A Topical Formulation of Melatoninergic Compounds Exerts Strong Hypotensive and Neuroprotective Effects in a Rat Model of Hypertensive Glaucoma

 , , and
, , and 
Abstract
:1. Introduction
2. Results
2.1. Melatonin/Agomelatine in the Eye
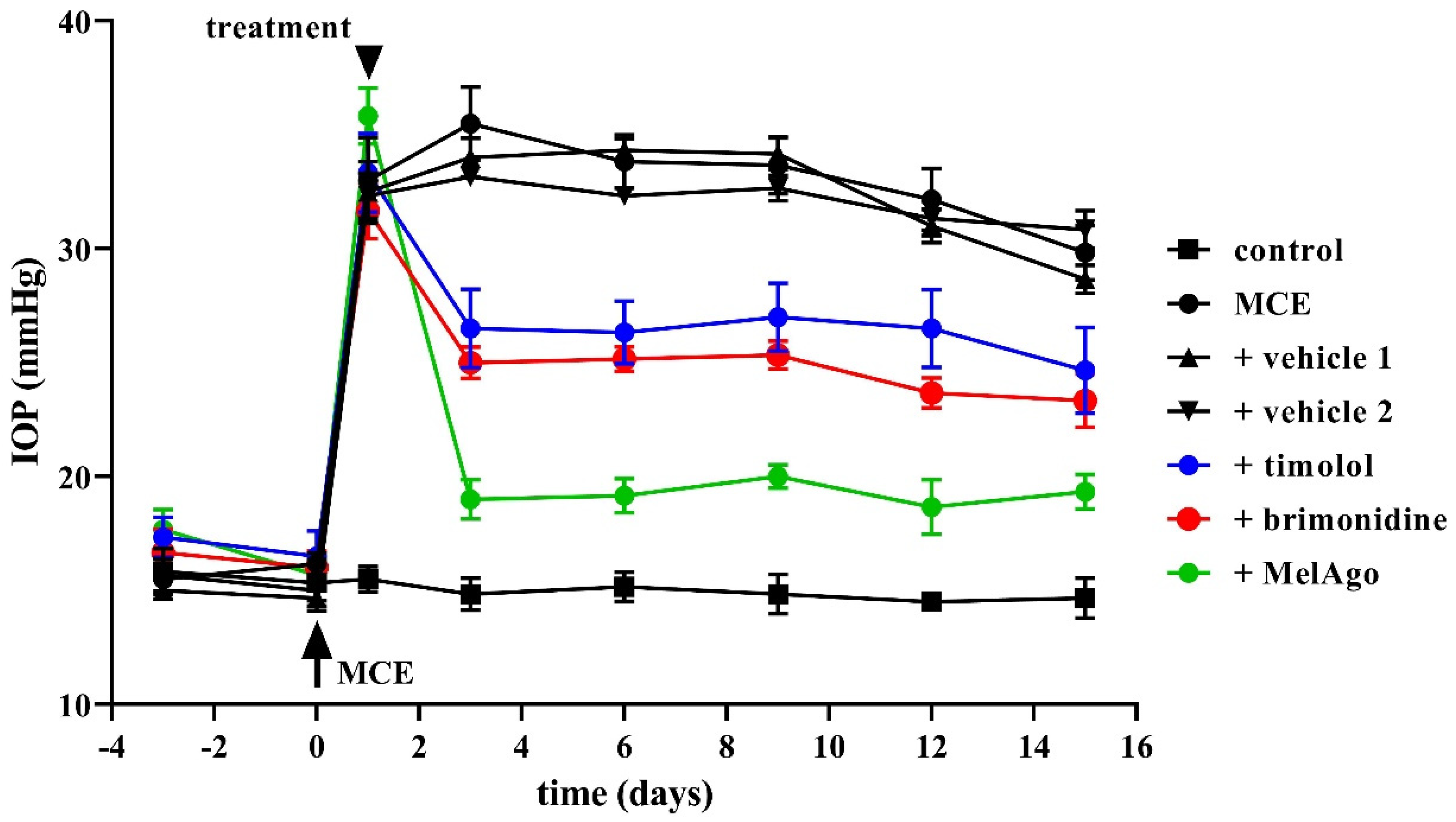
2.2. Effects on IOP Elevation
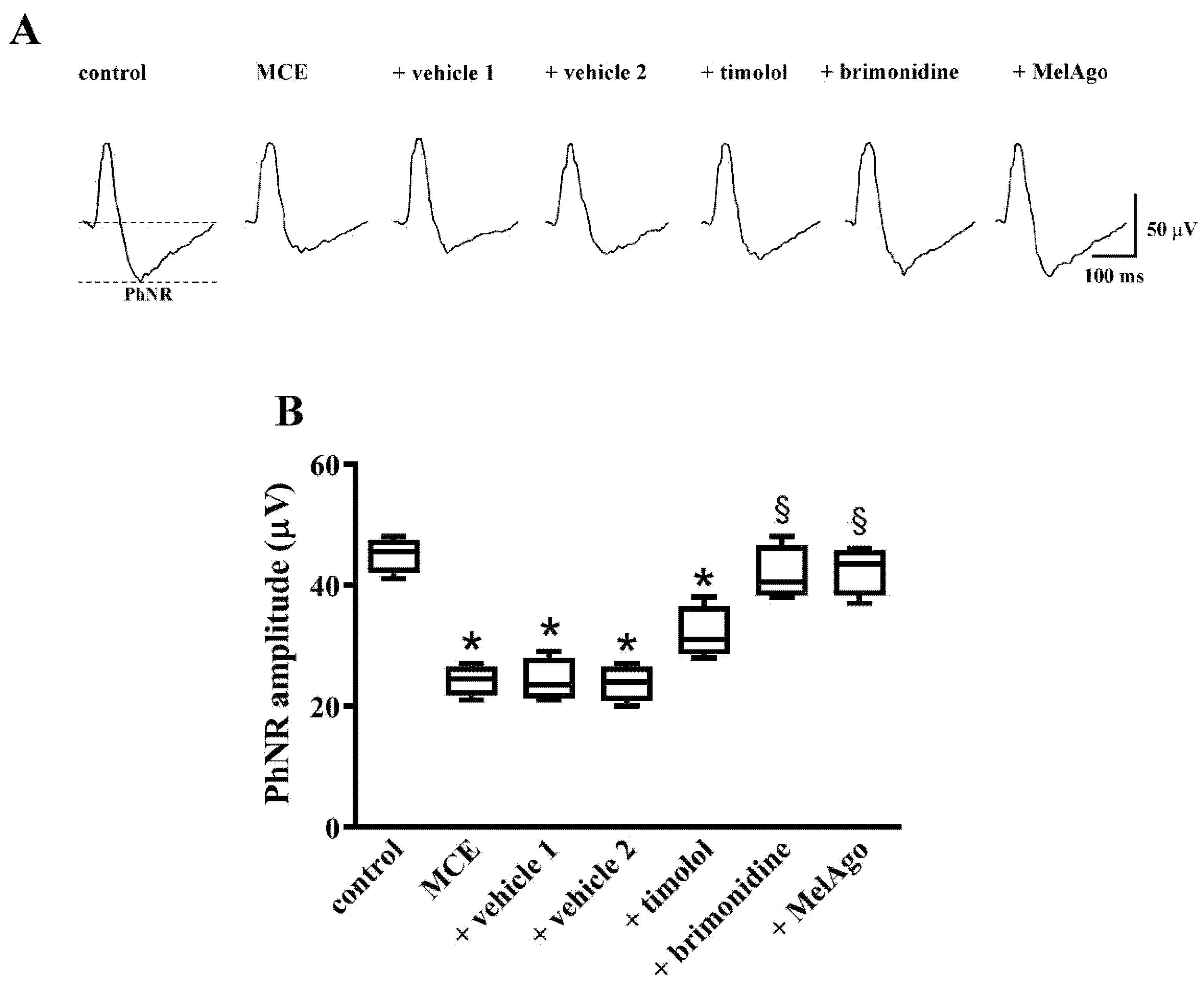
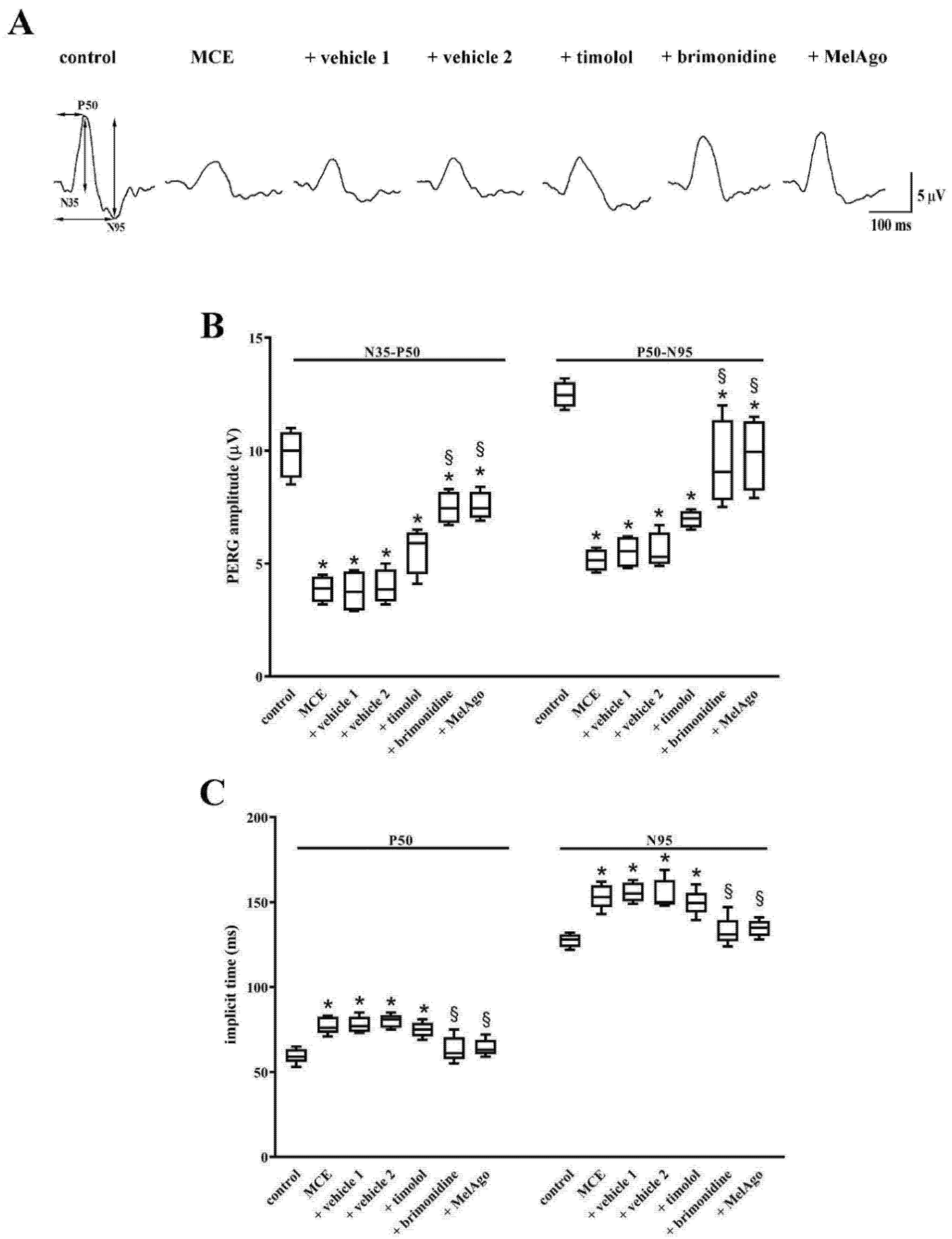
2.3. Effects on Retinal Dysfunction: PhNR and PERG
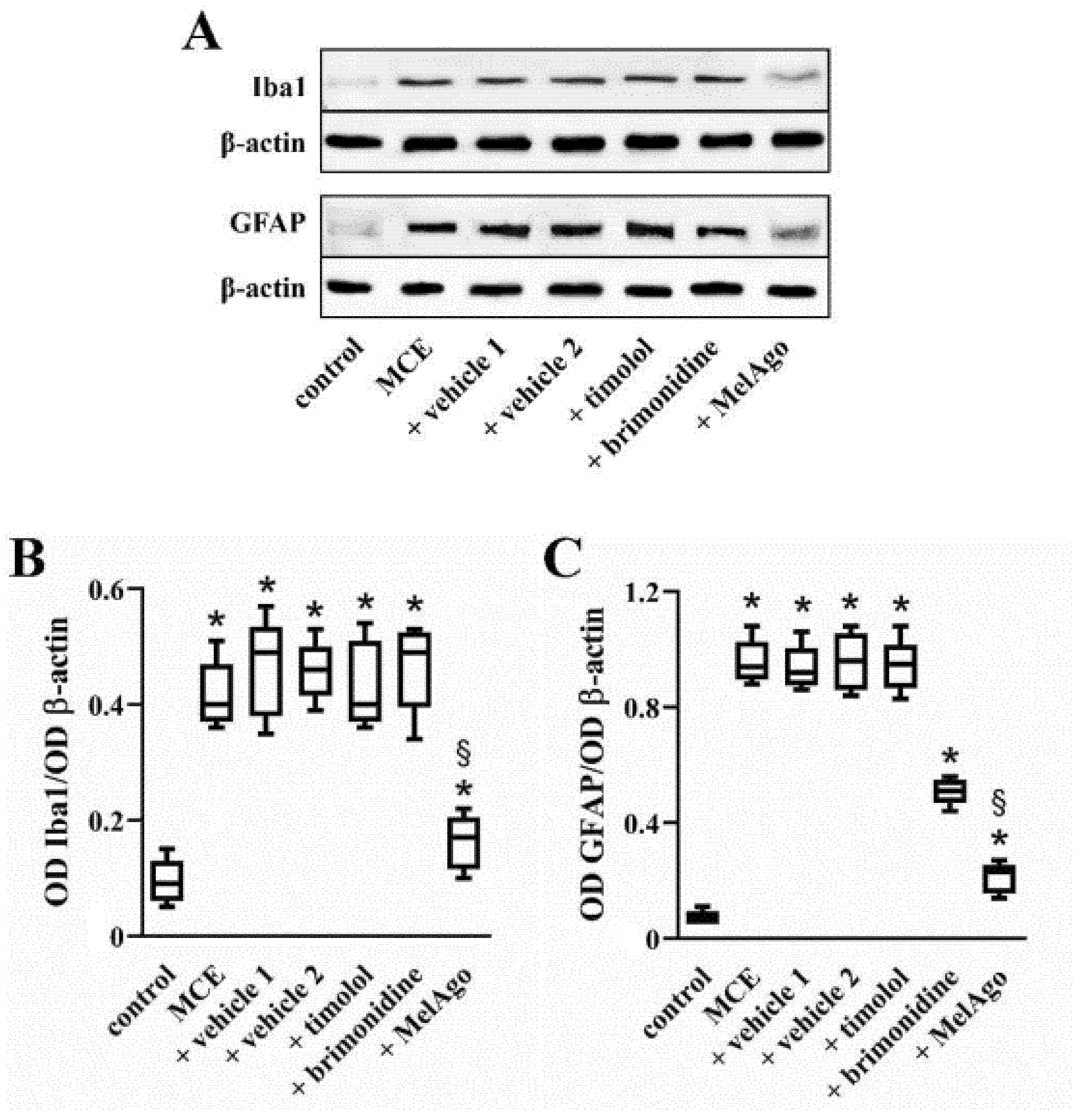
2.4. Effects on Gliosis-Related Inflammation
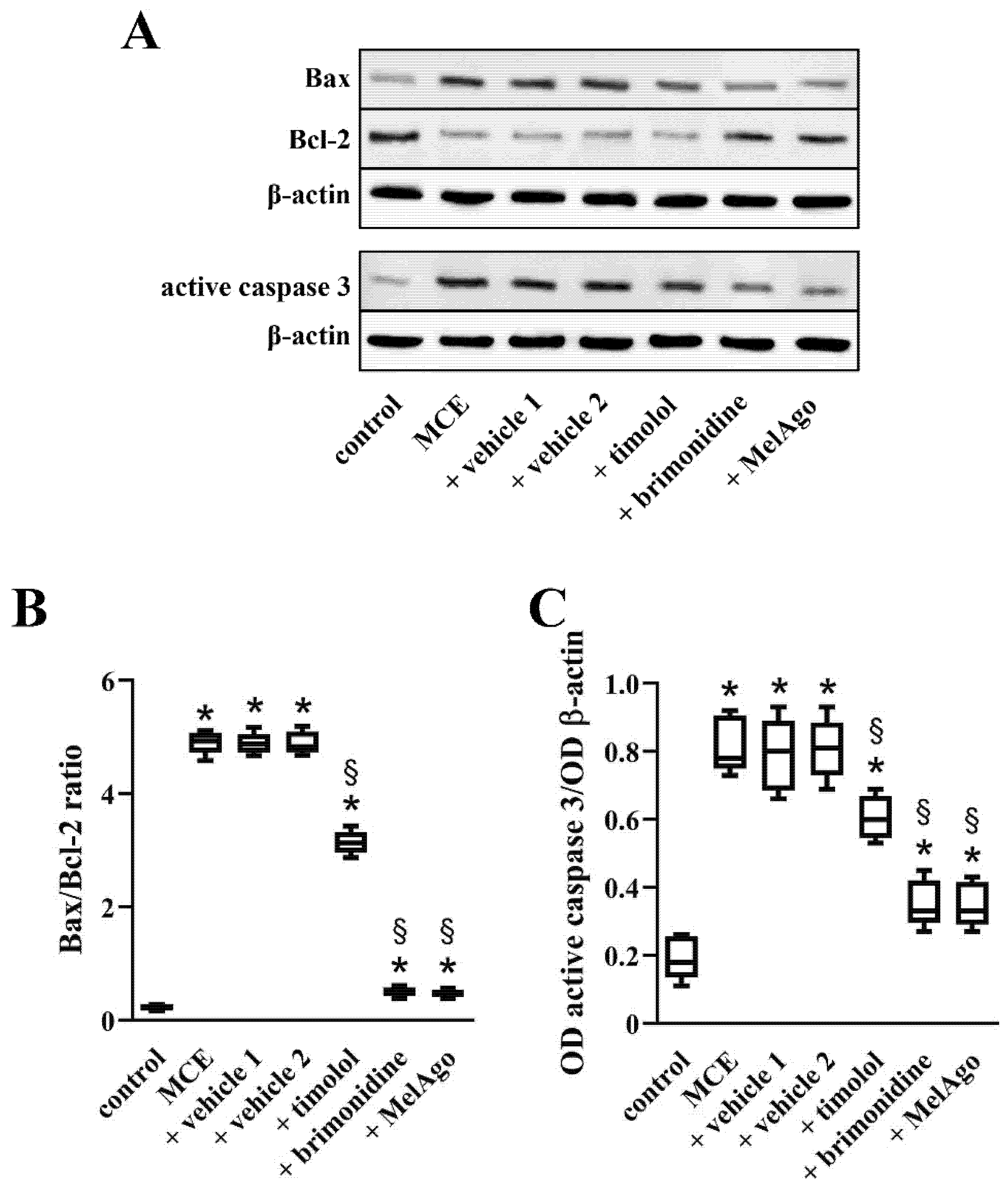
2.5. Effects on Bax-Caspase-3-Dependent Apoptotic Pathway
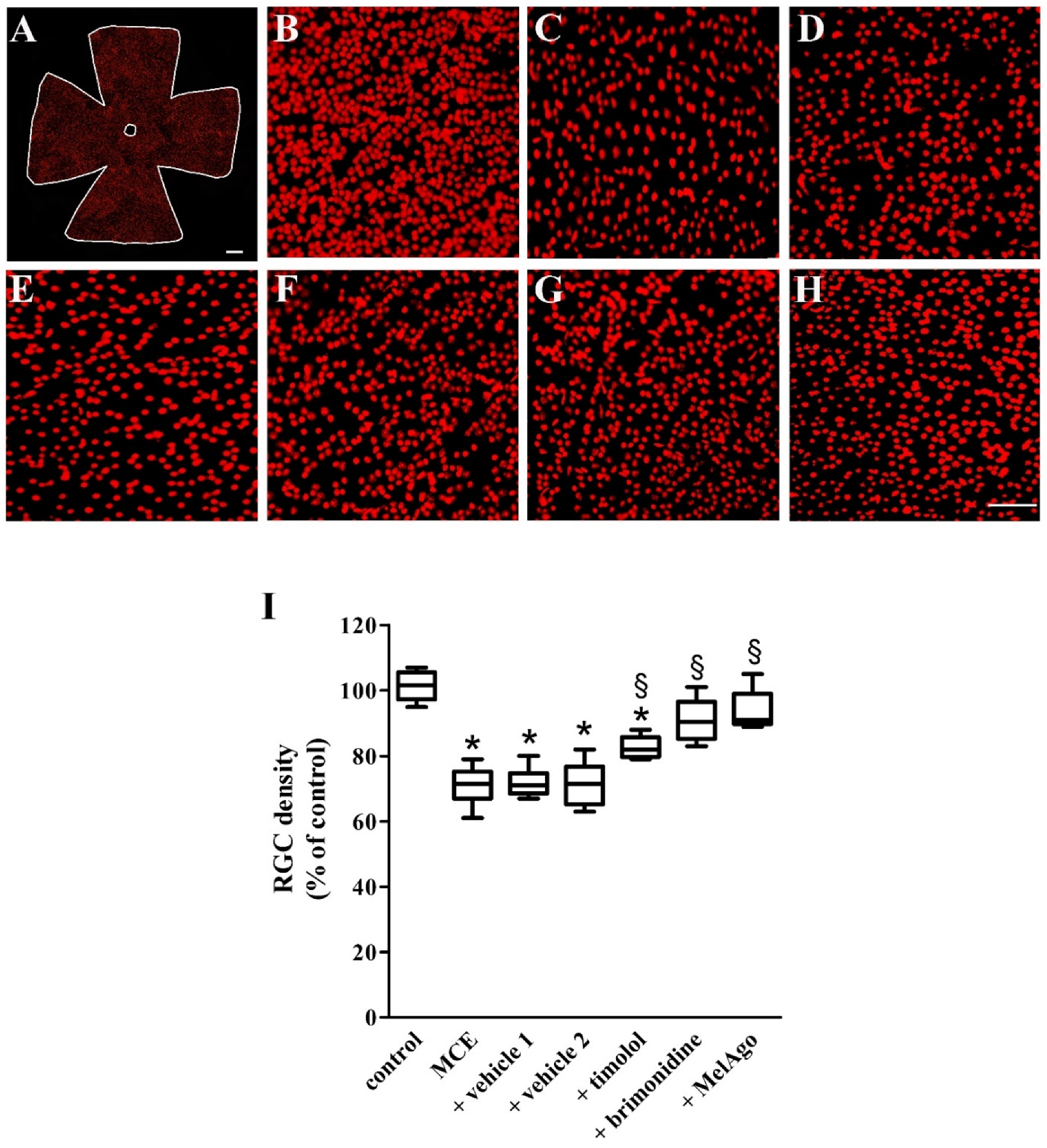
2.6. Effects on RGCs Loss
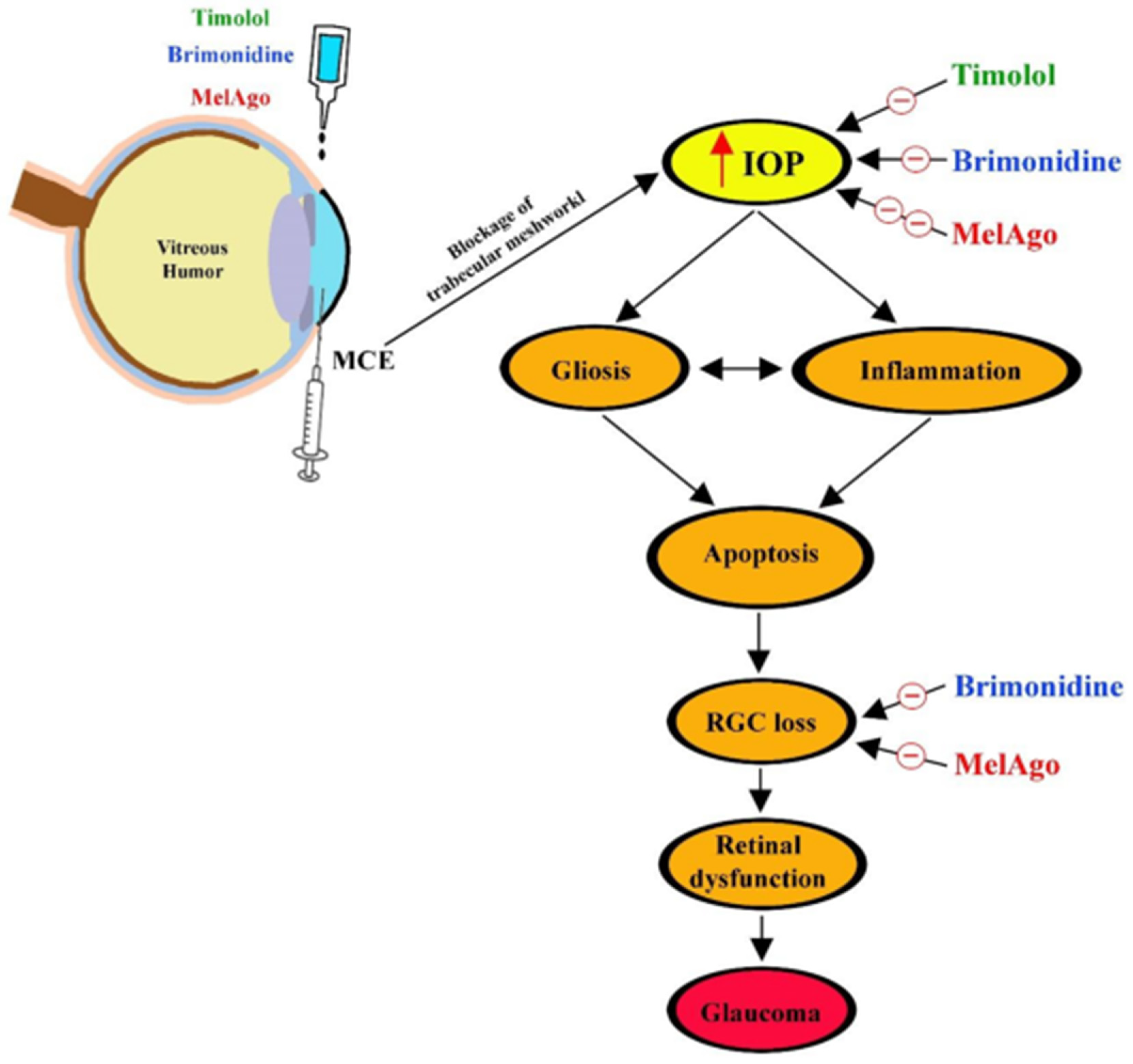
3. Discussion
3.1. The MCE Model
3.2. Hypotensive and Protective Efficacy of Timolol and Brimonidine
3.3. Distribution of Melatonin and Agomelatine
3.4. Hypotensive and Protective Efficacy of Melatonin/Agomelatine
3.5. Mechanisms Leading to RGC protection
4. Materials and Methods
4.1. Animals
4.2. Preparation of Eye Drops Containing Melatonin/Agomelatine
4.3. Pharmacokinetics Study
4.4. Experimental Model of Intraocular Hypertension
4.5. Eye Drop Administration
4.6. IOP Measurement and Quantitation
4.7. Evaluation of Retinal Function
4.8. Western Blot Analysis
4.9. Retinal Ganglion Cell Immunohistochemistry and Quantitation
4.10. Data Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| Brn3a | Brain-specific homeobox/POU domain protein 3A |
| ERG | Electroretinogram |
| GFAP | Glial fibrillary acidic protein |
| Iba1 | Ionized calcium binding adaptor molecule 1 |
| IL | Interleukin |
| IOP | Intraocular pressure |
| MCE | Methylcellulose |
| OD | Optical density |
| PB | Phosphate buffer |
| PERG | Pattern electroretinogram |
| PhNR | Photopic negative response |
| RGC | Retinal ganglion cell |
| TNF | Tumor necrosis factor |
References
- Samanta, S. Physiological and pharmacological perspectives of melatonin. Arch. Physiol. Biochem. 2020. [Google Scholar] [CrossRef] [PubMed]
- Ko, G.Y. Circadian regulation in the retina: From molecules to network. Eur. J. Neurosci. 2020, 51, 194–216. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Juybari, K.B.; Hosseinzadeh, A.; Ghaznavi, H.; Kamali, M.; Sedaghat, A.; Mehrzadi, S.; Naseripour, M. Melatonin as a modulator of degenerative and regenerative signaling pathways in injured retinal ganglion cells. Curr. Pharm. Des. 2019, 25, 3057–3073. [Google Scholar] [CrossRef] [PubMed]
- Alkozi, H.A.; Navarro, G.; Franco, R.; Pintor, J. Melatonin and the control of intraocular pressure. Prog. Retin. Eye Res. 2020, 75, 100798. [Google Scholar] [CrossRef] [PubMed]
- Kastner, A.; King, A.J. Advanced glaucoma at diagnosis: Current perspectives. Eye (Lond) 2020, 34, 116–128. [Google Scholar] [CrossRef] [PubMed]
- Kapetanakis, V.V.; Chan, M.P.; Foster, P.J.; Cook, D.G.; Owen, C.G.; Rudnicka, A.R. Global variations and time trends in the prevalence of primary open angle glaucoma (POAG): A systematic review and meta-analysis. Br. J. Ophthalmol. 2016, 100, 86–93. [Google Scholar] [CrossRef]
