Impact of the Stroma on the Biological Characteristics of the Parenchyma in Oral Squamous Cell Carcinoma

 , , ,
, , , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Properties of Various Types of Stromal Cells
2.2. 2D Co-Culture of HSC-2 Cells and Various Types of Stromal Cells
2.2.1. Effect of Stromal Conditioned Medium (CM) on the Growth of HSC-2 Cells
2.2.2. Ki-67 Labeling Index
2.2.3. Stromal Cells Enhance the Invasive Ability of Cancer Cells
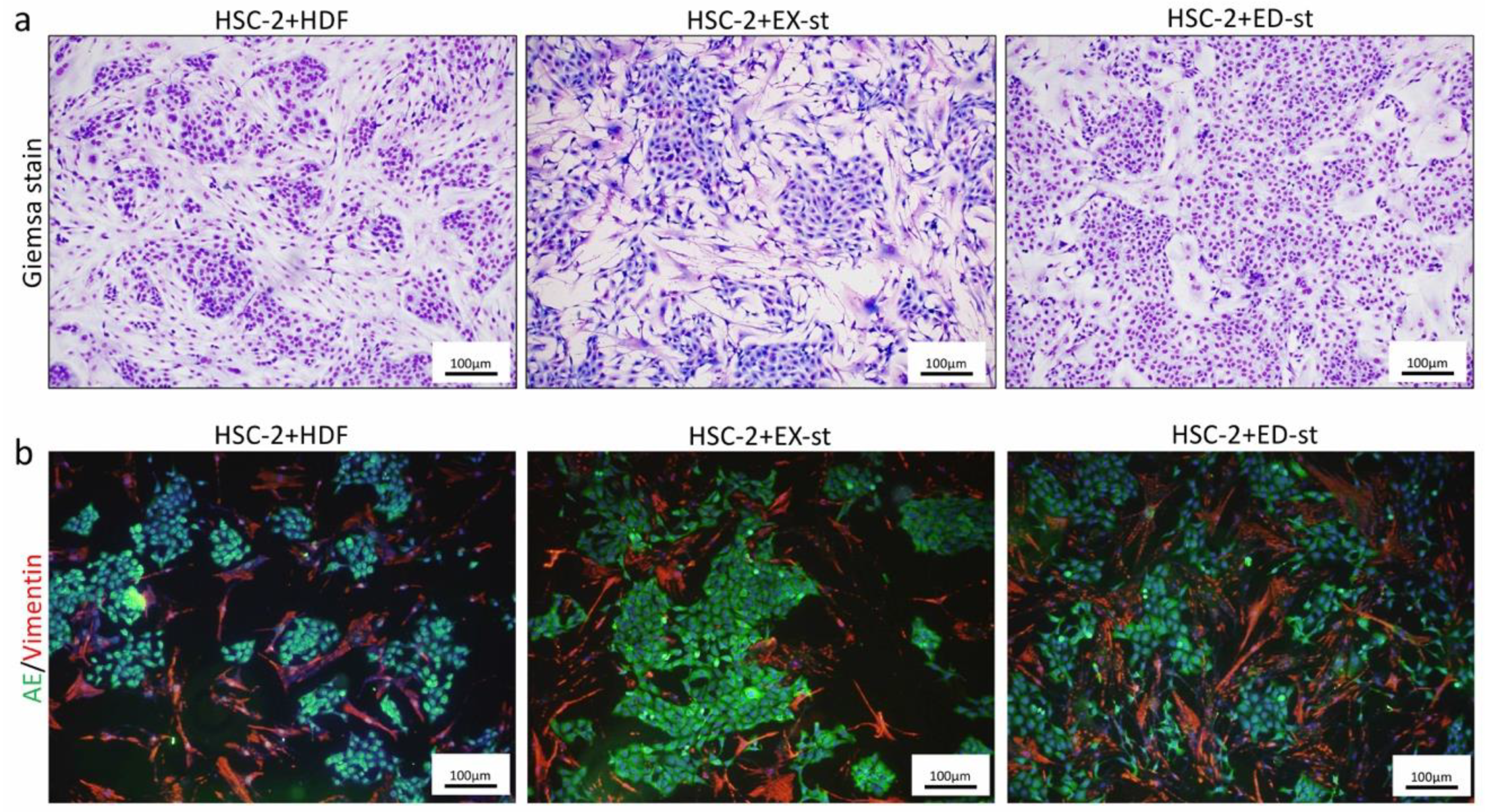
2.2.4. Comparison of the Morphology in 2D Co-Culture of HSC-2 Cells and Various Types of Stromal Cells
2.3. Tumor Xenograft Model
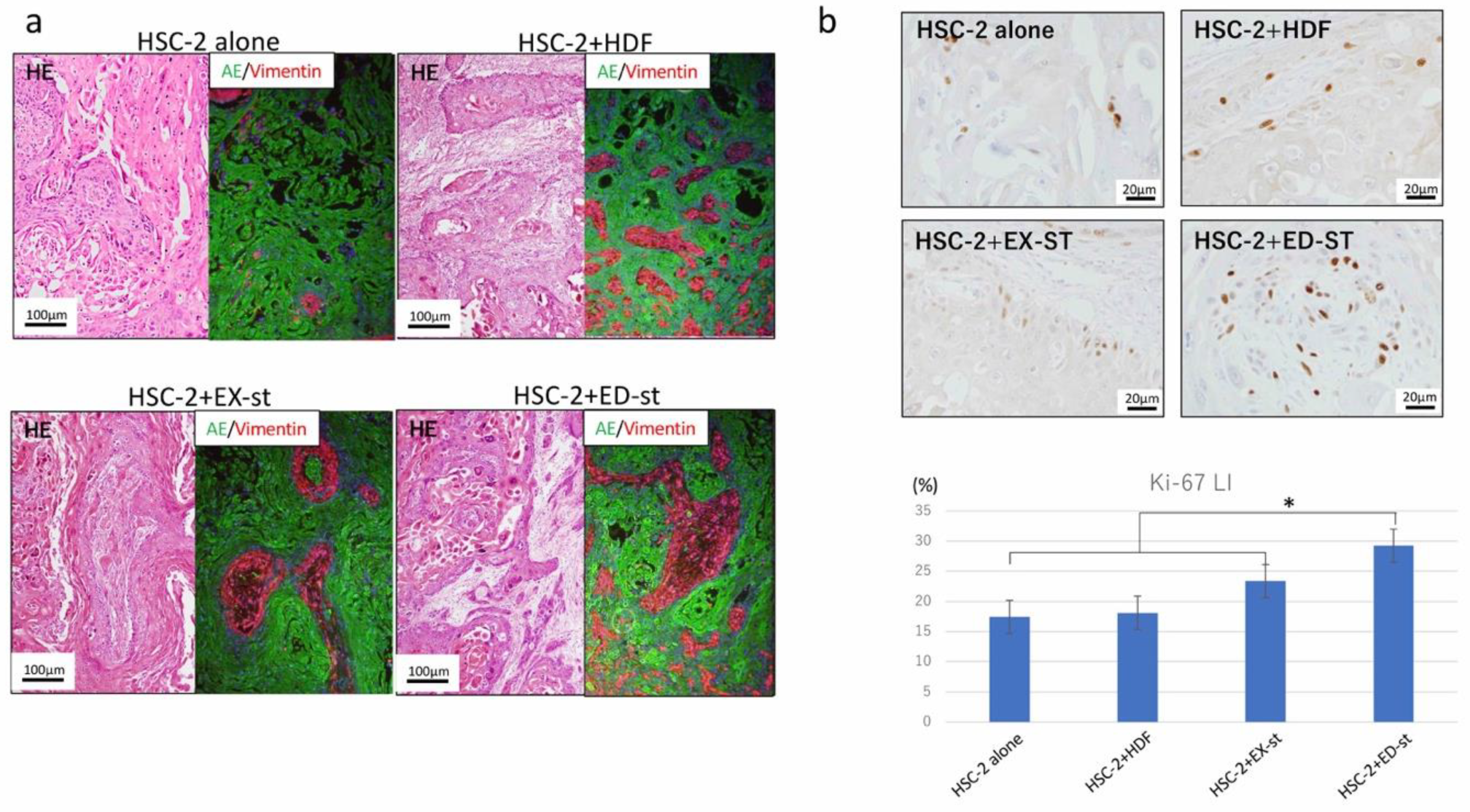
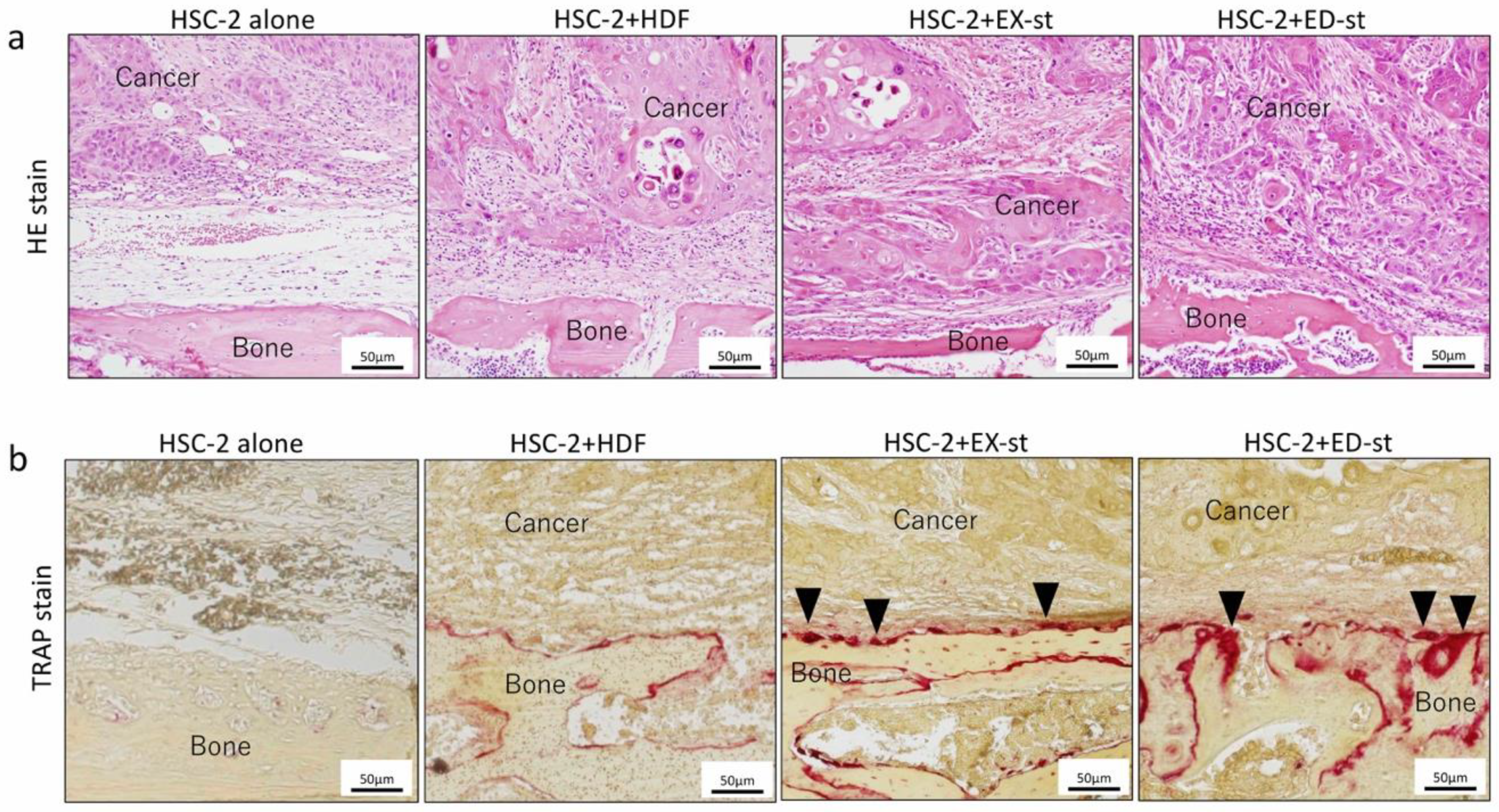
2.3.1. Histological Findings
2.3.2. Differences in the Ki-67-Positive Rate among Different Types of Stroma
2.3.3. Effect of Different Stroma Properties on Bone Resorption
3. Discussion
3.1. Properties of the Tumor Stroma Isolated from Different Subtypes of OSCCs
3.2. Effect of the Tumor Stroma on the Proliferative and Invasive Abilities of the Tumor Parenchyma
3.3. Effect of the Tumor Stroma on the Morphology of the Tumor Parenchyma
4. Materials and Methods
4.1. OSCC Cell Line
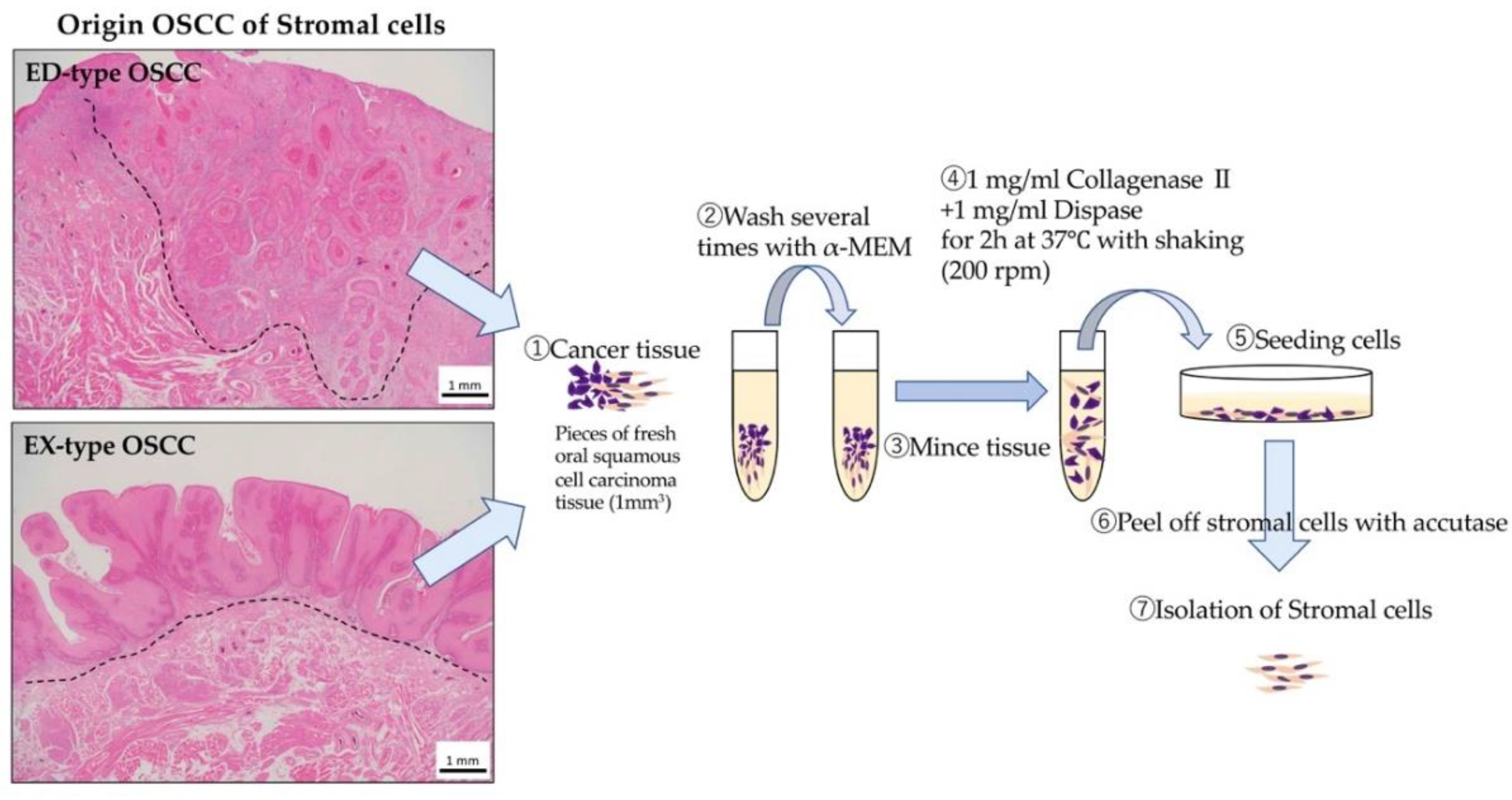
4.2. Stroma Isolation
4.3. In Vitro Experiment
4.3.1. Tumor Stromal Properties
Tumor Stromal Morphology
Tumor Stroma Proliferation Assay (MTS Assay)
4.3.2. Effect of Stromal Conditioned Medium (CM) on HSC-2 Cell Proliferation
4.4. Co-Culture In Vitro
4.4.1. Cell Morphology
4.4.2. Double-Fluorescent Immunohistochemical Staining
4.4.3. Invasion Assay
4.5. In Vivo Experiment (Tumor Xenografts)
4.5.1. Experimental Animals
4.5.2. Histological Examination
4.5.3. Cell Proliferation Assay (Ki-67 Labeling Index)
