Genome-Wide Association Studies and Transcriptome Changes during Acclimation and Deacclimation in Divergent Brassica napus Varieties

 , and
, and
Abstract
:1. Introduction
2. Results
2.1. Genotypic Differences in Response to Deacclimation Temperatures
2.2. RNAseq Analysis Identified Differential Expression between Treatments and between Varieties
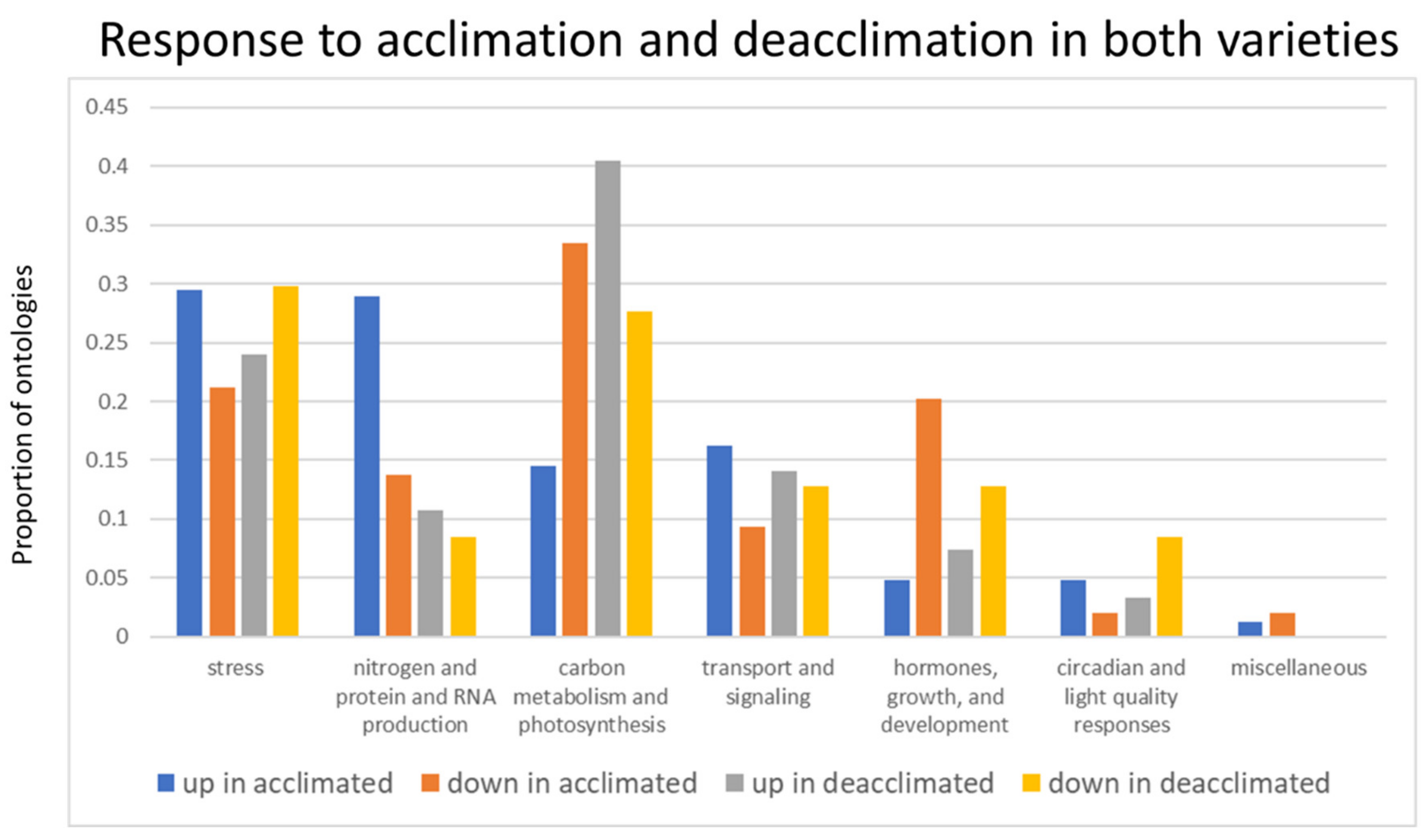
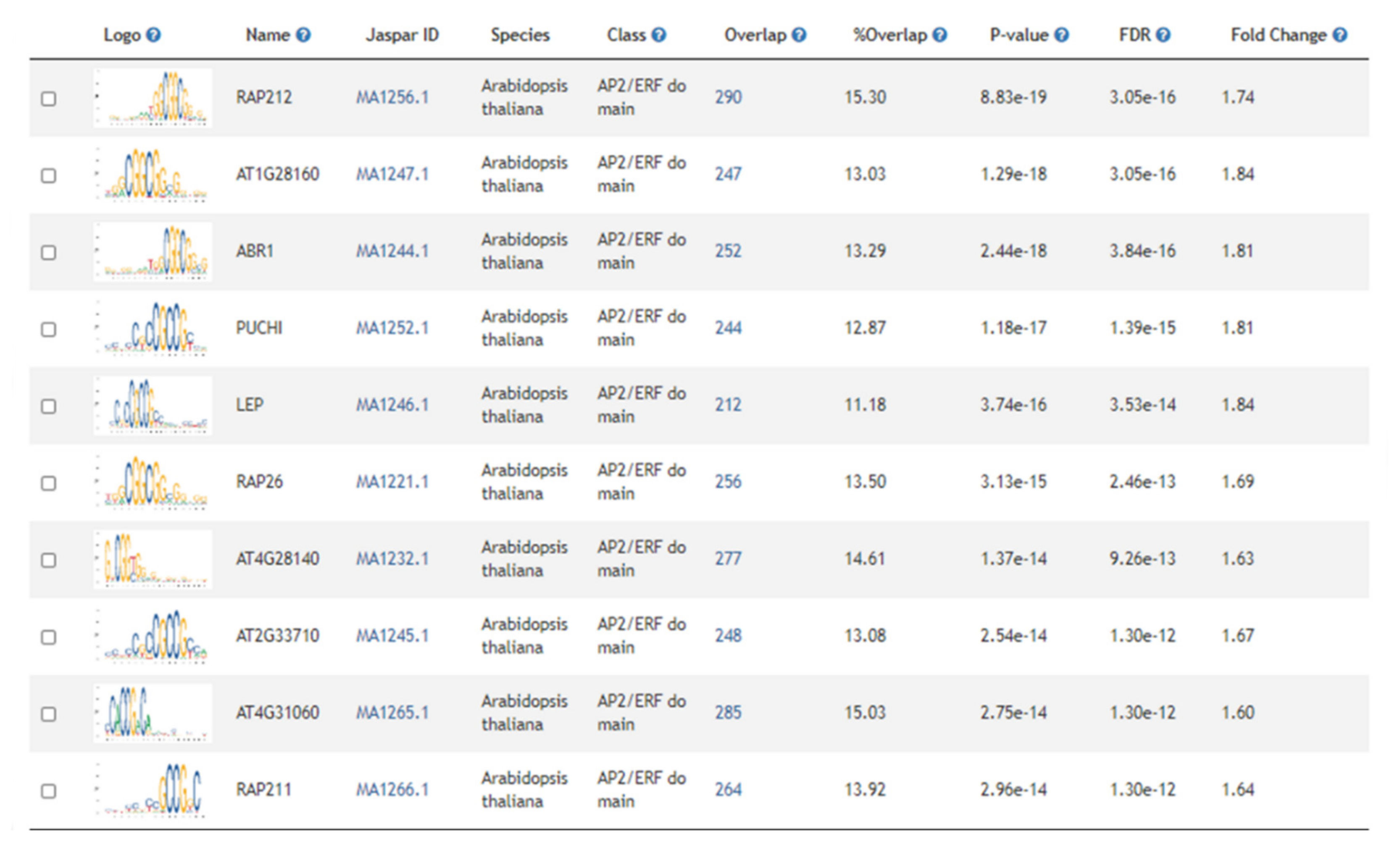
2.3. Gene Set Enrichment Analysis Identified Physiological Processes and Regulatory Components Associated with Freezing Tolerance
2.4. Differential Regulation of Circadian Responses, Photosynthetic Processes, and Hormones Associated with Abiotic Stress Are Associated with Deacclimation
2.5. Differences between Deacclimated and Non-Deacclimated Varieties Highlight Defense Responses in the Deacclimation-Sensitive Variety KS09068B-5-1
2.6. GWAS Identified Multiple Loci Associated with Freezing Tolerance after Deacclimation
3. Discussion
3.1. Genetic Analysis of Deacclimation Processes Identifies Numerous Candidate Genes
3.2. RNAseq Identifies Genes and Processes Previously Associated with Freezing Tolerance and Cold Acclimation
3.3. Similarities and Differences in Response to Deacclimation between Varieties Highlight Defense Responses and Photosynthetic Processes
4. Materials and Methods
4.1. Deacclimation Studies and Freezing Stress Assessment
4.2. RNAseq Analysis
4.3. Genome-Wide Association Study
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| GWAS | Genome-Wide Association Study |
| MLM | Mixed Linear Model |
| GLM | General Linear Model |
| FPKM | Fragment per Kilobase per Million |
| FDR | False Discovery Rate |
| CBF | CRT/DRE BINDING FACTOR |
| PCA | Principle Component Analysis |
References
- Boyles, M.; Peeper, T.; Stamm, M. Great Plains Canola Production Handbook; MF-2734; Kansas State University: Manhattan, KS, USA, 2006. [Google Scholar]
- Thomashow, M.F. Plant cold acclimation: Freezing tolerance genes and regulatory mechanisms. Annu. Rev. Plant Physiol. 1999, 50, 571–599. [Google Scholar] [CrossRef] [Green Version]
