In Vitro Evaluation of Gentamicin or Vancomycin Containing Bone Graft Substitute in the Prevention of Orthopedic Implant-Related Infections


Abstract
:1. Introduction
2. Results
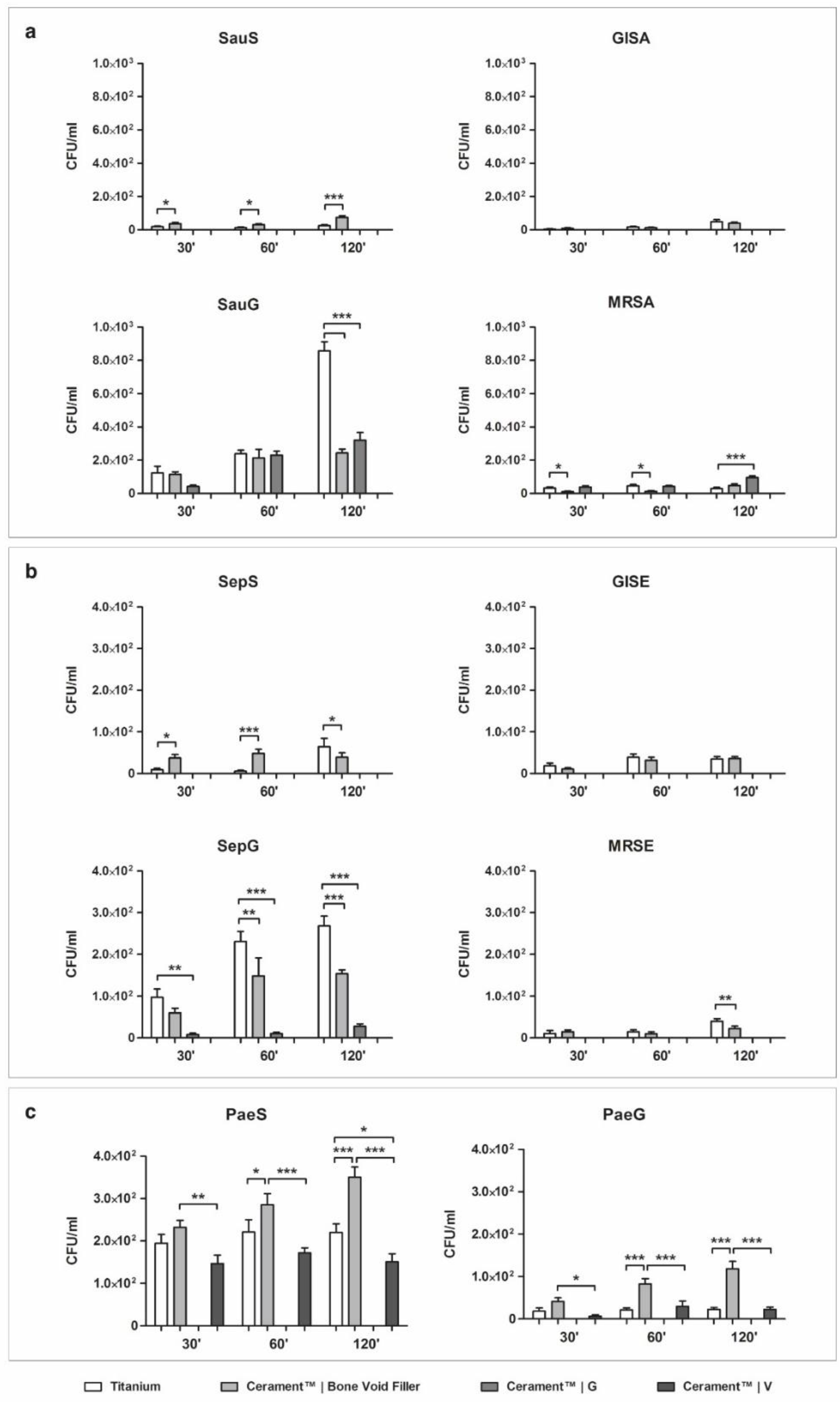
2.1. Bacterial Adhesion on Material Surface
2.1.1. Staphylococcus aureus
2.1.2. Staphylococcus epidermidis
2.1.3. Pseudomonas aeruginosa
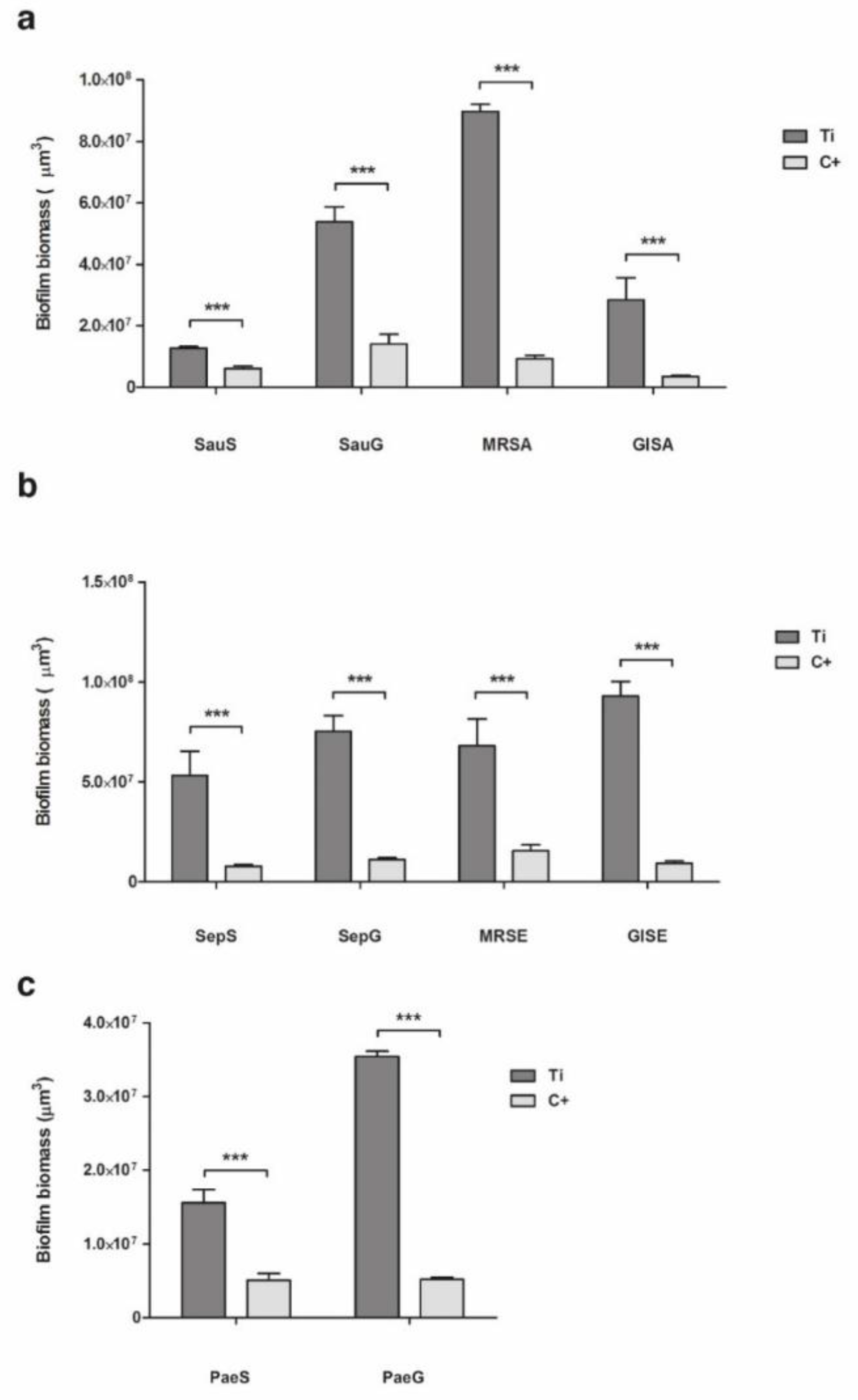
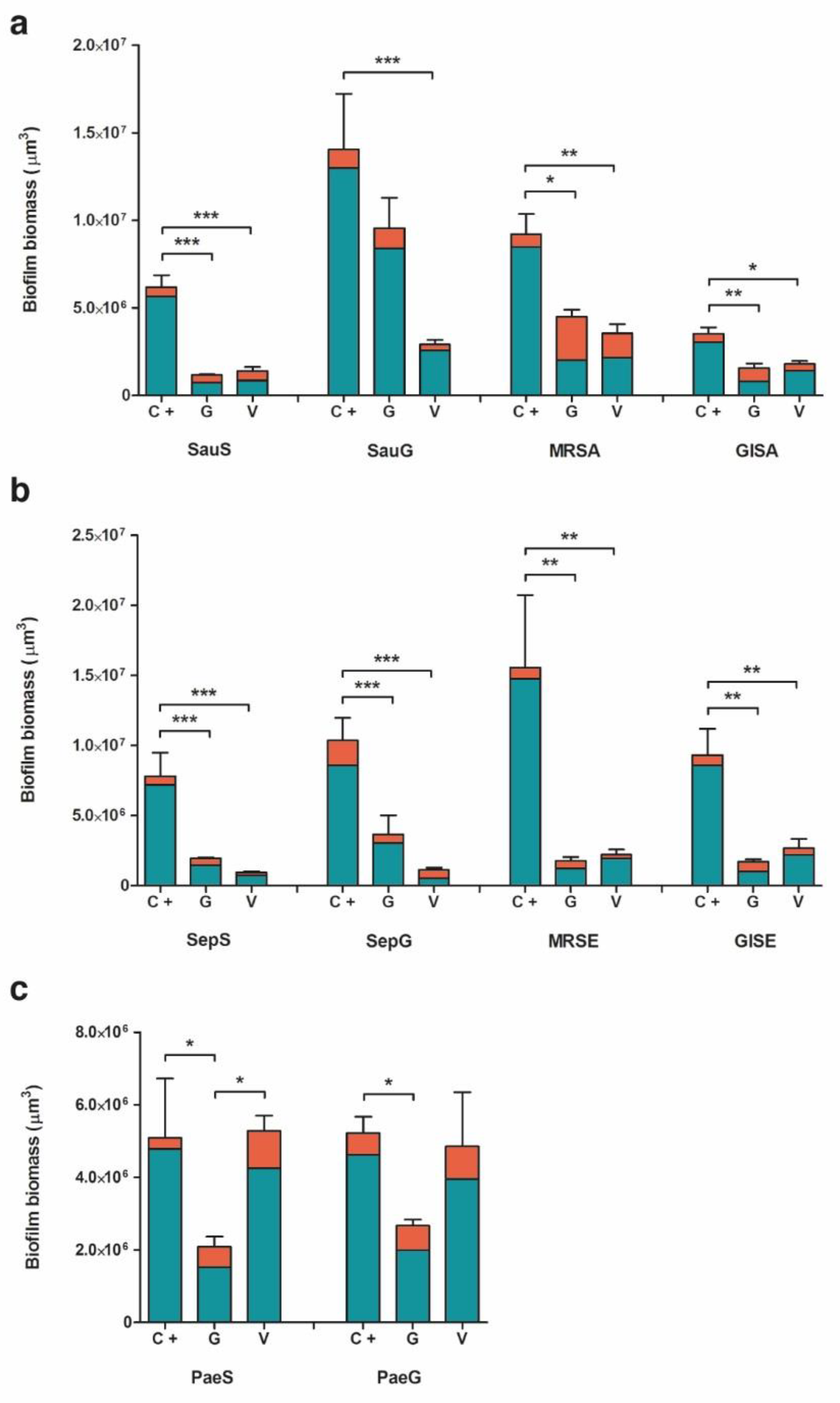
2.2. Biofilm Formation
2.2.1. Staphylococcus aureus
2.2.2. Staphylococcus epidermidis
2.2.3. Pseudomonas aeruginosa
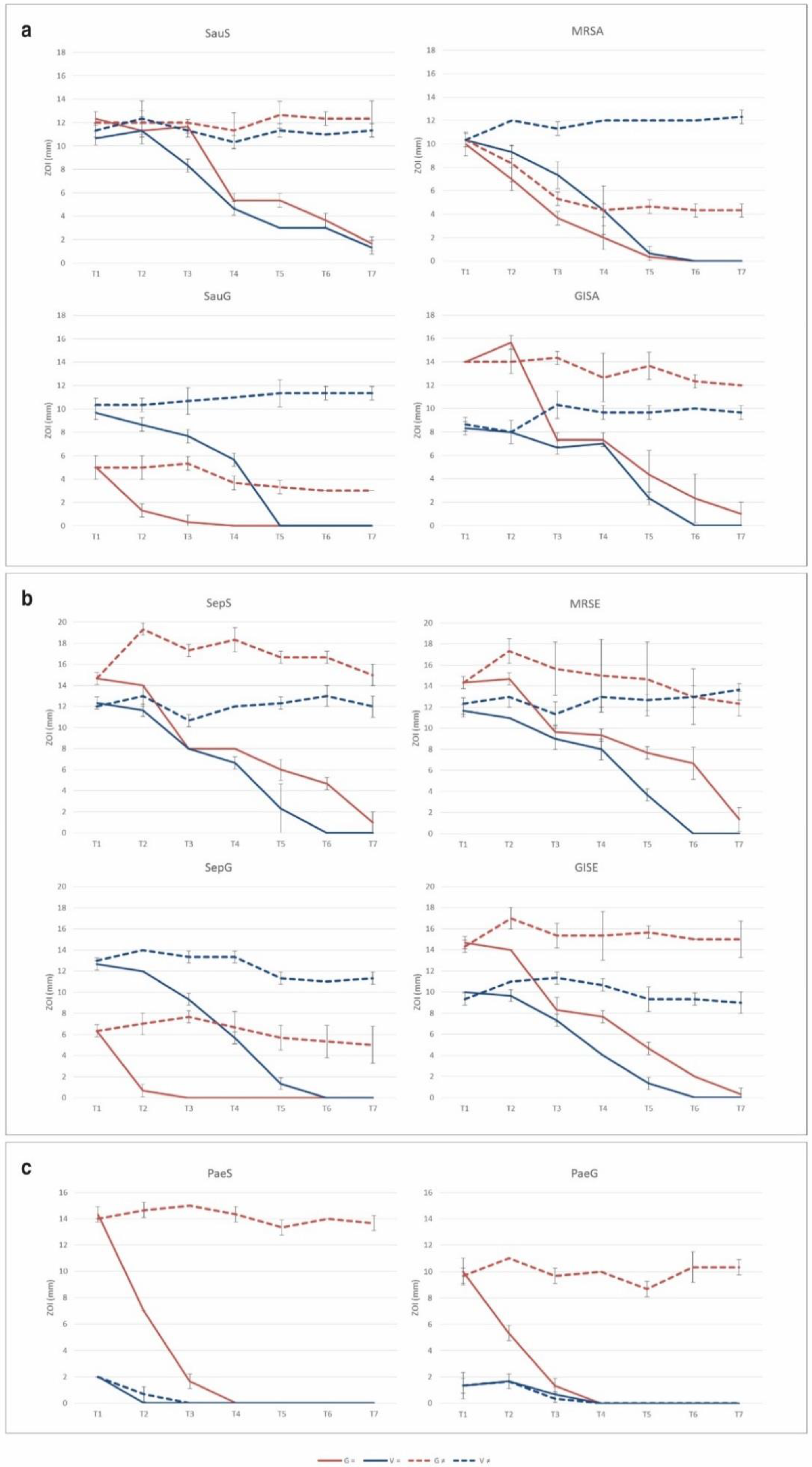
2.3. Resistance Selection to Eluted Vancomycin and Gentamicin
3. Discussion
3.1. Bacterial Adhesion
3.2. Evaluation of Biofilm Formation
3.3. Resistance Selection by Eluted Vancomycin and Gentamicin
4. Materials and Methods
4.1. Tested Bacterial Strains and Biomaterials
4.2. Evaluation of Bacterial Adhesion on Material Surface
4.3. Analysis of Biofilm Formation by CLSM
4.4. Determination of Minimum Inhibitory Concentration
4.5. Evaluation of Selection for Bacterial Resistance after Exposure to Eluted Antibiotics
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| BGS | Bone Graft Substitute |
| IRI | Implant-related Infection |
| PJI | Prosthetic Joint Infection |
| FRI | Fracture-related Infection |
| cOM | Chronic Osteomyelitis |
| DFO | Diabetic Foot Osteomyelitis |
| MIC | Minimum Inhibitory Concentrations |
| CBVF | Ceramic Bone Void Filler |
| CG | Ceramic Filler Containing Gentamicin |
| CV | Ceramic Filler Containing Vancomycin |
| CLSM | Confocal Laser Scan Microscopy |
| ZOI | Zone of Inhibition |
References
- Kapadia, B.H.; Berg, R.A.; Daley, J.A.; Fritz, J.; Bhave, A.; Mont, M.A. Periprosthetic joint infection. Lancet 2016, 387, 386–394. [Google Scholar] [CrossRef]
- Izakovicova, P.; Borens, O.; Trampuz, A. Periprosthetic joint infection: Current concepts and outlook. EFORT Open Rev. 2019, 4, 482–494. [Google Scholar] [CrossRef] [PubMed]
