Inhibitory Effects of Peptide Lunasin in Colorectal Cancer HCT-116 Cells and Their Tumorsphere-Derived Subpopulation

 ,
,  and
and 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results and Discussion
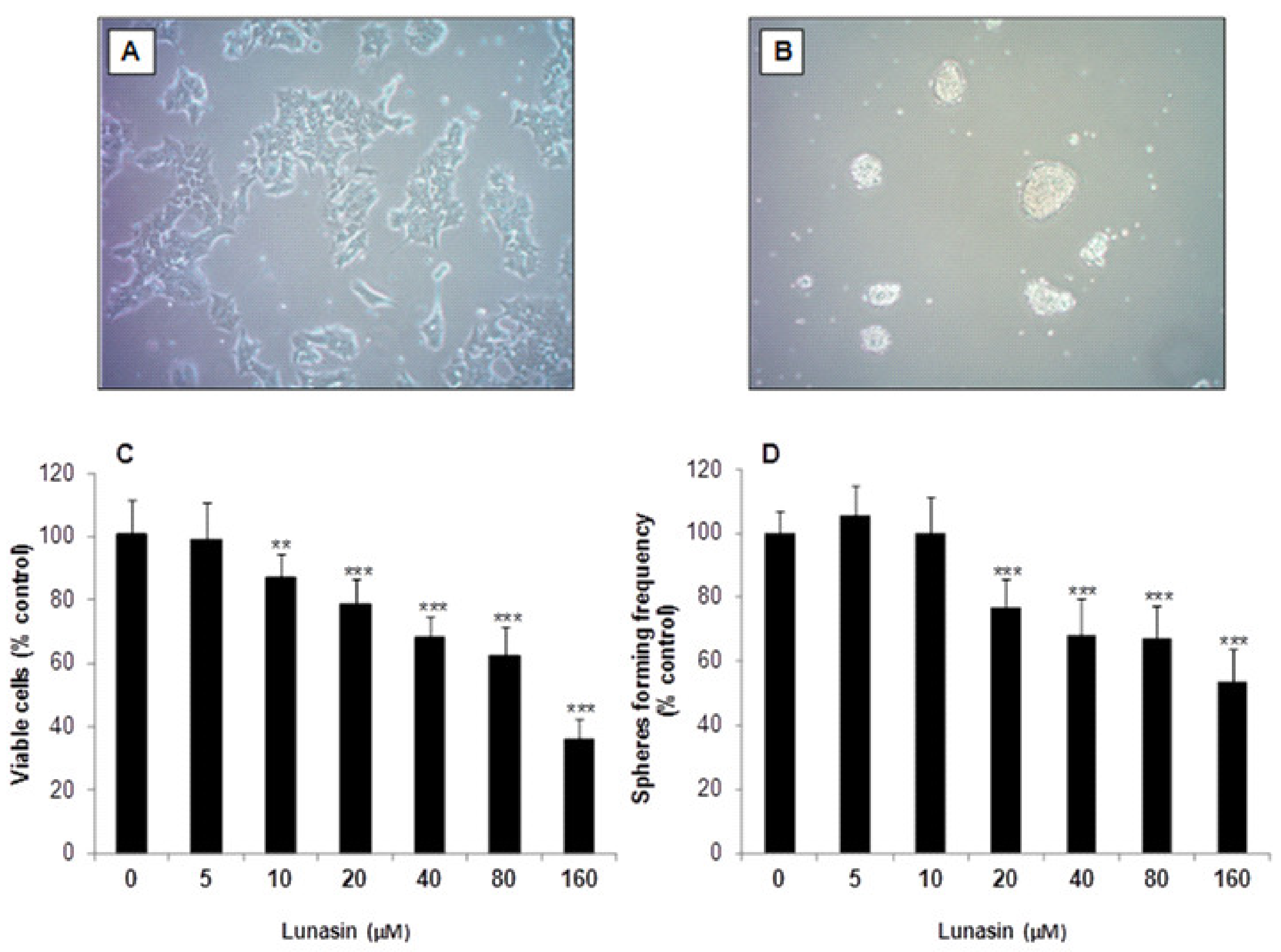
2.1. Inhibitory Effect of Lunasin on Cell Viability and Tumorsphere Formation
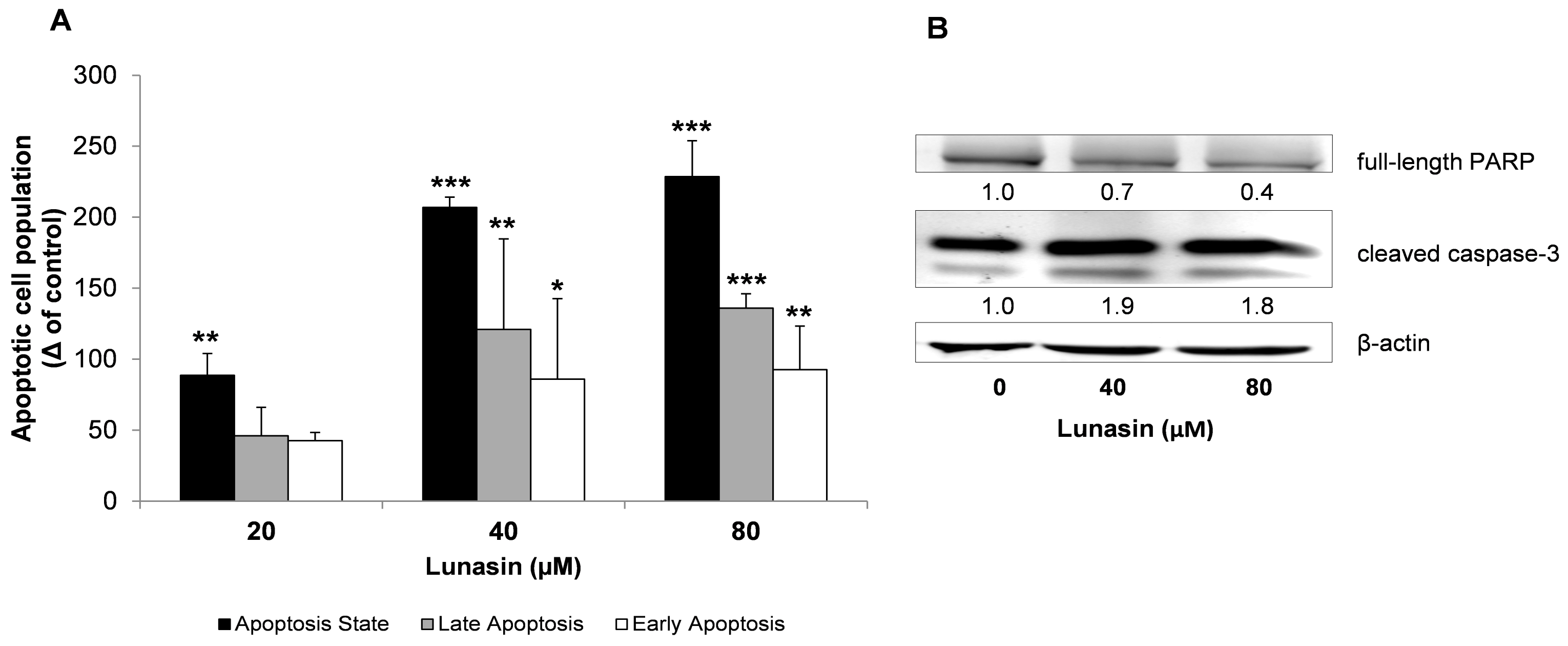
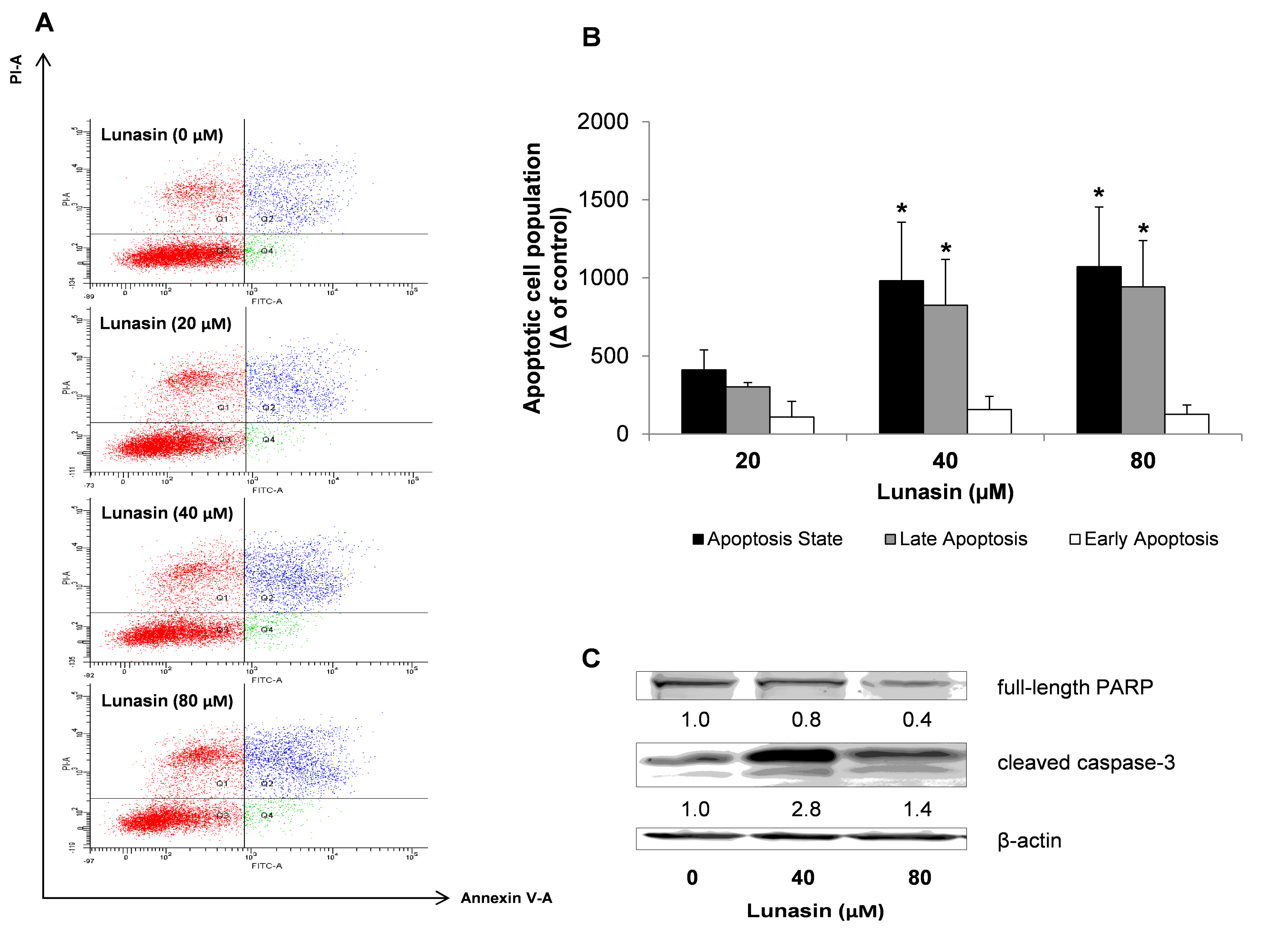
2.2. Apoptosis Analysis of Lunasin-Treated CRC Cells
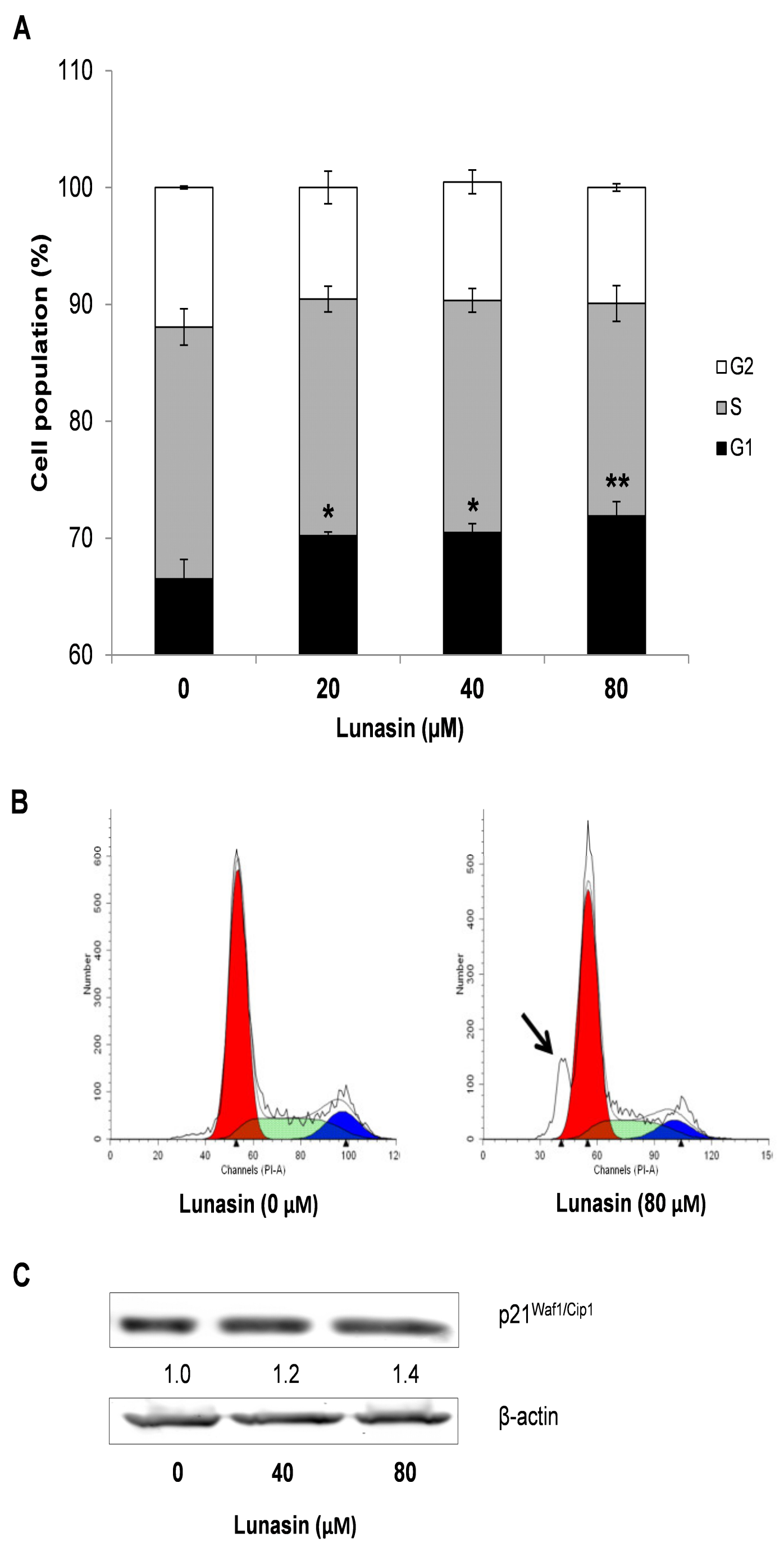
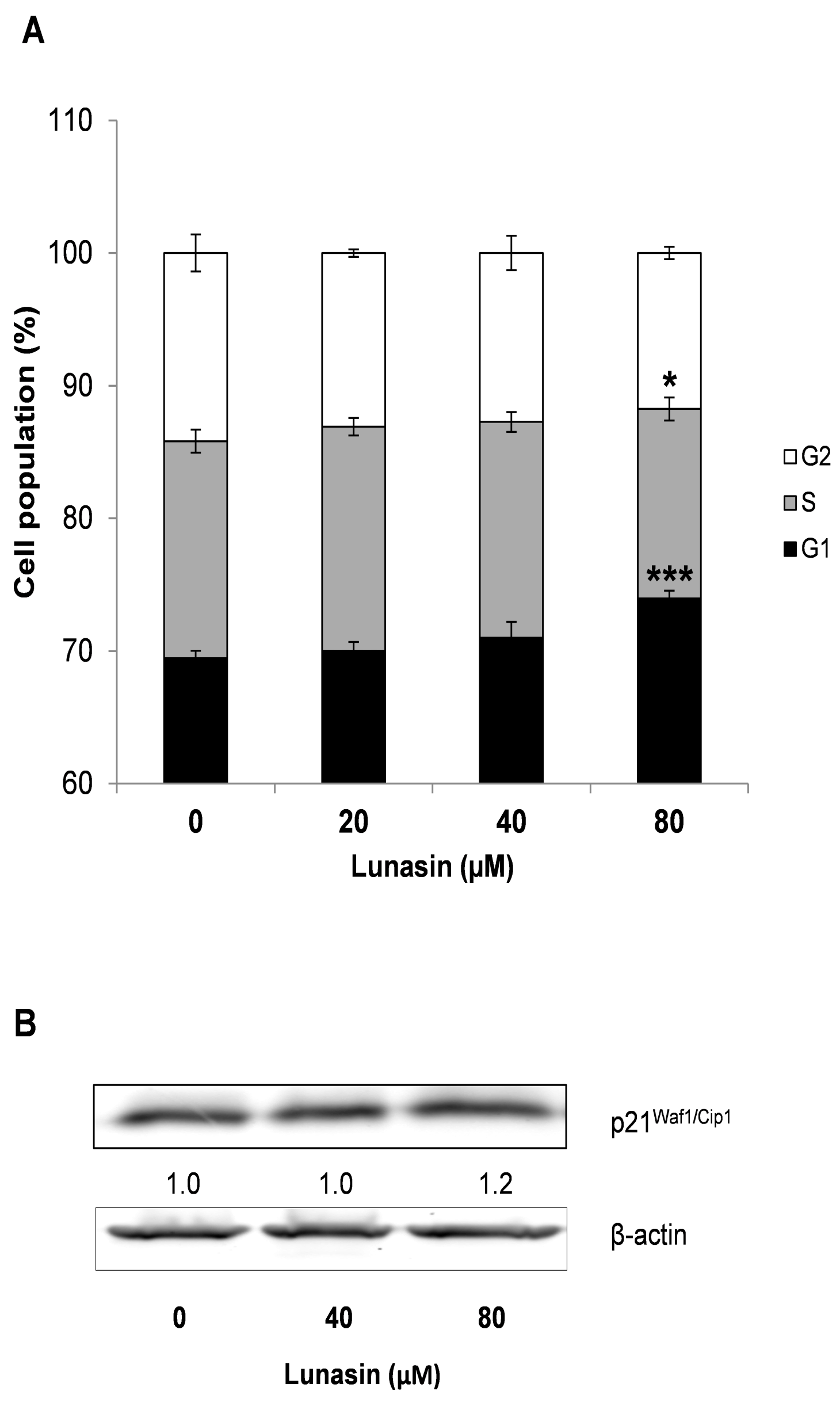
2.3. Effect of Lunasin on Cell Cycle Progression of CRC Cells
3. Materials and Methods
3.1. Materials
3.2. Cell Lines
3.3. Cell Proliferation Assay
3.4. Tumorsphere Formation Assay
3.5. Detection of Apoptosis
3.6. Cell Cycle Analyses
3.7. Immunoblotting
3.8. Statistical Analysis
4. Conclusions
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| ATCC | American Type Cell Collection |
| CDK | Cyclin-dependent kinase |
| CRC | Colorectal cancer |
| CSC | Cancer stem-like cells |
| DHA | Docosahexaenoic acid |
| EDTA | Ethylenediaminitetraacetic acid |
| FBS | Fetal bovine serum |
| MTT | 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyl tetrazolium bromide |
| PBS | Phosphate buffer saline |
| PI | Propidium iodide |
| SFM | Serum-free medium |
References
- Arnold, M.; Sierra, M.S.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global patterns and trends in colorectal cancer incidence and mortality. Gut 2017, 66, 683–691. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- O’Brien, C.A.; Pollett, A.; Gallinger, S.; Dick, J.E. A human colon cancer cell capable of initiating tumor growth in immunodeficient mice. Nature 2007, 445, 106–110. [Google Scholar] [CrossRef] [PubMed]
- Anderson, E.C.; Hessman, C.; Levin, T.G.; Monroe, M.M.; Wong, M.H. The role of colorectal cancer stem cells in metastatic disease and therapeutic response. Cancer 2011, 3, 319–339. [Google Scholar] [CrossRef] [PubMed]
