Description of a Novel Mechanism Possibly Explaining the Antiproliferative Properties of Glucocorticoids in Duchenne Muscular Dystrophy Fibroblasts Based on Glucocorticoid Receptor GR and NFAT5




Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Results
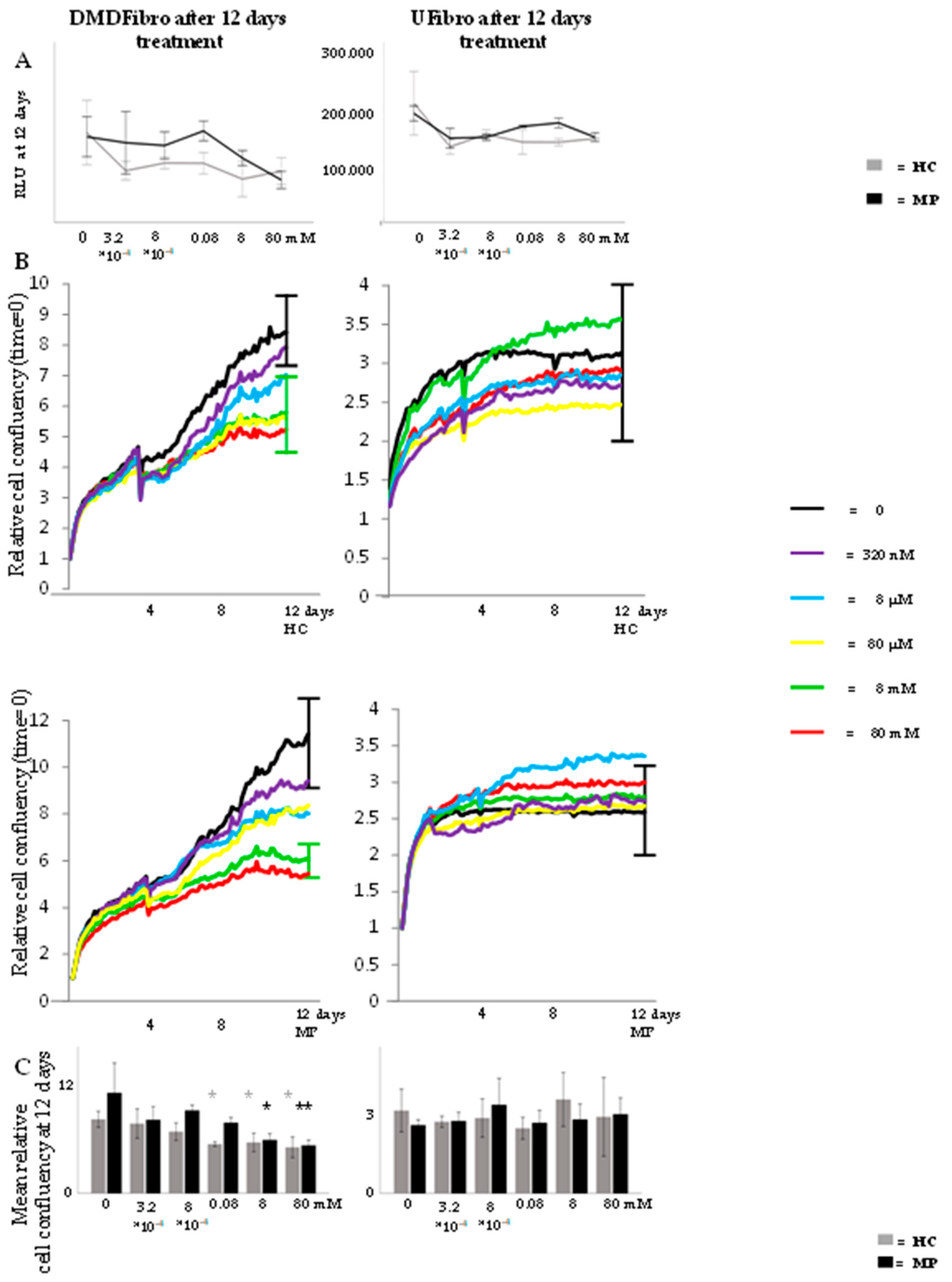
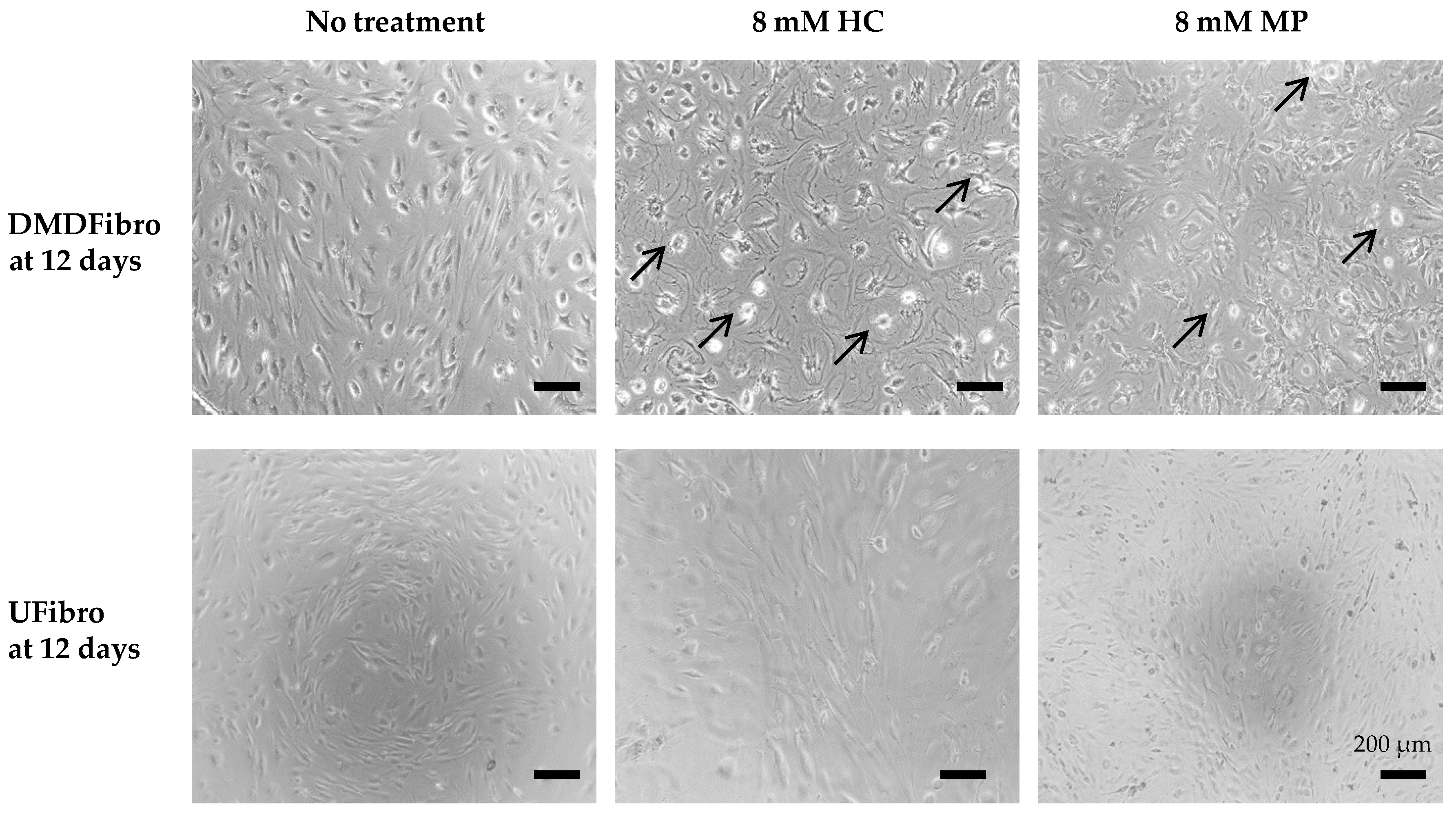
2.1. Hydrocortisone and Methylprednisolone only Decrease DMD Fibroblasts Growth
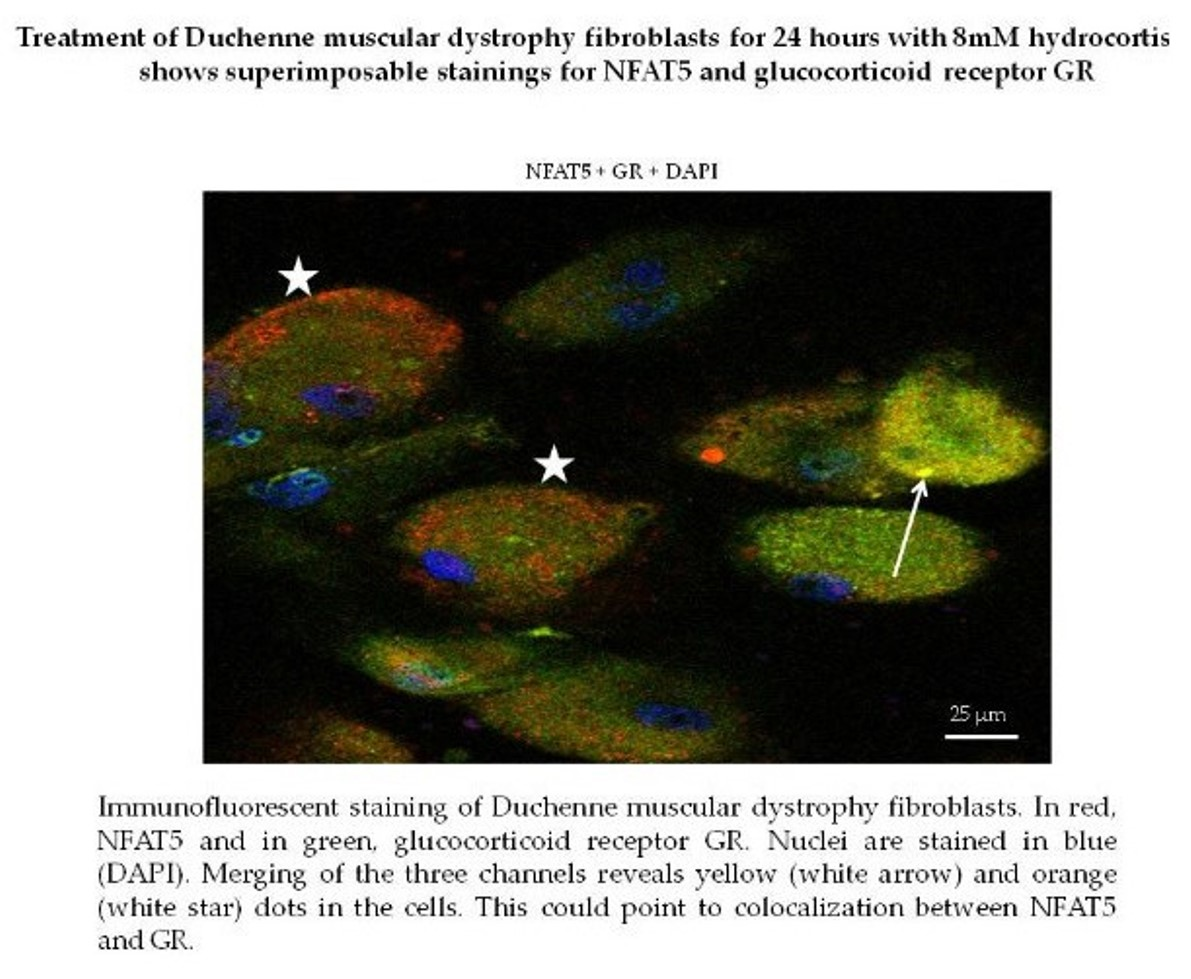
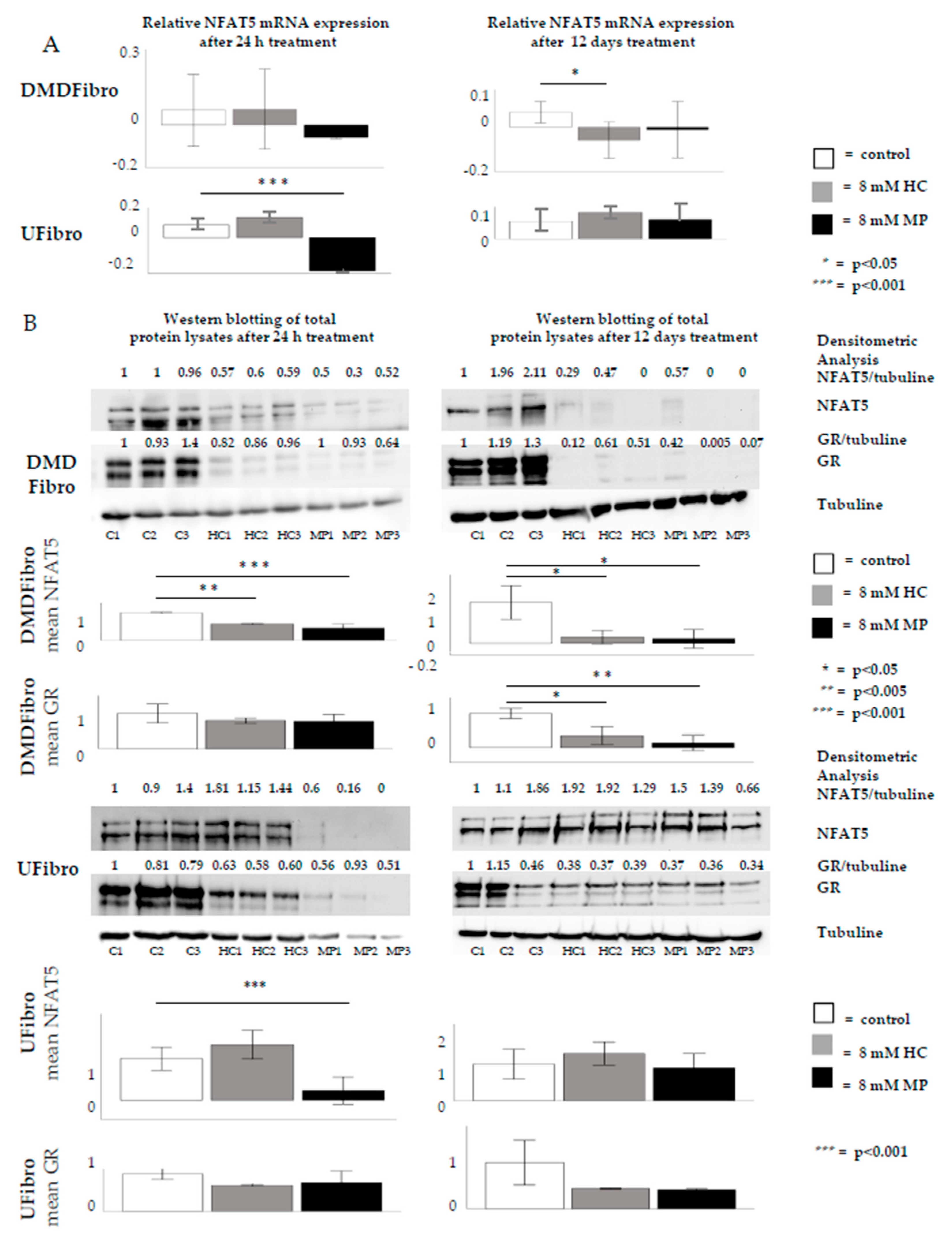
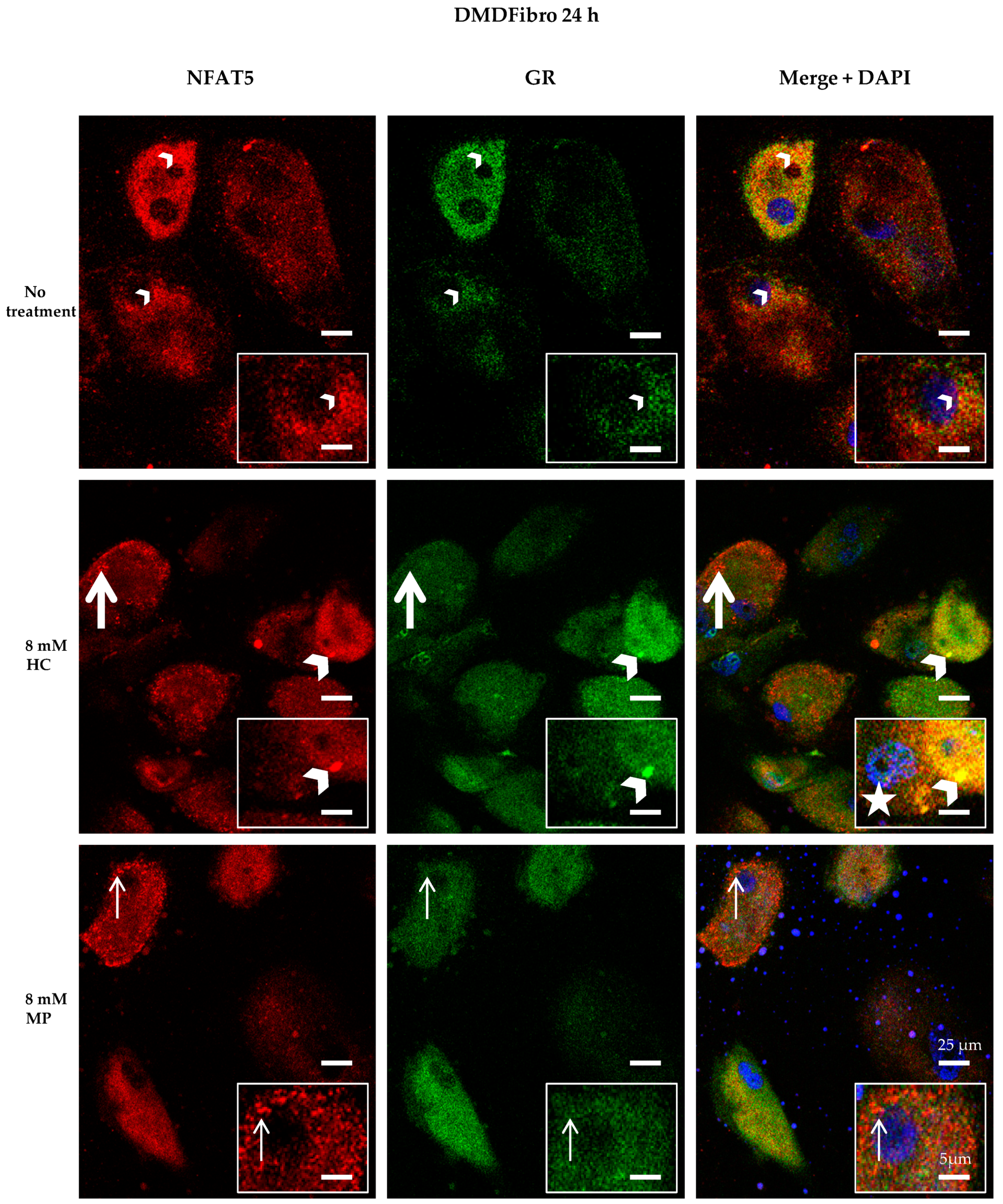
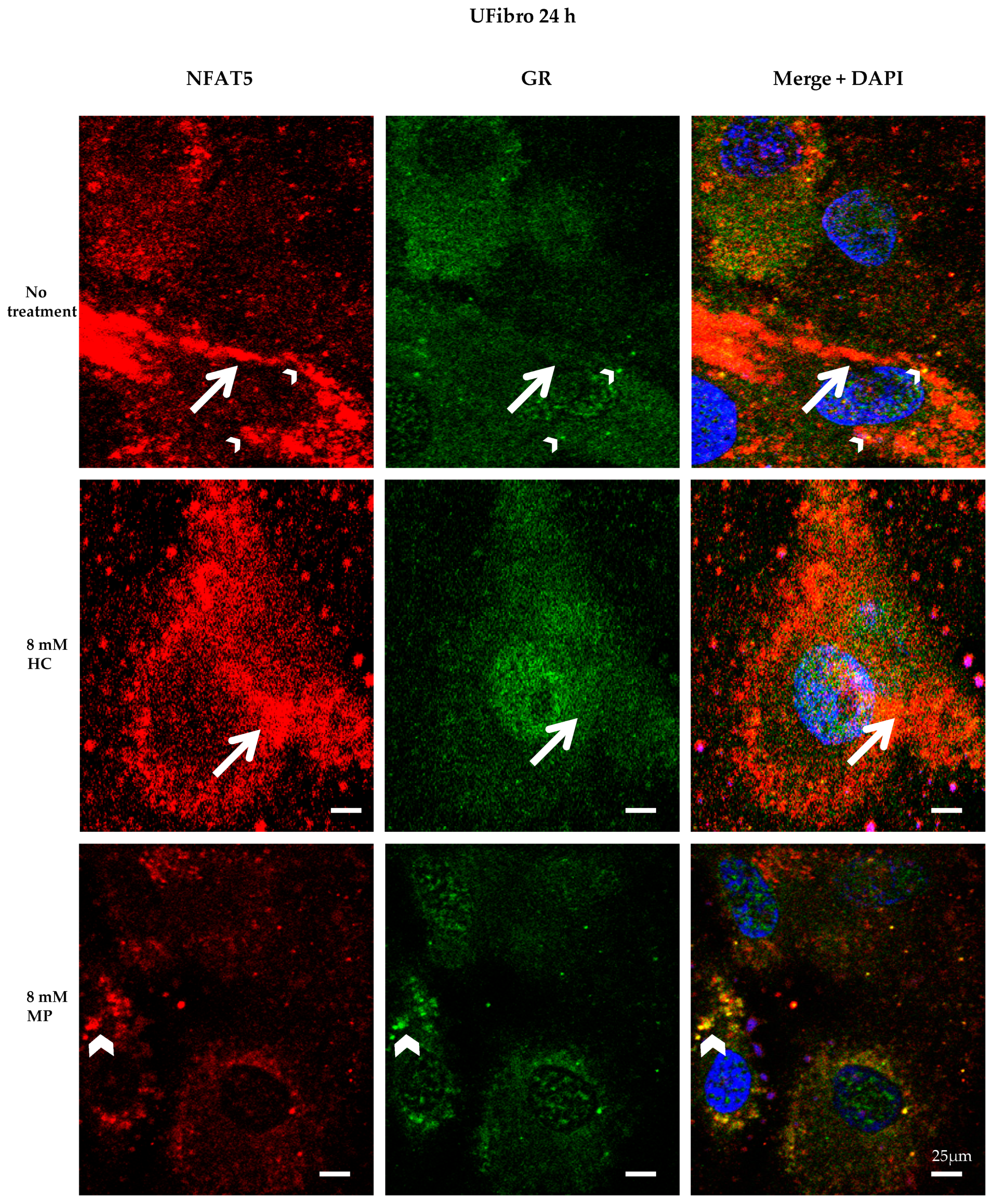
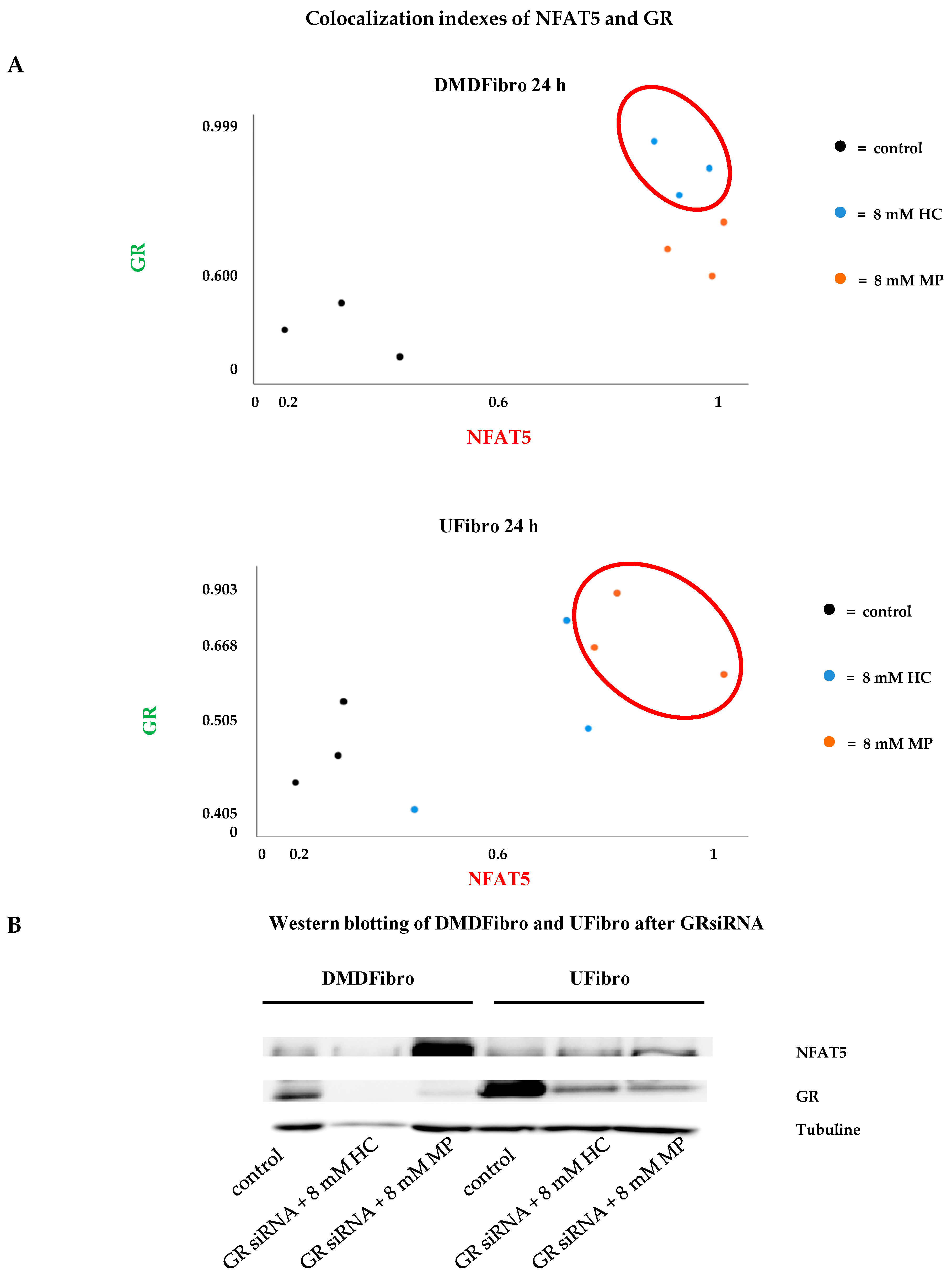
2.2. Insight into the Effect of Hydrocortisone and Methylprednisolone on DMD Fibroblast Growth through NFAT5-GR Interaction
3. Discussion
4. Materials and Methods
4.1. Cell Lines
4.2. Cell Viability and Life Imaging by IncuCyte ZOOM
4.3. Treatment with Methylprednisolone or Hydrocortisone
4.4. RT-qPCR
4.5. Quantitative Western Blotting (WB)
4.6. GR siRNA
4.7. Immunocytochemistry (ICC) and Confocal Microscopy (CM)
4.8. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| CBG | corticoid binding globulin |
| DAG | donkey anti goat |
| DAM | donkey anti mouse |
| DMD | Duchenne muscular dystrophy |
| DMDFibro | Duchenne muscular dystrophy fibroblasts |
| DMEM | Dulbecco’s modified Eagle medium |
| ECM | extracellular matrix |
| FITC | fluorescein isothiocyanate |
| GCs | glucocorticoids |
| GR | glucocorticoid receptor |
| HC | hydrocortisone |
| HCSS | hydrocortisone sodium succinate |
| HPA | hypothalamic-pituitary-adrenal |
| ICC | immunocytochemistry |
| IM | intramuscular |
| IV | intravenous |
| MIQE | minimum information for publication of quantitative real-time PCR experiments |
| MP | methylprednisolone |
| MPSS | methylprednisolone sodium succinate |