- Osmon, D.R.; Berbari, E.F.; Berendt, A.R.; Lew, D.; Zimmerli, W.; Steckelberg, J.M.; Rao, N.; Hanssen, A.; Wilson, W.R.; Infectious Diseases Society of America. Diagnosis and management of prosthetic joint infection: Clinical practice guidelines by the Infectious Diseases Society of America. Clin. Infect. Dis. 2013, 56, e1–e25. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zimmerli, W.; Trampuz, A.; Ochsner, P.E. Prosthetic-joint infections. N. Engl. J. Med. 2004, 351, 1645–1654. [Google Scholar] [CrossRef] [Green Version]
- Govaert, G.A.M.; Kuehl, R.; Atkins, B.L.; Trampuz, A.; Morgenstern, M.; Obremskey, W.T.; Verhofstad, M.H.J.; McNally, M.A.; Metsemakers, W.J.; Fracture-Related Infection Consensus Group. Diagnosing Fracture-Related Infection: Current Concepts and Recommendations. J. Orthop. Trauma 2020, 34, 8–17. [Google Scholar] [CrossRef]
- Metsemakers, W.J.; Morgenstern, M.; McNally, M.A.; Moriarty, T.F.; McFadyen, I.; Scarborough, M.; Athanasou, N.A.; Ochsner, P.E.; Kuehl, R.; Raschke, M.; et al. Fracture-related infection: A consensus on definition from an international expert group. Injury 2018, 49, 505–510. [Google Scholar] [CrossRef] [Green Version]
- Drago, L.; De Vecchi, E.; Bortolin, M.; Zagra, L.; Romano, C.L.; Cappelletti, L. Epidemiology and Antibiotic Resistance of Late Prosthetic Knee and Hip Infections. J. Arthroplast. 2017, 32, 2496–2500. [Google Scholar] [CrossRef]
- Bjarnsholt, T.; Ciofu, O.; Molin, S.; Givskov, M.; Hoiby, N. Applying insights from biofilm biology to drug development—Can a new approach be developed? Nat. Rev. Drug Discov. 2013, 12, 791–808. [Google Scholar] [CrossRef]
- Hall-Stoodley, L.; Stoodley, P. Evolving concepts in biofilm infections. Cell. Microbiol. 2009, 11, 1034–1043. [Google Scholar] [CrossRef]
- Brauner, A.; Fridman, O.; Gefen, O.; Balaban, N.Q. Distinguishing between resistance, tolerance and persistence to antibiotic treatment. Nat. Rev. Microbiol. 2016, 14, 320–330. [Google Scholar] [CrossRef]
