Scenarios of Genes-to-Terpenoids Network Led to the Identification of a Novel α/β-Farnesene/β-Ocimene Synthase in Camellia sinensis


Abstract
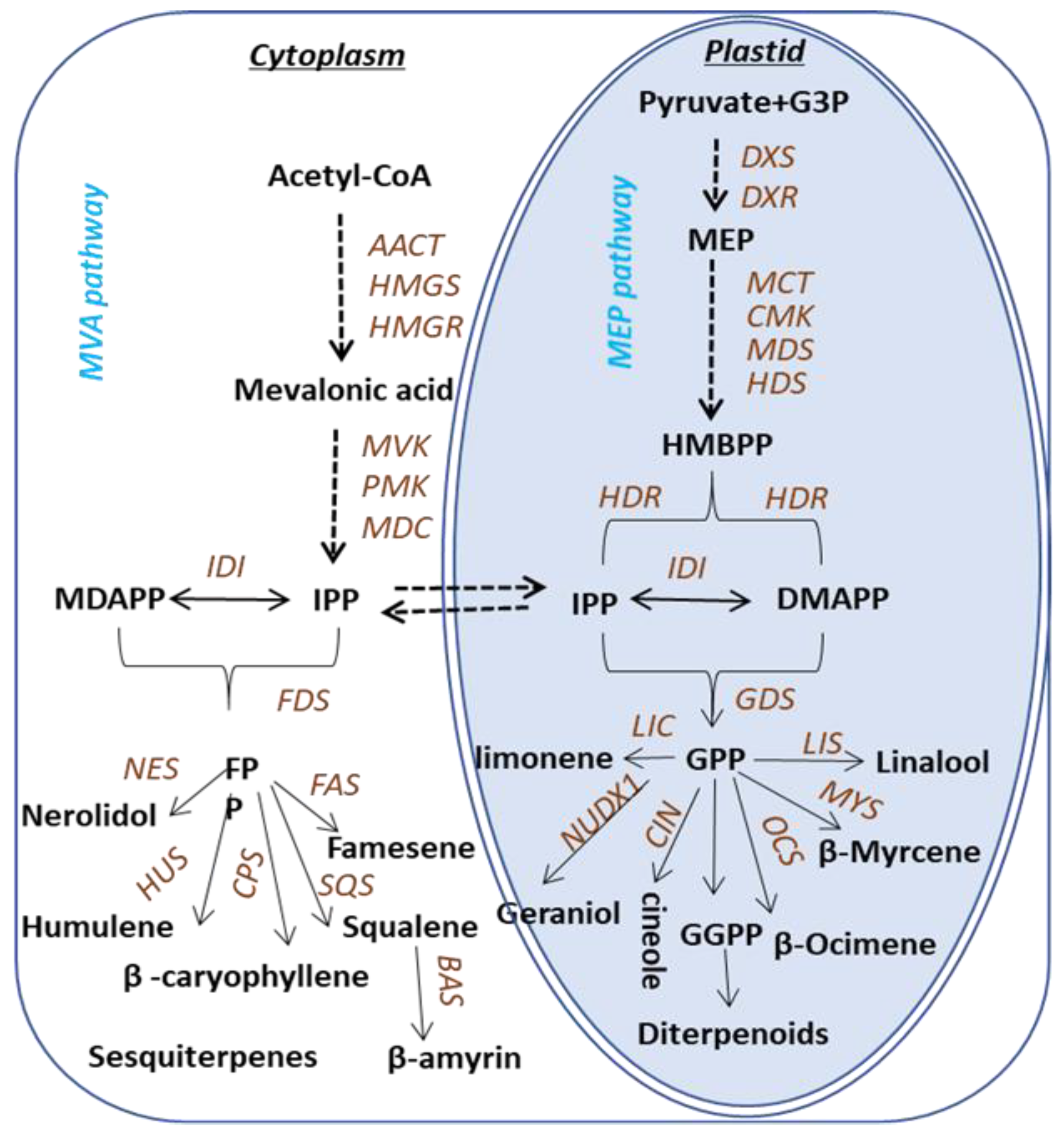
:1. Introduction
2. Results and Discussion
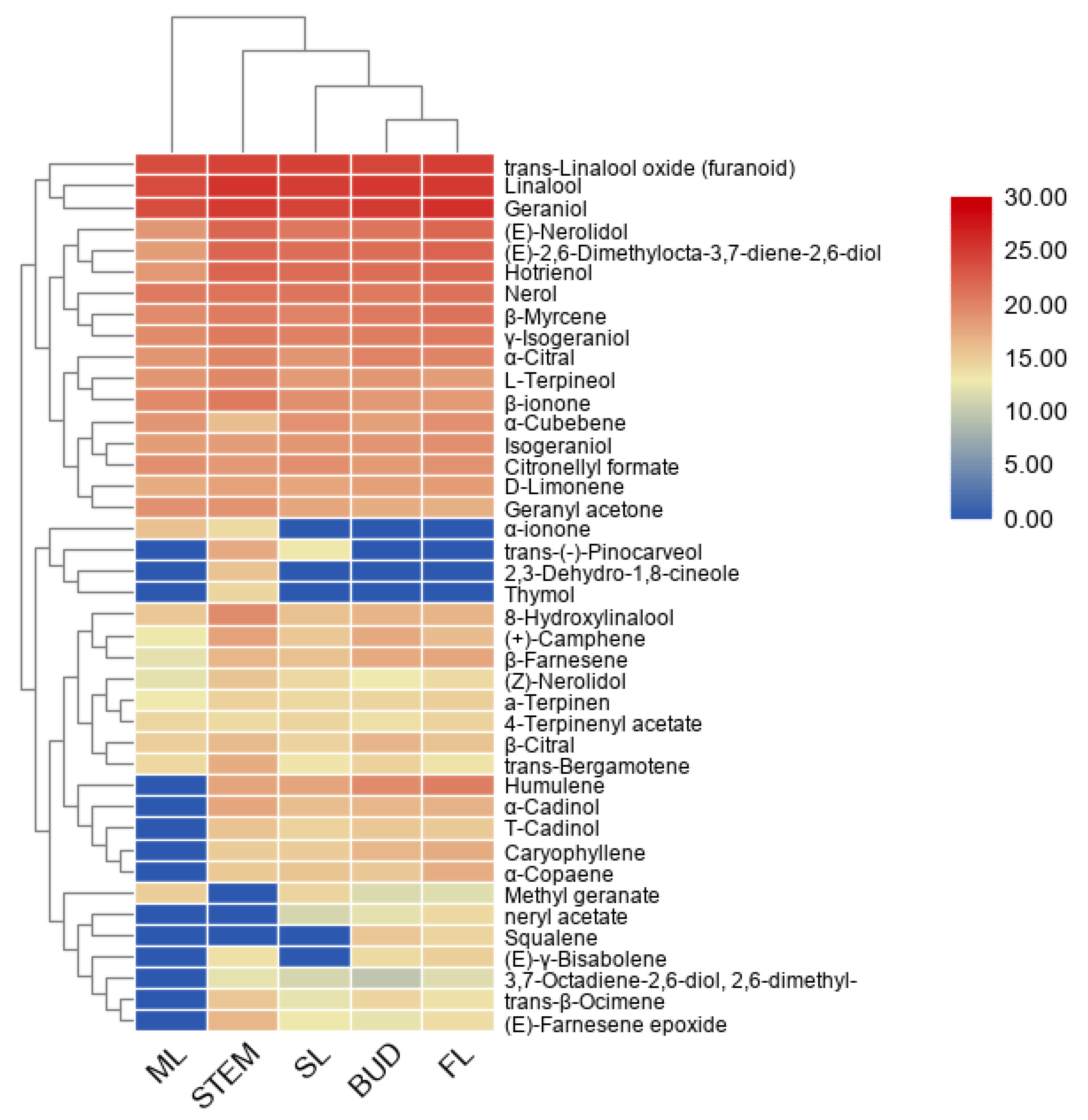
2.1. GC–MS-Based Metabolite Profiles
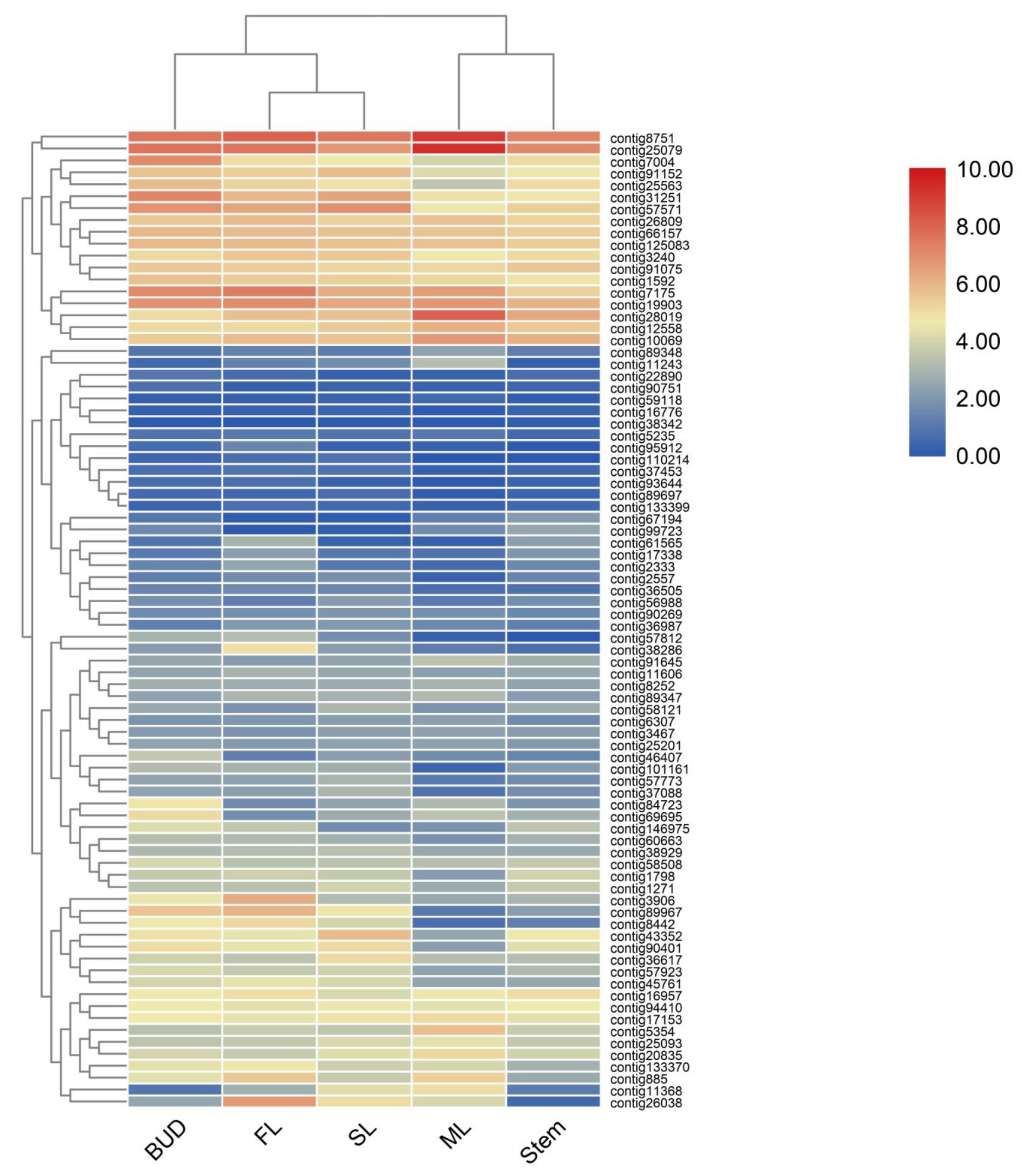
2.2. Transcriptome Sequencing
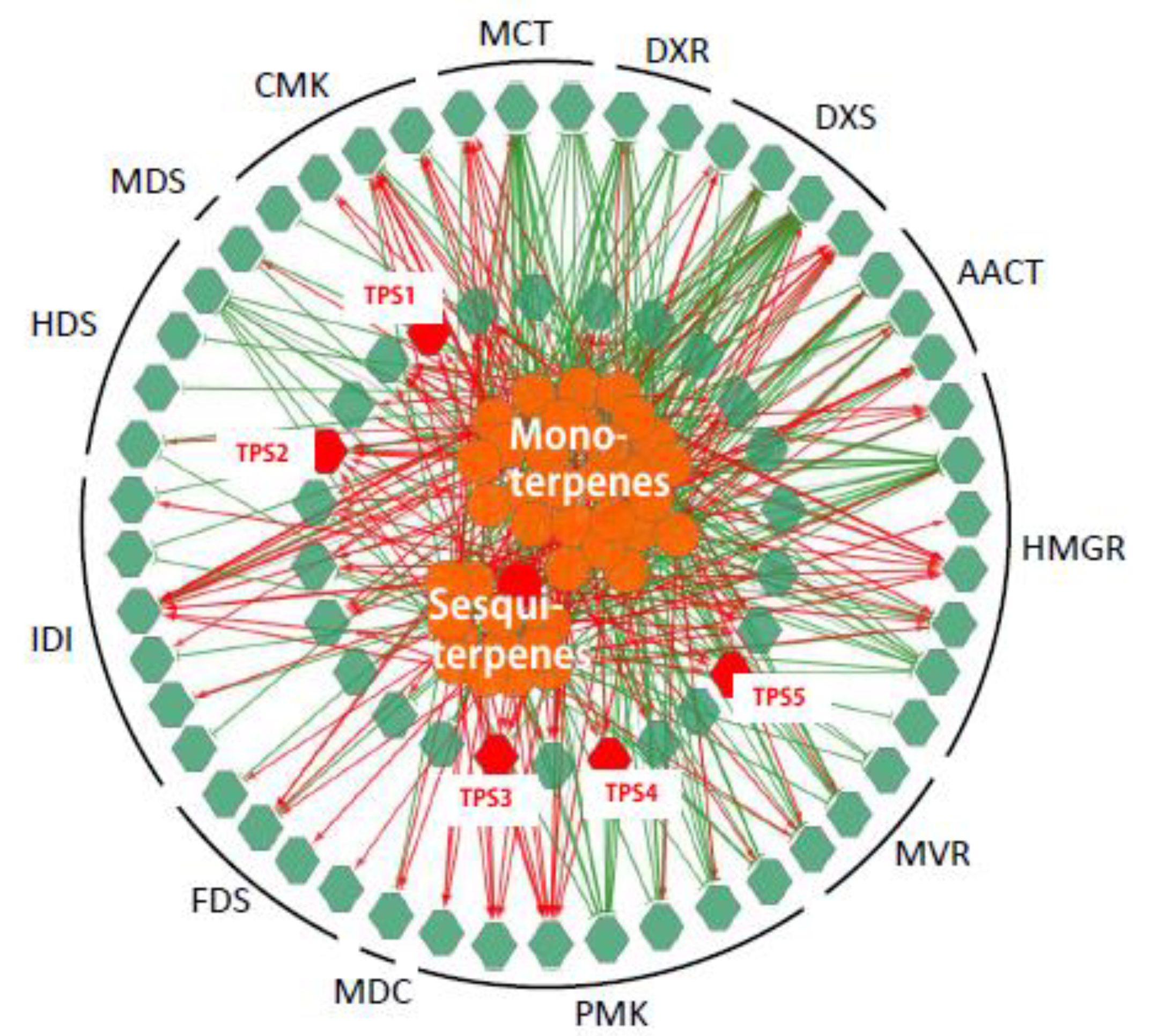
2.3. Integration of Gene Expression Levels and Terpenoid Profiles
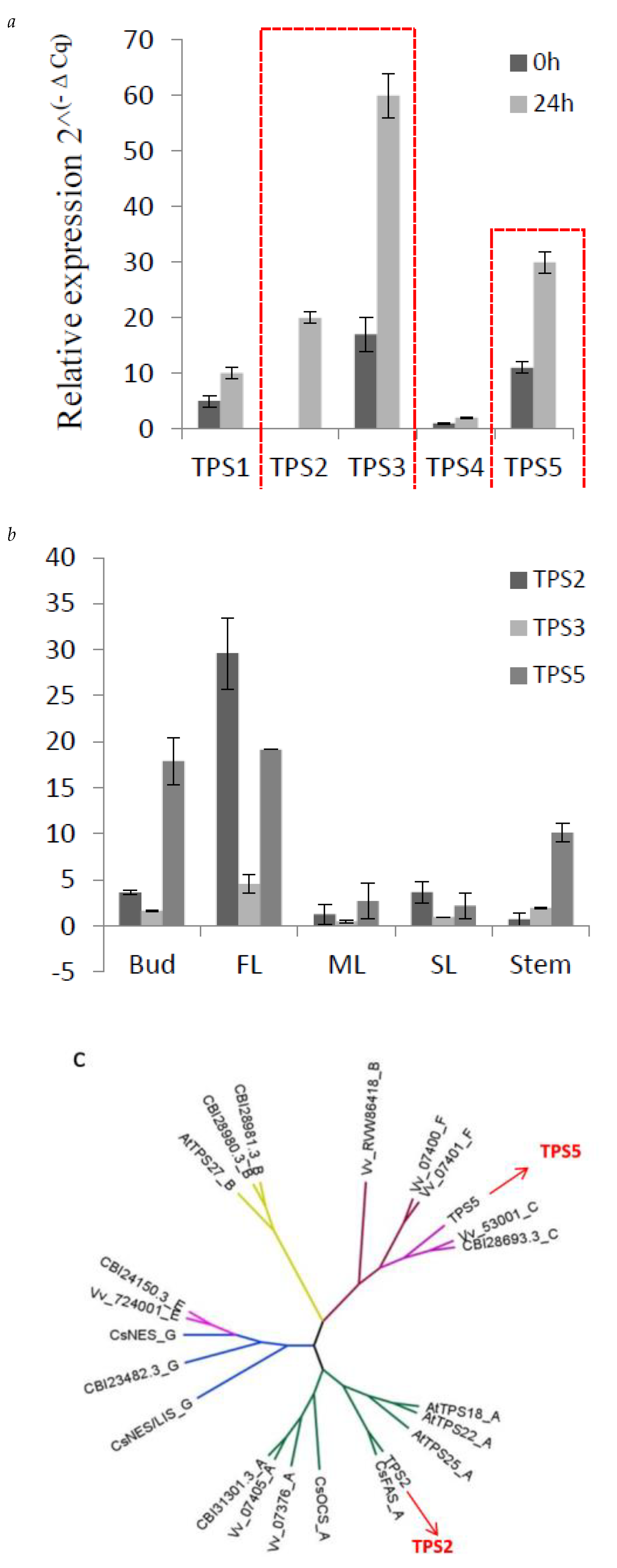
2.4. Expression of Three TPSs in Different Tissues and Their Response to MeJA
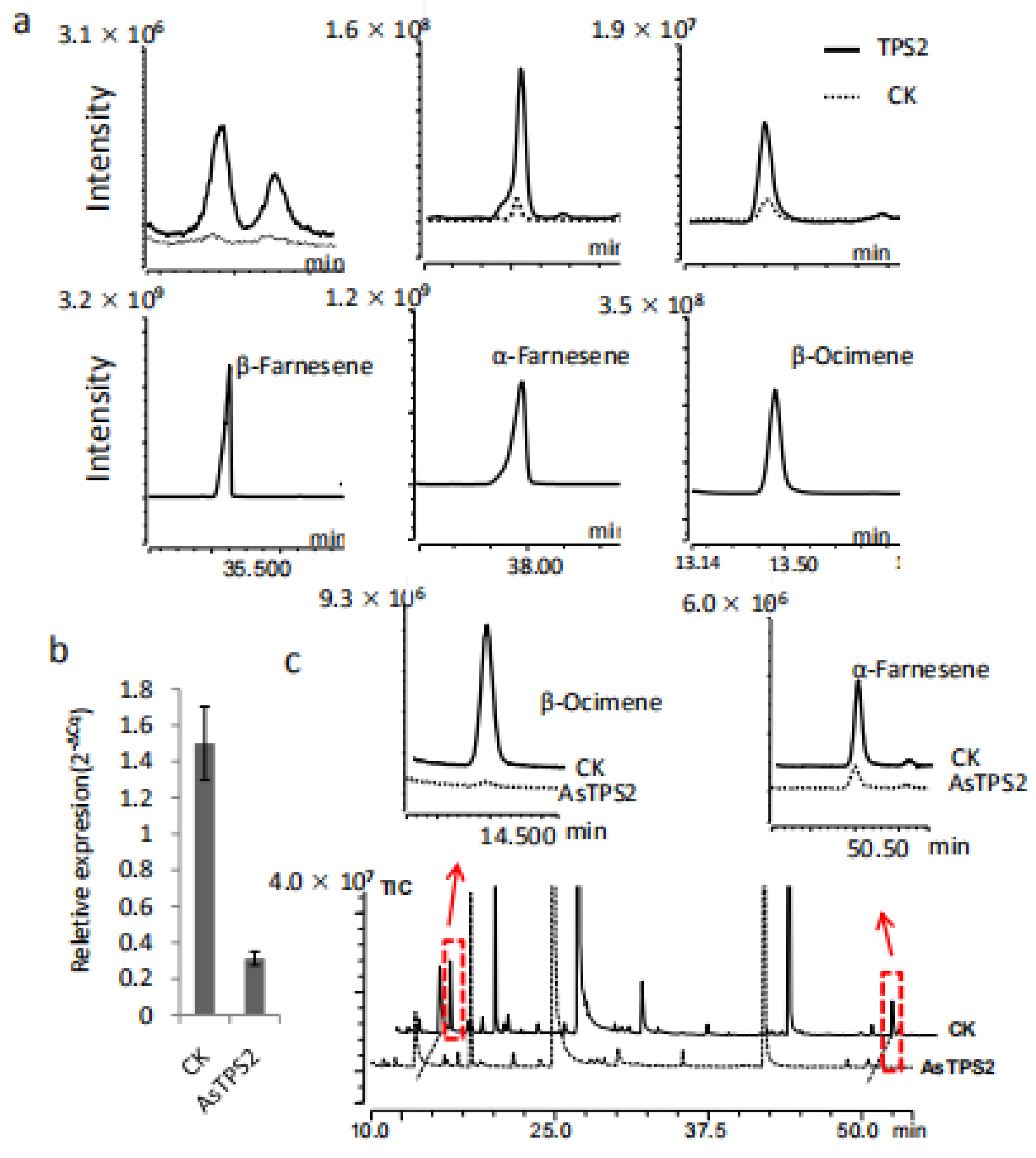
2.5. TPS2 Catalyze α/β-Frnesene and β-Ocimene Formation In Vitro
2.6. Functional Analysis of TPS2 in Tea Plants
3. Materials and Methods
3.1. Plant Materials and Chemicals
3.2. RNA Isolation and cDNA Library Construction
3.3. Transcript Assembly and Annotation
3.4. GC–MS Analysis of Volatile Compounds in Samples
3.5. Transcript Expression Analysis of Genes Involved in Terpenes
3.6. Integration of Gene Expression and Terpenoid Molecule Profiles
3.7. Methyl Jasmonate Treatments
3.8. Gene Cloning and Prokaryotic Expression
3.9. Enzyme Assay
3.10. TPS2 Enzyme Assay in Camellia Sinensis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| AACT | Acetoacetyl-CoA thiolase |
| AsODNs | Antisense oligonucleotides |
| At | Arabidopsis thaliana |
| CsAFS | α-farnesene synthase in Camellia sinensis |
| CMK | Cytidyl (4-diphospho)-2-C-methyl-d-erythritol kinase |
| CsNES | Nerolidol synthase in Camellia sinensis |
| CsNES/LIS | Nerolidol/linalool synthase in Camellia sinensis |
| CsOCS | β-Ocimene synthase in Camellia sinensis |
| DMAPP | Dimethylallyl pyrophosphate |
| DMNT | 4,8-Dimethylnona-1,3,7-triene |