- Thomashow, M.F.; Torii, K.U. SCREAMing Twist on the Role of ICE1 in Freezing Tolerance. Plant Cell 2020, 32, 816–819. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fürtauer, L.; Weiszmann, J.; Weckwerth, W.; Nägele, T. Dynamics of Plant Metabolism during Cold Acclimation. Int. J. Mol. Sci. 2019, 20, 5411. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ding, Y.; Shi, Y.; Yang, S. Advances and challenges in uncovering cold tolerance regulatory mechanisms in plants. New Phytol. 2019, 222, 1690–1704. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vyse, K.; Pagter, M.; Zuther, E.; Hincha, D.K. Deacclimation after cold acclimation—A crucial, but widely neglected part of plant winter survival. J. Exp. Bot. 2019, 70, 4595–4604. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, L.; Coulter, J.A.; Liu, L.; Zhao, Y.; Chang, Y.; Pu, Y.; Zeng, X.; Xu, Y.; Wu, J.; Fang, Y.; et al. Transcriptome Analysis Reveals Key Cold-Stress-Responsive Genes in Winter Rapeseed (Brassica rapa L.). Int. J. Mol. Sci. 2019, 20, 1071. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zuther, E.; Juszczak, I.; Lee, Y.P.; Baier, M.; Hincha, D.K. Time-dependent deacclimation after cold acclimation in Arabidopsis thaliana accessions. Sci. Rep. 2015, 5, 12199. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Juszczak, I.; Ecvetkovic, J.; Ezuther, E.; Hincha, D.K.; Baier, M. Natural Variation of Cold Deacclimation Correlates with Variation of Cold-Acclimation of the Plastid Antioxidant System in Arabidopsis thaliana Accessions. Front. Plant Sci. 2016, 7, 305. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Trischuk, R.G.; Schilling, B.S.; Low, N.H.; Gray, G.R.; Gusta, L.V. Cold acclimation, de-acclimation and re-acclimation of spring canola, winter canola and winter wheat: The role of carbohydrates, cold-induced stress proteins and vernalization. Environ. Exp. Bot. 2014, 106, 156–163. [Google Scholar] [CrossRef]
- Rapacz, M. Cold-deacclimation of Oilseed Rape (Brassica napus var. oleifera) in Response to Fluctuating Temperatures and Photoperiod. Ann. Bot. 2002, 89, 543–549. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Visscher, P.M.; Wray, N.R.; Zhang, Q.; Sklar, P.; McCarthy, M.I.; Brown, M.A.; Yang, J. 10 Years of GWAS Discovery: Biology, Function, and Translation. Am. J. Hum. Genet. 2017, 101, 5–22. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wurke, D.F.; Talukder, Z.I.; Rahma, M. Genome-wide association study for frost tolerance in rapeseed/canola (Brassica napus) under simulating freezing conditions. Plant Breed. 2020, 139, 356–367. [Google Scholar]
- Oakley, C.G.; Ågren, J.; Atchison, R.A.; Schemske, D.W. QTL mapping of freezing tolerance: Links to fitness and adaptive trade-offs. Mol. Ecol. 2014, 23, 4304–4315. [Google Scholar] [CrossRef] [PubMed]
