The Role of SHIP1 on Apoptosis and Autophagy in the Adipose Tissue of Obese Mice


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
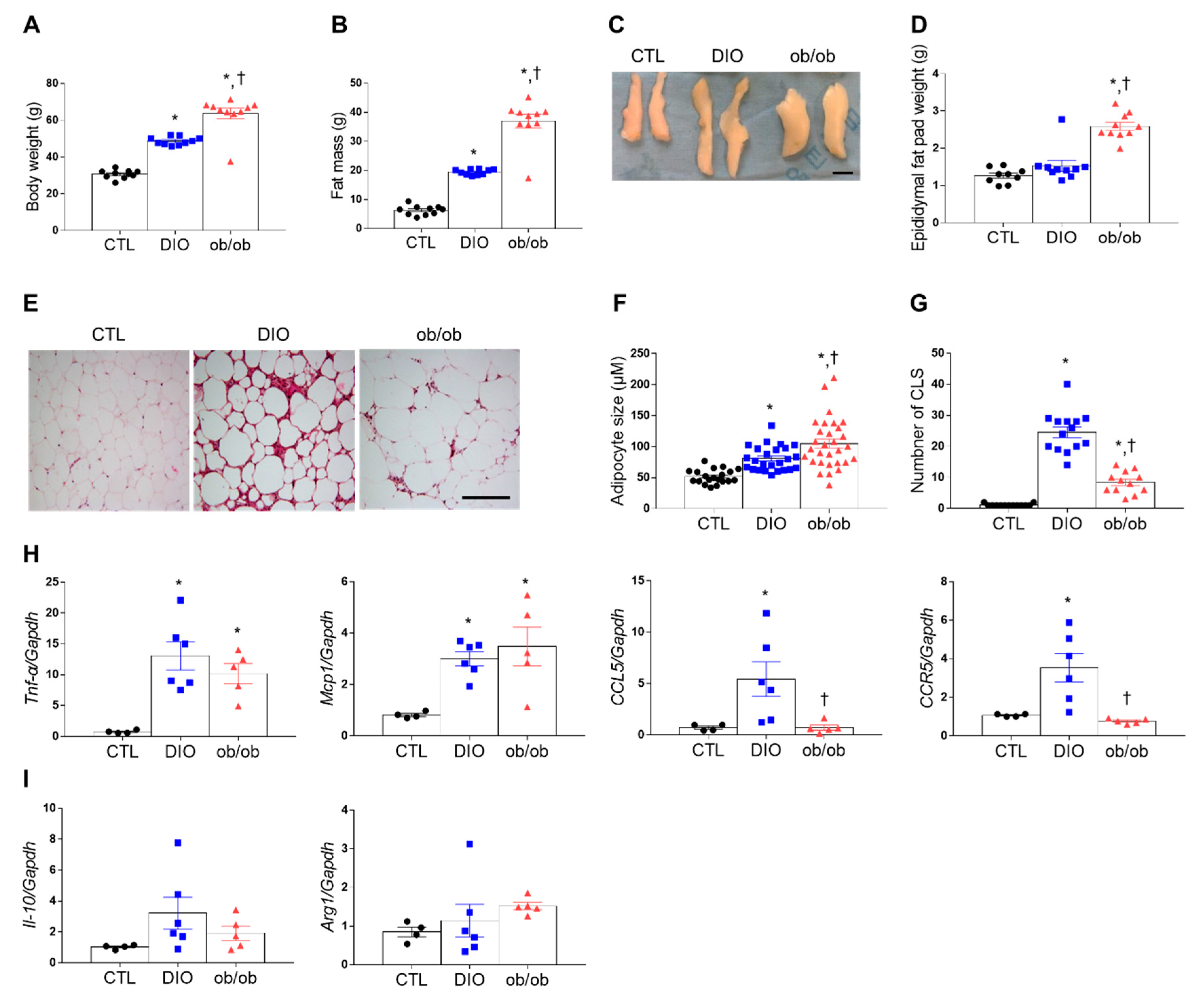
2.1. Macrophage Infiltration and Inflammation in the Adipose Tissue of DIO and ob/ob Mice
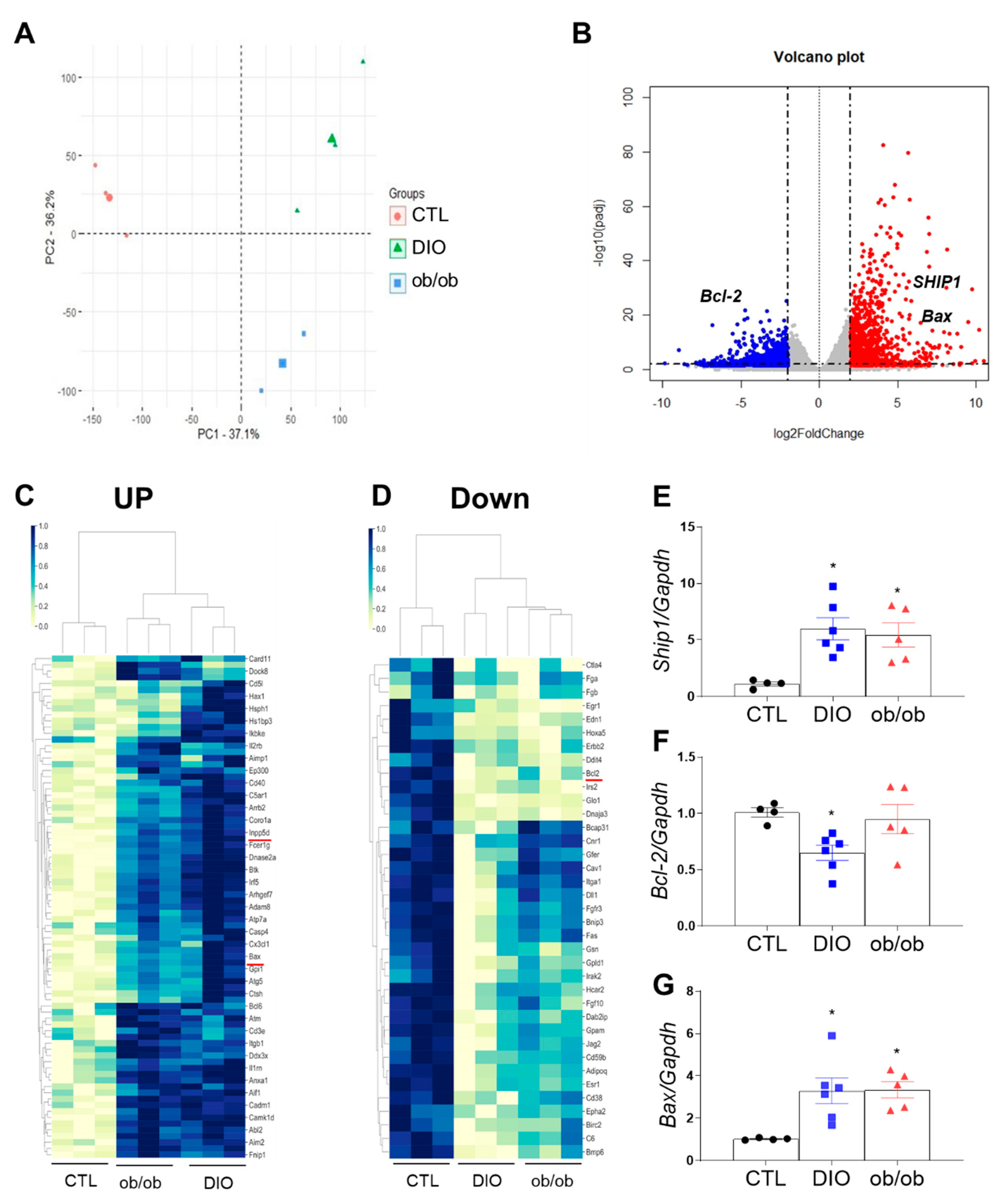
2.2. Differential Gene Expression in the Adipose Tissue of DIO and ob/ob Mice
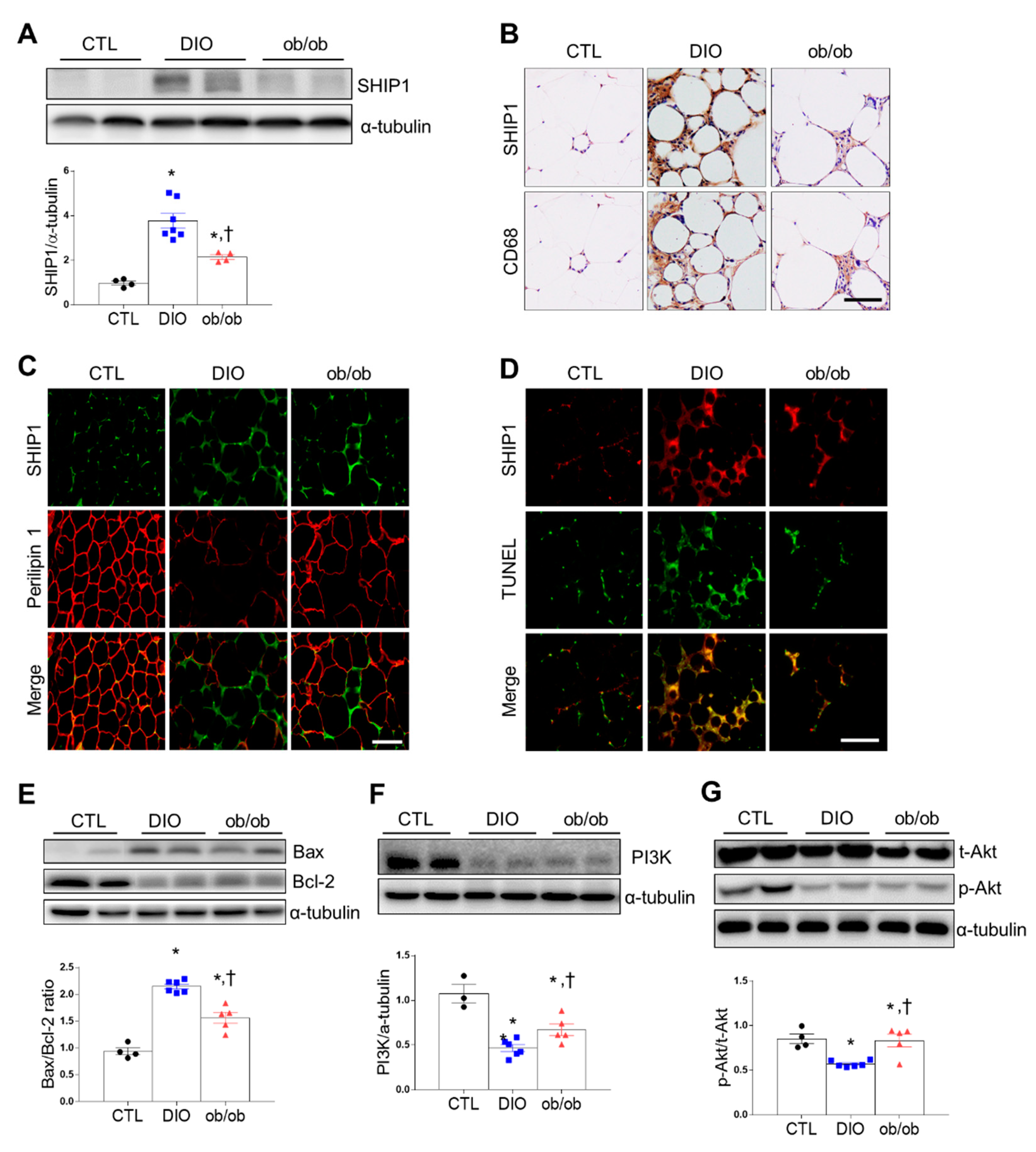
2.3. Effects of DIO and Leptin Deficiency on Inflammation and Apoptosis in Adipose Tissue
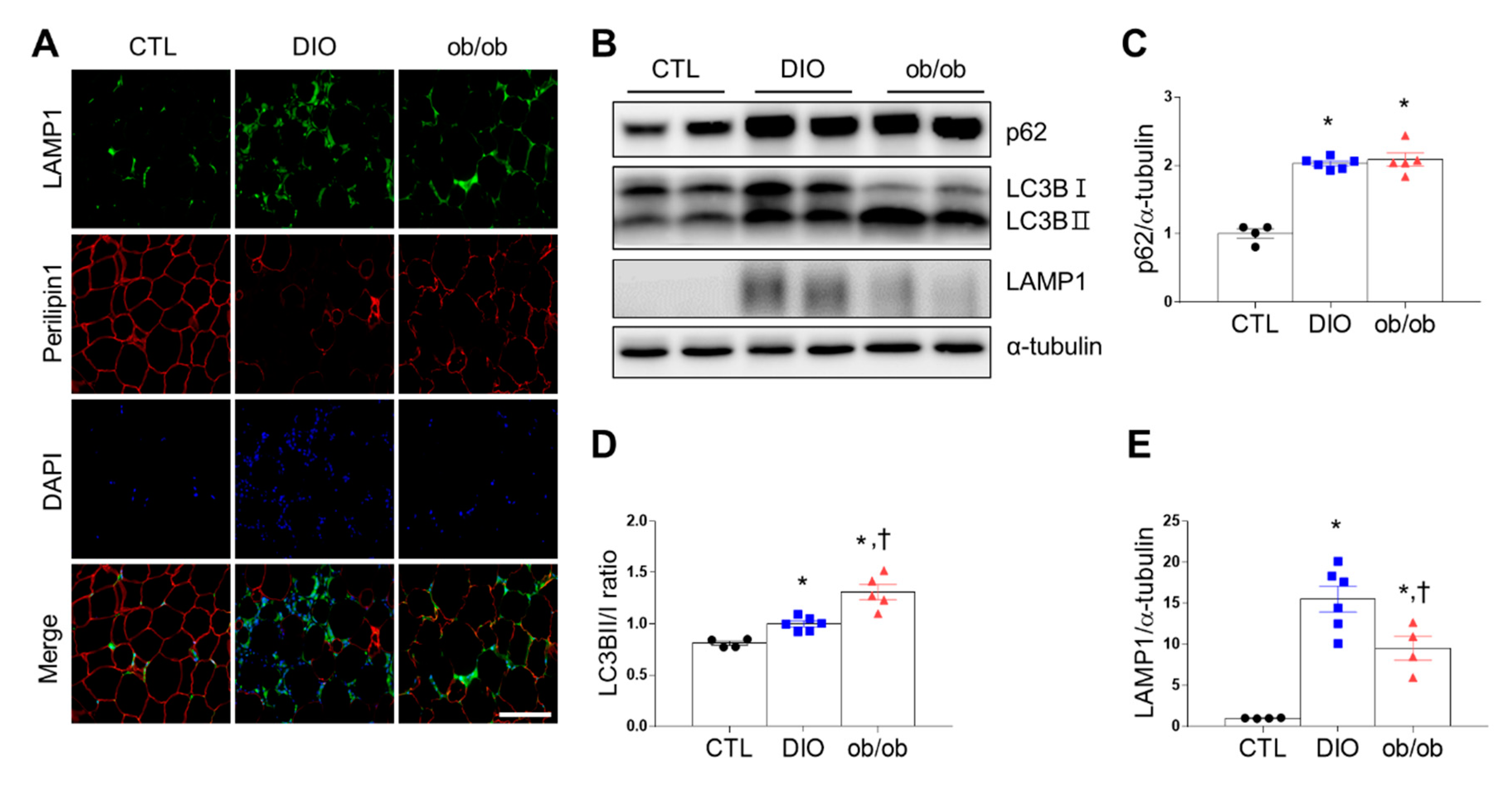
2.4. Effects of DIO and Leptin Deficiency on Autophagy in Adipose Tissue
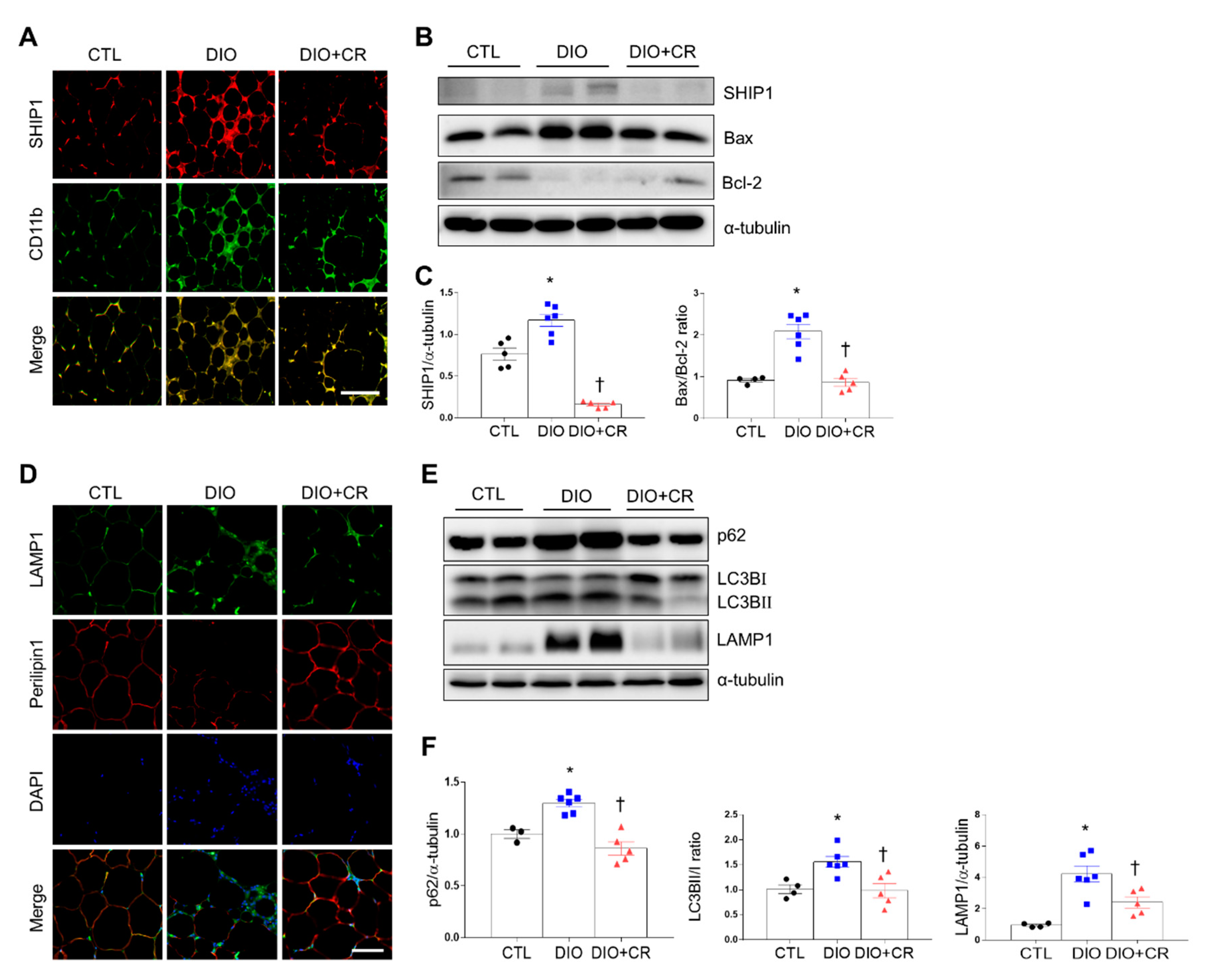
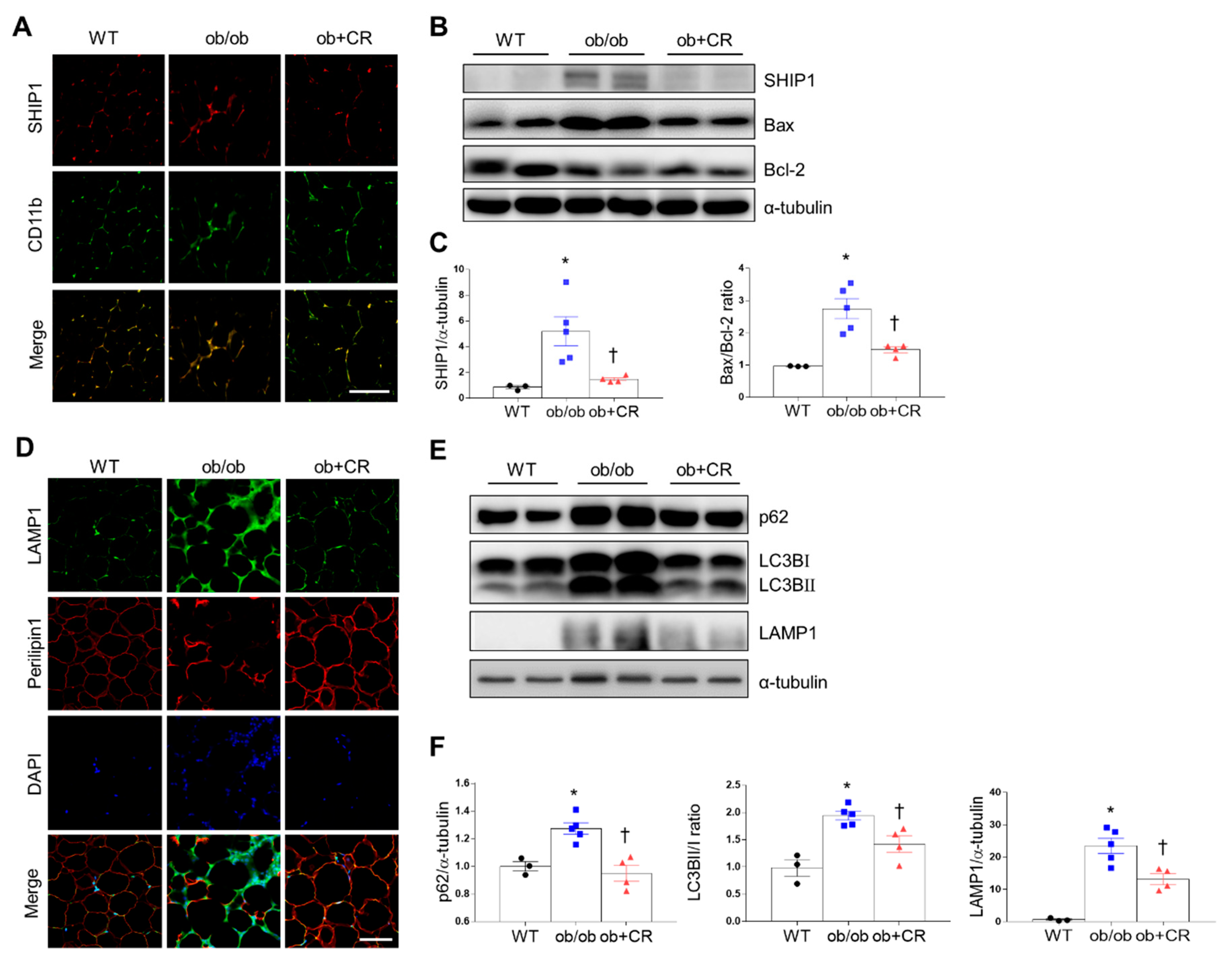
2.5. Effects of CR on SHIP1 Expression Related to Apoptosis and Autophagy in the Adipose Tissue of DIO and ob/ob Mice
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. Enzyme-Linked Immunosorbent Assay (ELISA)
4.3. RNA-seq Analysis
4.4. Real-time Reverse Transcriptase Polymerase Chain Reaction
4.5. Histological Analysis
4.6. Protein Extractions and Western Blot Analyses
4.7. Statistical Analyses
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| CCR5 | C-C motif receptor 5 |
| CCL5 | CC chemokine ligand 5 |
| CLS | Crown-like structure |
| CR | Calorie restriction |
| DAPI | 4′,6-Diamidino-2-phenylindole |
| DIO | Diet-induced obesity |
| HFD | High-fat diet |
| LAMP1 | Lysosomal-associated membrane protein 1 |
| PI3K | Phosphoinositide 3-kinase |
| SHIP1 | Src homology domain-containing inositol 5′-phosphatase 1 |