4.5.4. Tartrate-Resistant Acid Phosphatase (TRAP) Staining
4.6. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Joyce, J.A.; Pollard, J.W. Microenvironmental Regulation of Metastasis. Nat. Rev. Cancer 2009, 9, 239–252. [Google Scholar] [CrossRef]
- De Wever, O.; Mareel, M. Role of Tissue Stroma in Cancer Cell Invasion. J. Pathol. 2003, 200, 429–447. [Google Scholar] [CrossRef]
- Harper, J.; Sainson, R.C.A. Regulation of the Anti-Tumour Immune Response by Cancer-Associated Fibroblasts. Semin. Cancer Biol. 2014, 25, 69–77. [Google Scholar] [CrossRef]
- Tripathi, M.; Billet, S.; Bhowmick, N.A. Understanding the Role of Stromal Fibroblasts in Cancer Progression. Cell Adhes. Migr. 2012, 6, 231–235. [Google Scholar] [CrossRef]
- Fuyuhiro, Y.; Yashiro, M.; Noda, S.; Matsuoka, J.; Hasegawa, T.; Kato, Y.; Sawada, T.; Hirakawa, K. Cancer-associated Orthotopic Myofibroblasts Stimulates the Motility of Gastric Carcinoma Cells. Cancer Sci. 2012, 103, 797–805. [Google Scholar] [CrossRef]
- Yashiro, M.; Hirakawa, K. Cancer-stromal Interactions in Scirrhous Gastric Carcinoma. Cancer Microenviron. 2010, 3, 127–135. [Google Scholar] [CrossRef] [Green Version]
- Mbeunkui, F.; Johann, D.J., Jr. Cancer and the Tumor Microenvironment: A Review of an Essential Relationship. Cancer Chemother. Pharmacol. 2009, 63, 571–582. [Google Scholar] [CrossRef] [Green Version]
- Costea, D.E.; Hills, A.; Osman, A.H.; Thurlow, J.; Kalna, G.; Huang, X.; Murillo, C.P.; Parajuli, H.; Suliman, S.; Kulasekara, K.K.; et al. Identification of Two Distinct Carcinoma-Associated Fibroblast Subtypes with Differential Tumor-Promoting Abilities in Oral Squamous Cell Carcinoma. Cancer Res. 2013, 73, 3888–3901. [Google Scholar] [CrossRef] [Green Version]