- Sheybani, A.; Scott, R.; Samuelson, T.W.; Kahook, M.Y.; Bettis, D.I.; Ahmed, I.I.K.; Stephens, J.D.; Kent, D.; Ferguson, T.J.; Herndon, L.W. Open-angle glaucoma: Burden of illness, current Therapies, and the management of nocturnal IOP variation. Ophthalmol. Ther. 2020, 9, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Zhang, H.J.; Mi, X.S.; So, K.F. Normal tension glaucoma: From the brain to the eye or the inverse? Neural Regen. Res. 2019, 14, 1845–1850. [Google Scholar] [CrossRef]
- Hu, X.; Xu, M.X.; Zhou, H.; Cheng, S.; Li, F.; Miao, Y.; Wang, Z. Tumor necrosis factor-alpha aggravates gliosis and inflammation of activated retinal Müller cells. Biochem. Biophys. Res. Commun. 2020, 531, 383–389. [Google Scholar] [CrossRef]
- Alqawlaq, S.; Flanagan, J.G.; Sivak, J.M. All roads lead to glaucoma: Induced retinal injury cascades contribute to a common neurodegenerative outcome. Exp. Eye Res. 2019, 183, 88–97. [Google Scholar] [CrossRef]
- Lusthaus, J.; Goldberg, I. Current management of glaucoma. Med. J. Aust. 2019, 210, 180–187. [Google Scholar] [CrossRef] [PubMed]
- Chua, B.; Goldberg, I. Neuroprotective agents in glaucoma therapy: Recent developments and future directions. Expert Rev. Ophthalmol. 2010, 5, 627–636. [Google Scholar] [CrossRef]
- Oh, D.J.; Chen, J.L.; Vajaranant, T.S.; Dikopf, M.S. Brimonidine tartrate for the treatment of glaucoma. Expert Opin. Pharmacother. 2019, 20, 115–122. [Google Scholar] [CrossRef] [PubMed]
- Krupin, T.; Liebmann, J.M.; Greenfield, D.S.; Ritch, R.; Gardiner, S.; Low-Pressure Glaucoma Study Group. A randomized trial of brimonidine versus timolol in preserving visual function: Results from the low-pressure glaucoma treatment study. Am. J. Ophthalmol. 2011, 151, 671–681. [Google Scholar] [CrossRef] [PubMed]
- Morrone, L.A.; Rombola, L.; Adornetto, A.; Corasaniti, M.T.; Russo, R. Rational basis for nutraceuticals in the treatment of glaucoma. Curr. Neuropharmacol. 2018, 16, 1004–1017. [Google Scholar] [CrossRef]
- Rusciano, D.; Pezzino, S.; Mutolo, M.G.; Giannotti, R.; Librando, A.; Pescosolido, N. Neuroprotection in glaucoma: Old and new promising treatments. Adv. Pharmacol. Sci. 2017, 2017, 4320408. [Google Scholar] [CrossRef] [PubMed]
- Dal Monte, M.; Cammalleri, M.; Pezzino, S.; Corsaro, R.; Pescosolido, N.; Bagnoli, P.; Rusciano, D. Hypotensive effect of nanomicellar formulation of melatonin and agomelatine in a rat model: Significance for glaucoma therapy. Diagnostics (Basel) 2020, 10, 138. [Google Scholar] [CrossRef] [Green Version]
- Martínez-Águila, A.; Fonseca, B.; Pérez de Lara, M.J.; Pintor, J. Effect of melatonin and 5-Methoxycarbonylamino-N-Acetyltryptamine on the intraocular pressure of normal and glaucomatous mice. J. Pharmacol. Exp. Ther. 2016, 357, 293–299. [Google Scholar] [CrossRef] [Green Version]
- Crooke, A.; Huete-Toral, F.; Martínez-Águila, A.; Martín-Gil, A.; Pintor, J. Melatonin and its analog 5-methoxycarbonylamino-N-acetyltryptamine potentiate adrenergic receptor-mediated ocular hypotensive effects in rabbits: Significance for combination therapy in glaucoma. J. Pharmacol. Exp. Ther. 2013, 346, 138–145. [Google Scholar] [CrossRef] [Green Version]