| TUNEL | Terminal deoxynucleotidyl transferase dUTP nick-end labeling |
References
- Schenk, S.; Saberi, M.; Olefsky, J.M. Insulin sensitivity: Modulation by nutrients and inflammation. J. Clin. Investig. 2008, 118, 2992–3002. [Google Scholar] [CrossRef] [Green Version]
- Lee, B.C.; Lee, J. Cellular and molecular players in adipose tissue inflammation in the development of obesity-induced insulin resistance. Biochim. Biophys. Acta 2014, 1842, 446–462. [Google Scholar] [CrossRef] [Green Version]
- Gordon, S. Alternative activation of macrophages. Nat. Rev. Immunol. 2003, 3, 23–35. [Google Scholar] [CrossRef]
- Lumeng, C.N.; Bodzin, J.L.; Saltiel, A.R. Obesity induces a phenotypic switch in adipose tissue macrophage polarization. J. Clin. Investig. 2007, 117, 175–184. [Google Scholar] [CrossRef] [Green Version]
- Weisberg, S.P.; McCann, D.; Desai, M.; Rosenbaum, M.; Leibel, R.L.; Ferrante, A.W., Jr. Obesity is associated with macrophage accumulation in adipose tissue. J. Clin. Investig. 2003, 112, 1796–1808. [Google Scholar] [CrossRef]
- Kang, Y.H.; Cho, M.H.; Kim, J.Y.; Kwon, M.S.; Peak, J.J.; Kang, S.W.; Yoon, S.Y.; Song, Y. Impaired macrophage autophagy induces systemic insulin resistance in obesity. Oncotarget 2016, 7, 35577–35591. [Google Scholar] [CrossRef] [Green Version]
- Martinez, F.O. Regulators of macrophage activation. Eur. J. Immunol. 2011, 41, 1531–1534. [Google Scholar] [CrossRef] [PubMed]
- Geier, S.J.; Algate, P.A.; Carlberg, K.; Flowers, D.; Friedman, C.; Trask, B.; Rohrschneider, L.R. The human SHIP gene is differentially expressed in cell lineages of the bone marrow and blood. Blood 1997, 89, 1876–1885. [Google Scholar] [CrossRef] [PubMed]