- Routray, S.; Sunkavali, A.; Bari, K. Carcinoma-associated Fibroblasts, Its Implication in Head and Neck Squamous Cell Carcinoma: A Mini Review. Oral Dis. 2014, 20, 246–253. [Google Scholar] [CrossRef]
- Elaoui, D.; Movahedi, K.; Van Overmeire, E.; Bossche, J.V.D.; Schouppe, E.; Mommer, C.; Nikolaou, A.; Morias, Y.; De Baetselier, P.; Van Ginderachter, J.A. Tumor-associated Macrophages in Breast Cancer: Distinct Subsets, Distinct Functions. Int. J. Dev. Biol. 2011, 55, 861–867. [Google Scholar]
- Piao, Y.; Park, S.Y.; Henry, V.; Smith, B.D.; Tiao, N.; Flynn, D.L.; de Groot, J.F. Novel MET/TIE2/VEGFR2 Inhibitor Altiratinib Inhibits Tumor Growth and Invasiveness in Bevacizumab-Resistant Glioblastoma Mouse Models. Neuro-Oncology 2016, 18, 1230–1241. [Google Scholar] [CrossRef] [Green Version]
- Rothschild, E.; Banerjee, D. Subverting Subversion: A Review on the Breast Cancer Microenvironment and Therapeutic Opportunities. Breast Cancer Basic Clin. Res. 2015, 9, 7–15. [Google Scholar] [CrossRef]
- World Health Organization. Classification of Head and Neck Tumours; World Health Organization: Geneva, Switzerland, 2017; pp. 105–131. [Google Scholar]
- Jakobsson, P.Å.; Eneroth, C.M.; Killander, D.; Moberger, G.; Mårtensson, B. Histological classification and grading of malignancy in carcinoma of the larynx. Acta Radiol. Ther. Phys. Biol. 1973, 12, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Yılmaz, T.; Hoşal, A.Ş.; Gedikoglu, G.; Kaya, S.; Yılmaz, T. Prognostic significance of histopathological parameters in cancer of larynx. Eur. Arch. Otorhinolaryngol. 1999, 256, 139–144. [Google Scholar] [CrossRef]