| DXR | 1-Deoxy-d-xylulose5-phosphate reductase |
| DXS | 1-Deoxy-d-xylulose-5-phosphate synthase |
| FAS | Farnesene synthase |
| FDS | Farnesyl diphosphate |
| FPP | Farnesyl pyrophosphate |
| FL | First leaf |
| GC–MS | Gas chromatography-mass spectrometry |
| GDS | Geranyl diphosphate |
| GPP | Geranyl pyrophosphate |
| HDR | 4-Hydroxy-3-methylbut-2-enyldiphosphate reductase |
| HDS | 1-Hydroxyl-2-methyl-2-(E)-butenyl-diphosphate synthase |
| HMGS | 3-Hydroxy-3-methylglutaryl-coenzyme (a synthase) |
| HMGR | 3-Hydroxy-3-methylglutaryl coenzyme (a reductase) |
| HUS | Humulene synthase |
| IDI | Isopentenyl pyrophosphate isomerase |
| IPP | Isopentenyl pyrophosphate |
| LIS | Linalool synthase |
| LS | Limonene synthase |
| MCT | 2-C-Methyl-d-erythritol-4-phosphate cytidyltransferase |
| MDC | Mevalonate-5-phosphate decarboxylase |
| MDS | 2-C-Methyl-d-erythritol-2,4-cyclodiphohate synthase |
| MEP | Methylerythritol phosphate |
| MeJA | Methyl jasmonate |
| ML | Mature leaf |
| MVA | Mevalonic acid |
| MVK | Mevalonate kinase |
| MYS | β-Myrcene synthase |
| NUDX1 | Nudix hydrolase 1 |
| NES | Nerolidol synthase |
| OCS | β-Ocimene synthase |
| PMK | 5-Phosphatemevalonate kinase |
| PCA | Principle component analysis |
| Q-PCR | Quantitative PCR |
| RPKM | Reads per kilobase per million |
| SQS | Squalene synthase |
| SL | Second leaf |
| SPME | Solid-phase microextraction |
| TPSs | Terpene synthases |
| Vv | Vitis vinifera |
References
- Yang, Z.; Baldermann, S.; Watanabe, N. Recent Studies of the Volatile Compounds in Tea. Food Res. Int. 2013, 53, 585–599. [Google Scholar] [CrossRef]
- Wang, D.; Ando, K.; Morita, K.; Kubota, K.; Kobayashi, A. Optical Isomers of Linalool and Linalool Oxides in Tea Aroma. Biosci. Biotechnol. Biochem. 1994, 58, 2050–2053. [Google Scholar] [CrossRef] [Green Version]