| N | number of passages |
| NFAT5 | nuclear factor of activated T-cells 5 |
| NF-κB | nuclear factor kappa-light-chain-enhancer of activated B cells |
| RLU | relative light units |
| RT-qPCR | real-time quantitative PCR |
| scRNA | scrambled RNA |
| siRNA | silencing RNA |
| UFibro | unaffected skeletal muscle fibroblasts |
| UMyo1 | unaffected myoblasts |
| WB | Western blotting |
References
- Ray, A.; Prefontaine, K.E. Physical association and functional antagonism between the p65 subunit of transcription factor NF-kappa B and the glucocorticoid receptor. Proc. Natl. Acad. Sci. USA 1994, 91, 752–756. [Google Scholar] [CrossRef] [Green Version]
- Goya, L.; Maiyar, A.C.; Ge, Y.; Firestone, G.L. Glucocorticoids induce a G1/G0 cell cycle arrest of Con8 rat mammary tumor cells that is synchronously reversed by steroid withdrawal or addition of transforming growth factor-alpha. Mol. Endocrinol. 1993, 7, 1121–1132. [Google Scholar] [PubMed] [Green Version]
- Frost, G.H.; Rhee, K.; Ma, T.; Thompson, E.A. Expression of c-Myc in glucocorticoid-treated fibroblastic cells. J. Steroid Biochem. Mol. Biol. 1994, 50, 109–119. [Google Scholar] [CrossRef]
- Oakley, R.H.; Cidlowski, J.A. The biology of the glucorticoid receptor: New signaling mechanisms in health and disease. J. Allergy Clin. Immunol. 2013, 132, 1033–1044. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Herbelet, S.; Herregods, L.; Coppens, M. Exploring strategies to reduce time span to bystander CPR in sudden cardiac arrest based on the mechanism of the witness acute stress response. Resuscitation 2018, 131, e7–e8. [Google Scholar] [CrossRef]
