iPSC-Derived MSCs Versus Originating Jaw Periosteal Cells: Comparison of Resulting Phenotype and Stem Cell Potential

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Tri-Lineage Differentiation of iMSCs and JPCs
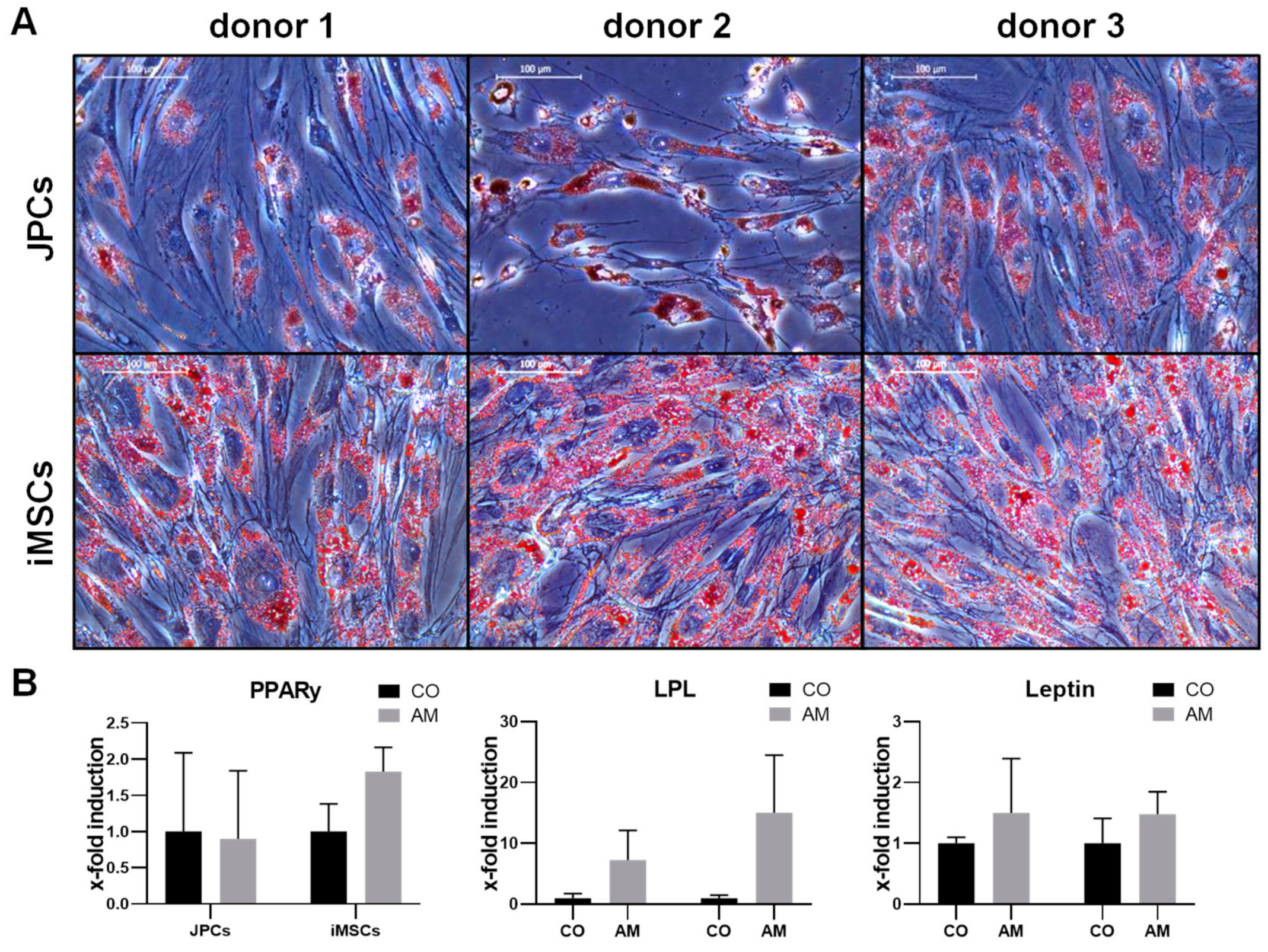
2.1.1. Adipogenic Differentiation
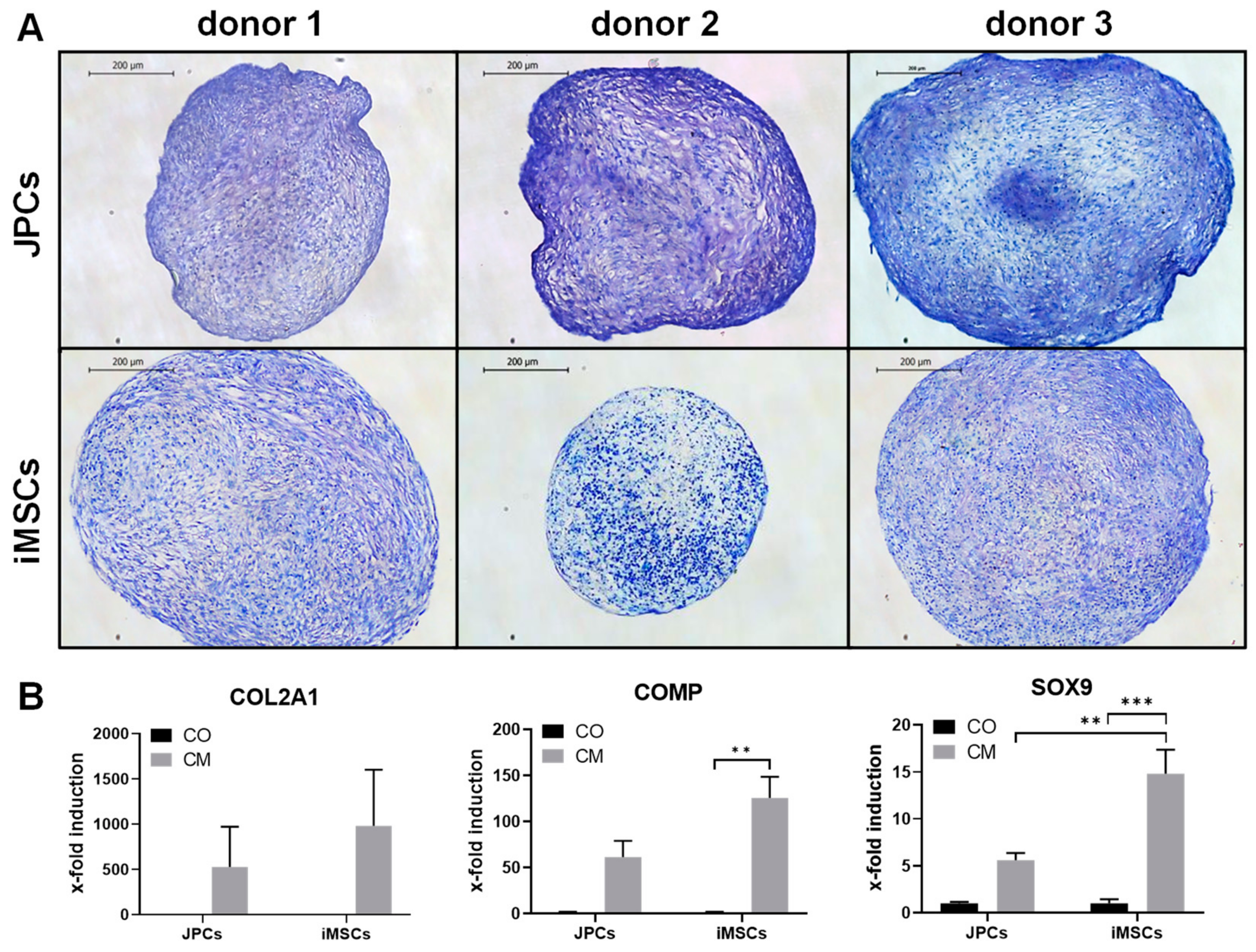
2.1.2. Chondrogenic Differentiation
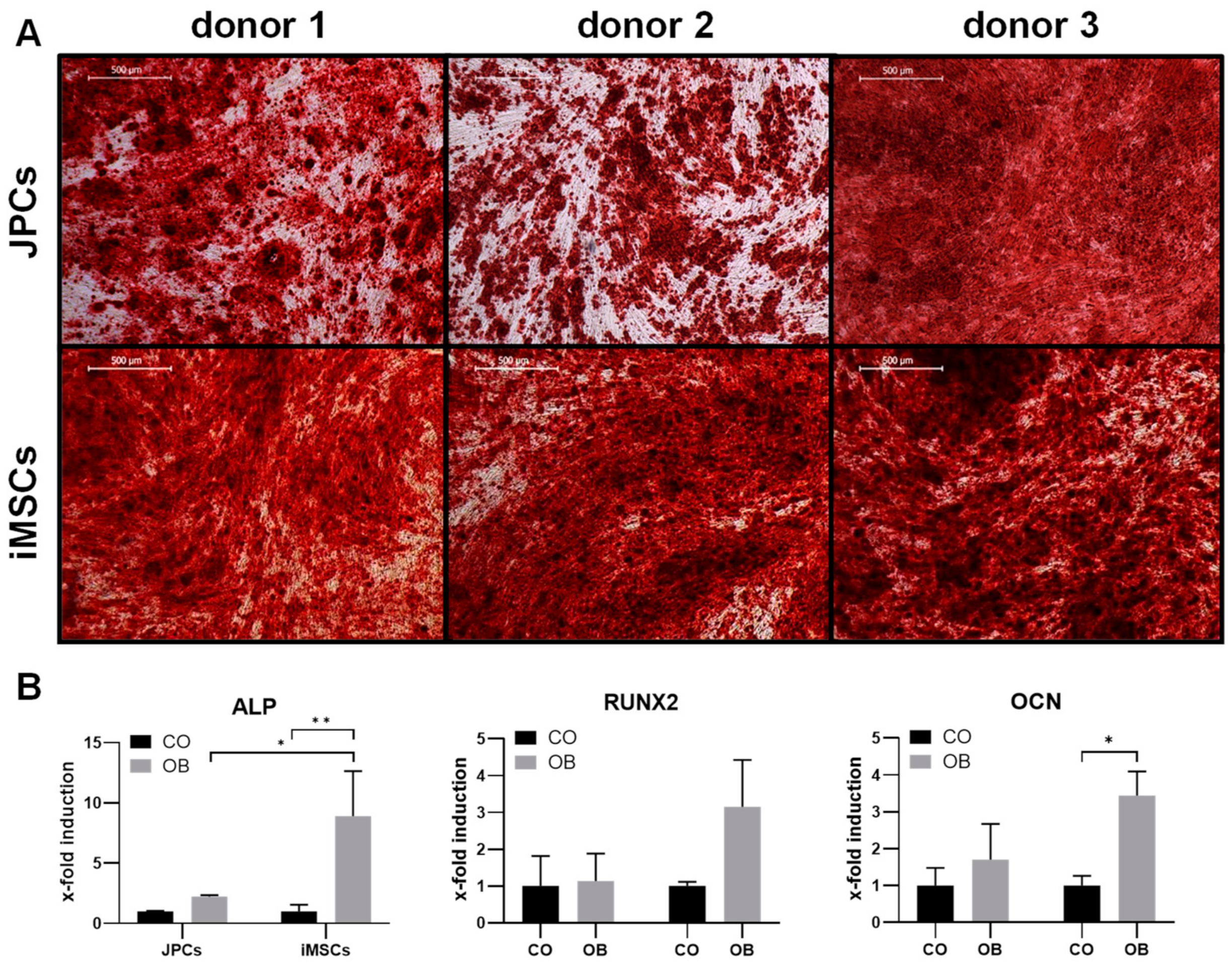
2.1.3. Osteogenic Differentiation
2.2. Rejuvenation and Senescence
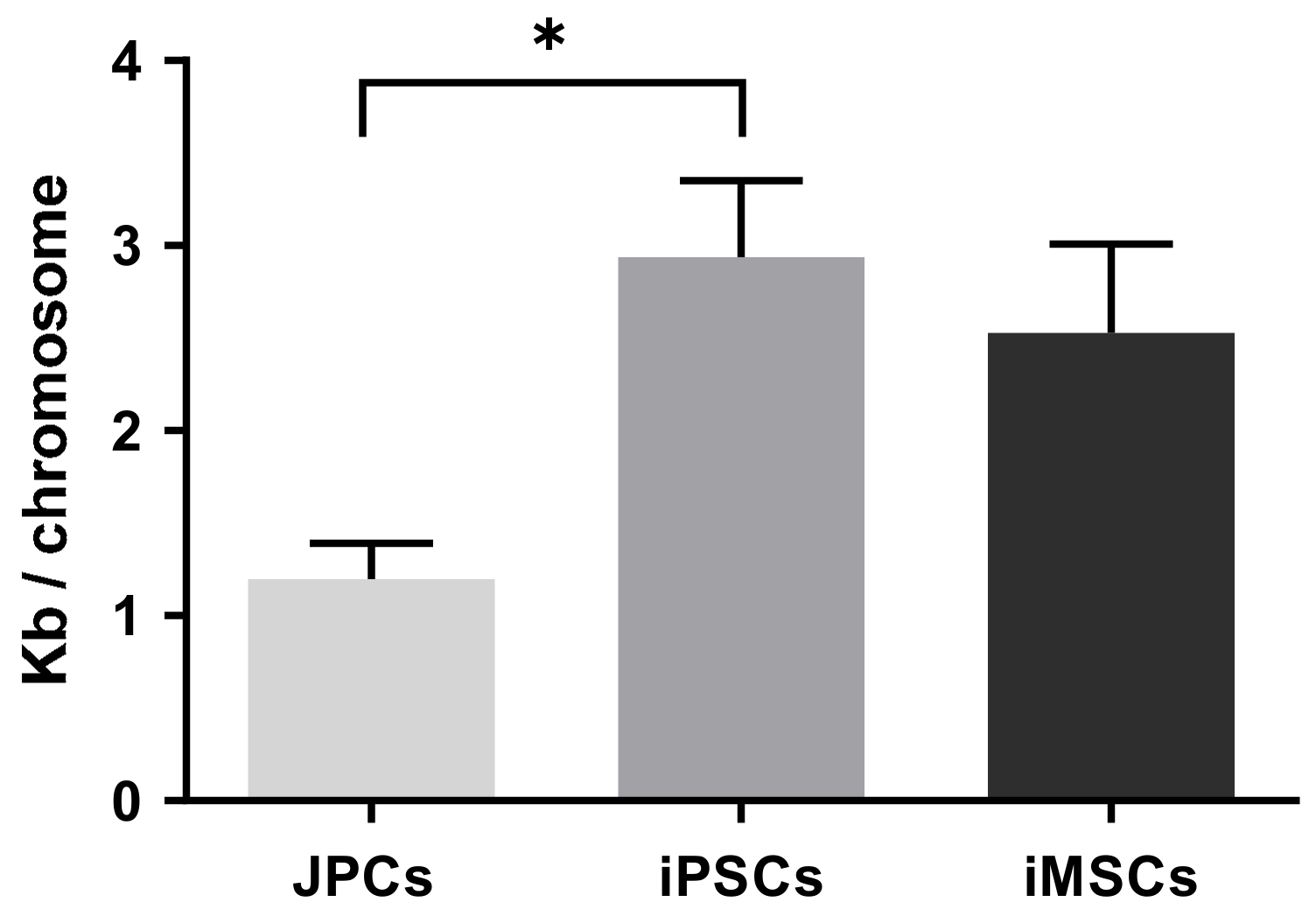
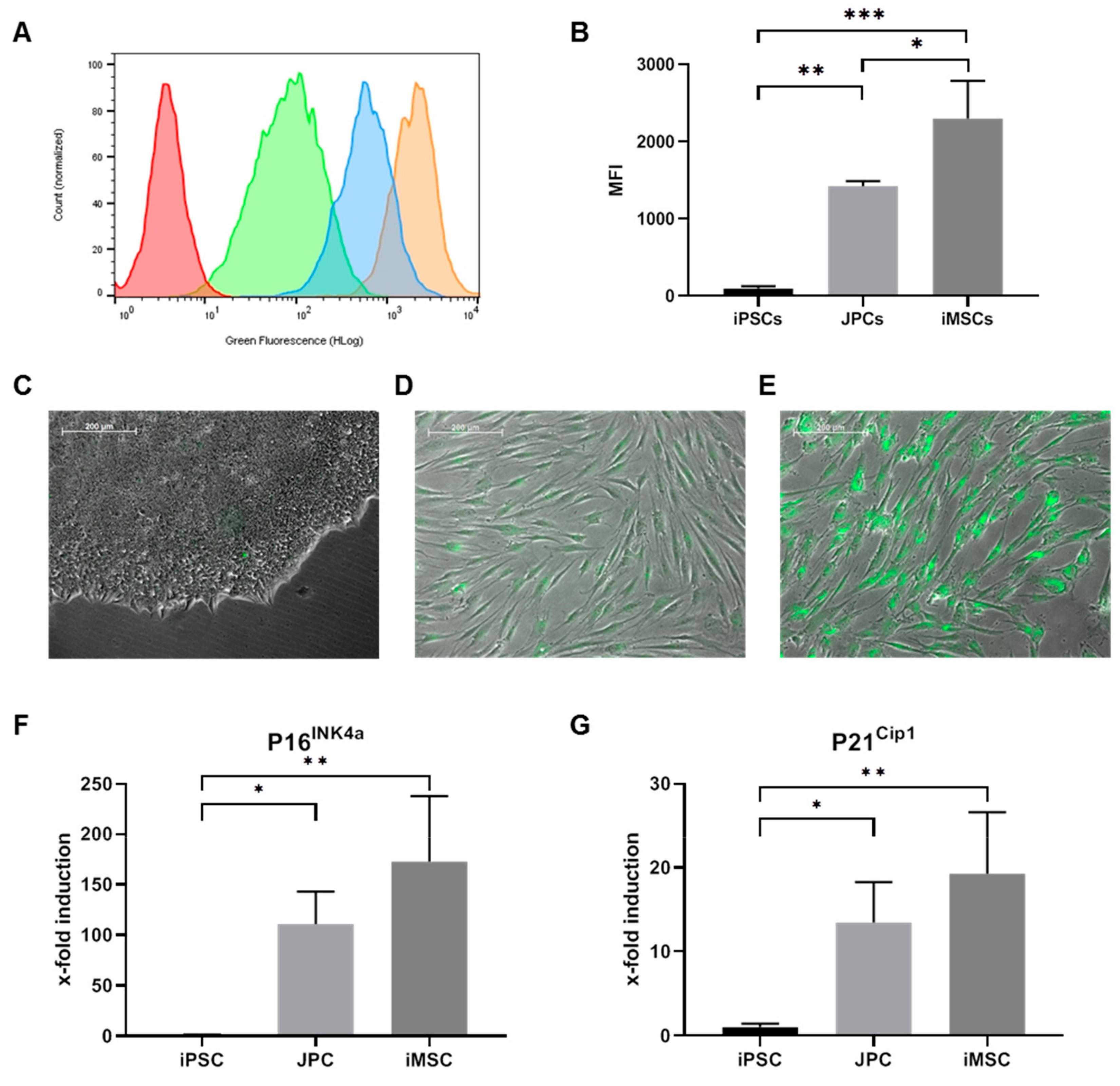
2.2.1. Telomere Length Assay
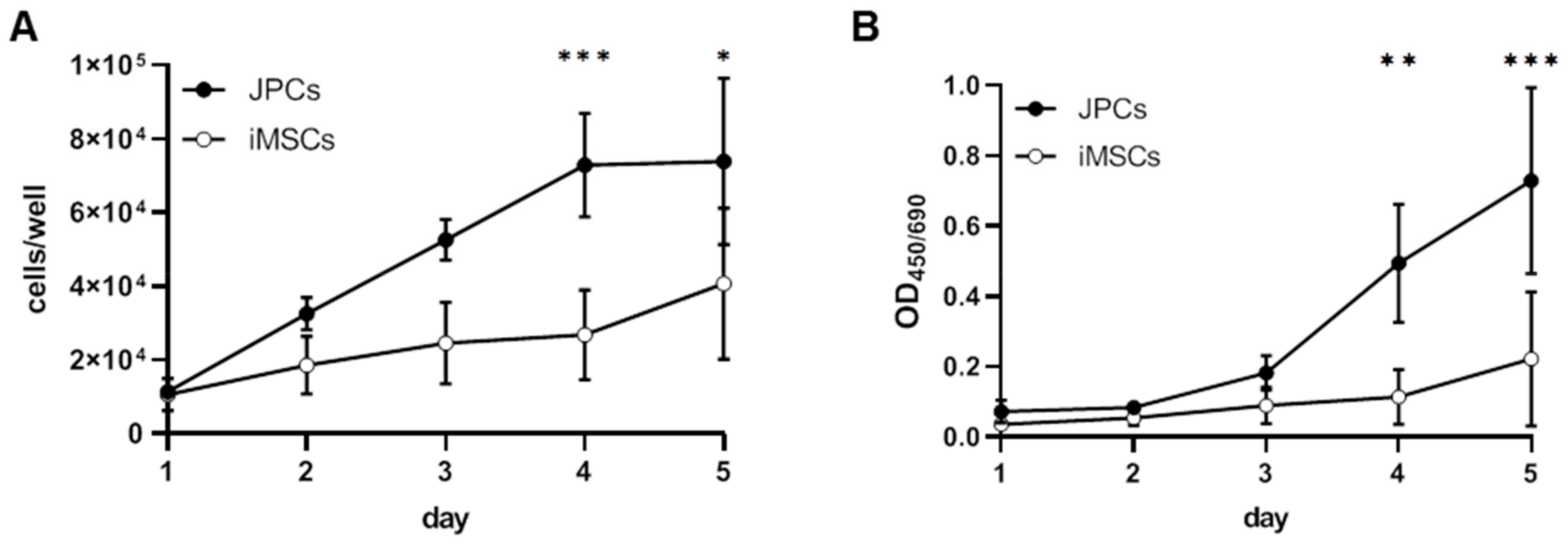
2.2.2. Proliferation, Mitochondrial Activity, and SA-β-Galactosidase Activity
3. Discussion
4. Materials and Methods
4.1. Cell Culture
4.2. Generation of Integration-Free iPSCs from JPCs Using srRNA
4.3. Differentiation of iPSCs to iMSCs
4.4. Tri-Lineage Differentiation of iMSCs
4.4.1. Osteogenic Differentiation
4.4.2. Adipogenic Differentiation
4.4.3. Chondrogenic Differentiation
4.5. Mitochondrial Activity Assay
4.6. Growth Kinetics
4.7. Senescence Assay
4.8. Telomere Length Measurement
4.9. Gene Expression Analysis of JPCs and iMSCs
4.10. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| AM | adipogenic medium |
| ANOVA | analysis of variance |