- Chieberle, P.E.S. Characterization of the Key Aroma Compounds in the Beverage Prepared from Darjeeling Black Tea : Quantitative Differences between Tea Leaves and Infusion AND. J. Agric. Food Chem. 2006, 54, 916–924. [Google Scholar]
- Joshi, R.; Gulati, A. Scientia Horticulturae Biochemical Attributes of Tea Flowers (Camellia Sinensis) at Different Developmental Stages in the Kangra Region of India. Sci. Horticulturae. 2011, 130, 266–274. [Google Scholar] [CrossRef]
- Song, C.; Hartl, K.; Kata, M.; Hoffmann, T.; Schwab, W. Attractive but Toxic: Emerging Roles of Glycosyltransferases Involved in Their Formation. Mol. Plant 2018, 11, 1225–1236. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pichersky, E.; Gershenzon, J. The Formation and Function of Plant Volatiles : Perfumes for Pollinator Attraction and Defense. Curr. Opin. Plant Biol. 2002, 5, 237–243. [Google Scholar] [CrossRef]
- Dicke, M.; Baldwin, I.T. The Evolutionary Context for Herbivore-Induced Plant Volatiles : Beyond the ‘Cry for Help’. Trends Plant Sci. 2010, 15, 167–175. [Google Scholar] [CrossRef]
- Godard, K.; White, R.; Bohlmann, J. Phytochemistry Monoterpene -Induced Molecular Responses in Arabidopsis Thaliana. Phytochemistry 2008, 69, 1838–1849. [Google Scholar] [CrossRef]
- Dong, F.; Yang, Z.; Baldermann, S.; Sato, Y.; Asai, T.; Watanabe, N. Herbivore-Induced Volatiles from Tea Camellia Sinensis) Plants and Their Involvement in Intraplant Communication and Changes in Endogenous Nonvolatile Metabolites. J. Agric. Food Chem. 2011, 59, 13131–13135. [Google Scholar] [CrossRef]
- Lee, S.; Badieyan, S.; Bevan, D.R.; Herde, M.; Gatz, C.; Tholl, D. Herbivore-Induced and Fl Oral Homoterpene Volatiles Are Biosynthesized by a Single P450 Enzyme (CYP82G1) in Arabidopsis. Proc. Natl. Acad. Sci. USA 2010, 107, 21205–21210. [Google Scholar] [CrossRef] [Green Version]