- Lalidou, F.; Kolios, G.; Drosos, G.I. Bone infections and bone graft substitutes for local antibiotic therapy. Surg. Technol. Int. 2014, 24, 353–362. [Google Scholar] [PubMed]
- Metsemakers, W.J.; Fragomen, A.T.; Moriarty, T.F.; Morgenstern, M.; Egol, K.A.; Zalavras, C.; Obremskey, W.T.; Raschke, M.; McNally, M.A.; Fracture-Related Infection (FRI) Consensus Group. Evidence-Based Recommendations for Local Antimicrobial Strategies and Dead Space Management in Fracture-Related Infection. J. Orthop. Trauma 2020, 34, 18–29. [Google Scholar] [CrossRef] [PubMed]
- Trampuz, A.; Zimmerli, W. Diagnosis and treatment of implant-associated septic arthritis and osteomyelitis. Curr. Infect. Dis. Rep. 2008, 10, 394–403. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Winkler, H. Treatment of chronic orthopaedic infection. EFORT Open Rev. 2017, 2, 110–116. [Google Scholar] [CrossRef]
- Inzana, J.A.; Schwarz, E.M.; Kates, S.L.; Awad, H.A. Biomaterials approaches to treating implant-associated osteomyelitis. Biomaterials 2016, 81, 58–71. [Google Scholar] [CrossRef] [Green Version]
- Stravinskas, M.; Horstmann, P.; Ferguson, J.; Hettwer, W.; Nilsson, M.; Tarasevicius, S.; Petersen, M.M.; McNally, M.A.; Lidgren, L. Pharmacokinetics of gentamicin eluted from a regenerating bone graft substitute: In vitro and clinical release studies. Bone Jt. Res. 2016, 5, 427–435. [Google Scholar] [CrossRef]
- Stravinskas, M.; Nilsson, M.; Horstmann, P.; Petersen, M.M.; Tarasevicius, S.; Lidgren, L. Antibiotic Containing Bone Substitute in Major Hip Surgery: A Long Term Gentamicin Elution Study. J. Bone Jt. Infect. 2018, 3, 68–72. [Google Scholar] [CrossRef] [Green Version]
- Stravinskas, M.; Nilsson, M.; Vitkauskiene, A.; Tarasevicius, S.; Lidgren, L. Vancomycin elution from a biphasic ceramic bone substitute. Bone Jt. Res. 2019, 8, 49–54. [Google Scholar] [CrossRef]