- Martínez-Águila, A.; Fonseca, B.; Bergua, A.; Pintor, J. Melatonin analogue agomelatine reduces rabbit’s intraocular pressure in normotensive and hypertensive conditions. Eur. J. Pharmacol. 2013, 701, 213–217. [Google Scholar] [CrossRef]
- Alkozi, H.; Sánchez-Naves, J.; de Lara, M.J.; Carracedo, G.; Fonseca, B.; Martinez-Aguila, A.; Pintor, J. Elevated intraocular pressure increases melatonin levels in the aqueous humour. Acta Ophthalmol. 2017, 95, e185–e189. [Google Scholar] [CrossRef] [PubMed]
- Pescosolido, N.; Gatto, V.; Stefanucci, A.; Rusciano, D. Oral treatment with the melatonin agonist agomelatine lowers the intraocular pressure of glaucoma patients. Ophthalmic. Physiol. Opt. 2015, 35, 201–205. [Google Scholar] [CrossRef] [PubMed]
- Diéguez, H.H.; González Fleitas, M.F.; Aranda, M.L.; Calanni, J.S.; Keller Sarmiento, M.I.; Chianelli, M.S.; Alaimo, A.; Sande, P.H.; Romeo, H.E.; Rosenstein, R.E.; et al. Melatonin protects the retina from experimental nonexudative age-related macular degeneration in mice. J. Pineal Res. 2020, 68, e12643. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Tian, Y.; Yao, A.; Zha, X.; Zhang, J.; Tao, Y. Intravitreal delivery of melatonin is protective against the photoreceptor loss in mice: A potential therapeutic strategy for degenerative retinopathy. Front. Pharmacol. 2020, 10, 1633. [Google Scholar] [CrossRef] [Green Version]
- Xu, X.J.; Wang, S.M.; Jin, Y.; Hu, Y.T.; Feng, K.; Ma, Z.Z. Melatonin delays photoreceptor degeneration in a mouse model of autosomal recessive retinitis pigmentosa. J. Pineal Res. 2017, 63, e12428. [Google Scholar] [CrossRef]
- Jiang, T.; Chang, Q.; Cai, J.; Fan, J.; Zhang, X.; Xu, G. Protective effects of melatonin on retinal inflammation and oxidative stress in experimental diabetic retinopathy. Oxid. Med. Cell. Longev. 2016, 2016, 3528274. [Google Scholar] [CrossRef] [Green Version]
- Park, S.W.; Lee, H.S.; Sung, M.S.; Kim, S.J. The effect of melatonin on retinal ganglion cell survival in ischemic retina. Chonnam. Med. J. 2012, 48, 116–122. [Google Scholar] [CrossRef] [Green Version]
- Del Valle Bessone, C.; Fajreldines, H.D.; de Barboza, G.E.D.; Tolosa de Talamoni, N.G.; Allemandi, D.A.; Carpentieri, A.R.; Quinteros, D.A. Protective role of melatonin on retinal ganglionar cell: In vitro an in vivo evidences. Life Sci. 2019, 218, 233–240. [Google Scholar] [CrossRef]
- Belforte, N.A.; Moreno, M.C.; de Zavalía, N.; Sande, P.H.; Chianelli, M.S.; Keller Sarmiento, M.I.; Rosenstein, R.E. Melatonin: A novel neuroprotectant for the treatment of glaucoma. J. Pineal Res. 2010, 48, 353–364. [Google Scholar] [CrossRef]
- Marangoz, D.; Guzel, E.; Eyuboglu, S.; Gumusel, A.; Seckin, I.; Ciftci, F.; Yilmaz, B.; Yalvac, I. Comparison of the neuroprotective effects of brimonidine tartrate and melatonin on retinal ganglion cells. Int. Ophthalmol. 2018, 38, 2553–2562. [Google Scholar] [CrossRef]