| BcM | B18R-conditioned medium |
| CM | chondrogenic medium |
| CO | control |
| FBS | fetal bovine serum |
| GAGs | glycosaminoglycans |
| GFP | green fluorescent protein |
| hPL | human platelet lysate |
| iMSC | iPSC-derived mesenchymal stem/stromal like cell |
| iPSC | induced pluripotent stem cell |
| JPC | jaw periosteal cell |
| kb | kilo basepairs |
| MFI | median fluorescence index |
| mRNA | messenger RNA |
| MSC | mesenchymal stem/stromal cell |
| NaB | sodium butyrate |
| OB | osteoblast medium |
| OSKM | OKT4, SOX2, KLF4, cMYC |
| PBS | phosphate buffered saline |
| PCR | polymerase chain reaction |
| Puro | puromycin |
| qRT-PCR | quantitative real-time polymerase chain reaction |
| ROCK | rho-associated protein kinase |
| SA-β-gal | senescence-associated beta-galactosidase |
| SD | standard deviation |
| SEM | standard error of mean |
| srRNA | self-replicating ribonucleic acid |
References
- Čamernik, K.; Barlič, A.; Drobnic, M.; Marc, J.; Jeras, M.; Zupan, J. Mesenchymal Stem Cells in the Musculoskeletal System: From Animal Models to Human Tissue Regeneration? Stem Cell Rev. Rep. 2018, 14, 346–369. [Google Scholar] [CrossRef] [PubMed]