- Munro, M.J.; Wickremesekera, S.K.; Peng, L.; Tan, S.T.; Itinteang, T. Cancer stem cells in colorectal cancer: A review. J. Clin. Pathol. 2018, 71, 110–116. [Google Scholar] [CrossRef]
- Reya, T.; Morrison, S.J.; Clarke, M.F.; Weissman, I.L. Stem cells, cancer, and cancer stem cells. Nature 2001, 414, 105–111. [Google Scholar] [CrossRef] [Green Version]
- Visvader, J.E.; Lindeman, G.J. Cancer stem cells in solid tumors: Accumulating evidence and unresolved questions. Nat. Rev. Cancer 2008, 8, 755–768. [Google Scholar] [CrossRef]
- Zhou, B.B.S.; Zhang, H.Y.; Damelin, M.; Geles, K.G.; Grindley, J.C.; Dirks, P.B. Tumor-initiating cells: Challenges and opportunities for anticancer drug discovery. Nat. Rev. Drug Discov. 2009, 8, 806–823. [Google Scholar] [CrossRef]
- Clevers, H. The cancer stem cell: Premises, promises and challenges. Nat. Med. 2011, 17, 313–319. [Google Scholar] [CrossRef]
- Spector, D.; Anthony, M.; Alexander, D.; Arab, L. Soy consumption and colorectal cancer. Nutr. Cancer 2003, 47, 1–12. [Google Scholar] [CrossRef]
- Yang, G.; Shu, X.O.; Li, H.L.; Chow, W.H.; Cai, H.; Zhang, X.L.; Gao, Y.T.; Zheng, W. Prospective cohort study of soy food intake and colorectal cancer risk in women. Am. J. Clin. Nutr. 2009, 89, 577–583. [Google Scholar] [CrossRef] [Green Version]
- Kim, Y.S.; Farrar, W.; Colburn, N.H.; Milner, J.A. Cancer stem cells: Potential target for bioactive food components. J. Nutr. Biochem. 2012, 23, 691–698. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsai, C.Y.; Chen, Y.H.; Chien, Y.W.; Huang, W.H.; Lin, S.H. Effect of soy saponin on the growth of human colon cancer cells. World J. Gastroenterol. 2010, 16, 3371–3376. [Google Scholar] [CrossRef] [PubMed]
- De Mejia, E.G.; Bradford, T.; Hasler, C. The anticarcinogenic potential of soybean lectin and lunasin. Nutr. Rev. 2003, 61, 239–246. [Google Scholar] [CrossRef] [PubMed]
