Telocytes/CD34+ Stromal Cells in Pathologically Affected White Adipose Tissue

 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
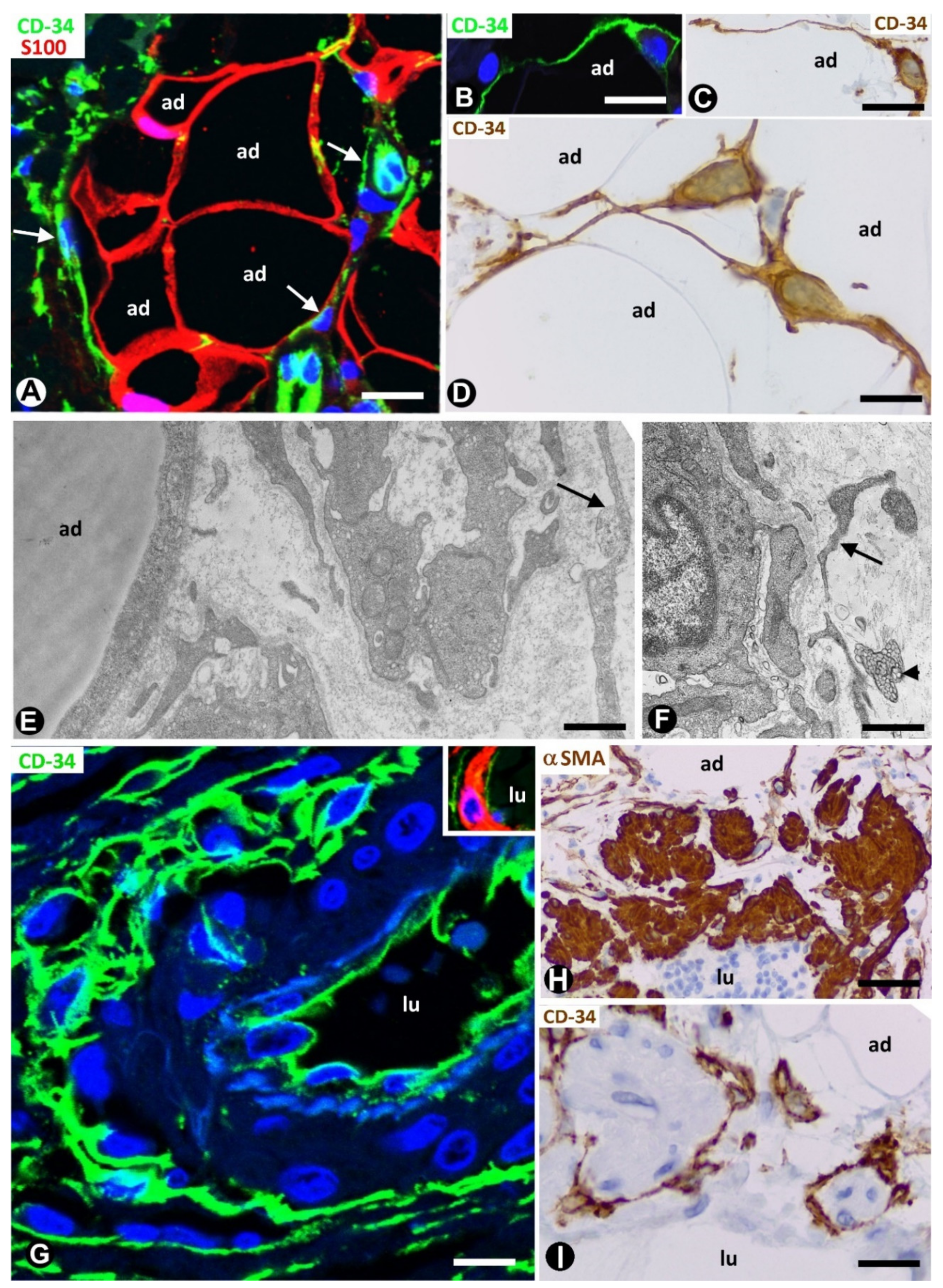
2. TCs/CD34+SCs in Normal Adipose Tissue
3. TCs/CD34+SCs in Pathologically Affected Adipose Tissue
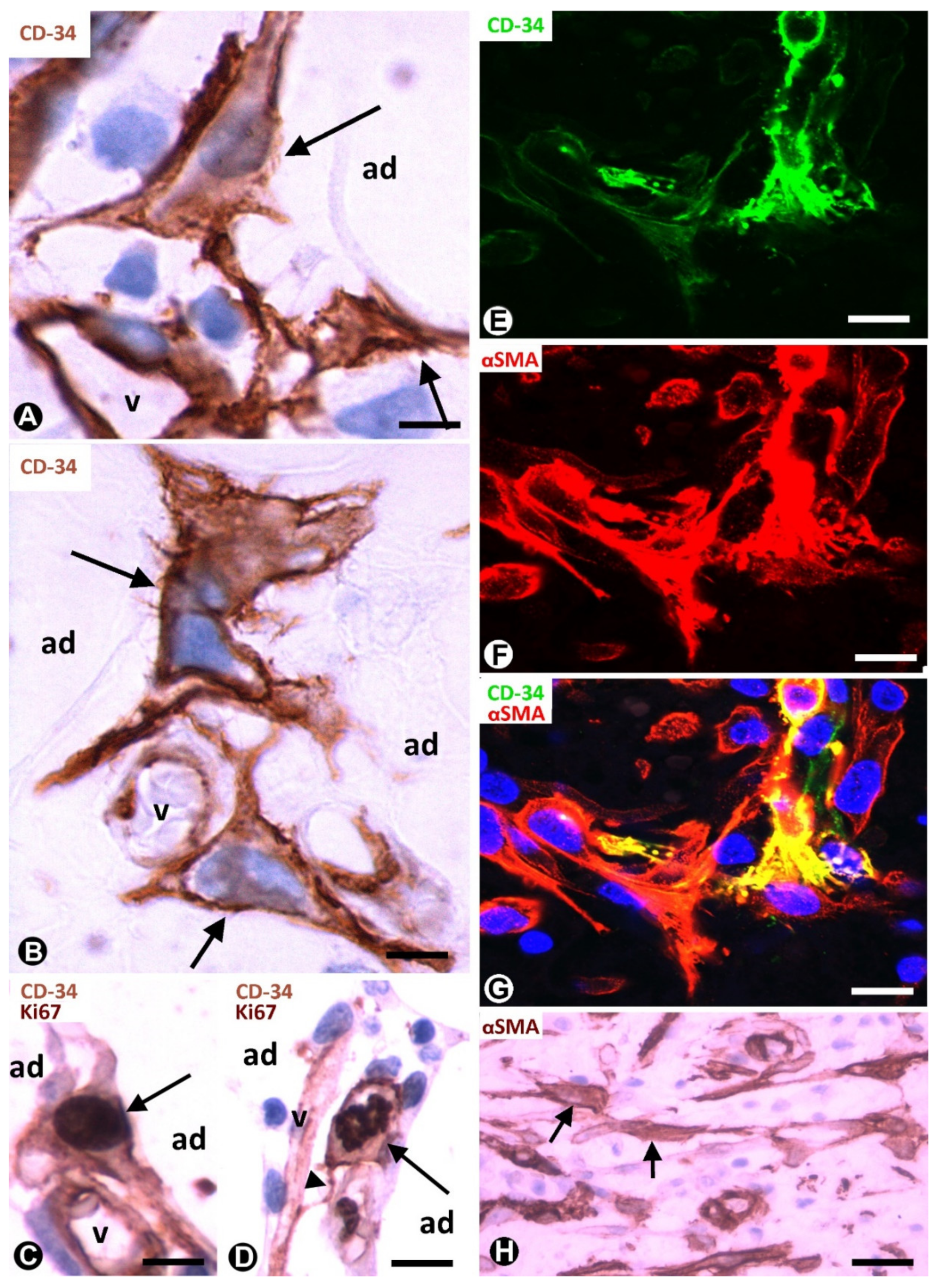
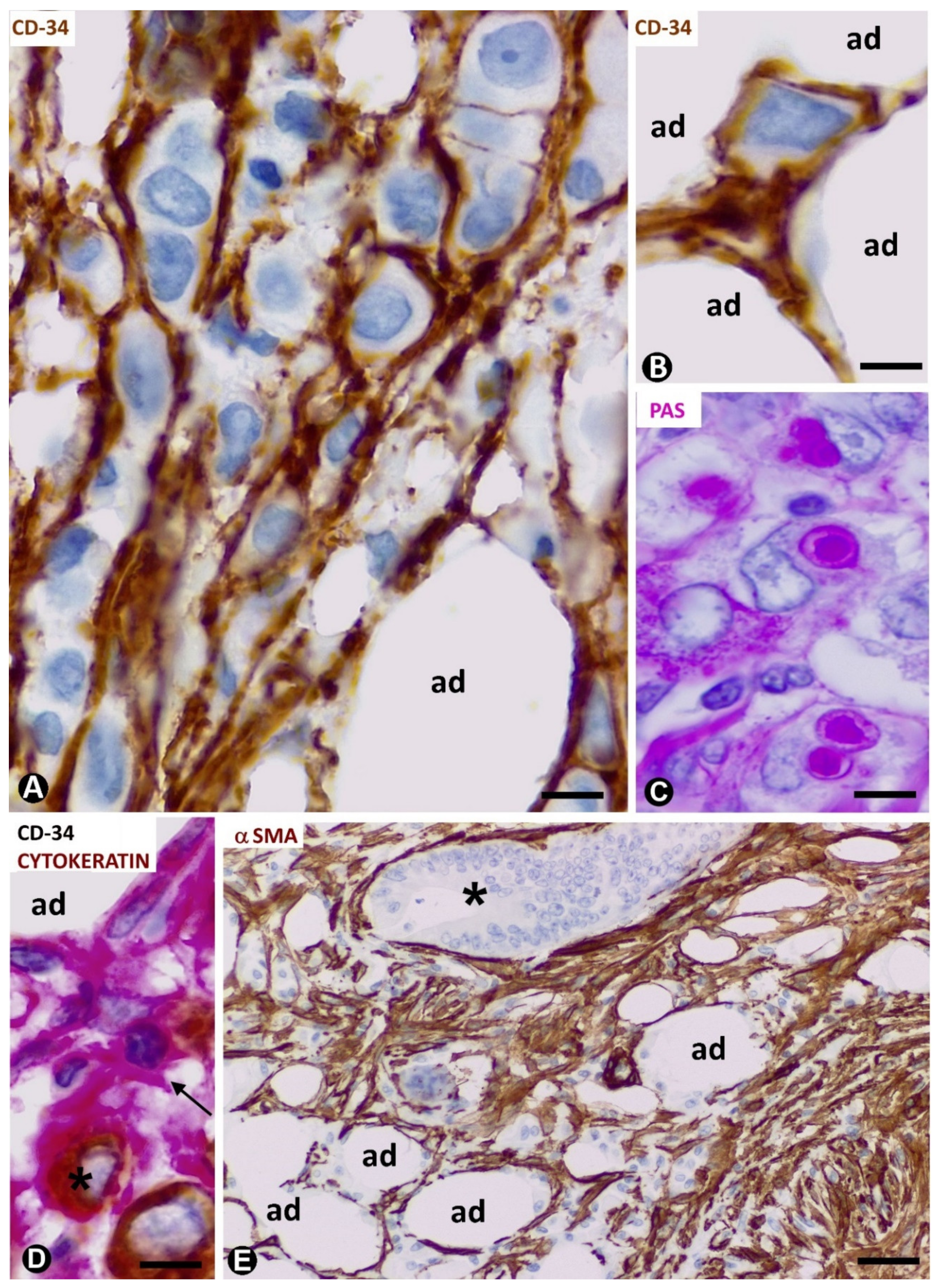
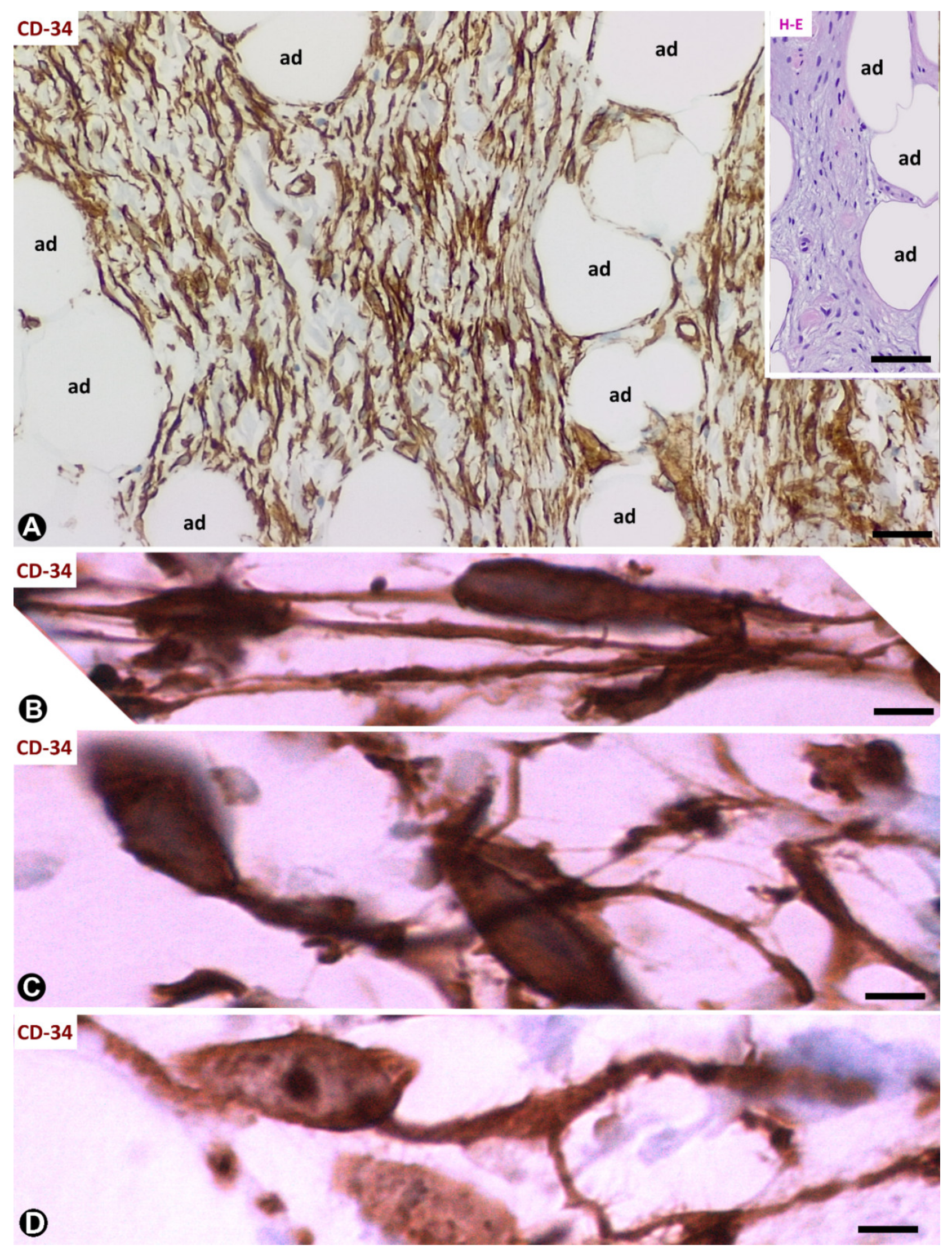
3.1. TCs/CD34+SCs of Adipose Tissue in Early and Most Advanced Stages of Inflammation and Repair through Granulation Tissue