- Ware, M.D.; Rosten, P.; Damen, J.E.; Liu, L.; Humphries, R.K.; Krystal, G. Cloning and characterization of human SHIP, the 145-kD inositol 5-phosphatase that associates with SHC after cytokine stimulation. Blood 1996, 88, 2833–2840. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rauh, M.J.; Ho, V.; Pereira, C.; Sham, A.; Sly, L.M.; Lam, V.; Huxham, L.; Minchinton, A.I.; Mui, A.; Krystal, G. SHIP represses the generation of alternatively activated macrophages. Immunity 2005, 23, 361–374. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kamen, L.A.; Levinsohn, J.; Cadwallader, A.; Tridandapani, S.; Swanson, J.A. SHIP-1 increases early oxidative burst and regulates phagosome maturation in macrophages. J. Immunol. 2008, 180, 7497–7505. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vachharajani, V.; Granger, D.N. Adipose tissue: A motor for the inflammation associated with obesity. IUBMB Life 2009, 61, 424–430. [Google Scholar] [CrossRef] [Green Version]
- Herold, C.; Rennekampff, H.O.; Engeli, S. Apoptotic pathways in adipose tissue. Apoptosis 2013, 18, 911–916. [Google Scholar] [CrossRef] [PubMed]
- Cinti, S.; Mitchell, G.; Barbatelli, G.; Murano, I.; Ceresi, E.; Faloia, E.; Wang, S.; Fortier, M.; Greenberg, A.S.; Obin, M.S. Adipocyte death defines macrophage localization and function in adipose tissue of obese mice and humans. J. Lipid Res. 2005, 46, 2347–2355. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matsuzawa-Ishimoto, Y.; Hwang, S.; Cadwell, K. Autophagy and Inflammation. Annu. Rev. Immunol. 2018, 36, 73–101. [Google Scholar] [CrossRef] [PubMed]