- Honoré, P.M.; Jacobs, R.; De Waele, E.; De Regt, J.; Rose, T.; Van Gorp, V.; Joannes-Boyau, O.; Boer, W.; Spapen, H.D. What do we know about steroids metabolism and ‘PK/PD approach’ in AKI and CKD especially while on RRT-Current status in 2014. Blood Purif. 2014, 38, 154–157. [Google Scholar] [CrossRef]
- Al-Habet, S.M.H.; Rogers, H.J. Methylprednisolone and pharmacokinetics after intravenous and oral administration. Br. J. Clin. Pharmac. 1989, 27, 285–290. [Google Scholar] [CrossRef] [Green Version]
- Daley-Yates, P.T.; Gregory, A.J.; Brooks, C.D. Pharmacokinetic and pharmacodynamic assessment of bioavailability for two prodrugs of methylprednisolone. Br. J. Clin. Pharm. 1997, 43, 593–601. [Google Scholar] [CrossRef] [Green Version]
- Behar-Cohen, F.F.; Gauthier, S.; El Aouni, A.; Chapon, P.; Parel, J.M.; Renard, G.; Chauvaud, D. Methylprednisolone concentrations in the vitreous and the serum after pulse therapy. Retina 2001, 21, 48–53. [Google Scholar] [CrossRef]
- Sklar, R.M.; Brown, R.H. Methylprednisolone increases dystrophin levels by inhibiting myotube death during myogenesis of normal human muscle in vitro. J. Neurol. Sci. 1991, 101, 73–81. [Google Scholar] [CrossRef]
- Sugiyama, K.; Kawada, T.; Sato, H.; Hirano, T. Comparison of suppressive potency between prednisolone and prednisolone sodium succinate against mitogen-induced blastogenesis of human peripheral blood mononuclear cells in-vitro. J. Pharm. Pharmacol. 2001, 53, 727–733. [Google Scholar] [CrossRef]
- Jung, C.; Greco, S.; Nguyen, H.H.T.; Ho, J.T.; Lewis, J.G.; Torpy, D.J.; Inder, W.J. Plasma, salivary and urinary cortisol levels following physiological stress doses of hydrocortisone in normal volunteers. BMC Endocr. Disord. 2014, 14, 91. Available online: http://0-www-biomedcentral-com.brum.beds.ac.uk/1472-6823/14/91 (accessed on 6 May 2020). [CrossRef] [Green Version]