- Chem, F. Composition of the Volatiles from Intact and Mechanically Pierced Tea Aphid − Tea Shoot Complexes and Their Attraction. J. Agric. Food Chem. 2002, 50, 2571–2575. [Google Scholar]
- Parks, W.P.; Todaro, G.J.; Aaronson, S.A.; Virol, J. Aphid Alarm Pheromone : Isolation, Identification, Synthesis Relation between REM Sleep and Intracranial Self-Stimulation. Science 1972, 177, 1121–1122. [Google Scholar]
- Boo, K.S.; Chung, I.B.; Han, K.S.; Pickett, J.A.; Wadhams, L.J. RESPONSE OF THE LACEWING Chrysopa Cognata TO PHEROMONES OF ITS APHID PREY. J. Chem. Ecol. 1998, 24, 631–643. [Google Scholar] [CrossRef]
- Gibson, R.W.; Pickett, J.A. Wild Potato Repels Aphids by Release of Aphid Alarm Pheromone. Nature 1983, 302, 608–609. [Google Scholar] [CrossRef]
- Rohmer, M.; Knani, T.M.; Simonin, P.; Sutter, B.; Sahmt, H.; Nationale, E.; De Chimie, S.; Werner, A.; Cedex, F.M. Isoprenoid Biosynthesis in Bacteria : A Novel Pathway for the Early Steps Leading to Isopentenyl Diphosphate. Biochem. J. 1993, 295, 517–524. [Google Scholar] [CrossRef]
- Pathway, C.; Newman, J.D.; Chappell, J. Isoprenoid Biosynthesis in Plants : Carbon Partitioning Within the Cytoplasmic Pathway. Crit. Rev. Biochem. Mol. Biol. 1999, 34, 95–106. [Google Scholar]
- Zhou, Y.; Zeng, L.; Liu, X.; Gui, J.; Mei, X.; Fu, X.; Dong, F.; Tang, J.; Zhang, L.; Yang, Z. Formation of (E) -Nerolidol in Tea (Camellia Sinensis) Leaves Exposed to Multiple Stresses during Tea Manufacturing. Food Chem. 2017, 231, 78–86. [Google Scholar] [CrossRef]