- Park, K.A.; Jin, Z.; An, H.S.; Lee, J.Y.; Jeong, E.A.; Choi, E.B.; Kim, K.E.; Shin, H.J.; Lee, J.E.; Roh, G.S. Effects of caloric restriction on the expression of lipocalin-2 and its receptor in the brown adipose tissue of high-fat diet-fed mice. Korean J. Physiol. Pharmacol. 2019, 23, 335–344. [Google Scholar] [CrossRef] [Green Version]
- An, H.S.; Lee, J.Y.; Choi, E.B.; Jeong, E.A.; Shin, H.J.; Kim, K.E.; Park, K.A.; Jin, Z.; Lee, J.E.; Koh, J.S.; et al. Caloric restriction reverses left ventricular hypertrophy through the regulation of cardiac iron homeostasis in impaired leptin signaling mice. Sci. Rep. 2020, 10, 7176. [Google Scholar] [CrossRef]
- Alkhouri, N.; Gornicka, A.; Berk, M.P.; Thapaliya, S.; Dixon, L.J.; Kashyap, S.; Schauer, P.R.; Feldstein, A.E. Adipocyte apoptosis, a link between obesity, insulin resistance, and hepatic steatosis. J. Biol. Chem. 2010, 285, 3428–3438. [Google Scholar] [CrossRef] [Green Version]
- Helgason, C.D.; Damen, J.E.; Rosten, P.; Grewal, R.; Sorensen, P.; Chappel, S.M.; Borowski, A.; Jirik, F.; Krystal, G.; Humphries, R.K. Targeted disruption of SHIP leads to hemopoietic perturbations, lung pathology, and a shortened life span. Genes Dev. 1998, 12, 1610–1620. [Google Scholar] [CrossRef] [Green Version]
- Srivastava, N.; Iyer, S.; Sudan, R.; Youngs, C.; Engelman, R.W.; Howard, K.T.; Russo, C.M.; Chisholm, J.D.; Kerr, W.G. A small-molecule inhibitor of SHIP1 reverses age- and diet-associated obesity and metabolic syndrome. JCI Insight 2016, 1. [Google Scholar] [CrossRef]