- Patel, S.G.; Shah, J.P. TNM staging of cancers of the head and neck: Striving for uniformity amoung diversity. CA Cancer J. Clin. 2005, 55, 242–258. [Google Scholar] [CrossRef] [Green Version]
- Spiro, R.H.; Guillamondegui, O.; Paulino, A.F.; Huvos, A.G. Pattern of invasion and margin assessment in patients with oral tongue cancer. Head Neck. 1999, 21, 408–413. [Google Scholar] [CrossRef]
- Hussein, M.R.; Cullen, K. Molecular biomarkers in HNSCC: Prognostic and therapeutic implications. Expert Rev. Anticancer Ther. 2002, 1, 116–124. [Google Scholar] [CrossRef]
- Robinson, C.M.; Prime, S.S.; Paterson, I.C.; Guest, P.G.; Eveson, J.W. Expression of Ki-67 and p53 in cutaneous free flaps used to reconstruct soft tissue defects following resection of oral squamous cell carcinoma. Oral Oncol. 2007, 43, 263–271. [Google Scholar] [CrossRef]
- Cymerman, J.A.; Kulkarni, R.; Gouldesbrough, D.; McCaul, J.A. First report of squamous cell carcinoma arising within an intraoral radial forearm free flap. Int. J. Surg. Case Rep. 2013, 4, 731–734. [Google Scholar] [CrossRef] [Green Version]
- Hanahan, D.; Coussens, L.M. Accessories to the Crime: Functions of Cells Recruited to the Tumor Microenvironment. Cancer Cell 2012, 21, 309–322. [Google Scholar] [CrossRef] [Green Version]
- Kalluri, R.; Zeisberg, M. Fibroblasts in Cancer. Nat. Rev. Cancer 2006, 6, 392–401. [Google Scholar] [CrossRef]
- Ostman, A.; Eaugsten, M. Cancer-associated Fibroblasts and Tumor Growth--Bystanders Turning into Key Players. Curr. Opin. Genet. Dev. 2009, 19, 67–73. [Google Scholar] [CrossRef]
- Anderberg, C.; Pietras, K. On the Origin of Cancer-Associated Fibroblasts. Cell Cycle 2009, 8, 1461–1462. [Google Scholar] [CrossRef] [Green Version]