- Wilsey, L.J.; Fortune, B. Electroretinography in glaucoma diagnosis. Curr. Opin. Ophthalmol. 2016, 27, 118–124. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haider, A.; Shehzad, A.; Wahid, F.; Kumar, A.; Rao, K.M.; Han, S.S. The multi regulatory role of signal transducer and activator of transcription factor Brn-3a. J. Neurol. Neurosci. 2016, 7, 93. [Google Scholar] [CrossRef]
- Hernández, M.; Urcola, J.H.; Vecino, E. Retinal ganglion cell neuroprotection in a rat model of glaucoma following brimonidine, latanoprost or combined treatments. Exp. Eye Res. 2008, 86, 798–806. [Google Scholar] [CrossRef] [PubMed]
- Cammalleri, M.; Dal Monte, M.; Amato, R.; Bagnoli, P.; Rusciano, D. A dietary combination of forskolin with homotaurine, spearmint and B vitamins protects injured retinal ganglion cells in a rodent model of hypertensive glaucoma. Nutrients 2020, 12, 1189. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pescosolido, N.; Scarsella, G.; Rusciano, D. Oral administration of forskolin decreases retinal damage after experimental induction of ocular hypertension in the rat. In Forskolin—Sources, Mechanisms of Action and Health Effects; Walker, M.H., Ed.; Novinka: New York, NY, USA, 2015; pp. 31–50. [Google Scholar]
- Zhu, M.D.; Cai, F.Y. Development of experimental chronic intraocular hypertension in the rabbit. Aust. N. Z. J. Ophthalmol. 1992, 20, 225–234. [Google Scholar] [CrossRef]
- Huang, W.; Gao, F.; Hu, F.; Huang, J.; Wang, M.; Xu, P.; Zhang, R.; Chen, J.; Sun, X.; Zhang, S.; et al. Asiatic acid prevents retinal ganglion cell apoptosis in a rat model of glaucoma. J. Front. Neurosci. 2018, 12, 489. [Google Scholar] [CrossRef] [Green Version]
- Crowston, J.G.; Kong, Y.X.; Trounce, I.A.; Dang, T.M.; Fahy, E.T.; Bui, B.V.; Morrison, J.C.; Chrysostomou, V. An acute intraocular pressure challenge to assess retinal ganglion cell injury and recovery in the mouse. Exp. Eye Res. 2015, 141, 3–8. [Google Scholar] [CrossRef]
- Mélik Parsadaniantz, S.; Réaux-le Goazigo, A.; Sapienza, A.; Habas, C.; Baudouin, C. Glaucoma: A degenerative optic neuropathy related to neuroinflammation? Cells 2020, 9, 535. [Google Scholar] [CrossRef] [Green Version]
- Maes, M.E.; Schlamp, C.L.; Nickells, R.W. BAX to basics: How the BCL2 gene family controls the death of retinal ganglion cells. Prog. Retin. Eye Res. 2017, 57, 1–25. [Google Scholar] [CrossRef] [Green Version]
- Dai, Y.; Hu, X.; Sun, X. Overexpression of parkin protects retinal ganglion cells in experimental glaucoma. Cell Death Dis. 2018, 9, 88. [Google Scholar] [CrossRef] [Green Version]
- Bach, M.; Cuno, A.K.; Hoffmann, M.B. Retinal conduction speed analysis reveals different origins of the P50 and N95 components of the (multifocal) pattern electroretinogram. Exp. Eye Res. 2018, 169, 48–53. [Google Scholar] [CrossRef] [PubMed]
- Cvenkel, B.; Sustar, M.; Perovšek, D. Ganglion cell loss in early glaucoma, as assessed by photopic negative response, pattern electroretinogram, and spectral-domain optical coherence tomography. Doc. Ophthalmol. 2017, 135, 17–28. [Google Scholar] [CrossRef]