- Fitzsimmons, R.E.B.; Mazurek, M.S.; Soos, A.; Simmons, C.A. Mesenchymal Stromal/Stem Cells in Regenerative Medicine and Tissue Engineering. Stem Cells Int. 2018, 2018, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Gebler, A.; Zabel, O.; Seliger, B. The immunomodulatory capacity of mesenchymal stem cells. Trends Mol. Med. 2012, 18, 128–134. [Google Scholar] [CrossRef]
- Zachar, L.; Bačenková, D.; Rosocha, J. Activation, homing, and role of the mesenchymal stem cells in the inflammatory environment. J. Inflamm. Res. 2016, 9, 231–240. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wagner, W.; Ho, A.D. Mesenchymal Stem Cell Preparations—Comparing Apples and Oranges. Stem Cell Rev. Rep. 2007, 3, 239–248. [Google Scholar] [CrossRef]
- Baker, N.; Boyette, L.B.; Tuan, R.S. Characterization of bone marrow-derived mesenchymal stem cells in aging. Bone 2015, 70, 37–47. [Google Scholar] [CrossRef]
- Yu, J.; Thomson, J.A. Induced Pluripotent Stem Cells. In Principles of Tissue Engineering; Lanza, R., Langer, R., Vacanti, J., Eds.; Academic Press: Boston, MA, USA, 2014; pp. 581–594. [Google Scholar]
- Jung, Y.; Bauer, G.; Nolta, J.A. Concise review: Induced pluripotent stem cell-derived mesenchymal stem cells: Progress toward safe clinical products. Stem Cells 2012, 30, 42–47. [Google Scholar] [CrossRef] [Green Version]
