Linking ABCC6 Deficiency in Primary Human Dermal Fibroblasts of PXE Patients to p21-Mediated Premature Cellular Senescence and the Development of a Proinflammatory Secretory Phenotype

 , ,
, ,
Abstract
:1. Introduction
2. Results
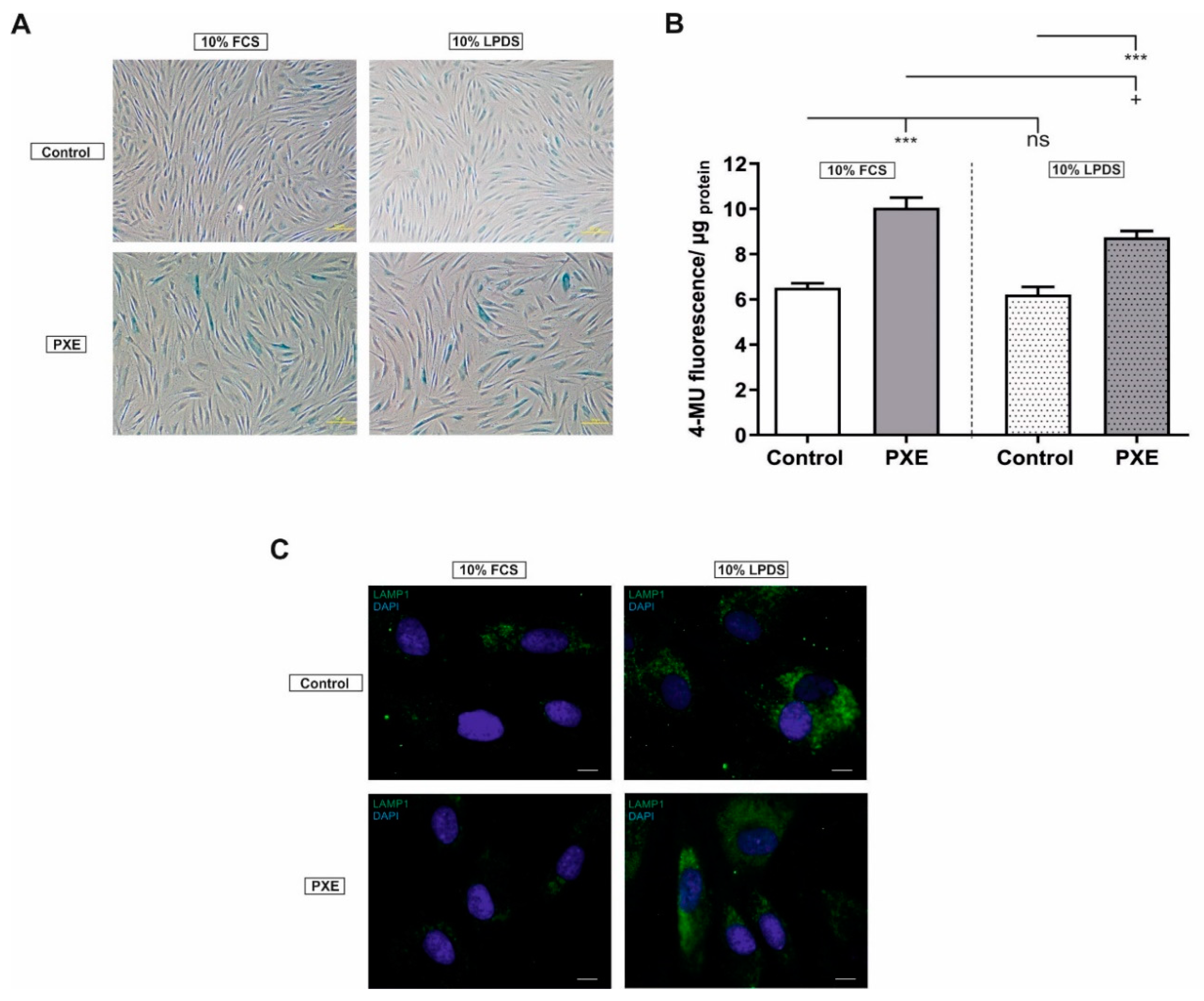
2.1. Altered Cell Morphology and Increased Senescence-Associated-β-Gal (SA-β-Gal) Activity
2.2. Aberrant Lamin B1 (LMNB1) Gene, but No Changes in Protein Expression
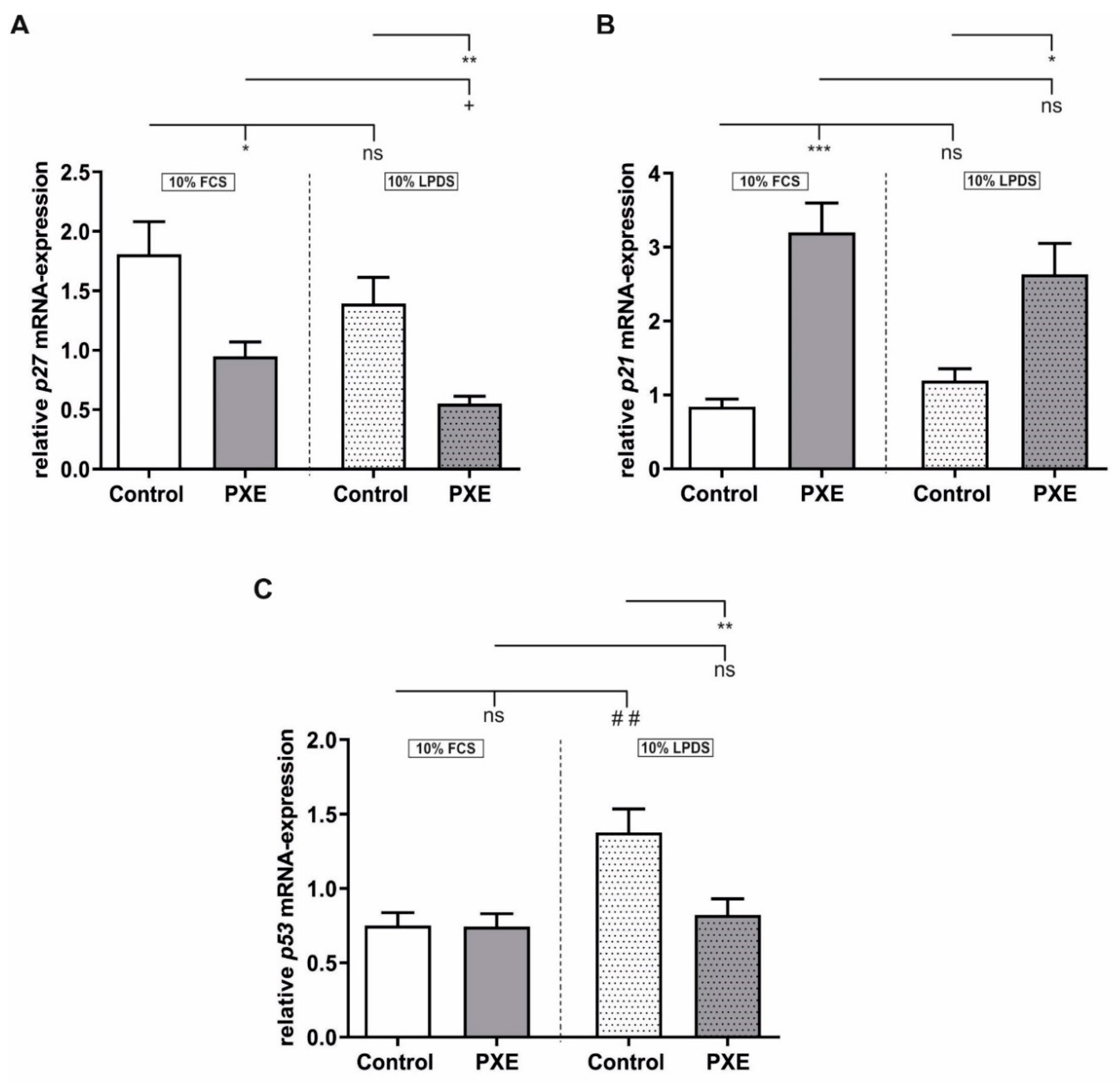
2.3. Alteration in Gene Expression of the Cyclin-Dependent Kinase Inhibitors (CDKI) p21 and p27
2.4. Strong Increase in Interleukin 6 (IL6) Gene and Protein Expression
3. Discussion
4. Materials and Methods
4.1. Experimental Design
4.2. Cell Culture
4.3. Delipidation of FCS
4.4. Nucleic Acid Isolation
4.5. Bicinchoninic Acid Assay
4.6. Gene Expression Analysis
4.7. Immunofluorescence Microscopy
4.8. Qualitative Senescence Assay
4.9. Quantitative Senescence Assay
4.10. Immunoassays for Evaluation of SASP Factors in Cell Culture Supernatants
4.11. Cell Cycle Antibody Array
4.12. Statistical Analysis
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| ABCC6 | ATP-binding cassette sub-family C member 6 |
| CDK | Cyclin-dependent kinase |
| CDKI | Cyclin-dependent kinase inhibitor |
| FCS | Fetal calf serum |
| HGPS | Hutchinson–Gilford progeria syndrome |
| ICAM1 | Intercellular adhesion molecule 1 |
| IL6 | Interleukin 6 |
| JAK | Janus Kinase |
| LAMP1 | Lysosomal-associated membrane protein 1 |
| LMNB1 | Lamin B1 |
| LPDS | Lipoprotein-deficient fetal calf serum |
| MCP1 | Monocyte chemoattractant protein-1 |
| MMP | Matrix metalloproteinase |
| NHDF | Normal human dermal fibroblasts |
| PPi | Pyrophosphate |
| pRB | Retinoblastoma protein |
| PXE | Pseudoxanthoma elasticum |
| SA-β-Gal | Senescence-associated β-galactosidase |
| SASP | Senescence-associated secretory phenotype |
| STAT | Signal transducer and activator of transcription |
References
- Germain, D.P. Pseudoxanthoma elasticum. Orphanet J. Rare Dis. 2017, 12, 85. [Google Scholar] [CrossRef] [PubMed]
- Le Saux, O.; Urban, Z.; Tschuch, C.; Csiszar, K.; Bacchelli, B.; Quaglino, D.; Pasquali-Ronchetti, I.; Pope, F.M.; Richards, A.; Terry, S.; et al. Mutations in a gene encoding an ABC transporter cause pseudoxanthoma elasticum. Nat. Genet. 2000, 25, 223–227. [Google Scholar] [CrossRef] [PubMed]