- Clemente, A.; Moreno, F.J.; Marín-Manzano, M.C.; Jiménez, E.; Domoney, C. The cytotoxic effect of Bowman-Birk isoinhibitors, IBB1 and IBBD2, from soybean (Glycine max) on HT29 human colorectal cancer cells is related to their intrinsic ability to inhibit serine proteases. Mol. Nutr. Food Res. 2010, 54, 396–405. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fernández-Tomé, S.; Hernández-Ledesma, B. Current state of art after twenty years of the discovery of bioactive peptide lunasin. Food Res. Int. 2019, 116, 71–78. [Google Scholar] [CrossRef]
- Dia, V.P.; de Mejia, E.G. Lunasin induces apoptosis and modifies the expression of genes associated with extracellular matrix and cell adhesion in human metastatic colon cancer cells. Mol. Nutr. Food Res. 2011, 55, 623–634. [Google Scholar] [CrossRef]
- Dia, V.P.; de Mejia, E.G. Lunasin promotes apoptosis in human colon cancer cells by mitochondrial pathway activation and induction of nuclear clusterin expression. Cancer Lett. 2010, 295, 44–53. [Google Scholar] [CrossRef]
- Dia, V.P.; de Mejia, E.G. Lunasin potentiates the effect of oxaliplatin preventing outgrowth of colon cancer metastasis, binds to α5β1 integrin and suppresses FAK/ERK/NF-κB signaling. Cancer Lett. 2011, 313, 167–180. [Google Scholar] [CrossRef]
- García-Nebot, M.J.; Recio, I.; Hernández-Ledesma, B. Antioxidant activity and protective effects of peptide lunasin against oxidative stress in intestinal Caco-2 cells. Food Chem. Toxicol. 2014, 65, 155–161. [Google Scholar] [CrossRef] [Green Version]
- Kawasaki, B.T.; Hurt, E.M.; Mistree, T.; Farrar, W.L. Targeting cancer stem cells with phytochemicals. Mol. Interv. 2008, 8, 174–184. [Google Scholar] [CrossRef]
- Li, Y.; Wicha, M.S.; Schwartz, S.J.; Sun, D. Implications of cancer stem cell theory for cancer chemoprevention by natural dietary compounds. J. Nutr. Biochem. 2011, 22, 799–806. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, G.N.; Song, J.H.; Kim, E.S.; Choi, H.T.; Jang, H.D. Isoflavone content and apoptotic effect in HT-29 cancer cells of a soy germ extract. Food Chem. 2012, 130, 404–407. [Google Scholar] [CrossRef]