- Boras, V.V.; Fucic, A.; Virag, M.; Gabrić, D.; Blivajs, I.; Tomasovic-Loncaric, C.; Rakusić, Z.; Bišof, V.; Le Novere, N.; Vrdoljak, D.V. Significance of Stroma in Biology of Oral Squamous Cell Carcinoma. Tumori J. 2018, 104, 9–14. [Google Scholar] [CrossRef]
- Thuwajit, C.C.P.; Paupairoj, A.; Chau-In, S.; Suthiphongchai, T.; Thuwajit, C. Alpha-smooth Muscle Actin-Positive Fibroblasts Promote Biliary Cell Proliferation and Correlate with Poor Survival in Cholangiocarcinoma. Oncol. Rep. 2009, 21, 957–969. [Google Scholar] [CrossRef] [Green Version]
- Parikh, J.G.; Kulkarni, A.; Johns, C. α-Smooth Muscle Actin-Positive Fibroblasts Correlate with Poor Survival in Hepatocellular Carcinoma. Oncol. Lett. 2014, 7, 573–575. [Google Scholar] [CrossRef] [Green Version]
- Sinn, M.; Denkert, C.; Striefler, J.K.; Pelzer, U.; Stieler, J.M.; Bahra, M.; Lohneis, P.; Dörken, B.; Oettle, H.; Riess, H.; et al. α-Smooth Muscle Actin Expression and Desmoplastic Stromal Reaction in Pancreatic Cancer: Results From the CONKO-001 Study. Br. J. Cancer 2014, 111, 1917–1923. [Google Scholar] [CrossRef] [Green Version]
- Ding, L.; Zhang, Z.; Shang, D.; Cheng, J.; Yuan, H.; Wu, Y.; Song, X.; Jiang, H. α-Smooth Muscle Actin-Positive Myofibroblasts, in Association with Epithelial-Mesenchymal Transition and Lymphogenesis, Is a Critical Prognostic Parameter in Patients With Oral Tongue Squamous Cell Carcinoma. J. Oral Pathol. Med. 2014, 43, 335–343. [Google Scholar] [CrossRef]
- Wang, W.-Q.; Liu, L.; Xu, H.-X.; Luo, G.-P.; Chen, T.; Wu, C.-T.; Xu, Y.-F.; Xu, J.; Liu, C.; Zhang, B.; et al. Intratumoral α-SMA Enhances the Prognostic Potency of CD34 Associated with Maintenance of Microvessel Integrity in Hepatocellular Carcinoma and Pancreatic Cancer. PLoS ONE 2013, 8, e71189. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.; Zou, L.; Zhang, Y.; Chen, Y.; Xing, P.; Yang, W.; Li, F.; Ji, X.; Liu, F.; Lu, X. Transforming Growth factor-β1 and α-Smooth Muscle Actin in Stromal Fibroblasts Are Associated with a Poor Prognosis in Patients with Clinical Stage I-IIIA Nonsmall Cell Lung Cancer After Curative Resection. Tumor Biol. 2014, 35, 6707–6713. [Google Scholar] [CrossRef]
- Choi, S.-Y.; Sung, R.; Lee, S.-J.; Lee, T.-G.; Kim, N.; Yoon, S.M.; Lee, E.J.; Chae, H.B.; Youn, S.J.; Park, S.M. Podoplanin, α-Smooth Muscle Actin or S100A4 Expressing Cancer-Associated Fibroblasts Are Associated With Different Prognosis in Colorectal Cancers. J. Korean Med. Sci. 2013, 28, 1293–1301. [Google Scholar] [CrossRef] [Green Version]
- Hanahan, D.; Weinberg, R.A. Hallmarks of Cancer: The Next Generation. Cell 2011, 144, 646–667. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yurugi, Y.; Wakahara, M.; Matsuoka, Y.; Sakabe, T.; Kubouch, Y.; Haruki, T.; Nosaka, K.; Miwa, K.; Araki, K.; Taniguchi, Y.; et al. Podoplanin Expression in Cancer-associated Fibroblasts Predicts Poor Prognosis in Patients with Squamous Cell Carcinoma of the Lung. Anticancer Res. 2017, 37, 207–213. [Google Scholar] [CrossRef]
- Kuemmerle-Deschner, J.B.; Wittkowski, H.; Tyrrell, P.N.; Koetter, I.; Lohse, P.; Ummenhofer, K.; Reess, F.; Hansmann, S.; Koitschev, A.; Deuter, C.; et al. Treatment of Muckle-Wells Syndrome: Analysis of Two IL-1-blocking Regimens. Treatment of Muckle-Wells Syndrome: Analysis of Two IL-1-blocking Regimens. Arthritis Res. Ther. 2013, 15, R64. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kato, T.; Noma, K.; Ohara, T.; Kashima, H.; Katsura, Y.; Sato, H.; Komoto, S.; Katsube, R.; Ninomiya, T.; Tazawa, H.; et al. Cancer-Associated Fibroblasts Affect Intratumoral CD8 + and FoxP3 + T Cells Via IL6 in the Tumor Microenvironment. Clin. Cancer Res. 2018, 24, 4820–4833. [Google Scholar] [CrossRef] [Green Version]