- Legrand, A.; Cornez, L.; Samkari, W.; Mazzella, J.-M.; Venisse, A.; Boccio, V.; Auribault, K.; Keren, B.; Benistan, K.; Germain, D.P.; et al. Mutation spectrum in the ABCC6 gene and genotype–phenotype correlations in a French cohort with pseudoxanthoma elasticum. Genet. Med. 2017, 19, 909–917. [Google Scholar] [CrossRef] [PubMed]
- Ronchetti, I.; Boraldi, F.; Annovi, G.; Cianciulli, P.; Quaglino, D. Fibroblast involvement in soft connective tissue calcification. Front. Genet. 2013, 4. [Google Scholar] [CrossRef] [Green Version]
- Boraldi, F.; Annovi, G.; Vermeer, C.; Schurgers, L.J.; Trenti, T.; Tiozzo, R.; Guerra, D.; Quaglino, D. Matrix gla protein and alkaline phosphatase are differently modulated in human dermal fibroblasts from PXE patients and controls. J. Investig. Dermatol. 2013, 133, 946–954. [Google Scholar] [CrossRef] [Green Version]
- Boraldi, F.; Annovi, G.; Bartolomeo, A.; Quaglino, D. Fibroblasts from patients affected by Pseudoxanthoma elasticum exhibit an altered PPi metabolism and are more responsive to pro-calcifying stimuli. J. Dermatol. Sci. 2014, 74, 72–80. [Google Scholar] [CrossRef] [Green Version]
- Villa-Bellosta, R.; Rivera-Torres, J.; Osorio, F.G.; Acín-Pérez, R.; Enriquez, J.A.; López-Otín, C.; Andrés, V. Defective extracellular pyrophosphate metabolism promotes vascular calcification in a mouse model of Hutchinson-Gilford progeria syndrome that is ameliorated on pyrophosphate treatment. Circulation 2013, 127, 2442–2451. [Google Scholar] [CrossRef] [Green Version]
- Guo, H.; Li, Q.; Chou, D.W.; Uitto, J. Atorvastatin counteracts aberrant soft tissue mineralization in a mouse model of pseudoxanthoma elasticum (Abcc6 −/−). J. Mol. Med. 2013, 91, 1177–1184. [Google Scholar] [CrossRef] [Green Version]
- Li, Q.; Sundberg, J.P.; Levine, M.A.; Terry, S.F.; Uitto, J. The effects of bisphosphonates on ectopic soft tissue mineralization caused by mutations in the ABCC6 gene. Cell Cycle 2015, 14, 1082–1089. [Google Scholar] [CrossRef] [Green Version]
- Luft, F.C. Pseudoxanthoma elasticum and statin prophylaxis. J. Mol. Med. 2013, 91, 1129–1130. [Google Scholar] [CrossRef] [Green Version]
- Wheaton, K.; Campuzano, D.; Ma, W.; Sheinis, M.; Ho, B.; Brown, G.W.; Benchimol, S. Progerin-Induced Replication Stress Facilitates Premature Senescence in Hutchinson-Gilford Progeria Syndrome. Mol. Cell. Biol. 2017, 37. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- López-Otín, C.; Blasco, M.A.; Partridge, L.; Serrano, M.; Kroemer, G. The Hallmarks of Aging. Cell 2013, 153, 1194–1217. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Demaria, M.; Ohtani, N.; Youssef, S.A.; Rodier, F.; Toussaint, W.; Mitchell, J.R.; Laberge, R.-M.; Vijg, J.; Van Steeg, H.; Dollé, M.E.T.; et al. An Essential Role for Senescent Cells in Optimal Wound Healing through Secretion of PDGF-AA. Dev. Cell 2014, 31, 722–733. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Faust, I.; Donhauser, E.; Fischer, B.; Ibold, B.; Kuhn, J.; Knabbe, C.; Hendig, D. Characterization of dermal myofibroblast differentiation in pseudoxanthoma elasticum. Exp. Cell Res. 2017, 360, 153–162. [Google Scholar] [CrossRef]
- Diekmann, U.; Zarbock, R.; Hendig, D.; Szliska, C.; Kleesiek, K.; Götting, C. Elevated circulating levels of matrix metalloproteinases MMP-2 and MMP-9 in pseudoxanthoma elasticum patients. J. Mol. Med. 2009, 87, 965–970. [Google Scholar] [CrossRef]
- Chau, K.Y.; Sivaprasad, S.; Patel, N.; Donaldson, T.A.; Luthert, P.J.; Chong, N.V. Plasma levels of matrix metalloproteinase-2 and -9 (MMP-2 and MMP-9) in age-related macular degeneration. Eye 2008, 22, 855–859. [Google Scholar] [CrossRef]
- Gliem, M.; Zaeytijd, J.D.; Finger, R.P.; Holz, F.G.; Leroy, B.P.; Charbel Issa, P. An update on the ocular phenotype in patients with pseudoxanthoma elasticum. Front. Genet. 2013, 4, 14. [Google Scholar] [CrossRef] [Green Version]
- Miglionico, R.; Ostuni, A.; Armentano, M.F.; Milella, L.; Crescenzi, E.; Carmosino, M.; Bisaccia, F. ABCC6 knockdown in HepG2 cells induces a senescent-like cell phenotype. Cell. Mol. Biol. Lett. 2017, 22, 7. [Google Scholar] [CrossRef] [Green Version]
- Finger, R.P.; Charbel Issa, P.; Ladewig, M.S.; Götting, C.; Szliska, C.; Scholl, H.P.N.; Holz, F.G. Pseudoxanthoma elasticum: Genetics, clinical manifestations and therapeutic approaches. Surv. Ophthalmol. 2009, 54, 272–285. [Google Scholar] [CrossRef]
- Kuzaj, P.; Kuhn, J.; Dabisch-Ruthe, M.; Faust, I.; Götting, C.; Knabbe, C.; Hendig, D. ABCC6-A new player in cellular cholesterol and lipoprotein metabolism? Lipids Health Dis. 2014, 13, 118. [Google Scholar] [CrossRef] [Green Version]
- Freund, A.; Laberge, R.-M.; Demaria, M.; Campisi, J. Lamin B1 loss is a senescence-associated biomarker. Mol. Biol. Cell 2012, 23, 2066–2075. [Google Scholar] [CrossRef] [PubMed]
- Shimi, T.; Butin-Israeli, V.; Adam, S.A.; Hamanaka, R.B.; Goldman, A.E.; Lucas, C.A.; Shumaker, D.K.; Kosak, S.T.; Chandel, N.S.; Goldman, R.D. The role of nuclear lamin B1 in cell proliferation and senescence. Genes Dev. 2011, 25, 2579–2593. [Google Scholar] [CrossRef] [Green Version]