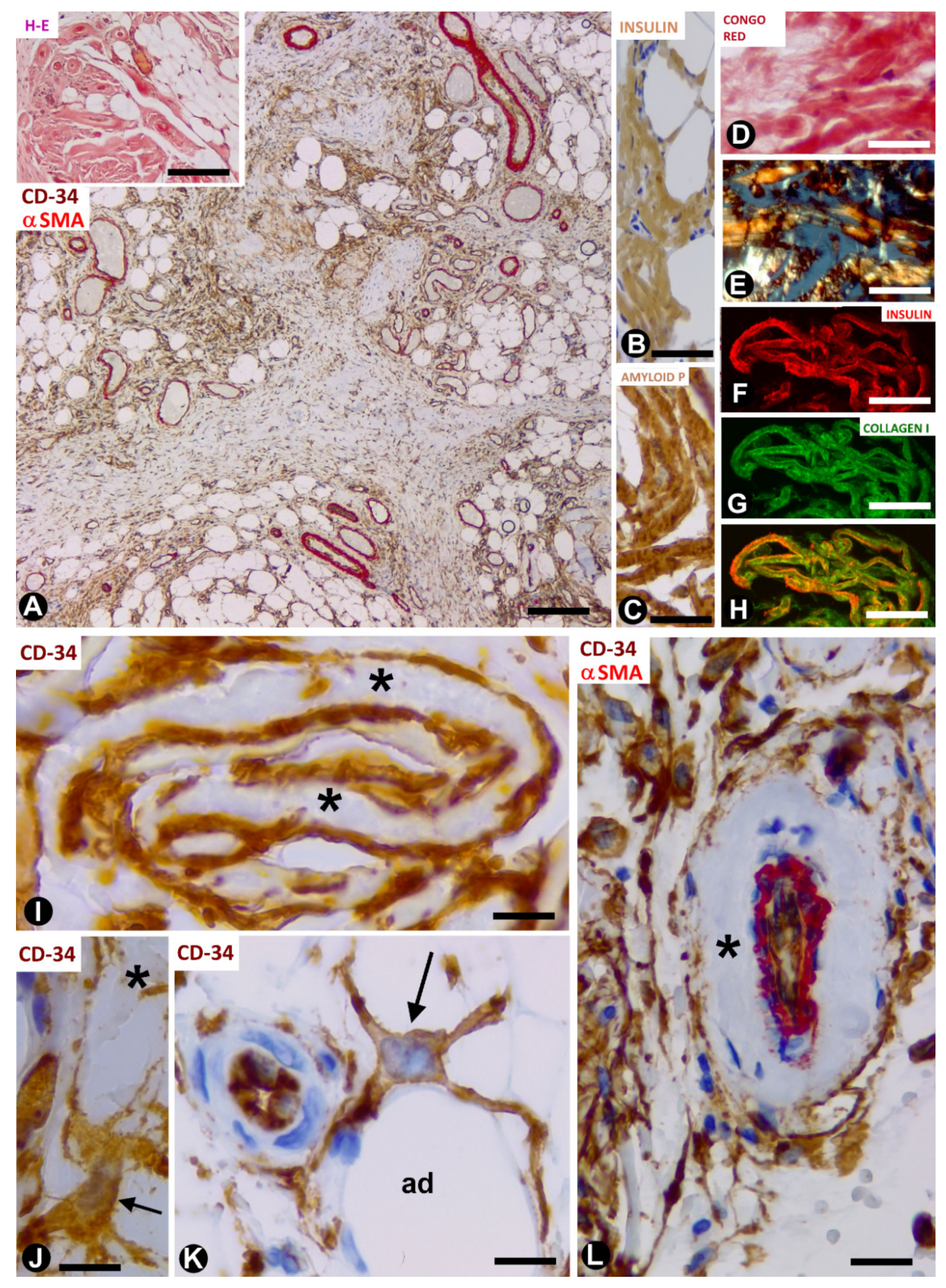
3.2. TCs/CD34+SCs in Adipose Tissue with Iatrogenic Insulin-Amyloid Type Amyloidosis
3.3. TCs/CD34+SCs in Non-Adipose Pathologic Processes Growing in Adipose Tissue
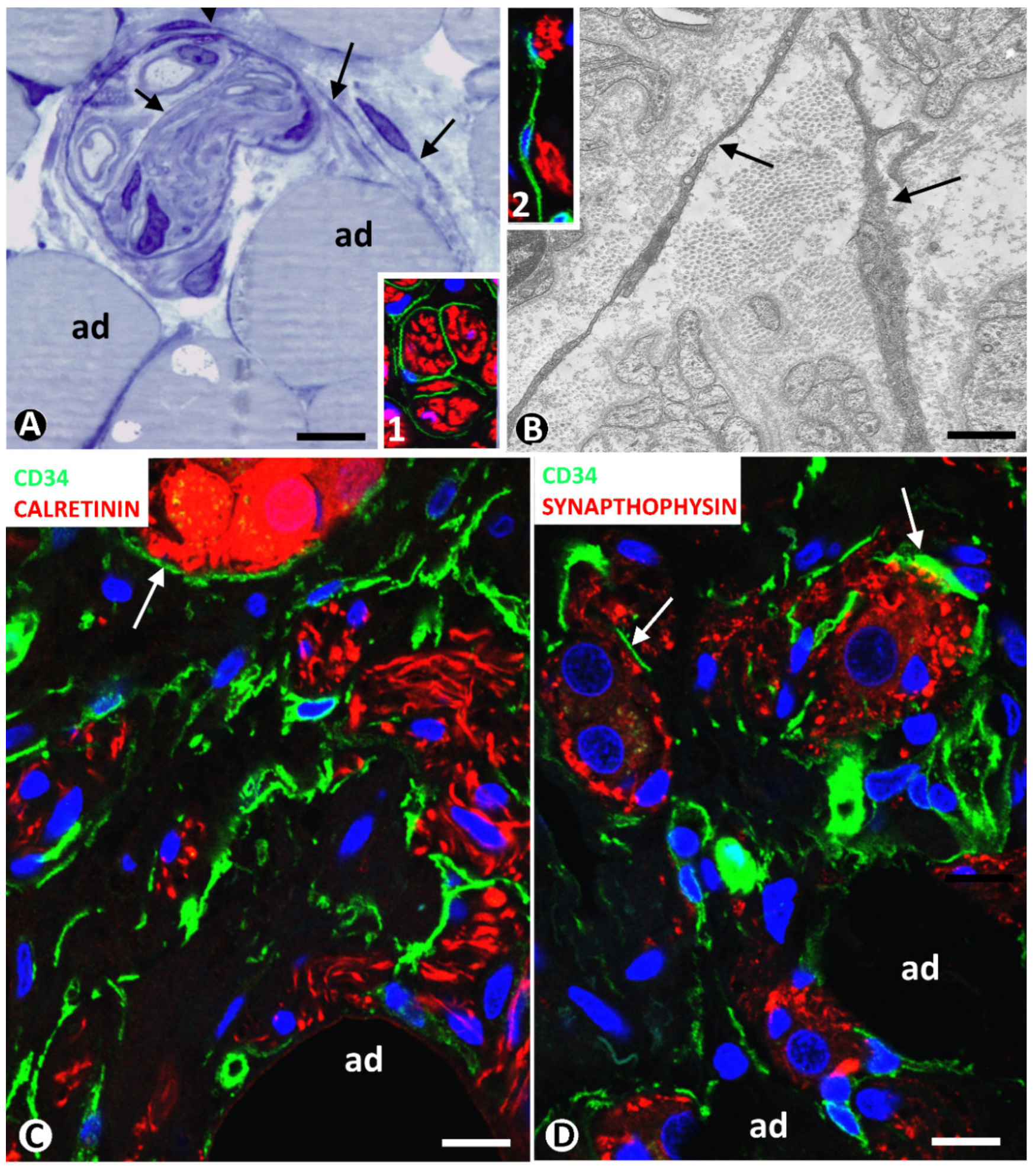
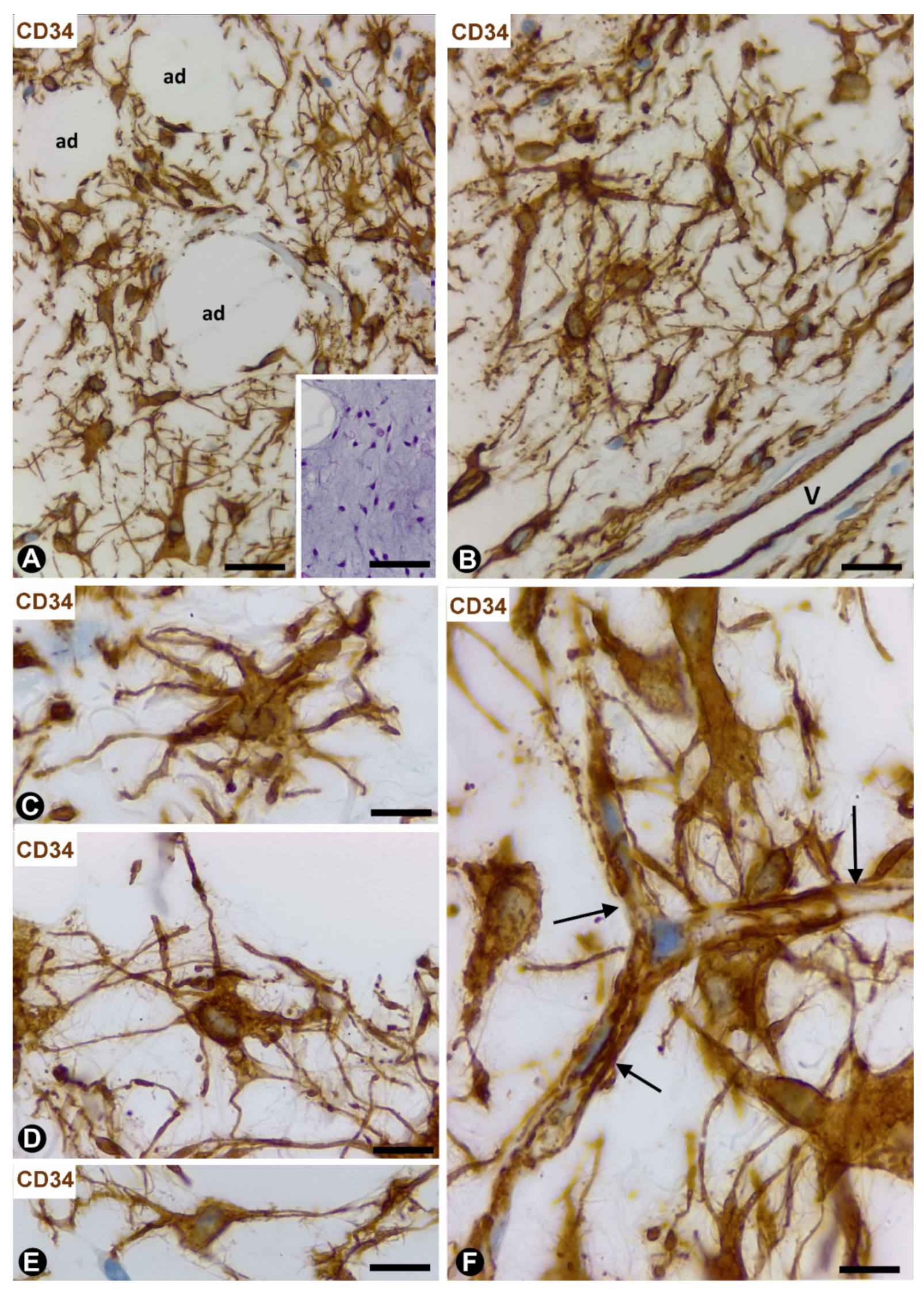
3.3.1. TCs/CD34+SCs in Neuromas and Hyperplastic Neurogenic Processes Affecting Adipose Tissue
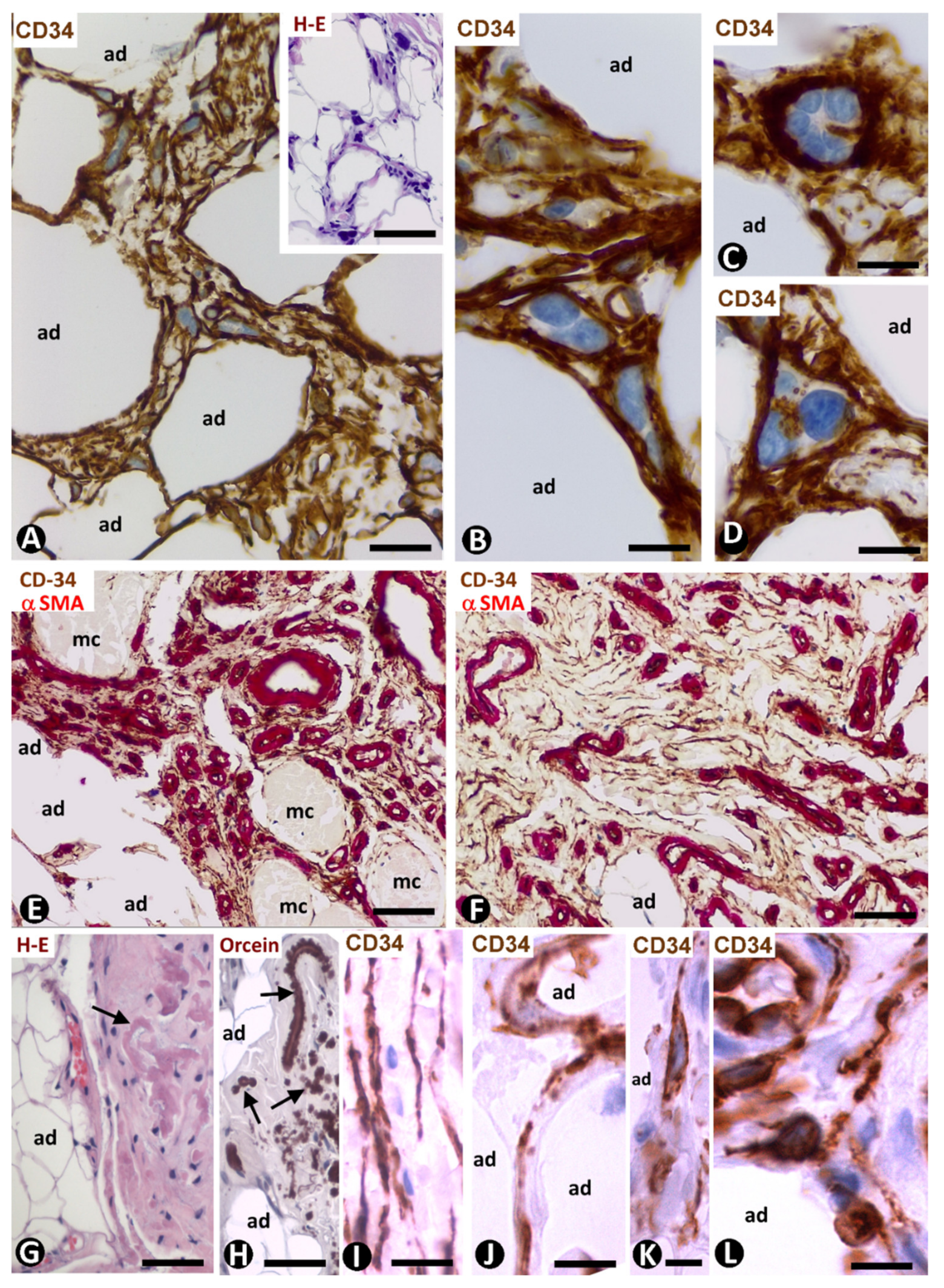