- Chalhoub, B.; Denoeud, F.; Liu, S.; Parkin, I.A.P.; Tang, H.; Wang, X.; Chiquet, J.; Belcram, H.; Tong, C.; Samans, B.; et al. Early allopolyploid evolution in the post-Neolithic Brassica napus oilseed genome. Science 2014, 345, 950–953. [Google Scholar] [CrossRef] [Green Version]
- Horvath, D.P.; Stamm, M.; Talukder, Z.; Fiedler, J.; Horvath, A.P.; Ashok, M.; Horvath, G.A.; Chao, W.S.; Anderson, J.V. A new diversity panel for winter rapeseed (Brassica napus L.) genome wide association studies. Agronomy 2020. submitted. [Google Scholar]
- Van Leene, J.; Han, C.; Gadeyne, A.; Eeckhout, D.; Matthijs, C.; Cannoot, B.; De Winne, N.; Persiau, G.; Van De Slijke, E.; Van De Cotte, B.; et al. Capturing the phosphorylation and protein interaction landscape of the plant TOR kinase. Nat. Plants 2019, 5, 316–327. [Google Scholar] [CrossRef]
- Chao, W.S.; Horvath, D.P.; Stam, M.; Anderson, J.V. Genome-Wide Association Mapping of Freezing Tolerance in B. napus (Brassica napus L.). Agronomy 2020. submitted. [Google Scholar]
- Horton, M.W.; Hancock, A.M.; Huang, Y.S.; Toomajian, C.; Atwell, S.; Auton, A.; Muliyati, N.W.; Platt, A.; Sperone, F.G.; Vilhjálmsson, B.J.; et al. Genome-wide patterns of genetic variation in worldwide Arabidopsis thaliana accessions from the RegMap panel. Nat. Genet. 2012, 44, 212–216. [Google Scholar] [CrossRef] [Green Version]
- Creelman, R.A.; Mullet, J.E. Jasmonic acid distribution and action in plants: Regulation during development and response to biotic and abiotic stress. Proc. Natl. Acad. Sci. USA 1995, 92, 4114–4119. [Google Scholar] [CrossRef] [Green Version]
- Margalha, L.; Confraria, A.; Baena, E. SnRK1 and TOR: Modulating growth–defense trade-offs in plant stress responses. J. Exp. Bot. 2019, 70, 2261–2274. [Google Scholar] [CrossRef]
- Kim, Y.S.; Jane, G.S.; Park, S.; Gilmour, S.J.; Wang, L.; Renna, L.; Brandizzi, F.; Grumet, R.; Thomashow, M.F. CAMTA-Mediated Regulation of Salicylic Acid Immunity Pathway Genes in Arabidopsis Exposed to Low Temperature and Pathogen Infection. Plant Cell 2017, 29, 2465–2477. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Feng, Y.; Yin, Y.; Fei, S.-Z. BdVRN1 Expression Confers Flowering Competency and Is Negatively Correlated with Freezing Tolerance in Brachypodium distachyon. Front. Plant Sci. 2017, 8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fowler, S.G.; Cook, D.; Thomashow, M.F. Low temperature induction of Arabidopsis CBF1, 2, and 3 is gated by the circadian clock. Plant Physiol. 