M.; Horinouchi, S.; Beppu, T. Trichostatin A induces morphological changes and gelsolin expression by inhibiting histone deacetylase in human carcinoma cell lines. Exp. Cell Res. 1994, 214, 189–197. [Google Scholar] [CrossRef]
- Li, W.; Xie, S.; Qian, L.; Chang, B.; Zou, B.; Zhou, W.; Zhao, R.; Wang, G. Large-scale synthesis of aligned carbon nanotubes. Science 1996, 274, 1701–1703. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, M.; Suda, T.; Takahashi, T.; Nagata, S. Expression of the functional soluble form of human fas ligand in activated lymphocytes. EMBO J. 1995, 14, 1129–1135. [Google Scholar] [CrossRef] [PubMed]
- Evans, C.A.; Tonge, R.; Blinco, D.; Pierce, A.; Shaw, J.; Lu, Y.; Hamzah, H.G.; Gray, A.; Downes, C.P.; Gaskell, S.J.; et al. Comparative proteomics of primitive hematopoietic cell populations reveals differences in expression of proteins regulating motility. Blood 2004, 103, 3751–3759. [Google Scholar] [CrossRef] [PubMed]
- Shin, J.Y.; Hu, W.; Naramura, M.; Park, C.Y. High c-Kit expression identifies hematopoietic stem cells with impaired self-renewal and megakaryocytic bias. J. Exp. Med. 2014, 211, 217–231. [Google Scholar] [CrossRef] [Green Version]
- Bucki, R.; Kulakowska, A.; Byfield, F.J.; Zendzian-Piotrowska, M.; Baranowski, M.; Marzec, M.; Winer, J.P.; Ciccarelli, N.J.; Górski, J.; Drozdowski, W.; et al. Plasma gelsolin modulates cellular response to sphingosine 1-phosphate. Am. J. Physiol. Cell Physiol. 2010, 299, C1516–C1523. [Google Scholar] [CrossRef] [Green Version]
- Yang, C.; Hashimoto, M.; Lin, Q.X.X.; Tan, D.Q.; Suda, T. Sphingosine-1-phosphate signaling modulates terminal erythroid differentiation through the regulation of mitophagy. Exp. Hematol. 2019, 72, 47–59.e41. [Google Scholar] [CrossRef] [Green Version]
- Brendt, P.; Rehfeld, I.; Kamphausen, A.; Kreissig, C.; Peters, J. Lipopolysaccharide interference in erythropoiesis in mice. Anaesthesia 2012, 67, 493–500. [Google Scholar] [CrossRef]
- Osborn, T.M.; Dahlgren, C.; Hartwig, J.H.; Stossel, T.P. Modifications of cellular responses to lysophosphatidic acid and platelet-activating factor by plasma gelsolin. Am. J. Physiol. Cell Physiol. 2007, 292, C1323–C1330. [Google Scholar] [CrossRef] [Green Version]
- Baek, E.J.; Kim, H.S.; Kim, J.H.; Kim, N.J.; Kim, H.O. Stroma-free mass production of clinical-grade red blood cells (RBCs) by using poloxamer 188 as an RBC survival enhancer. Transfusion 2009, 49, 2285–2295. [Google Scholar] [CrossRef]
- Lee, E.; Han, S.Y.; Choi, H.S.; Chun, B.; Hwang, B.; Baek, E.J. Red blood cell generation by three-dimensional aggregate cultivation of late erythroblasts. Tissue Eng. Part. A 2015, 21, 817–828. [Google Scholar] [CrossRef]
- Choi, H.S.; Lee, E.M.; Kim, H.O.; Park, M.I.; Baek, E.J. Autonomous control of terminal erythropoiesis via physical interactions among erythroid cells. Stem Cell Res. 2013, 10, 442–453. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koya, R.C.; Fujita, H.; Shimizu, S.; Ohtsu, M.; Takimoto, M.; Tsujimoto, Y.; Kuzumaki, N. Gelsolin inhibits apoptosis by blocking mitochondrial membrane potential loss and cytochrome c release. J. Biol. Chem. 2000, 275, 15343–15349. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peddada, N.; Sagar, A.; Ashish; Garg, R. Plasma gelsolin: A general prognostic marker of health. Med. Hypotheses 2012, 78, 203–210. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Diagnosis | No. of Patients |
|---|---|
| Leukemia Acute myeloid leukemia Acute lymphoblastic leukemia Adult T-cell leukemia T-cell prolymphocytic leukemia Myeloproliferative neoplasm (MPN) | 10 2 1 1 |
| Chronic myeloid leukemia (CML) | 4 |
| Essential thrombocythemia | 2 |
| Polycythemia vera | 1 |
| No hematopoietic malignancy | |
| Aplastic anemia (AA) | 4 |
| Thrombotic thrombocytopenia purpura (TTP) | 3 |
| Immune thrombocytopenia purpura (ITP) | 4 |
| Case No. | Age (Year) | Sex | Diagnosis | Isolated Sample |
|---|---|---|---|---|
| 1 | 55 | M | MDS with multi lineage dysplasia | Buffy coat |
| 2 | 78 | F | MDS-EB-2 | Buffy coat |
| 3 | 70 | M | MDS with multi lineage dysplasia | Mononuclear cells |
| 4 | 70 | M | MDS-EB-1 | Mononuclear cells |
| 5 | 76 | M | MDS-EB-1 | Mononuclear cells |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Han, S.Y.; Lee, E.M.; Kim, S.; Kwon, A.M.; Baek, E.J. Role of Plasma Gelsolin Protein in the Final Stage of Erythropoiesis and in Correction of Erythroid Dysplasia In Vitro. Int. J. Mol. Sci. 2020, 21, 7132. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21197132
Han SY, Lee EM, Kim S, Kwon AM, Baek EJ. Role of Plasma Gelsolin Protein in the Final Stage of Erythropoiesis and in Correction of Erythroid Dysplasia In Vitro. International Journal of Molecular Sciences. 2020; 21(19):7132. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21197132
Chicago/Turabian StyleHan, So Yeon, Eun Mi Lee, Suyeon Kim, Amy M. Kwon, and Eun Jung Baek. 2020. "Role of Plasma Gelsolin Protein in the Final Stage of Erythropoiesis and in Correction of Erythroid Dysplasia In Vitro" International Journal of Molecular Sciences 21, no. 19: 7132. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21197132