- Wilsey, L.; Gowrisankaran, S.; Cull, G.; Hardin, C.; Burgoyne, C.F.; Fortune, B. Comparing three different modes of electroretinography in experimental glaucoma: Diagnostic performance and correlation to structure. Doc. Ophthalmol. 2017, 134, 111–128. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sah, A.K.; Suresh, P.K. Medical management of glaucoma: Focus on ophthalmologic drug delivery systems of timolol maleate. Artif. Cells Nanomed. Biotechnol. 2017, 45, 448–459. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, K.; Xu, L.; Zhang, H.; Zhang, S.; Pu, M.; Jonas, J.B. Effect of brimonidine on retinal ganglion cell survival in an optic nerve crush model. Am. J. Ophthalmol. 2009, 147, 326–331. [Google Scholar] [CrossRef]
- Semba, K.; Namekata, K.; Kimura, A.; Harada, C.; Mitamura, Y.; Harada, T. Brimonidine prevents neurodegeneration in a mouse model of normal tension glaucoma. Cell Death Dis. 2014, 5, e1341. [Google Scholar] [CrossRef] [Green Version]
- Wheeler, L.A.; WoldeMussie, E. Alpha-2 adrenergic receptor agonists are neuroprotective in experimental models of glaucoma. Eur. J. Ophthalmol. 2001, 11 (Suppl. S2), S30–S35. [Google Scholar] [CrossRef]
- Takamura, Y.; Tomomatsu, T.; Matsumura, T.; Takihara, Y.; Kozai, S.; Arimura, S.; Yokota, S.; Inatani, M. Vitreous and aqueous concentrations of brimonidine following topical application of brimonidine tartrate 0.1% ophthalmic solution in humans. J. Ocul. Pharmacol. Ther. 2015, 31, 282–285. [Google Scholar] [CrossRef]
- Del Amo, E.M.; Urtti, A. Current and future ophthalmic drug delivery systems. A shift to the posterior segment. Drug Discov. Today 2008, 13, 135–143. [Google Scholar] [CrossRef] [PubMed]
- Acheampong, A.A.; Shackleton, M.; John, B.; Burke, J.; Wheeler, L.; Tang-Liu, D. Distribution of brimonidine into anterior and posterior tissues of monkey, rabbit, and rat eyes. Drug Metab. Dispos. 2002, 30, 421–429. [Google Scholar] [CrossRef] [Green Version]
- Liu, J.; Clough, S.J.; Hutchinson, A.J.; Adamah-Biassi, E.B.; Popovska-Gorevski, M.; Dubocovich, M.L. MT1 and MT2 melatonin receptors: A therapeutic perspective. Annu. Rev. Pharmacol. Toxicol. 2016, 56, 361–383. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dehdashtian, E.; Mehrzadi, S.; Yousefi, B.; Hosseinzadeh, A.; Reiter, R.J.; Safa, M.; Ghaznavi, H.; Naseripour, M. Diabetic retinopathy pathogenesis and the ameliorating effects of melatonin; involvement of autophagy, inflammation and oxidative stress. Life Sci. 2018, 193, 20–33. [Google Scholar] [CrossRef] [PubMed]
- Domenici, L.; Origlia, N.; Falsini, B.; Cerri, E.; Barloscio, D.; Fabiani, C.; Sansò, M.; Giovannini, L. Rescue of retinal function by BDNF in a mouse model of glaucoma. PLoS ONE. 2014, 9, e115579. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Parisi, V.; Oddone, F.; Roberti, G.; Tanga, L.; Carnevale, C.; Ziccardi, L.; Manni, G. Enhancement of retinal function and of neural conduction along the visual pathway induced by treatment with citicoline eye drops in liposomal formulation in open angle glaucoma: A pilot electrofunctional study. Adv. Ther. 2019, 36, 987–996. [Google Scholar] [CrossRef] [PubMed]