- Donato, J., Jr.; Frazão, R.; Elias, C.F. The PI3K signaling pathway mediates the biological effects of leptin. Arq. Bras. Endocrinol. Metabol. 2010, 54, 591–602. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Collins, S.; Martin, T.L.; Surwit, R.S.; Robidoux, J. Genetic vulnerability to diet-induced obesity in the C57BL/6J mouse: Physiological and molecular characteristics. Physiol. Behav. 2004, 81, 243–248. [Google Scholar] [CrossRef] [PubMed]
- Gardai, S.; Whitlock, B.B.; Helgason, C.; Ambruso, D.; Fadok, V.; Bratton, D.; Henson, P.M. Activation of SHIP by NADPH oxidase-stimulated Lyn leads to enhanced apoptosis in neutrophils. J. Biol. Chem. 2002, 277, 5236–5246. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- An, H.; Xu, H.; Zhang, M.; Zhou, J.; Feng, T.; Qian, C.; Qi, R.; Cao, X. Src homology 2 domain-containing inositol-5-phosphatase 1 (SHIP1) negatively regulates TLR4-mediated LPS response primarily through a phosphatase activity- and PI-3K-independent mechanism. Blood 2005, 105, 4685–4692. [Google Scholar] [CrossRef] [PubMed]
- Zhang, T.Y.; Daynes, R.A. Glucocorticoid conditioning of myeloid progenitors enhances TLR4 signaling via negative regulation of the phosphatidylinositol 3-kinase-Akt pathway. J. Immunol. 2007, 178, 2517–2526. [Google Scholar] [CrossRef] [Green Version]
- Cani, P.D.; Amar, J.; Iglesias, M.A.; Poggi, M.; Knauf, C.; Bastelica, D.; Neyrinck, A.M.; Fava, F.; Tuohy, K.M.; Chabo, C.; et al. Metabolic endotoxemia initiates obesity and insulin resistance. Diabetes 2007, 56, 1761–1772. [Google Scholar] [CrossRef] [Green Version]