- Butini, M.E.; Cabric, S.; Trampuz, A.; Di Luca, M. In vitro anti-biofilm activity of a biphasic gentamicin-loaded calcium sulfate/hydroxyapatite bone graft substitute. Colloids Surf. B Biointerfaces 2018, 161, 252–260. [Google Scholar] [CrossRef]
- Ferguson, J.; Athanasou, N.; Diefenbeck, M.; McNally, M. Radiographic and Histological Analysis of a Synthetic Bone Graft Substitute Eluting Gentamicin in the Treatment of Chronic Osteomyelitis. J. Bone Jt. Infect. 2019, 4, 76–84. [Google Scholar] [CrossRef] [Green Version]
- McNally, M.A.; Ferguson, J.Y.; Lau, A.C.; Diefenbeck, M.; Scarborough, M.; Ramsden, A.J.; Atkins, B.L. Single-stage treatment of chronic osteomyelitis with a new absorbable, gentamicin-loaded, calcium sulphate/hydroxyapatite biocomposite: A prospective series of 100 cases. Bone Jt. J. 2016, 98, 1289–1296. [Google Scholar] [CrossRef] [Green Version]
- Logoluso, N.; Drago, L.; Gallazzi, E.; George, D.A.; Morelli, I.; Romano, C.L. Calcium-Based, Antibiotic-Loaded Bone Substitute as an Implant Coating: A Pilot Clinical Study. J. Bone Jt. Infect. 2016, 1, 59–64. [Google Scholar] [CrossRef] [PubMed]
- Trampuz, A.; Widmer, A.F. Infections associated with orthopedic implants. Curr. Opin. Infect. Dis. 2006, 19, 349–356. [Google Scholar] [CrossRef] [PubMed]
- Aljawadi, A.; Islam, A.; Jahangir, N.; Niazi, N.; Ferguson, Z.; Sephton, B.; Elmajee, M.; Reid, A.; Wong, J.; Pillai, A. Adjuvant Local Antibiotic Hydroxyapatite Bio-Composite in the management of open Gustilo Anderson IIIB fractures. Prospective Review of 80 Patients from the Manchester Ortho-Plastic Unit. J. Orthop. 2020, 18, 261–266. [Google Scholar] [CrossRef]
- Drampalos, E.; Mohammad, H.R.; Pillai, A. Augmented debridement for implant related chronic osteomyelitis with an absorbable, gentamycin loaded calcium sulfate/hydroxyapatite biocomposite. J. Orthop. 2020, 17, 173–179. [Google Scholar] [CrossRef]