- Wang, A.S.; Ong, P.F.; Chojnowski, A.; Clavel, C.; Dreesen, O. Loss of lamin B1 is a biomarker to quantify cellular senescence in photoaged skin. Sci. Rep. 2017, 7, 15678. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Giacinti, C.; Giordano, A. RB and cell cycle progression. Oncogene 2006, 25, 5220–5227. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weinberg, R.A. The retinoblastoma protein and cell cycle control. Cell 1995, 81, 323–330. [Google Scholar] [CrossRef] [Green Version]
- Chen, Q.M.; Tu, V.C.; Catania, J.; Burton, M.; Toussaint, O.; Dilley, T. Involvement of Rb family proteins, focal adhesion proteins and protein synthesis in senescent morphogenesis induced by hydrogen peroxide. J. Cell. Sci. 2000, 113, 4087–4097. [Google Scholar]
- Hayflick, L.; Moorhead, P.S. The serial cultivation of human diploid cell strains. Exp. Cell Res. 1961, 25, 585–621. [Google Scholar] [CrossRef]
- Dimri, G.P.; Lee, X.; Basile, G.; Acosta, M.; Scott, G.; Roskelley, C.; Medrano, E.E.; Linskens, M.; Rubelj, I.; Pereira-Smith, O. A biomarker that identifies senescent human cells in culture and in aging skin in vivo. Proc. Natl. Acad. Sci. USA 1995, 92, 9363–9367. [Google Scholar] [CrossRef] [Green Version]
- Kurz, D.J.; Decary, S.; Hong, Y.; Erusalimsky, J.D. Senescence-associated (beta)-galactosidase reflects an increase in lysosomal mass during replicative ageing of human endothelial cells. J. Cell. Sci. 2000, 113, 3613–3622. [Google Scholar]
- Lee, B.Y.; Han, J.A.; Im, J.S.; Morrone, A.; Johung, K.; Goodwin, E.C.; Kleijer, W.J.; DiMaio, D.; Hwang, E.S. Senescence-associated β-galactosidase is lysosomal β-galactosidase. Aging Cell 2006, 5, 187–195. [Google Scholar] [CrossRef]
- Jung, H.-J.; Nobumori, C.; Goulbourne, C.N.; Tu, Y.; Lee, J.M.; Tatar, A.; Wu, D.; Yoshinaga, Y.; de Jong, P.J.; Coffinier, C.; et al. Farnesylation of lamin B1 is important for retention of nuclear chromatin during neuronal migration. Proc. Natl. Acad. Sci. USA 2013, 110, E1923–E1932. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gartel, A.L.; Tyner, A.L. Transcriptional regulation of the p21((WAF1/CIP1)) gene. Exp. Cell Res. 1999, 246, 280–289. [Google Scholar] [CrossRef] [PubMed]
- Das, U.N. Statins and the prevention of dementia. Can. Med. Assoc. J. 2001, 165, 908–909. [Google Scholar]
- Magee, T.; Marshall, C. New Insights into the Interaction of Ras with the Plasma Membrane. Cell 1999, 98, 9–12. [Google Scholar] [CrossRef] [Green Version]