- Deuter-Reinhard, M.; Apell, G.; Pot, D.; Klippel, A.; Williams, L.T.; Kavanaugh, W.M. SIP/SHIP inhibits Xenopus oocyte maturation induced by insulin and phosphatidylinositol 3-kinase. Mol. Cell Biol. 1997, 17, 2559–2565. [Google Scholar] [CrossRef] [Green Version]
- Liang, W.C.; Wang, Y.; Wan, D.C.; Yeung, V.S.; Waye, M.M. Characterization of miR-210 in 3T3-L1 adipogenesis. J. Cell Biochem. 2013, 114, 2699–2707. [Google Scholar] [CrossRef]
- Tai, E.S.; Ordovas, J.M. The role of perilipin in human obesity and insulin resistance. Curr. Opin. Lipidol. 2007, 18, 152–156. [Google Scholar] [CrossRef]
- Kolak, M.; Westerbacka, J.; Velagapudi, V.R.; Wågsäter, D.; Yetukuri, L.; Makkonen, J.; Rissanen, A.; Häkkinen, A.M.; Lindell, M.; Bergholm, R.; et al. Adipose tissue inflammation and increased ceramide content characterize subjects with high liver fat content independent of obesity. Diabetes 2007, 56, 1960–1968. [Google Scholar] [CrossRef] [Green Version]
- Mizushima, N.; Komatsu, M. Autophagy: Renovation of cells and tissues. Cell 2011, 147, 728–741. [Google Scholar] [CrossRef] [Green Version]
- Kosacka, J.; Kern, M.; Klöting, N.; Paeschke, S.; Rudich, A.; Haim, Y.; Gericke, M.; Serke, H.; Stumvoll, M.; Bechmann, I.; et al. Autophagy in adipose tissue of patients with obesity and type 2 diabetes. Mol. Cell Endocrinol. 2015, 409, 21–32. [Google Scholar] [CrossRef] [PubMed]