2005, 137, 961–968. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Johnson-Flanagan, A.; Barran, L.; Singh, J. L-Methionine Transport during the Induction of Freezing Hardiness by Abscisic Acid in Brassica napus Cell Suspension Cultures. J. Plant Physiol. 1986, 124, 309–319. [Google Scholar] [CrossRef]
- Kurepin, L.V.; Keshav, P.; Dahal, K.P.; Savitch, L.V.; Singh, J.; Bode, R.; Ivanov, A.G.; Hurry, V.; Hüner, N.P.A. Role of CBFs as integrators of chloroplast redox, phytochrome and plant hormone signaling during cold acclimation. Int. J. Mol. Sci. 2013, 14, 12729–12763. [Google Scholar] [CrossRef] [PubMed]
- Xin, H.; Ni, X.C.; Pan, X.; Wei, L.; Min, Y.; Yu, K.; Qin, L.W.; Wei, H. Comparative transcriptome analyses revealed conserved and novel responses to cold and freezing stress in Brassica napus L. G3 Genes Genom. Genet. 2019, 9, 2723–2737. [Google Scholar] [CrossRef] [Green Version]
- Xian, M.; Luo, T.; Khan, M.N.; Hu, L.; Xu, Z. Identifying Differentially Expressed Genes Associated with Tolerance against Low Temperature Stress in Brassica napus through Transcriptome Analysis. Int. J. Agric. Biol. 2017, 19, 273–281. [Google Scholar] [CrossRef]
- Gilmour, S.J.; Zarka, D.G.; Stockinger, E.J.; Salazar, M.P.; Houghton, J.M.; Thomashow, M.F. Low temperature regulation of the Arabidopsis CBF family of AP2 transcriptional activators as an early step in cold-induced COR gene expression. Plant J. 1998, 16, 433–442. [Google Scholar] [CrossRef]
- Fowler, S.; Thomashow, M.F. Arabidopsis transcriptome profiling indicates that multiple regulatory pathways are activated during cold acclimation in addition to the CBF cold response pathway. Plant Cell 2002, 14, 1675–1690. [Google Scholar] [CrossRef] [Green Version]
- Markovskaya, E.F.; Kosobryukhov, A.A.; Kreslavski, V.D. Photosynthetic machinery response to low temperature stress. In Photosynthesis: New Approaches to the Molecular, Cellular, and Organismal Levels; Allakhverdiev, S.I., Ed.; Scrivener Publishing LLC: Beverly, MA, USA, 2016; pp. 355–382. [Google Scholar]
- Shi, Y.; Ding, Y.; Yang, S. Molecular Regulation of CBF Signaling in Cold Acclimation. Trends Plant Sci. 2018, 23, 623–637. [Google Scholar] [CrossRef]