- Molnár, G.; Dagher, M.-C.; Geiszt, M.; Settleman, J.; Ligeti, E. Role of Prenylation in the Interaction of Rho-Family Small GTPases with GTPase Activating Proteins. Biochemistry 2001, 40, 10542–10549. [Google Scholar] [CrossRef]
- Omoigui, S. The Interleukin-6 inflammation pathway from cholesterol to aging-Role of statins, bisphosphonates and plant polyphenols in aging and age-related diseases. Immun. Ageing 2007, 4, 1. [Google Scholar] [CrossRef] [Green Version]
- Pereira-Leal, J.B.; Hume, A.N.; Seabra, M.C. Prenylation of Rab GTPases: Molecular mechanisms and involvement in genetic disease. FEBS Lett. 2001, 498, 197–200. [Google Scholar] [CrossRef]
- Dulic, V. Senescence Regulation by mTOR. In Cell Senescence; Galluzzi, L., Vitale, I., Kepp, O., Kroemer, G., Eds.; Humana Press: Totowa, NJ, USA, 2013; Volume 965, pp. 15–35. [Google Scholar]
- Polyak, K.; Kato, J.Y.; Solomon, M.J.; Sherr, C.J.; Massague, J.; Roberts, J.M.; Koff, A. p27Kip1, a cyclin-Cdk inhibitor, links transforming growth factor-Beta and contact inhibition to cell cycle arrest. Genes Dev. 1994, 8, 9–22. [Google Scholar] [CrossRef] [Green Version]
- Tiemann, J.; Wagner, T.; Vanakker, O.M.; van Gils, M.; Cabrera, J.-L.B.; Ibold, B.; Faust, I.; Knabbe, C.; Hendig, D. Cellular and Molecular Biomarkers Indicate Premature Aging in Pseudoxanthoma Elasticum Patients. Aging Dis. 2020, 11, 536. [Google Scholar] [CrossRef]
- Bellido, T.; O’Brien, C.A.; Roberson, P.K.; Manolagas, S.C. Transcriptional activation of the p21(WAF1,CIP1,SDI1) gene by interleukin-6 type cytokines. A prerequisite for their pro-differentiating and anti-apoptotic effects on human osteoblastic cells. J. Biol. Chem. 1998, 273, 21137–21144. [Google Scholar] [CrossRef] [Green Version]
- Choi, J.M.; Rotimi, O.O.; O’Carroll, S.J.; Nicholson, L.F.B. IL-6 stimulates a concentration-dependent increase in MCP-1 in immortalised human brain endothelial cells. F1000Research 2016, 5, 270. [Google Scholar] [CrossRef] [Green Version]
- Marino, M.; Scuderi, F.; Provenzano, C.; Scheller, J.; Rose-John, S.; Bartoccioni, E. IL-6 regulates MCP-1, ICAM-1 and IL-6 expression in human myoblasts. J. Neuroimmunol. 2008, 196, 41–48. [Google Scholar] [CrossRef]
- Spörri, B.; Müller, K.M.; Wiesmann, U.; Bickel, M. Soluble IL-6 receptor induces calcium flux and selectively modulates chemokine expression in human dermal fibroblasts. Int. Immunol. 1999, 11, 1053–1058. [Google Scholar] [CrossRef] [Green Version]