- Gan, L.; Liu, Z.; Luo, D.; Ren, Q.; Wu, H.; Li, C.; Sun, C. Reduced Endoplasmic Reticulum Stress-Mediated Autophagy Is Required for Leptin Alleviating Inflammation in Adipose Tissue. Front. Immunol. 2017, 8, 1507. [Google Scholar] [CrossRef] [Green Version]
- Xu, X.; Grijalva, A.; Skowronski, A.; van Eijk, M.; Serlie, M.J.; Ferrante, A.W., Jr. Obesity activates a program of lysosomal-dependent lipid metabolism in adipose tissue macrophages independently of classic activation. Cell Metab. 2013, 18, 816–830. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ju, L.; Han, J.; Zhang, X.; Deng, Y.; Yan, H.; Wang, C.; Li, X.; Chen, S.; Alimujiang, M.; Li, X.; et al. Obesity-associated inflammation triggers an autophagy-lysosomal response in adipocytes and causes degradation of perilipin 1. Cell Death Dis. 2019, 10, 121. [Google Scholar] [CrossRef]
- Kim, H.; Kang, H.; Heo, R.W.; Jeon, B.T.; Yi, C.O.; Shin, H.J.; Kim, J.; Jeong, S.Y.; Kwak, W.; Kim, W.H.; et al. Caloric restriction improves diabetes-induced cognitive deficits by attenuating neurogranin-associated calcium signaling in high-fat diet-fed mice. J. Cereb. Blood Flow. Metab. 2016, 36, 1098–1110. [Google Scholar] [CrossRef] [Green Version]
- Jeon, B.T.; Heo, R.W.; Jeong, E.A.; Yi, C.O.; Lee, J.Y.; Kim, K.E.; Kim, H.; Roh, G.S. Effects of caloric restriction on O-GlcNAcylation, Ca(2+) signaling, and learning impairment in the hippocampus of ob/ob mice. Neurobiol. Aging 2016, 44, 127–137. [Google Scholar] [CrossRef]