- Ng, J.; Hynes, K.; White, G.; Sivanathan, K.N.; VanDyke, K.; Bartold, P.M.; Gronthos, S. Immunomodulatory Properties of Induced Pluripotent Stem Cell-Derived Mesenchymal Cells. J. Cell. Biochem. 2016, 117, 2844–2853. [Google Scholar] [CrossRef] [Green Version]
- Lapasset, L.; Milhavet, O.; Prieur, A.; Besnard, E.; Babled, A.; Aït-Hamou, N.; Leschik, J.; Pellestor, F.; Ramirez, J.-M.; De Vos, J.; et al. Rejuvenating senescent and centenarian human cells by reprogramming through the pluripotent state. Genes Dev. 2011, 25, 2248–2253. [Google Scholar] [CrossRef] [Green Version]
- Spitzhorn, L.-S.; Megges, M.; Wruck, W.; Rahman, S.; Otte, J.; Degistirici, Ö.; Meisel, R.; Sorg, R.V.; Oreffo, R.O.C.; Adjaye, J. Human iPSC-derived MSCs (iMSCs) from aged individuals acquire a rejuvenation signature. Stem Cell Res. Ther. 2019, 10, 100. [Google Scholar] [CrossRef] [Green Version]
- Turinetto, V.; Vitale, E.; Giachino, C. Senescence in Human Mesenchymal Stem Cells: Functional Changes and Implications in Stem Cell-Based Therapy. Int. J. Mol. Sci. 2016, 17, 1164. [Google Scholar] [CrossRef] [PubMed]