3.3.2. CD34+ and αSMA+SCs in Tumours Infiltrating Adipose Tissue
4. TCs/CD34+SCs in Tumours/Pseudo-Tumours of Adipose Tissue
4.1. TCs/CD34+SCs in Spindle Cell Lipoma
4.2. TCs/CD34+SCs in Dendritic Fibromyxolipoma
4.3. TCs/CD34+SCs in Pleomorphic Lipoma
4.4. TCs/CD34+SCs in Infiltrating Angiolipoma of Skeletal Muscle
4.5. TCs/CD34+SCs in the Elastofibrolipoma
5. TCs/CD34+SCs in Other Processes Affecting Adipose Tissue
5.1. TCs/CD34+SCs in Lipomatous Hypertrophy of the Interatrial Septum
5.2. TCs/CD34+SCs in the Nevus Lipomatosus Cutaneous Superficialis of Hoffman–Zurhelle
5.3. TCs/CD34+SCs in Irradiated Adipose Tissue
6. General Considerations about the Behaviour of TCs/CD34+SCs According to the Pathological Process in Adipose Tissue, Conclusions and Required Future Studies
Funding
Conflicts of Interest
References
- Zwick, R.K.; Guerrero-Juarez, C.F.; Horsley, V.; Plikus, M.V. Anatomical, physiological, and functional diversity of adipose tissue. Cell Metab. 