- Byun, Y.J.; Koo, M.Y.; Joo, H.J.; Ha-Lee, Y.M.; Lee, D.H. Comparative analysis of gene expression under cold acclimation, deacclimation and reacclimation in Arabidopsis. Physiol. Plant. 2014, 152, 256–274. [Google Scholar] [CrossRef] [PubMed]
- Pagter, M.; Alpers, J.; Erban, A.; Kopka, J.; Zuther, E.; Hincha, D.K. Rapid transcriptional and metabolic regulation of the deacclimation process in cold acclimated Arabidopsis thaliana. BMC Genom. 2017, 18, 731. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oliver, S.L.; Lenards, A.J.; Barthelson, R.A.; Merchant, N.; McKay, S.J. Using the iPlant Collaborative Discovery Environment. Curr. Protoc. Bioinform. 2013, 42, 1.22.1–1.22.26. [Google Scholar] [CrossRef] [PubMed]
- Ran, X.; Zhao, F.; Wang, Y.; Liu, J.; Zhuang, Y.; Ye, L.; Qi, M.; Cheng, J.; Zhang, Y. Plant Regulomics: A data-driven interface for retrieving upstream regulators from plant multi-omics data. Plant J. 2019, 101, 237–248. [Google Scholar] [CrossRef] [PubMed]
- Mamidi, S.; Chikara, S.; Goos, R.J.; Hyten, D.L.; Annam, D.; Moghaddam, S.M.; Lee, R.K.; Cregan, P.B.; McClean, P.E. Genome-Wide Association Analysis Identifies Candidate Genes Associated with Iron Deficiency Chlorosis in Soybean. Plant Genome 2011, 4, 154–164. [Google Scholar] [CrossRef] [Green Version]
- Liu, X.; Huang, M.; Fan, B.; Buckler, E.S.; Zhang, Z. Iterative Usage of Fixed and Random Effect Models for Powerful and Efficient Genome-Wide Association Studies. PLoS Genet. 2016, 12, e1005767. [Google Scholar] [CrossRef]
- Huang, M.; Liu, X.; Zhou, Y.; Summers, R.M.; Zhang, Z. BLINK: A package for the next level of genome-wide association studies with both individuals and markers in the millions. GigaScience 2019, 8, 154. [Google Scholar] [CrossRef]
- Wang, Q.; Tian, F.; Pan, Y.; Buckler, E.S.; Zhang, Z. A SUPER powerful method for genome wide association study. PLoS ONE 2014, 9, e107684. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Marker | Position | LD Interval | B. napus Gene Model | Gene Model Map Position | Arabidopsis Orthologue | Probable Gene Function |
|---|---|---|---|---|---|---|
| S1_750314091 | Ann_Random 37666286 | 37666186-37666386 | DMBras.unigeneT00180796001 | 109261544 to 109263434 | NA | NA |
| S1_698489879 | C06_random 1114643 | 1094643-1134643 | BnaC06g41810D | 1113495 to 1114219 | NA | NA |
| BnaC06g41820D | 1126313 to 1126528 | NA | NA | |||
| BnaC06g41830D | 1133565 to 1134202 | AT1G52720 | unknown function | |||
| BnaC06g41840D | 1135605 to 1138056 | AT1G52730 | Transducin/WD40 repeat-like superfamily protein | |||