- Niazi, N.S.; Drampalos, E.; Morrissey, N.; Jahangir, N.; Wee, A.; Pillai, A. Adjuvant antibiotic loaded bio composite in the management of diabetic foot osteomyelitis—A multicentre study. Foot 2019, 39, 22–27. [Google Scholar] [CrossRef]
- Pesch, S.; Hanschen, M.; Greve, F.; Zyskowski, M.; Seidl, F.; Kirchhoff, C.; Biberthaler, P.; Huber-Wagner, S. Treatment of fracture-related infection of the lower extremity with antibiotic-eluting ceramic bone substitutes: Case series of 35 patients and literature review. Infection 2020, 48, 333–344. [Google Scholar] [CrossRef]
- Hutting, K.; van Netten, J.; Dening, J.; Cate, W.T.; van Baal, J. Surgical debridement and gentamicin-loaded calcium sulphate/hydroxyapatite bone void filling to treat diabetic foot osteomyelitis. Diabet. Foot J. 2019, 22, 22–27. [Google Scholar]
- Labby, K.J.; Garneau-Tsodikova, S. Strategies to overcome the action of aminoglycoside-modifying enzymes for treating resistant bacterial infections. Future Med. Chem. 2013, 5, 1285–1309. [Google Scholar] [CrossRef] [Green Version]
- Carniello, V.; Peterson, B.W.; van der Mei, H.C.; Busscher, H.J. Physico-chemistry from initial bacterial adhesion to surface-programmed biofilm growth. Adv. Colloid Interface Sci. 2018, 261, 1–14. [Google Scholar] [CrossRef]
- Torbert, J.T.; Joshi, M.; Moraff, A.; Matuszewski, P.E.; Holmes, A.; Pollak, A.N.; O’Toole, R.V. Current bacterial speciation and antibiotic resistance in deep infections after operative fixation of fractures. J. Orthop. Trauma 2015, 29, 7–17. [Google Scholar] [CrossRef] [PubMed]
- Stokes, J.M.; French, S.; Ovchinnikova, O.G.; Bouwman, C.; Whitfield, C.; Brown, E.D. Cold Stress Makes Escherichia coli Susceptible to Glycopeptide Antibiotics by Altering Outer Membrane Integrity. Cell Chem. Biol. 2016, 23, 267–277. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hausner, M.; Wuertz, S. High rates of conjugation in bacterial biofilms as determined by quantitative in situ analysis. Appl. Environ. Microbiol. 