- Zhao, M.; Zhang, N.; Gao, T.; Jin, J.; Jing, T.; Wang, J.; Wu, Y.; Wan, X.; Schwab, W.; Song, C. Sesquiterpene Glucosylation mediated by glucosyltransferase UGT91Q2 is involved in the modulation of cold stress tolerance in tea plants. New Phytol. 2019. [Google Scholar] [CrossRef]
- Xu, Q.; He, Y.; Yan, X.; Zhao, S.; Zhu, J.; Wei, C. Unraveling a Crosstalk Regulatory Network of Temporal Aroma Accumulation in Tea Plant (Camellia Sinensis) Leaves by Integration of Metabolomics and Transcriptomics. Environ. Exp. Bot. 2018, 149, 81–94. [Google Scholar] [CrossRef]
- Wang, X.; Zeng, L.; Liao, Y.; Li, J.; Tang, J. Formation of α -Farnesene in Tea (Camellia Sinensis) Leaves Induced by Herbivore-Derived Wounding and Its Effect on Neighboring Tea Plants. Int. J. Mol. Sci. 2019, 20, 4151. [Google Scholar] [CrossRef] [Green Version]
- Falara, V.; Akhtar, T.A.; Nguyen, T.T.H.; Spyropoulou, E.A.; Bleeker, P.M.; Schauvinhold, I.; Matsuba, Y.; Bonini, M.E.; Schilmiller, A.L.; Last, R.L.; et al. The Tomato Terpene Synthase Gene Family. Plant Physiol. 2011, 157, 770–789. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meyer, O.; Tritsch, D.; Kagan, I.A.; Grosdemange-billiard, C.; Rohmer, M.; Bach, T.J. Cross-Talk between the Cytosolic Mevalonate and the Plastidial Methylerythritol Phosphate Pathways in Tobacco Bright Yellow-2 Cells. J. Biol. Chem. 2003, 278, 26666–26676. [Google Scholar]
- Dudareva, N.; Andersson, S.; Orlova, I.; Gatto, N.; Reichelt, M.; Rhodes, D.; Boland, W.; Gershenzon, J. The Nonmevalonate Pathway Supports Both Monoterpene and Sesquiterpene Formation in Snapdragon Flowers. Proc. Natl. Acad. Sci. USA 2005, 102, 933–938. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nagegowda, D.A. Plant Volatile Terpenoid Metabolism : Biosynthetic Genes, Transcriptional Regulation and Subcellular Compartmentation. FEBS Lett. 2010, 584, 2965–2973. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nagegowda, D.; Plants, A. Plant Volatiles : Recent Advances and Future Perspectives Critical Reviews in Plant Sciences Plant Volatiles : Recent Advances and Future. Crit. Rev. Plant Sci. 2015, 25, 417–440. [Google Scholar]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular Evolutionary Genetics Analysis Version 7.0 for Bigger Datasets. Mol. Biol. Evolution. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [Green Version]
- Bacteria, B.; Metabolic, P.A.; Achieved, M.; Rodrı, M. Elucidation of the Methylerythritol Phosphate Pathway for Isoprenoid Update on Isoprenoid Biosynthesis Elucidation in Bacteria and Plastids. A Metabolic Milestone Achieved through Genomics. Plant Physiol. 2002, 130, 1079–1089. [Google Scholar]
- Fu, X.; Chen, Y.; Mei, X.; Katsuno, T.; Kobayashi, E.; Dong, F.; Watanabe, N.; Yang, Z. Regulation of Formation of Volatile Compounds of Tea (Camellia Sinensis) Leaves by Single Light Wavelength. Sci. Rep. 2015, 5, 16858. [Google Scholar] [CrossRef]
- De la Peña Moreno, F.; Blanch, G.P.; del Castillo, M.L.R. Effect of (−)- and (+)-Methyl Jasmonate on the Bioformation of Aroma-Active Esters in Strawberry Fruits. Eur. Food Res. Technol. 2010, 231, 829–834. [Google Scholar] [CrossRef]
- Matarese, F.; Cuzzola, A.; Scalabrelli, G.; D’Onofrio, C. Expression of terpene synthase genes associated with the formation of volatiles in different organs of Vitis iniera. Phytochemistry 2014, 105, 12–24. [Google Scholar] [CrossRef]
- Magrane, M.; Consortium, U. Original Article UniProt Knowledgebase : A Hub of Integrated Protein Data. Database 2011, 2011, bar009. [Google Scholar] [CrossRef] [Green Version]