- Parisi, V.; Centofanti, M.; Gandolfi, S.; Marangoni, D.; Rossetti, L.; Tanga, L.; Tardini, M.; Traina, S.; Ungaro, N.; Vetrugno, M.; et al. Effects of coenzyme Q10 in conjunction with vitamin E on retinal-evoked and cortical-evoked responses in patients with open-angle glaucoma. J. Glaucoma 2014, 23, 391–404. [Google Scholar] [CrossRef] [PubMed]
- Cazevieille, C.; Osborne, N.N. Retinal neurones containing kainate receptors are influenced by exogenous kainate and ischaemia while neurones lacking these receptors are not—melatonin counteracts the effects of ischaemia and kainate. Brain Res. 1997, 755, 91–100. [Google Scholar] [CrossRef]
- WoldeMussie, E.; Ruiz, G.; Wijono, M.; Wheeler, L.A. Neuroprotection of retinal ganglion cells by brimonidine in rats with laser-induced chronic ocular hypertension. Investig. Ophthalmol. Vis. Sci. 2001, 42, 2849–2855. [Google Scholar]
- Bertino, B.; Blanchet-Réthoré, S.; Thibaut de Ménonville, S.; Reynier, P.; Méhul, B.; Bogouch, A.; Gamboa, B.; Dugaret, A.S.; Zugaj, D.; Petit, L.; et al. Brimonidine displays anti-inflammatory properties in the skin through the modulation of the vascular barrier function. Exp. Dermatol. 2018, 27, 1378–1387. [Google Scholar] [CrossRef]
- Alonso, R.S.; Solari, H.P.; de França Damasceno, E.; Burnier, M.N.N., Jr.; Ventura, M.P. The chemotactic properties of various topical brimonidine tartrate ophthalmic preparations. BMC Pharmacol. Toxicol. 2020, 21, 24. [Google Scholar] [CrossRef] [Green Version]
- Beltz, J.; Zamir, E. Brimonidine induced anterior uveitis. Ocul. Immunol. Inflamm. 2016, 24, 128–133. [Google Scholar] [CrossRef]
- Levkovitch-Verbin, H.; Harris-Cerruti, C.; Groner, Y.; Wheeler, L.A.; Schwartz, M.; Yoles, E. RGC death in mice after optic nerve crush injury: Oxidative stress and neuroprotection. Investig. Ophthalmol. Vis. Sci. 2000, 41, 4169–4174. [Google Scholar] [PubMed]
- Yoles, E.; Wheeler, L.A.; Schwartz, M. Alpha2-adrenoreceptor agonists are neuroprotective in a rat model of optic nerve degeneration. Investig. Ophthalmol. Vis. Sci. 1999, 40, 65–73. [Google Scholar] [PubMed]
- Zhou, X.; Zhang, T.; Wu, J. Brimonidine enhances inhibitory postsynaptic activity of OFF- and ON-type retinal ganglion cells in a Wistar rat chronic glaucoma model. Exp. Eye Res. 2019, 189, 107833. [Google Scholar] [CrossRef] [PubMed]
- Dong, C.J.; Guo, Y.; Agey, P.; Wheeler, L.; Hare, W.A. Alpha2 adrenergic modulation of NMDA receptor function as a major mechanism of RGC protection in experimental glaucoma and retinal excitotoxicity. Investig. Ophthalmol. Vis. Sci. 2008, 49, 4515–4522. [Google Scholar] [CrossRef]
- Lai, R.K.; Chun, T.; Hasson, D.; Lee, S.; Mehrbod, F.; Wheeler, L. Alpha-2 adrenoceptor agonist protects retinal function after acute retinal ischemic injury in the rat. Vis. Neurosci. 2002, 19, 175–185. [Google Scholar] [CrossRef]
- Gao, H.; Qiao, X.; Hefti, F.; Hollyfield, J.G.; Knusel, B. Elevated mRNA expression of brain-derived neurotrophic factor in retinal ganglion cell layer after optic nerve injury. Investig. Ophthalmol. Vis. Sci. 1997, 38, 1840–1847. [Google Scholar]