1999, 65, 3710–3713. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Madsen, J.S.; Burmolle, M.; Hansen, L.H.; Sorensen, S.J. The interconnection between biofilm formation and horizontal gene transfer. FEMS Immunol. Med Microbiol. 2012, 65, 183–195. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Molin, S.; Tolker-Nielsen, T. Gene transfer occurs with enhanced efficiency in biofilms and induces enhanced stabilisation of the biofilm structure. Curr. Opin. Biotechnol. 2003, 14, 255–261. [Google Scholar] [CrossRef]
- Savage, V.J.; Chopra, I.; O’Neill, A.J. Staphylococcus aureus biofilms promote horizontal transfer of antibiotic resistance. Antimicrob. Agents Chemother. 2013, 57, 1968–1970. [Google Scholar] [CrossRef] [Green Version]
- Boles, B.R.; Singh, P.K. Endogenous oxidative stress produces diversity and adaptability in biofilm communities. Proc. Natl. Acad. Sci. USA 2008, 105, 12503–12508. [Google Scholar] [CrossRef] [Green Version]
- Driffield, K.; Miller, K.; Bostock, J.M.; O’Neill, A.J.; Chopra, I. Increased mutability of Pseudomonas aeruginosa in biofilms. J. Antimicrob. Chemother. 2008, 61, 1053–1056. [Google Scholar] [CrossRef] [Green Version]
- Vaudaux, P.; Ferry, T.; Uckay, I.; Francois, P.; Schrenzel, J.; Harbarth, S.; Renzoni, A. Prevalence of isolates with reduced glycopeptide susceptibility in orthopedic device-related infections due to methicillin-resistant Staphylococcus aureus. Eur. J. Clin. Microbiol. Infect. Dis. 2012, 31, 3367–3374. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Li, X.; Jiang, L.; Han, W.; Xie, X.; Jin, Y.; He, X.; Wu, R. Novel Mutation Sites in the Development of Vancomycin- Intermediate Resistance in Staphylococcus aureus. Front. Microbiol. 2016, 7, 2163. [Google Scholar] [CrossRef] [Green Version]