| S1_770664926 | Cnn_random 9358695 | 9328695-9388695 | BnaCnng10210D | 9333223 to 9335001 | AT5G04270 | DHHC-type zinc finger family protein |
| BnaCnng10220D | 9335248 to 9336428 | AT5G04260 | WCRKC thioredoxin 2 | |||
| BnaCnng10230D | 9336609 to 9338164 | AT5G04250 | Cysteine proteinases superfamily protein | |||
| BnaCnng10240D | 9339664 to 9344492 | AT5G04240 | transcription factor jumonji (jmj) family protein | |||
| BnaCnng10250D | 9353589 to 9356426 | AT5G04220 | deacclimation-regulated Calcium-dependent lipid-binding (CaLB domain) | |||
| BnaCnng10260D | 9356627 to 9356920 | AT5G04200 | metacaspase 9 | |||
| BnaCnng10270D | 9356994 to 9357596 | AT5G04200 | metacaspase 9 | |||
| BnaCnng10280D | 9358250 to 9359464 | AT5G04190 | phytochrome kinase substrate 4 | |||
| BnaCnng10290D | 9360809 to 9362430 | AT5G04180 | alpha carbonic anhydrase 3 | |||
| BnaCnng10300D | 9365237 to 9367298 | AT5G04170 | Calcium-binding EF-hand family protein | |||
| BnaCnng10310D | 9368412 to 9369733 | AT1G14800 | Nucleic acid-binding, OB-fold-like protein | |||
| BnaCnng10320D | 9372342 to 9374120 | AT5G04160 | Nucleotide-sugar transporter family protein | |||
| BnaCnng10330D | 9374957 to 9376012 | AT4G36840 | Galactose oxidase/kelch repeat superfamily protein | |||
| BnaCnng10340D | 9376962 to 9384925 | AT5G04140 | glutamate synthase 1 | |||
| BnaCnng10350D | 9386245 to 9387697 | AT5G04120 | Phosphoglycerate mutase family protein | |||
| S1_38158858 | C03 57859505 | 57759505-5799505 | BnaC03g68090D | 57742606 to 57744811 | AT2G19090 | Protein of unknown function (DUF630 and DUF632) |
| BnaC03g68100D | 57751536 to 57752049 | AT4G30074 | low-molecular-weight cysteine-rich 19 | |||
| BnaC03g68110D | 57761207 to 57764215 | AT4G30060 | Core-2/I-branching beta-1,6-N-acetylglucosaminyltransferase protein | |||
| BnaC03g68120D | 57765406 to 57766578 | AT4G30010 | LOCATED IN: mitochondrion, plastid | |||
| BnaC03g68130D | 57768296 to 57772432 | AT4G29960 | LOCATED IN: plasma membrane | |||
| BnaC03g68140D | 57772494 to 57774352 | AT4G29950 | Ypt/Rab-GAP domain of gyp1p superfamily protein | |||
| BnaC03g68150D | 57774688 to 57778115 | AT4G29940 | pathogenesis related homeodomain protein A | |||
| BnaC03g68160D | 57788448 to 57789017 | AT4G29930 | basic helix-loop-helix (bHLH) DNA-binding superfamily protein | |||
| BnaC03g68170D | 57789892 to 57792284 | AT4G29930 | basic helix-loop-helix (bHLH) DNA-binding superfamily protein | |||