- Oh, J.; Hlatky, L.; Jeong, Y.S.; Kim, D. Therapeutic effectiveness of anticancer phytochemicals on cancer stem cells. Toxins 2016, 8, 199. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Mejia, E.G.; Dia, V.P. The role of nutraceutical proteins and peptides in apoptosis, angiogenesis, and metastasis of cancer cells. Cancer Metastasis Rev. 2010, 29, 511–528. [Google Scholar] [CrossRef] [PubMed]
- Hernández-Ledesma, B.; Hsieh, C.-C. Chemopreventive role of food-derived proteins and peptides: A review. Crit. Rev. Food Sci. 2017, 57, 2358–2376. [Google Scholar] [CrossRef] [PubMed]
- Hernández-Ledesma, B.; Hsieh, C.-C.; de Lumen, B.O. Relationship between lunasin’s sequence and its inhibitory activity of histones H3 and H4 acetylation. Mol. Nutr. Food Res. 2011, 55, 989–998. [Google Scholar] [CrossRef] [PubMed]
- Kanwar, S.S.; Yu, Y.J.; Nautiyal, J.; Patel, B.B.; Majumdar, A.P.N. The Wnt/beta-catenin pathway regulates growth and maintenance of colonospheres. Mol. Cancer 2010, 9, 212. [Google Scholar] [CrossRef] [Green Version]
- Qiu, P.; Guan, H.; Dong, P.; Guo, S.; Zheng, J.; Li, S.; Chen, Y.; Ho, C.-T.; Pan, M.-H.; McClements, D.J.; et al. The inhibitory effects of 5-hydroxy-3, 6, 7, 8, 3′, 4′-hexamethoxyflavone on human colon cancer cells. Mol. Nutr. Food Res. 2011, 55, 1523–1532. [Google Scholar] [CrossRef] [Green Version]
- Pabona, J.M.P.; Dave, B.; Su, Y.; Montales, M.T.E.; de Lumen, B.O.; de Mejia, E.G.; Rahal, O.M.; Simmen, R.C.M. The soybean peptide lunasin promotes apoptosis of mammary epithelial cells via induction of tumor suppressor PTEN: Similarities and distinct actions from soy isoflavone genistein. Genes Nutr. 2013, 8, 79–90. [Google Scholar] [CrossRef] [Green Version]