- Wang, R.-F.; Zhang, L.-H.; Shan, L.-H.; Sun, W.-G.; Chai, C.-C.; Wu, H.-M.; Ibla, J.C.; Wang, L.-F.; Liu, J.-R. Effects of the Fibroblast Activation Protein on the Invasion and Migration of Gastric Cancer. Exp. Mol. Pathol. 2013, 95, 350–356. [Google Scholar] [CrossRef] [PubMed]
- Hwang, R.F.; Moore, T.; Arumugam, T.; Ramachandran, V.; Amos, K.D.; Rivera, A.; Ji, B.; Evans, D.B.; Logsdon, C.D. Cancer-associated Stromal Fibroblasts Promote Pancreatic Tumor Progression. Cancer Res. 2008, 68, 918–926. [Google Scholar] [CrossRef] [Green Version]
- Orimo, A.; Gupta, P.B.; Sgroi, D.C.; Arenzana-Seisdedos, F.; Delaunay, T.; Naeem, R.; Carey, V.J.; Richardson, A.L.; Weinberg, R.A. Stromal Fibroblasts Present in Invasive Human Breast Carcinomas Promote Tumor Growth and Angiogenesis Through Elevated SDF-1/CXCL12 Secretion. Cell 2005, 121, 335–348. [Google Scholar] [CrossRef]
- Sha, M.; Jeong, S.; Qiu, B.; Tong, Y.; Xia, L.; Xu, N.; Zhang, J.; Xia, Q. Isolation of Cancer-Associated Fibroblasts and Its Promotion to the Progression of Intrahepatic Cholangiocarcinoma. Cancer Med. 2018, 7, 4665–4677. [Google Scholar] [CrossRef]
- Okabe, H.; Beppu, T.; Hayashi, H.; Horino, K.; Masuda, T.; Komori, H.; Ishikawa, S.; Watanabe, M.; Takamori, H.; Iyama, K.-I.; et al. Hepatic Stellate Cells May Relate to Progression of Intrahepatic Cholangiocarcinoma. Ann. Surg. Oncol. 2009, 16, 2555–2564. [Google Scholar] [CrossRef]