- Umrath, F.; Steinle, H.; Weber, M.; Wendel, H.-P.; Reinert, S.; Alexander, D.; Avci-Adali, M. Generation of iPSCs from Jaw Periosteal Cells Using Self-Replicating RNA. Int. J. Mol. Sci. 2019, 20, 1648. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alexander, D.; Hoffmann, J.; Munz, A.; Friedrich, B.; Geis-Gerstorfer, J.; Reinert, S. Analysis of OPLA scaffolds for bone engineering constructs using human jaw periosteal cells. J. Mater. Sci. Mater. Med. 2008, 19, 965–974. [Google Scholar] [CrossRef] [PubMed]
- Ferretti, C.; Mattioli-Belmonte, M. Periosteum derived stem cells for regenerative medicine proposals: Boosting current knowledge. World J. Stem Cells 2014, 6, 266–277. [Google Scholar] [CrossRef]
- Roberts, S.J.; Van Gastel, N.; Carmeliet, G.; Luyten, F.P. Uncovering the periosteum for skeletal regeneration: The stem cell that lies beneath. Bone 2015, 70, 10–18. [Google Scholar] [CrossRef] [PubMed]
- Frobel, J.; Hemeda, H.; Lenz, M.; Abagnale, G.; Joussen, S.; Denecke, B.; Saric, T.; Zenke, M.; Wagner, W. Epigenetic rejuvenation of mesenchymal stromal cells derived from induced pluripotent stem cells. Stem Cell Rep. 2014, 3, 414–422. [Google Scholar] [CrossRef] [Green Version]
- Marión, R.M.; Strati, K.; Li, H.; Tejera, A.; Schoeftner, S.; Ortega, S.; Serrano, M.; Blasco, M.A. Telomeres Acquire Embryonic Stem Cell Characteristics in Induced Pluripotent Stem Cells. Cell Stem Cell 2009, 4, 141–154. [Google Scholar] [CrossRef] [Green Version]