- Yu, Y.; Nangia-Makker, P.; Farhana, L.; Rajendra, G.S.; Levi, E.; Majumdar, A.P. miR-21 and miR-145 cooperation in regulation of colon cancer stem cells. Mol. Cancer 2015, 14, 98. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.; Wang, X.-Q.; Zhang, Q.; Zhu, J.-Y.; Li, Y.; Xie, C.-F.; Li, X.-T.; Wu, J.-S.; Geng, S.-S.; Zhong, C.-Y.; et al. (-)-epigallocatechin-3-gallate inhibits colorectal cancer stem cells by suppressing Wnt/β-catenin pathway. Nutrients 2017, 9, 572. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, T.I. Chemopreventive drugs: Mechanisms via inhibition of cancer stem cells in colorectal cancer. World J. Gastroenterol. 2014, 20, 3835–3846. [Google Scholar] [CrossRef]
- Yang, T.; Fang, S.; Zhang, H.X.; Xu, L.X.; Zhang, Z.Q.; Yuan, K.T.; Xue, C.L.; Yu, H.L.; Zhang, S.; Li, Y.F.; et al. N-3 PUFAs have antiproliferative and apoptotic effects on human colorectal cancer stem-like cells in vitro. J. Nutr. Biochem. 2013, 24, 744–753. [Google Scholar] [CrossRef] [PubMed]
- McConnell, E.J.; Devapatla, B.; Yaddanapudi, K.; Davis, K.R. The soybean-derived peptide lunasin inhibits non-small cell lung cancer cell proliferation by suppressing phosphorylation of the retinoblastoma protein. Oncotarget 2015, 6, 4649–4662. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shidal, C.; Al-Rayyan, N.; Yaddanapudi, K.; Davis, K.R. Lunasin is a novel therapeutic agent for targeting melanoma cancer stem cells. Oncotarget 2016, 7, 84128–84141. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shidal, C.; Inaba, J.-I.; Yaddanapudi, K.; Davis, K.R. The soy-derived peptide Lunasin inhibits potential of melanoma initiating cells. Oncotarget 2017, 8, 25525–25541. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hanahan, D.; Weinberg, R.A. Hallmarks of cancer: The next generation. Cell 2011, 144, 646–674. [Google Scholar] [CrossRef] [Green Version]