- Lee, W.; Jo, S.; Lee, M.; Won, C.; Lee, M.; Choi, J.; Chang, S. The Effect of MCP-1/CCR2 on the Proliferation and Senescence of Epidermal Constituent Cells in Solar Lentigo. Int. J. Mol. Sci. 2016, 17, 948. [Google Scholar] [CrossRef] [Green Version]
- Marino, M.; Scuderi, F.; Mazzarelli, P.; Mannella, F.; Provenzano, C.; Bartoccioni, E. Constitutive and cytokine-induced expression of MHC and intercellular adhesion molecule-1 (ICAM-1) on human myoblasts. J. Neuroimmunol. 2001, 116, 94–101. [Google Scholar] [CrossRef]
- Shelton, D.N.; Chang, E.; Whittier, P.S.; Choi, D.; Funk, W.D. Microarray analysis of replicative senescence. Curr. Biol. 1999, 9, 939–945. [Google Scholar] [CrossRef] [Green Version]
- Bauer, C.; le Saux, O.; Pomozi, V.; Aherrahrou, R.; Kriesen, R.; Stölting, S.; Liebers, A.; Kessler, T.; Schunkert, H.; Erdmann, J.; et al. Etidronate prevents dystrophic cardiac calcification by inhibiting macrophage aggregation. Sci. Rep. 2018, 8, 5812. [Google Scholar] [CrossRef] [Green Version]
- Hendig, D.; Adam, A.; Zarbock, R.; Szliska, C.; Kleesiek, K.; Götting, C. Elevated serum levels of intercellular adhesion molecule ICAM-1 in Pseudoxanthoma elasticum. Clin. Chim. Acta 2008, 394, 54–58. [Google Scholar] [CrossRef] [PubMed]
- Gorgoulis, V.G.; Pratsinis, H.; Zacharatos, P.; Demoliou, C.; Sigala, F.; Asimacopoulos, P.J.; Papavassiliou, A.G.; Kletsas, D. p53-dependent ICAM-1 overexpression in senescent human cells identified in atherosclerotic lesions. Lab. Investig. 2005, 85, 502–511. [Google Scholar] [CrossRef] [PubMed]
- Milan-Mattos, J.C.; Anibal, F.F.; Perseguini, N.M.; Minatel, V.; Rehder-Santos, P.; Castro, C.A.; Vasilceac, F.A.; Mattiello, S.M.; Faccioli, L.H.; Catai, A.M. Effects of natural aging and gender on pro-inflammatory markers. Braz. J. Med. Biol. Res. 2019, 52, e8392. [Google Scholar] [CrossRef] [Green Version]
- Puzianowska-Kuźnicka, M.; Owczarz, M.; Wieczorowska-Tobis, K.; Nadrowski, P.; Chudek, J.; Slusarczyk, P.; Skalska, A.; Jonas, M.; Franek, E.; Mossakowska, M. Interleukin-6 and C-reactive protein, successful aging, and mortality: The PolSenior study. Immun. Ageing 2016, 13, 21. [Google Scholar] [CrossRef] [Green Version]
- Hendig, D.; Langmann, T.; Kocken, S.; Zarbock, R.; Szliska, C.; Schmitz, G.; Kleesiek, K.; Götting, C. Gene expression profiling of ABC transporters in dermal fibroblasts of pseudoxanthoma elasticum patients identifies new candidates involved in PXE pathogenesis. Lab. Investig. 2008, 88, 1303–1315. [Google Scholar] [CrossRef] [Green Version]