- Hettwer, W.; Horstmann, P.F.; Bischoff, S.; Gullmar, D.; Reichenbach, J.R.; Poh, P.S.P.; van Griensven, M.; Gras, F.; Diefenbeck, M. Establishment and effects of allograft and synthetic bone graft substitute treatment of a critical size metaphyseal bone defect model in the sheep femur. APMIS Acta Pathol. Microbiol. Immunol. Scand. 2019, 127, 53–63. [Google Scholar] [CrossRef] [PubMed]
- Kyriacou, H.; Kamaraj, A.; Khan, W.S. Developments in Antibiotic-Eluting Scaffolds for the Treatment of Osteomyelitis. Appl. Sci. 2020, 10, 2244. [Google Scholar] [CrossRef] [Green Version]
- Azeredo, J.; Azevedo, N.F.; Briandet, R.; Cerca, N.; Coenye, T.; Costa, A.R.; Desvaux, M.; Di Bonaventura, G.; Hebraud, M.; Jaglic, Z.; et al. Critical review on biofilm methods. Crit. Rev. Microbiol. 2017, 43, 313–351. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bidossi, A.; Bottagisio, M.; De Grandi, R.; De Vecchi, E. Ability of adhesion and biofilm formation of pathogens of periprosthetic joint infections on titanium-niobium nitride (TiNbN) ceramic coatings. J. Orthop. Surg. Res. 2020, 15, 90. [Google Scholar] [CrossRef] [Green Version]
- De Vecchi, E.; Bottagisio, M.; Bortolin, M.; Toscano, M.; Lovati, A.B.; Drago, L. Improving the Bacterial Recovery by Using Dithiothreitol with Aerobic and Anaerobic Broth in Biofilm-Related Prosthetic and Joint Infections. Adv. Exp. Med. Biol. 2017, 973, 31–39. [Google Scholar] [CrossRef]
- Herigstad, B.; Hamilton, M.; Heersink, J. How to optimize the drop plate method for enumerating bacteria. J. Microbiol. Methods 2001, 44, 121–129. [Google Scholar] [CrossRef]
- Bidossi, A.; De Grandi, R.; Toscano, M.; Bottagisio, M.; De Vecchi, E.; Gelardi, M.; Drago, L. Probiotics Streptococcus salivarius 24SMB and Streptococcus oralis 89a interfere with biofilm formation of pathogens of the upper respiratory tract. BMC Infect. Dis. 2018, 18, 653. [Google Scholar] [CrossRef]
- EUCAST Guidelines. Available online: http://www.eucast.org/fileadmin/src/media/PDFs/EUCAST_files/MIC_testing/Edis5.1_broth_dilution.pdf (accessed on 3 December 2020).