- Satoh, M.S.; Lindahl, T. Role of poly (ADP-ribose) formation in DNA repair. Nature 1992, 356, 356–358. [Google Scholar] [CrossRef]
- Oliver, F.J.; de la Rubia, G.; Rolli, V.; Ruiz-Ruiz, M.C.; de Murcia, G.; Menissier-de Murcia, J. Importance of poly(ADP-ribose) polymerase and its cleavage in apoptosis—Lesson from an uncleavable mutant. J. Biol. Chem. 1998, 273, 33533–33539. [Google Scholar] [CrossRef] [Green Version]
- Charoensinphon, N.; Qiu, P.; Dong, P.; Zheng, J.; Ngauv, P.; Cao, Y.; Li, S.; Ho, C.-T.; Xiao, H. 5-demethyltangeretin inhibits human non-small lung cancer cell growth by inducing G2/M cell cycle arrest and apoptosis. Mol. Nutr. Food Res. 2013, 57, 2103–2111. [Google Scholar] [CrossRef]
- De Mejia, E.G.; Wang, W.; Dia, V.P. Lunasin, with an arginine-glycine-aspartic acid motif, causes apoptosis to L1210 leukemia cells by activation of caspase-3. Mol. Nutr. Food Res. 2010, 54, 406–414. [Google Scholar] [CrossRef] [PubMed]
- Hsieh, C.-C.; Hernández-Ledesma, B.; de Lumen, B.O. Cell proliferation inhibitory and apoptosis-inducing properties of anacardic acid and lunasin in human breast cancer MDA-MB-231 cells. Food Chem. 2011, 125, 630–636. [Google Scholar] [CrossRef]
- Lin, L.; Liu, Y.; Li, H.; Li, P.K.; Fuchs, J.; Shibata, H.; Iwabuchi, Y.; Lin, J. Targeting colon cancer stem cells using a new curcumin analogue, GO-Y030. Br. J. Cancer 2011, 105, 212–220. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Massey, A.R.; Reddivari, L.; Vanamala, J. The dermal layer of sweet sorghum (Shorgum bicolor) stalk, a byproduct of biofuel production and source of unique 3-deoxyanthocyanidins, has more antiproliferative and proapoptotic activity than the pith in p53 variants of HCT116 and colon cancer stem cells. J. Agric. Food Chem. 2014, 62, 3150–3159. [Google Scholar] [PubMed]