- Gibson, K.M.; Hoffmann, G.; Schwall, A.; Broock, R.L.; Aramaki, S.; Sweetman, L.; Nyhan, W.L.; Brandt, I.K.; Wappner, R.S.; Lehnert, W. 3-Hydroxy-3-methylglutaryl coenzyme A reductase activity in cultured fibroblasts from patients with mevalonate kinase deficiency: Differential response to lipid supplied by fetal bovine serum in tissue culture medium. J. Lipid Res. 1990, 31, 515–521. [Google Scholar] [PubMed]
- Gary, R.K.; Kindell, S.M. Quantitative assay of senescence-associated beta-galactosidase activity in mammalian cell extracts. Anal. Biochem. 2005, 343, 329–334. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Array Position | Protein | Mean Signal Intensity of Spots | Ratio PXE/Control | |
|---|---|---|---|---|
| Control | PXE | |||
| Block 1, No. 5 | Cyclin B1 | 585 | 492 | 0.84 |
| Block 1, No. 6 | Cyclin C | 284 | 260 | 0.92 |
| Block 1, No. 7 | Cyclin E | 2237 | 1715 | 0.77 |
| Block 1, No. 8 | Cdk3 | 964 | 887 | 0.92 |
| Block 1, No. 9 | Cdk8 | 560 | 514 | 0.92 |
| Block 1, No. 10 | CDC37 | 284 | 262 | 0.92 |
| Block 1, No. 18 | APC11 | 401 | 380 | 0.95 |
| Block 2 No. 8 | Cdk1/p34cdc2 | 901 | 772 | 0.86 |
| Block 2, No. 9 | Cyclin D1 | 259 | 228 | 0.88 |
| Block 2, No. 14 | p21WAF1 | 240 | 262 | 1.09 |
| Block 2, No. 17 | E2F-2 | 302 | 335 | 1.11 |
| Block 3, No. 10 | p130cas | 284 | 332 | 1.17 |
| Block 4, No. 5 | Ki67 | 304 | 257 | 0.85 |
| Block 4, No. 6 | Chk1 | 518 | 424 | 0.82 |
| Block 4, No. 7 | 14.3.3, Pan | 682 | 866 | 1.27 |
| Block 4, No. 8 | Cullin-3 (CUL-3) | 4227 | 3821 | 0.90 |
| Block 4, No. 12 | GSK3b | 1733 | 1153 | 0.67 |
| Block 4, No. 13 | p19ARF | 2247 | 1994 | 0.89 |
| Block 4, No. 14 | p57Kip2 | 310 | 258 | 0.83 |
| Sample ID | Gender | Age 1 | Biopsy Source | ABCC6 Genotype 2 | Genotype Status | Phenodex Score 3 | |
|---|---|---|---|---|---|---|---|
| PXE patients | |||||||
| P3M a | male | 57 | Neck | c.3421C>T (p.Arg1141*) | c.3883-6G>A (SSM) | cht | V2; C0 |
| P128M a | male | 51 | Neck | c.3769_3770insC (p.L1259fsX1277) | c.3769_3770insC (p.L1259fsX1277) | hm | S2; E2; G0; C1 |
| P255F a | female | 48 | Arm | c.3421C>T (p.Arg1141*) | c.2787+1G>T | cht | S3; E2; G0; C0 |
| Healthy controls | |||||||
| M57A b (AG13145) | male | 57 | Arm | - | - | wt | Not applicable |
| M52A b (AG11482) | male | 52 | Arm | - | - | wt | Not applicable |
| F48A b (AG14284) | female | 48 | Arm | - | - | wt | Not applicable |