- Nizari, S.; Guo, L.; Davis, B.M.; Normando, E.M.; Galvao, J.; Turner, L.A.; Bizrah, M.; Dehabadi, M.; Tian, K.; Cordeiro, M.F. Non-amyloidogenic effects of α2 adrenergic agonists: Implications for brimonidine-mediated neuroprotection. Cell Death Dis. 2016, 7, e2514. [Google Scholar] [CrossRef] [Green Version]
- Adornetto, A.; Russo, R.; Parisi, V. Neuroinflammation as a target for glaucoma therapy. Neural Regen. Res. 2019, 14, 391–394. [Google Scholar] [CrossRef]
- Zhang, X.; Zhang, R.; Chen, J.; Wu, J. Neuroprotective effects of DAAO are mediated via the ERK1/2 signaling pathway in a glaucomatous animal model. Exp. Eye Res. 2020, 190, 107892. [Google Scholar] [CrossRef]
- Jiang, T.; Chang, Q.; Zhao, Z.; Yan, S.; Wang, L.; Cai, J.; Xu, G. Melatonin-mediated cytoprotection against hyperglycemic injury in Müller cells. PLoS ONE. 2012, 7, e50661. [Google Scholar] [CrossRef] [Green Version]
- Aranda, M.L.; González Fleitas, M.F.; De Laurentiis, A.; Keller Sarmiento, M.I.; Chianelli, M.; Sande, P.H.; Dorfman, D.; Rosenstein, R.E. Neuroprotective effect of melatonin in experimental optic neuritis in rats. J. Pineal Res. 2016, 60, 360–372. [Google Scholar] [CrossRef] [PubMed]
- Kaur, C.; Sivakumar, V.; Robinson, R.; Foulds, W.S.; Luu, C.D.; Ling, E.A. Neuroprotective effect of melatonin against hypoxia-induced retinal ganglion cell death in neonatal rats. J. Pineal Res. 2013, 54, 190–206. [Google Scholar] [CrossRef] [PubMed]
- Xie, M.; Hu, A.; Luo, Y.; Sun, W.; Hu, X.; Tang, S. Interleukin-4 and melatonin ameliorate high glucose and interleukin-1β stimulated inflammatory reaction in human retinal endothelial cells and retinal pigment epithelial cells. Mol. Vis. 2014, 20, 921–928. [Google Scholar] [PubMed]
- Yan, G.; Yu, L.; Jiang, S.; Zhu, J. Melatonin antagonizes oxidative stress-induced mitochondrial dysfunction in retinal pigmented epithelium cells via melatonin receptor 1 (MT1). J. Toxicol. Sci. 2018, 43, 659–669. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rosen, R.B.; Hu, D.N.; Chen, M.; McCormick, S.A.; Walsh, J.; Roberts, J.E. Effects of melatonin and its receptor antagonist on retinal pigment epithelial cells against hydrogen peroxide damage. Mol. Vis. 2012, 18, 1640–1648. [Google Scholar]
- Mandal, A.; Cholkar, K.; Khurana, V.; Shah, A.; Agrahari, V.; Bisht, R.; Pal, D.; Mitra, A.K. Topical formulation of self-assembled antiviral prodrug nanomicelles for targeted retinal delivery. Mol. Pharm. 2017, 14, 2056–2069. [Google Scholar] [CrossRef]
- Mukherjee, R.; Banerjee, S.; Joshi, N.; Singh, P.K.; Baxi, D.; Ramachandran, A.V. A combination of melatonin and alpha lipoic acid has greater cardioprotective effect than either of them singly against cadmium-induced oxidative damage. Cardiovasc. Toxicol. 2011, 11, 78–88. [Google Scholar] [CrossRef]
- Jiménez-Ortega, V.; Barquilla, P.C.; Pagano, E.S.; Fernández-Mateos, P.; Esquifino, A.I.; Cardinali, D.P. Melatonin supplementation decreases prolactin synthesis and release in rat adenohypophysis: Correlation with anterior pituitary redox state and circadian clock mechanisms. Chronobiol. Int. 2012, 29, 1021–1035. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Zhang, D.; Liu, M.; Zhao, H.; Du, A.; Meng, L.; Liu, H. LC-MS/MS method for the determination of agomelatine in human plasma and its application to a pharmacokinetic study. Biomed. Chromatogr. 2014, 28, 218–222. [Google Scholar] [CrossRef]