- Ottinger, S.; Klöppel, A.; Rausch, V.; Liu, L.; Kallifatidis, G.; Gross, W.; Gebhard, M.M.; Brümmer, F.; Herr, I. Targeting of pancreatic and prostatic cancer stem cell characteristics by Crambe crambe marine sponge extract. Int. J. Cancer 2012, 130, 1671–1681. [Google Scholar] [CrossRef] [PubMed]
- Soner, B.C.; Aktug, H.; Acikgoz, E.; Duzagac, F.; Guven, U.; Ayla, S.; Cal, G.; Oktem, G. Induced growth inhibition, cell cycle arrest and apoptosis in CD133+/CD44+ prostate cancer stem cells by flavopiridol. Int. J. Mol. Med. 2014, 34, 1249–1256. [Google Scholar] [CrossRef] [Green Version]
- Nagata, S.; Nagase, H.; Kawane, K.; Mukae, N.; Fukuyama, H. Degradation of chromosomal DNA during apoptosis. Cell Death Differ. 2003, 10, 108–116. [Google Scholar] [CrossRef] [Green Version]
- Yang, W.; Meng, L.; Wang, H.; Chen, R.; Wang, R.; Ma, X.; Xu, G.; Zhou, J.; Wang, Y.; Lu, Y.; et al. Inhibition of proliferative and invasive capacities of breast cancer cells by arginine-glycine-aspartic acid peptide in vitro. Oncol. Rep. 2006, 15, 113–117. [Google Scholar] [CrossRef]
- Abukhdeir, A.M.; Park, B.H. p21 and p27: Roles in carcinogenesis and drug resistance. Expert Rev. Mol. Med. 2009, 10, e19. [Google Scholar] [CrossRef] [Green Version]
- Vuyyuri, S.B.; Shidal, C.; David, K.R. Development of the plant-derived peptide lunasin as an anticancer agent. Curr. Opin. Pharmacol. 2018, 41, 27–33. [Google Scholar] [CrossRef]
- Inaba, J.; McConnell, E.J.; Davis, K.R. Lunasin sensitivity in non-small lung cancer cells is linked to suppression of integrin signaling and changes in histone acetylation. Int. J. Mol. Sci. 2014, 15, 23705–23724. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, Q.; Pan, Y.; Cheng, Y.; Li, H.; Liu, D. Lunasin suppresses the migration and invasion of breast cancer cells by inhibiting matrix metalloproteinase-2/-9 via the FAK/Akt/ERK and NF-κB signaling pathways. Oncol. Rep. 2016, 36, 253–262. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dia, V.P.; de Mejia, E.G. Potential of lunasin orally-administered in comparison to intraperitoneal injection to inhibit colon cancer metastasis in vivo. J. Cancer Ther. 2013, 4, 34–43. [Google Scholar] [CrossRef] [Green Version]