- Herbelet, S.; Rodenbach, A.; De Paepe, B.; De Bleecker, J.L. Anti-inflammatory and general glucocorticoid physiology in skeletal muscles affected by Duchenne muscular dystrophy: Exploration of steroid-sparing agents. Int. J. Mol. Sci. 2020, 21, 4596. [Google Scholar] [CrossRef]
- Koenig, M.; Hoffman, E.P.; Bertelson, C.J.; Monaco, A.P.; Feener, C.; Kunkel, L.M. Complete cloning of the Duchenne muscular-dystrophy (DMD) cDNA and preliminary genomic organization of the DMD gene in normal and affected individuals. Cell 1987, 50, 509–517. [Google Scholar] [CrossRef]
- Hoffman, E.P.; Brown, R.H.; Kunke, L.M. Dystrophin—The protein product of the Duchenne muscular-dystrophy locus. Cell 1987, 51, 919–928. [Google Scholar] [CrossRef]
- Abdel-Salam, E.; Abdel-Mequi, I.; Korraa, S.S. Markers of degeneration and regeneration in Duchenne muscular dystrophy. Acta Myol. 2009, 28, 94–100. [Google Scholar]
- Banker, B.Q.; Engel, A.G. Basic reactions of muscle. In Myology; Engel, A.G., Franzini-Armstrong, C., Eds.; McGraw-Hill: New York, NY, USA, 2004; pp. 691–748. [Google Scholar]
- Mann, C.J.; Perdiguero, E.; Kharraz, Y.; Aguilar, S.; Pessina, P.; Serrano, A.L.; Muñoz-Cánoves, P. Aberrant repair and fibrosis development in skeletal muscle. Muscle Nerv. 2001, 1, 21. [Google Scholar] [CrossRef] [Green Version]
- Wynn, T.A.; Barron, L. Macrophages: Master regulators of inflammation and fibrosis. Semin. Liver Dis. 2010, 30, 245–257. [Google Scholar] [CrossRef] [Green Version]
- Yoon, H.-J.; You, S.; Yoo, S.-A.; Kim, N.-H.; Kwon, H.M.; Yoon, C.-H.; Cho, C.-S.; Hwang, D.; Kim, W.-U. NFAT5 is a critical regulator of inflammatory arthritis. Arthritis Rheum. 2011, 63. [Google Scholar] [CrossRef]