- Bodnar, A.G.; Ouellette, M.; Frolkis, M.; Holt, S.E.; Chiu, C.P.; Morin, G.B.; Harley, C.B.; Shay, J.W.; Lichtsteiner, S.; Wright, W.E. Extension of Life-Span by Introduction of Telomerase into Normal Human Cells. Science 1998, 279, 349–352. [Google Scholar] [CrossRef] [Green Version]
- Feng, Q.; Klimanskaya, I.; Gomes, I.; Kim, H.; Chung, Y.; Honig, G.R.; Kim, K.-S.; Lanza, R.; Lu, S.-J. Hemangioblastic Derivatives from Human Induced Pluripotent Stem Cells Exhibit Limited Expansion and Early Senescence. Stem Cells 2010, 28, 704–712. [Google Scholar] [CrossRef]
- Gokoh, M.; Nishio, M.; Nakamura, N.; Matsuyama, S.; Nakahara, M.; Suzuki, S.; Mitsumoto, M.; Akutsu, H.; Umezawa, A.; Yasuda, K.; et al. Early Senescence Is Not an Inevitable Fate of Human-Induced Pluripotent Stem-Derived Cells. Cell. Reprogramming 2011, 13, 361–370. [Google Scholar] [CrossRef]
- McHugh, D.; Gil, J. Senescence and aging: Causes, consequences, and therapeutic avenues. J. Cell Boil. 2017, 217, 65–77. [Google Scholar] [CrossRef] [PubMed]
- Jun, J.-I.; Lau, L.F. The matricellular protein CCN1 induces fibroblast senescence and restricts fibrosis in cutaneous wound healing. Nat. Cell Biol. 2010, 12, 676–685. [Google Scholar] [CrossRef] [PubMed]