| BnaC03g68180D | 57793363 to 57793613 | NA | NA | |||
| BnaC03g68190D | 57793691 to 57793750 | NA | NA | |||
| BnaC03g68200D | 57816672 to 57819918 | AT4G29920 | Double Clp-N motif-containing P-loop nucleoside triphosphate hydrolases | |||
| BnaC03g68210D | 57826932 to 57827135 | AT4G29905 | Unknown conserved | |||
| BnaC03g68220D | 57833058 to 57834309 | AT5G63820 | Protein of unknown function (DUF626) | |||
| BnaC03g68230D | 57839948 to 57841025 | AT5G63820 | Protein of unknown function (DUF626) | |||
| BnaC03g68240D | 57841738 to 57843321 | AT5G63820 | Protein of unknown function (DUF626) | |||
| BnaC03g68250D | 57858635 to 57860206 | AT4G29840 | Pyridoxal-5′-phosphate-dependent enzyme family protein | |||
| BnaC03g68260D | 57860353 to 57861992 | AT4G29830 | Transducin/WD40 repeat-like superfamily protein VIP3 | |||
| BnaC03g68270D | 57956109 to 57956560 | AT4G29700 | Alkaline-phosphatase-like family protein | |||
| BnaC03g68280D | 57961823 to 57962718 | AT4G29690 | Alkaline-phosphatase-like family protein | |||
| S1_122523793 | A06 2474952 | 2473952-2475952 | BnaA06g04060D | 2451590 to 2477036 | AT1G48090 | calcium-dependent lipid-binding family protein |
| S1_757283058 | Ann_random 44635253 | 44635153-44635353 | BnaAnng39250D | 44634379 to 44636475 | AT3G11510 | Ribosomal protein S11 family protein |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Horvath, D.P.; Zhang, J.; Chao, W.S.; Mandal, A.; Rahman, M.; Anderson, J.V. Genome-Wide Association Studies and Transcriptome Changes during Acclimation and Deacclimation in Divergent Brassica napus Varieties. Int. J. Mol. Sci. 2020, 21, 9148. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21239148
Horvath DP, Zhang J, Chao WS, Mandal A, Rahman M, Anderson JV. Genome-Wide Association Studies and Transcriptome Changes during Acclimation and Deacclimation in Divergent Brassica napus Varieties. International Journal of Molecular Sciences. 2020; 21(23):9148. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21239148
Chicago/Turabian StyleHorvath, David P., Jiaping Zhang, Wun S. Chao, Ashok Mandal, Mukhlesur Rahman, and James V. Anderson. 2020. "Genome-Wide Association Studies and Transcriptome Changes during Acclimation and Deacclimation in Divergent Brassica napus Varieties" International Journal of Molecular Sciences 21, no. 23: 9148. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21239148