- Lee, S.; Kong, J.-S.; You, S.; Kwon, H.M.; Yoo, S.-A.; Cho, C.-S.; Kim, W.-U. Transcription factor NFAT5 promotes migration and invasion of rheumatoid synoviocytes via coagulation factor III and CCL2. J. Immunol. 2018, 201. [Google Scholar] [CrossRef] [Green Version]
- Herbelet, S.; De Paepe, B.; De Bleecker, J.L. Abnormal NFAT5 physiology in Duchenne muscular dystrophy fibroblasts as a putative explanation for the permanent fibrosis formation in Duchenne muscular dystrophy. Int. J. Mol. Sci. 2020, 21, 7888. [Google Scholar] [CrossRef]
- López-Rodríguez, C.; Aramburu, J.; Jin, L.; Rakeman, A.S.; Michino, M.; Rao, A. Bridging the NFAT and NF-κB families: NFAT5 dimerization regulates cytokine gene transcription in response to osmotic stress. Immunity 2001, 15, 47–58. [Google Scholar] [CrossRef] [Green Version]
- Auphan, N.; DiDonato, J.A.; Rosette, C.; Helmberg, A.; Karin, M. Immunosuppression by glucocorticoids: Inhibition of NF-κB activity through induction of IκB synthesis. Science 1995, 270, 286–290. [Google Scholar] [CrossRef]
- Herbelet, S.; De Vlieghere, E.; Gonçalves, A.; De Paepe, B.; Schmidt, K.; Nys, E.; Weynants, L.; Weis, J.; van Peer, G.; Vandesompele, J.; et al. Localization and expression of nuclear factor of activated T-cells 5 in myoblasts exposed to pro-inflammatory cytokines or hyperosmolar stress and in biopsies from myositis patients. Front. Physiol. 2018, 9, 126. [Google Scholar] [CrossRef] [Green Version]
- Roth, I.; Leroy, V.; Kwon, H.M.; Martin, P.-Y.; Féraille, E.; Hasler, U. Osmoprotective transcription factor NFAT5/TonEBP modulates nuclear factor κB activity. Mol. Biol. Cell. 2010, 21, 3459–3474. [Google Scholar] [CrossRef] [Green Version]
- Yamamoto, Y.; Gaynor, R.B. Therapeutic potential of inhibition of the NF-κB pathway in the treatment of inflammation and cancer. JCI 2001, 107, 135–142. [Google Scholar] [CrossRef] [Green Version]