- Karr, J.C.; Lauretta, J.; Keriazes, G. In vitro antimicrobial activity of calcium sulfate and hydroxyapatite (Cerament Bone Void Filler) discs using heat-sensitive and non-heat-sensitive antibiotics against methicillin-resistant Staphylococcus aureus and Pseudomonas aeruginosa. J. Am. Podiatr. Med. Assoc. 2011, 101, 146–152. [Google Scholar] [CrossRef]
- Drago, L.; De Vecchi, E.; Nicola, L.; Tocalli, L.; Gismondo, M.R. In vitro selection of resistance in Pseudomonas aeruginosa and Acinetobacter spp. by levofloxacin and ciprofloxacin alone and in combination with beta-lactams and amikacin. J. Antimicrob. Chemother. 2005, 56, 353–359. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| CBVF | CG | CV | ||
|---|---|---|---|---|
| S. aureus | SauS | 8.6 | 36.7 | 39.5 |
| SauG | 7.7 | 12.1 | 11.6 | |
| MRSA | 7.9 | 55.3 | 38.6 | |
| GISA | 14.2 | 48.9 | 20.6 | |
| S. epidermidis | SepS | 7.7 | 26.2 | 20.6 |
| SepG | 15.8 | 16.8 | 52.3 | |
| MRSE | 5.0 | 31.2 | 12.4 | |
| GISE | 7.8 | 40.3 | 18.2 | |
| P. aeruginosa | PaeS | 6.0 | 26.8 | 19.6 |
| PaeG | 11.4 | 25.2 | 18.6 |
| T0 | T7 (=) | T14 (=) | T7 (≠) | T14 (≠) | ||
|---|---|---|---|---|---|---|
| S. aureus | SauS | 0.5 | 0.125–1 | 0.5 | 0.25–0.5 | 0.25 |
| MRSA | 0.5 | 0.125–1 | 0.5 | 1 | 1 | |
| SauG | 256 | 256–512 | 256–512 | 512 | 512 | |
| GISA | 0.25 | 0.25–0.5 | 0.25 | 0.5–1 | 0.5 | |
| S. epidermidis | SepS | 0.125 | 0.125–0.25 | 0.125 | 0.125–0.25 | 0.125–0.25 |
| MRSE | 0.25 | 0.125–0.25 | 0.125–0.25 | 0.25 | 0.125–0.25 | |
| SepG | 128 | 256 | 128 | 128–256 | 128–256 | |
| GISE | 0.125 | 0.125 | 0.125 | 0.125 | 0.125 | |
| P. aeruginosa | PaeS | 0.125 | 0.125 | 0.125 | 0.25 | 0.125–0.125 |
| PaeG | 16 | 8 | 8–16 | 8 | 8–16 |
| T0 | T7 (=) | T14 (=) | T7 (≠) | T14 (≠) | ||
|---|---|---|---|---|---|---|
| S. aureus | SauS | 0.25 | 0.125–0.25 | 0.25 | 0.5 | 0.25 |
| MRSA | 1 | 0.5–1 | 0.5 | 2 | 1–2 | |
| SauG | 0.5 | 0.5–1 | 0.5 | 1 | 0.5 | |
| GISA | 4 | 4 | 2–4 | 4 | 2–4 | |
| S. epidermidis | SepS | 1 | 2 | 1 | 1–2 | 1 |
| MRSE | 0.125 | 0.125 | 0.125 | 0.125 | 0.125 | |
| SepG | 1 | 1–2 | 1 | 2 | 2 | |
| GISE | 4 | 4–8 | 4 | 4–8 | 4 | |
| P. aeruginosa | PaeS | >1024 | >1024 | >1024 | >1024 | >1024 |
| PaeG | >1024 | >1024 | >1024 | >1024 | >1024 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bidossi, A.; Bottagisio, M.; Logoluso, N.; De Vecchi, E. In Vitro Evaluation of Gentamicin or Vancomycin Containing Bone Graft Substitute in the Prevention of Orthopedic Implant-Related Infections. Int. J. Mol. Sci. 2020, 21, 9250. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21239250
Bidossi A, Bottagisio M, Logoluso N, De Vecchi E. In Vitro Evaluation of Gentamicin or Vancomycin Containing Bone Graft Substitute in the Prevention of Orthopedic Implant-Related Infections. International Journal of Molecular Sciences. 2020; 21(23):9250. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21239250
Chicago/Turabian StyleBidossi, Alessandro, Marta Bottagisio, Nicola Logoluso, and Elena De Vecchi. 2020. "In Vitro Evaluation of Gentamicin or Vancomycin Containing Bone Graft Substitute in the Prevention of Orthopedic Implant-Related Infections" International Journal of Molecular Sciences 21, no. 23: 9250. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21239250