- Livak, K.J.; Schmittgen, T.D. Analysis of Relative Gene Expression Data Using Real- Time Quantitative PCR and the 2 –ΔΔCT Method. Methods. 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Jing, T.; Chen, Y.; Song, C. Glucosylation of (Z)-3-Hexenol Informs Intraspecies Interactions in Plants : A Case Study in Camellia Sinensis. Plant Cell Environ. 2018, 42, 1352–1367. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Monoterpenes(26) | Chemical Formulas | Tissues | Types and Number |
|---|---|---|---|
| β-Myrcene * | C10H16 | All tissues | Acyclic(17) |
| trans-β-Ocimene * | C10H16 | Bud, FL, SL, Stem | |
| Linalool * | C10H18O | All tissues | |
| Geraniol * | C10H18O | All tissues | |
| α-Citral * | C10H16O | All tissues | |
| β-Citral * | C10H16O | All tissues | |
| Methyl geranate * | C11H18O2 | ML, SL | |
| Nerol * | C10H18O | All tissues | |
| 8-Hydroxylinalool | C10H18O2 | All tissues | |
| 3,7-Octadiene-2,6-di-ol, 2,6-dimethyl | C10H18O2 | FL, SL, Stem | |
| neryl acetate * | C12H20O2 | FL | |
| Isogeraniol | C10H18O | All tissues | |
| γ-Isogeraniol | C10H18O | All tissues | |
| Geranyl acetone * | C13H22O | All tissues | |
| Citronellyl formate | C11H20O2 | All tissues | |
| DMDD | C10H18O2 | All tissues | |
| Hotrienol | C10H16O | All tissues | |
| D-Limonene * | C10H16 | All tissues | Monocyclic(6) |
| trans-Linalool oxide (furanoid) * | C10H18O2 | All tissues | |
| L-Terpineol * | C10H18O | All tissues | |
| α-Terpinen * | C10H16 | All tissues | |
| 4-Terpinenyl acetate | C12H20O2 | All tissues | |
| Thymol * | C10H14O | Stem | |
| 2,3-Dehydro-1,8-cineole | C10H16O | Stem | Bicyclic(3) |
| (+)-Camphene | C10H16 | All tissues | |
| trans-(−)-Pinocarveo | C10H16O | Stem |
| Sesquiterpenes(14) | Chemical Formulas | Tissues | Types and Number |
|---|---|---|---|
| (Z)-Nerolidol * | C15H26O | All tissues | Acyclic(3) |