- DeVito, L.; Klontzas, M.E.; Cvoro, A.; Galleu, A.; Simon, M.; Hobbs, C.; Dazzi, F.; Mantalaris, A.; Khalaf, Y.; Ilic, D. Comparison of human isogeneic Wharton’s jelly MSCs and iPSC-derived MSCs reveals differentiation-dependent metabolic responses to IFNG stimulation. Cell Death Dis. 2019, 10, 277. [Google Scholar] [CrossRef] [PubMed]
- Luzzani, C.D.; Miriuka, S.G. Pluripotent Stem Cells as a Robust Source of Mesenchymal Stem Cells. Stem Cell Rev. Rep. 2017, 13, 68–78. [Google Scholar]
- Nejadnik, H.; Diecke, S.; Lenkov, O.D.; Chapelin, F.; Donig, J.; Tong, X.; Derugin, N.; Chan, R.C.F.; Gaur, A.; Yang, F.; et al. Improved approach for chondrogenic differentiation of human induced pluripotent stem cells. Stem Cell Rev. Rep. 2015, 11, 242–253. [Google Scholar] [CrossRef] [Green Version]
- Sun, Y.-Q.; Zhang, Y.; Li, X.; Deng, M.-X.; Gao, W.-X.; Yao, Y.; Chiu, S.-M.; Liang, X.; Gao, F.; Chan, C.W.; et al. Insensitivity of Human iPS Cells-Derived Mesenchymal Stem Cells to Interferon-γ-induced HLA Expression Potentiates Repair Efficiency of Hind Limb Ischemia in Immune Humanized NOD Scid Gamma Mice. Stem Cells 2015, 33, 3452–3467. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Chiu, S.; Liang, X.; Gao, F.; Zhang, Z.; Liao, S.; Liang, Y.; Chai, Y.-H.; Low, D.J.H.; Tse, H.