- Alkan, A.; Bulut, E.; Gunhan, O.; Ozden, B. Oral Verrucous Carcinoma: A Study of 12 Cases. Eur. J. Dent. 2010, 4, 202–207. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takabatake, K.; Shimo, T.; Murakami, J.; Anqi, C.; Kawai, H.; Yoshida, S.; Oo, M.W.; Haruka, O.; Sukegawa, S.; Tsujigiwa, H.; et al. The role of sonic hedgehog signaling in the tumor microenvironment of oral squamous cell carcinoma. Int. J. Mol. Sci. 2019, 20, 5779. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hanyu, S.; Sakuma, K.; Tanaka, A. A Study on the Effect of Human Dental Pulp Stem Cell Conditioned Medium on Human Oral Squamous Cell Carcinoma Cell Lines. J. Hard Tissue Biol. 2019, 28, 281–288. [Google Scholar] [CrossRef] [Green Version]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Takabatake, K.; Kawai, H.; Omori, H.; Qiusheng, S.; Oo, M.W.; Sukegawa, S.; Nakano, K.; Tsujigiwa, H.; Nagatsuka, H. Impact of the Stroma on the Biological Characteristics of the Parenchyma in Oral Squamous Cell Carcinoma. Int. J. Mol. Sci. 2020, 21, 7714. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21207714
Takabatake K, Kawai H, Omori H, Qiusheng S, Oo MW, Sukegawa S, Nakano K, Tsujigiwa H, Nagatsuka H. Impact of the Stroma on the Biological Characteristics of the Parenchyma in Oral Squamous Cell Carcinoma. International Journal of Molecular Sciences. 2020; 21(20):7714. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21207714
Chicago/Turabian StyleTakabatake, Kiyofumi, Hotaka Kawai, Haruka Omori, Shan Qiusheng, May Wathone Oo, Shintaro Sukegawa, Keisuke Nakano, Hidetsugu Tsujigiwa, and Hitoshi Nagatsuka. 2020. "Impact of the Stroma on the Biological Characteristics of the Parenchyma in Oral Squamous Cell Carcinoma" International Journal of Molecular Sciences 21, no. 20: 7714. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21207714