- Chen, L.-F.; Greene, W.C. Shaping the nuclear action of NF-κB. Nat. Rev. Mol. Cell Biol. 2004, 5, 392–401. [Google Scholar] [CrossRef] [PubMed]
- Woo, S.K.; Dahl, S.C.; Handler, J.S.; Kwon, H.M. Bidirectional regulation of tonicity-responsive enhancer binding protein in response to changes in tonicity. Am. J. Physiol. Ren. Physiol. 2000, 278, F1006–F1012. [Google Scholar] [CrossRef]
- Bonifati, D.M.; Witchel, S.F.; Ermani, M.; Hoffman, E.P.; Angelini, C.; Pegoraro, E. The glucocorticoid receptor N363S polymorphism and steroid response in Duchenne dystrophy. J. Neurol. Neurosurg. Psychiatry 2006, 77, 1177–1179. [Google Scholar] [CrossRef] [Green Version]
- Tardif, G.; Pelletier, J.-P.; Fahmi, H.; Hum, D.; Zhang, Y.; Kapoor, M.; Martel-Pelletier, J. NFAT3 and TGF-β/SMAD3 regulate the expression of miR-140 in osteoarthritis. Arthritis Res. Ther. 2013, 15, R197. [Google Scholar] [CrossRef] [Green Version]
- Meng, X.-M.; Nikolic-Paterson, D.J.; Lan, H. YTGF-β: The master regulator of fibrosis. Nat. Rev. Nephrol. 2016, 12, 325–338. [Google Scholar] [CrossRef]
- Hussein, M.R.; Hamed, S.A.; Mostafa, M.G.; Abu-Dief, E.E.; Kamel, N.F.; Kandil, M.R. The effects of glucocorticoid therapy on the inflammatory and dendritic cells in muscular dystrophies. Int. J. Exp. Pathol. 2006, 87, 451–461. [Google Scholar] [CrossRef]
- Angelini, C. The role of corticosteroids in muscular dystrophy: A critical appraisal. Muscle Nerve 2007, 36, 424–435. [Google Scholar] [CrossRef]
- Vidal, B.; Serrano, A.L.; Tjwa, M.; Suelves, M.; Ardite, E.; De Mori, R.; Baeza-Raja, B.; Martínez de Lagrán, M.; Lafuste, P.; Ruiz-Bonilla, V.; et al. Fibrinogen drives dystrophic muscle fibrosis via a TGFβ/ alternative macrophage activation pathway. Genes Dev. 2008, 22, 1747–1752. [Google Scholar] [CrossRef] [Green Version]