- O’Brien, C.A.; Kreso, A.; Ryan, P.; Hermans, K.G.; Gibson, L.; Wang, Y.D.; Tsatsanis, A.; Gallinger, S.; Dick, J.E. ID1 and ID3 regulate the self-renewal capacity of human colon cancer-initiating cells through p21. Cancer Cell 2012, 21, 777–792. [Google Scholar] [CrossRef] [Green Version]
- Qiu, P.; Dong, P.; Guan, H.; Li, S.; Ho, C.T.; Pan, M.H.; McClements, D.J.; Xiao, H. Inhibitory effects of 5-hydroxy polymethoxyflavones on colon cancer cells. Mol. Nutr. Food Res. 2010, 54, S244–S252. [Google Scholar] [CrossRef]
- Xiao, H.; Yang, C.S.; Li, S.; Jin, H.; Ho, C.T.; Patel, T. Monodemethylated polymethoxyflavones from sweet orange (Citrus sinensis) peel inhibit growth of human lung cancer cells by apoptosis. Mol. Nutr. Food Res. 2009, 53, 398–406. [Google Scholar] [CrossRef]





© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fernández-Tomé, S.; Xu, F.; Han, Y.; Hernández-Ledesma, B.; Xiao, H. Inhibitory Effects of Peptide Lunasin in Colorectal Cancer HCT-116 Cells and Their Tumorsphere-Derived Subpopulation. Int. J. Mol. Sci. 2020, 21, 537. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21020537
Fernández-Tomé S, Xu F, Han Y, Hernández-Ledesma B, Xiao H. Inhibitory Effects of Peptide Lunasin in Colorectal Cancer HCT-116 Cells and Their Tumorsphere-Derived Subpopulation. International Journal of Molecular Sciences. 2020; 21(2):537. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21020537
Chicago/Turabian StyleFernández-Tomé, Samuel, Fei Xu, Yanhui Han, Blanca Hernández-Ledesma, and Hang Xiao. 2020. "Inhibitory Effects of Peptide Lunasin in Colorectal Cancer HCT-116 Cells and Their Tumorsphere-Derived Subpopulation" International Journal of Molecular Sciences 21, no. 2: 537. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21020537