| α-Farnesene * | C15H24 | All tissues | |
| (E)-Nerolidol * | C15H26O | All tissues | |
| (E)-Farnesene epoxide * | C15H24O | Bud, FL, SL, Stem | Monocyclic(5) |
| Humulene * | C15H24 | Bud, FL, SL, Stem | |
| α-ionone * | C13H20O | ML, Stem | |
| β-ionone * | C13H20O | All tissues | |
| (E)-γ-Bisabolene * | C15H24 | Bud, FL, Stem | Bicyclic(4) |
| trans-Bergamotene | C15H24 | All tissues | |
| α-Cadinol | C15H26O | Bud, FL, SL, Stem | |
| T-Cadinol * | C15H26O | Bud, FL, SL, Stem | |
| Caryophyllene * | C15H24 | Bud, FL, SL, Stem | |
| α-Cubebene | C15H24 | All tissues | Tricyclic(2) |
| α-Copaene | C15H24 | Bud, FL, SL, Stem | |
| Squalene | C30H50 | Bud, FL | Triterpene(1) |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jin, J.; Zhang, S.; Zhao, M.; Jing, T.; Zhang, N.; Wang, J.; Wu, B.; Song, C. Scenarios of Genes-to-Terpenoids Network Led to the Identification of a Novel α/β-Farnesene/β-Ocimene Synthase in Camellia sinensis. Int. J. Mol. Sci. 2020, 21, 655. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21020655
Jin J, Zhang S, Zhao M, Jing T, Zhang N, Wang J, Wu B, Song C. Scenarios of Genes-to-Terpenoids Network Led to the Identification of a Novel α/β-Farnesene/β-Ocimene Synthase in Camellia sinensis. International Journal of Molecular Sciences. 2020; 21(2):655. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21020655
Chicago/Turabian StyleJin, Jieyang, Shangrui Zhang, Mingyue Zhao, Tingting Jing, Na Zhang, Jingming Wang, Bin Wu, and Chuankui Song. 2020. "Scenarios of Genes-to-Terpenoids Network Led to the Identification of a Novel α/β-Farnesene/β-Ocimene Synthase in Camellia sinensis" International Journal of Molecular Sciences 21, no. 2: 655. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21020655