| Gene | Protein | 5′-3′Sequence | Reference 1 | Annealing Temperature (°C) | Efficiency |
|---|---|---|---|---|---|
| β-ACTIN beta-Actin | β-Actin | CGCGAGAAGATGACCC ATTGCCAATGGTGATGAC | NM_001101 | 59 | 2.0 |
| GAPDH glycerinaldehyd-3-phosphat-dehydrogenase | GAPDH | AGGTCGGAGTCAACGGAT TCCTGGAAGATGGTGATG | NM_002046 | 59 | 1.8 |
| β2M beta-2- microglobulin | β2M | TGTGCTCGCGCTACTCTCTCTT CGGATGGATGAAACCCAGACA | NM_004048 | 59 | 2.0 |
| ICAM1 Intercellular adhesion molecule 1 | ICAM1 | ACCATCTACAGCTTTCCGGC CAATCCCTCTCGTCCAGTCG | NM_000201.3 | 63 | 1.9 |
| IL6 Interleukin 6 | IL6 | ACAGCCACTCACCTCTTCAG GTGCCTCTTTGCTGCTTTCAC | NM 000600.4 | 63 | 1.9 |
| LMNB1 Lamin B1 | Lamin B1 | GCAGACTTACCATGCCAAAC TCCCTTATTTCCGCCATCTC | NM 005573.3 | 63 | 1.9 |
| MCP1 Monocyte chemotactic Protein 1 | MCP1 | CTTCTGTGCCTGCTGCTCATA GGACACTTGTCGCTGGTGATT | NM 002982.3 | 66 | 2.0 |
| p21/CDKN1A cyclin-dependent kinase inhibitor 1A | p21 | GCAGACCAGCATGACAGATTTC ACCTCCGGGAGAGAGGAAAA | NM_000389.4 | 66 | 1.8 |
| p27/CDKN1B Cyclin-dependent kinase inhibitor 1B | p27 | CAGCTTGCCCGAGTTCTACT AGAAGAATCGTCGGTTGCAGG | NM_004064.4 | 66 | 2.0 |
| p53 tumor protein p53 | p53 | AGATAGCGATGGTCTGGC TTGGGCAGTGCTCGCTTAGT | NM_000546.5 | 63 | 2.0 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tiemann, J.; Wagner, T.; Lindenkamp, C.; Plümers, R.; Faust, I.; Knabbe, C.; Hendig, D. Linking ABCC6 Deficiency in Primary Human Dermal Fibroblasts of PXE Patients to p21-Mediated Premature Cellular Senescence and the Development of a Proinflammatory Secretory Phenotype. Int. J. Mol. Sci. 2020, 21, 9665. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21249665
Tiemann J, Wagner T, Lindenkamp C, Plümers R, Faust I, Knabbe C, Hendig D. Linking ABCC6 Deficiency in Primary Human Dermal Fibroblasts of PXE Patients to p21-Mediated Premature Cellular Senescence and the Development of a Proinflammatory Secretory Phenotype. International Journal of Molecular Sciences. 2020; 21(24):9665. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21249665
Chicago/Turabian StyleTiemann, Janina, Thomas Wagner, Christopher Lindenkamp, Ricarda Plümers, Isabel Faust, Cornelius Knabbe, and Doris Hendig. 2020. "Linking ABCC6 Deficiency in Primary Human Dermal Fibroblasts of PXE Patients to p21-Mediated Premature Cellular Senescence and the Development of a Proinflammatory Secretory Phenotype" International Journal of Molecular Sciences 21, no. 24: 9665. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21249665