-F.; et al. Rap1-mediated nuclear factor-kappaB (NF-κB) activity regulates the paracrine capacity of mesenchymal stem cells in heart repair following infarction. Cell Death Discov. 2015, 1, 15007. [Google Scholar] [CrossRef] [Green Version]






© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Umrath, F.; Weber, M.; Reinert, S.; Wendel, H.-P.; Avci-Adali, M.; Alexander, D. iPSC-Derived MSCs Versus Originating Jaw Periosteal Cells: Comparison of Resulting Phenotype and Stem Cell Potential. Int. J. Mol. Sci. 2020, 21, 587. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21020587
Umrath F, Weber M, Reinert S, Wendel H-P, Avci-Adali M, Alexander D. iPSC-Derived MSCs Versus Originating Jaw Periosteal Cells: Comparison of Resulting Phenotype and Stem Cell Potential. International Journal of Molecular Sciences. 2020; 21(2):587. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21020587
Chicago/Turabian StyleUmrath, Felix, Marbod Weber, Siegmar Reinert, Hans-Peter Wendel, Meltem Avci-Adali, and Dorothea Alexander. 2020. "iPSC-Derived MSCs Versus Originating Jaw Periosteal Cells: Comparison of Resulting Phenotype and Stem Cell Potential" International Journal of Molecular Sciences 21, no. 2: 587. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21020587