- Jeon, B.T.; Kim, K.E.; Heo, R.W.; Shin, H.J.; Yi, C.O.; Hah, Y.S.; Kim, W.H.; Lee, S.I.; Roh, G.S. Myeloid-specific deletion of SIRT1 increases hepatic steatosis and hypothalamic inflammation in mice fed a high-fat diet. Metab. Brain Dis. 2014, 29, 635–643. [Google Scholar] [CrossRef] [PubMed]
- Jeong, E.A.; Jeon, B.T.; Shin, H.J.; Kim, N.; Lee, D.H.; Kim, H.J.; Kang, S.S.; Cho, G.J.; Choi, W.S.; Roh, G.S. Ketogenic diet-induced peroxisome proliferator-activated receptor-gamma activation decreases neuroinflammation in the mouse hippocampus after kainic acid-induced seizures. Exp. Neurol. 2011, 232, 195–202. [Google Scholar] [CrossRef] [PubMed]






© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jeong, J.H.; Choi, E.B.; Jang, H.M.; Ahn, Y.J.; An, H.S.; Lee, J.Y.; Park, G.; Jeong, E.A.; Shin, H.J.; Lee, J.; et al. The Role of SHIP1 on Apoptosis and Autophagy in the Adipose Tissue of Obese Mice. Int. J. Mol. Sci. 2020, 21, 7225. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21197225
Jeong JH, Choi EB, Jang HM, Ahn YJ, An HS, Lee JY, Park G, Jeong EA, Shin HJ, Lee J, et al. The Role of SHIP1 on Apoptosis and Autophagy in the Adipose Tissue of Obese Mice. International Journal of Molecular Sciences. 2020; 21(19):7225. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21197225
Chicago/Turabian StyleJeong, Jae Hun, Eun Bee Choi, Hye Min Jang, Yu Jeong Ahn, Hyeong Seok An, Jong Youl Lee, Gyeongah Park, Eun Ae Jeong, Hyun Joo Shin, Jaewoong Lee, and et al. 2020. "The Role of SHIP1 on Apoptosis and Autophagy in the Adipose Tissue of Obese Mice" International Journal of Molecular Sciences 21, no. 19: 7225. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21197225