- Alarma-Estrany, P.; Guzman-Aranguez, A.; Huete, F.; Peral, A.; Plourde, R., Jr.; Pelaez, T.; Yerxa, B.; Pintor, J. Design of novel melatonin analogs for the reduction of intraocular pressure in normotensive rabbits. J. Pharmacol. Exp. Ther. 2011, 337, 703–709. [Google Scholar] [CrossRef] [Green Version]
- Porciatti, V. Electrophysiological assessment of retinal ganglion cell function. Exp. Eye Res. 2015, 141, 164–170. [Google Scholar] [CrossRef] [PubMed] [Green Version]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Melatonin | Agomelatine | |||||||
|---|---|---|---|---|---|---|---|---|
| Animal | Sample Type | Weight (mg) | Concentration (ng/mg) | Mean | SEM | Concentration (ng/mg) | Mean | SEM |
| 1 | Whole eye | 151 | 7.83 | 7.44 | 0.85 | 10.89 | 9.94 | 0.68 |
| 2 | Whole eye | 165 | 10.21 | 10.64 | ||||
| 3 | Whole eye | 176 | 7.87 | 11.35 | ||||
| 4 | Whole eye | 160 | 5.64 | 9.16 | ||||
| 5 | Whole eye | 150 | 5.67 | 7.67 | ||||
| 1 to 5 | Pool of retinas | 115 | 0.004 | - | - | 0.096 | ||
| 1 | Vitreous | 11 | 0.007 | 0.010 | 0.001 | 0.071 | 0.081 | 0.005 |
| 2 | Vitreous | 12 | 0.008 | 0.073 | ||||
| 3 | Vitreous | 10 | 0.013 | 0.099 | ||||
| 4 | Vitreous | 13 | 0.010 | 0.077 | ||||
| 5 | Vitreous | 11 | 0.014 | 0.084 | ||||
| RGC density (% of control) | |||
|---|---|---|---|
| Central | Middle | Peripheral | |
| Control | 100 ± 4 | 100 ± 5 | 100 ± 7 |
| MCE | 70 ± 6 * | 71 ± 4 * | 71 ± 6 * |
| + Vehicle 1 | 72 ± 5 * | 72 ± 3 * | 70 ± 4 * |
| + Vehicle 2 | 73 ± 7 * | 72 ± 4 * | 71 ± 6 * |
| + Timolol | 78 ± 4 *,§ | 83 ± 3 *,§ | 81 ± 4 *,§ |
| + Brimonidine | 88 ± 3 § | 91 ± 6 § | 91 ± 3 § |
| + MelAgo | 92 ± 5 § | 94 ± 5 § | 94 ± 3 § |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dal Monte, M.; Cammalleri, M.; Amato, R.; Pezzino, S.; Corsaro, R.; Bagnoli, P.; Rusciano, D. A Topical Formulation of Melatoninergic Compounds Exerts Strong Hypotensive and Neuroprotective Effects in a Rat Model of Hypertensive Glaucoma. Int. J. Mol. Sci. 2020, 21, 9267. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21239267
Dal Monte M, Cammalleri M, Amato R, Pezzino S, Corsaro R, Bagnoli P, Rusciano D. A Topical Formulation of Melatoninergic Compounds Exerts Strong Hypotensive and Neuroprotective Effects in a Rat Model of Hypertensive Glaucoma. International Journal of Molecular Sciences. 2020; 21(23):9267. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21239267
Chicago/Turabian StyleDal Monte, Massimo, Maurizio Cammalleri, Rosario Amato, Salvatore Pezzino, Roberta Corsaro, Paola Bagnoli, and Dario Rusciano. 2020. "A Topical Formulation of Melatoninergic Compounds Exerts Strong Hypotensive and Neuroprotective Effects in a Rat Model of Hypertensive Glaucoma" International Journal of Molecular Sciences 21, no. 23: 9267. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21239267