- Meikle, A.W.; Tyler, F.H. Potency and duration of action of glucocorticoids: Effects of hydrocortisone, prednisone and dexamethasone on human pituitary-adrenal function. Am. J. Med. 1977, 63, 200–207. [Google Scholar] [CrossRef]
- Kerrigan, J.R.; Veldhuis, J.D.; Leyo, S.A.; Iranmanesh, A.; Rogol, A.D. Estimation of daily cortisol production and clearance rates in normal pubertal males by deconvolution analysis. J. Clin. Endocrinol. Metab. 1993, 76, 1505–1510. [Google Scholar]
- Wood, A.J.; Lamberts, S.W.; Bruining, H.A.; de Jong, F.H. Corticosteroid therapy in severe illness. NEJM 1997, 337, 1285–1292. [Google Scholar]
- Yong, S.L.; Coulthard, P.; Wrzosek, A. Supplemental perioperative steroids for surgical patients with adrenal insufficiency. Cochrane Database Syst. Rev. 2012, 12. [Google Scholar] [CrossRef] [Green Version]
- Bowden, S.A.; Connolly, A.M.; Kinnett, K.; Zeitler, P.S. Management of adrenal insufficiency risk after long-term systemic glucocorticoid therapy in Duchenne muscular dystrophy: Clinical practice recommendation. J. Neuromuscl. Dis. 2019, 6, 31–41. [Google Scholar] [CrossRef]
- Masur, S.K.; Dewal, H.S.; Dinh, T.T.; Erenburg, I.; Petridou, S. Myofibroblasts differentiate from fibroblasts when plated at low density. Proc. Natl. Acad. Sci. USA 1996, 93, 4219–4223. [Google Scholar] [CrossRef] [Green Version]
- Vandesompele, J.; De Preter, K.; Pattyn, F.; Poppe, B.; Van Roy, N.; De Paepe, A.; Speleman, F. Accurate normalization of real-time quantitative RT-PCR data by geometric averaging of multiple internal control genes. Genome Biol. 2002, 3, RESEARCH0034. [Google Scholar] [CrossRef] [Green Version]
- Bustin, S.A.; Benes, V.; Garson, J.A.; Hellemans, J.; Huggett, J.; Kubista, M.; Mueller, R.; Nolan, T.; Pfaffl, M.W.; Shipley, G.L.; et al. The MIQE guidelines: Minimum information for publication of quantitative real-time PCR experiments. Clin. Chem. 2009, 55, 4. [Google Scholar] [CrossRef] [Green Version]
- Hellemans, J.; Mortier, G.; De Paepe, A.; Speleman, F.; Vandesompele, J. qBase relative quantification framework and software for management and automated analysis of real-time quantitative PCR data. Genome Biol. 2007, 8, R19. [Google Scholar] [CrossRef] [Green Version]
- Bolte, S.; Cordelieres, F.P. A guided tour into subcellular colocalization analysis in light microscopy. J. Microsc. 2006, 224, 213–232. [Google Scholar] [CrossRef]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Herbelet, S.; De Paepe, B.; De Bleecker, J.L. Description of a Novel Mechanism Possibly Explaining the Antiproliferative Properties of Glucocorticoids in Duchenne Muscular Dystrophy Fibroblasts Based on Glucocorticoid Receptor GR and NFAT5. Int. J. Mol. Sci. 2020, 21, 9225. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21239225
Herbelet S, De Paepe B, De Bleecker JL. Description of a Novel Mechanism Possibly Explaining the Antiproliferative Properties of Glucocorticoids in Duchenne Muscular Dystrophy Fibroblasts Based on Glucocorticoid Receptor GR and NFAT5. International Journal of Molecular Sciences. 2020; 21(23):9225. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21239225
Chicago/Turabian StyleHerbelet, Sandrine, Boel De Paepe, and Jan L. De Bleecker. 2020. "Description of a Novel Mechanism Possibly Explaining the Antiproliferative Properties of Glucocorticoids in Duchenne Muscular Dystrophy Fibroblasts Based on Glucocorticoid Receptor GR and NFAT5" International Journal of Molecular Sciences 21, no. 23: 9225. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21239225