2018, 27, 68–83. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Braun, J.; Kurtz, A.; Barutcu, N.; Bodo, J.; Thiel, A.; Dong, J. Concerted regulation of CD34 and CD105 accompanies mesenchymal stromal cell derivation from human adventitial stromal cell. Stem Cells Dev. 2013, 22, 815–827. [Google Scholar] [CrossRef] [PubMed]
- Lin, G.; Garcia, M.; Ning, H.; Banie, L.; Guo, Y.L.; Lue, T.F.; Lin, C.S. Defining stem and progenitor cells within adipose tissue. Stem Cells Dev. 2008, 17, 1053–1063. [Google Scholar] [CrossRef] [PubMed]
- Mitchell, J.B.; McIntosh, K.; Zvonic, S.; Garrett, S.; Floyd, Z.E.; Kloster, A.; Di Halvorsen, Y.; Storms, R.W.; Goh, B.; Kilroy, G.; et al. Immunophenotype of human adipose-derived cells: Temporal changes in stromal-associated and stem cell-associated markers. Stem Cells 2006, 24, 376–385. [Google Scholar] [CrossRef]
- Peng, Q.; Alipour, H.; Porsborg, S.; Fink, T.; Zachar, V. Evolution of ASC immunophenotypical subsets during expansion in vitro. Int. J. Mol. Sci. 2020, 21, 1408. [Google Scholar] [CrossRef] [Green Version]
- Faussone-Pellegrini, M.S.; Popescu, L.M. Telocytes. BioMol. Concepts 2011, 2, 481–489. [Google Scholar] [CrossRef]
- Popescu, L.M.; Faussone-Pellegrini, M.S. Telocytes—A case of serendipity: The winding way from Interstitial Cells of Cajal (ICC), via Interstitial Cajal-Like Cells (ICLC) to telocytes. J. Cell. Mol. Med. 2010, 14, 729–740. [Google Scholar] [CrossRef] [Green Version]
- Bani, D.; Formigli, L.; Gherghiceanu, M.; Faussone-Pellegrini, M.S. Telocytes as supporting cells for myocardial tissue organization in developing and adult heart. J. Cell. Mol. Med. 2010, 14, 2531–2538. [Google Scholar] [CrossRef] [Green Version]
- Cretoiu, S.M.; Popescu, L.M. Telocytes revisited. BioMol. Concepts 2014, 5, 353–369. [Google Scholar] [CrossRef]
- Díaz-Flores, L.; Gutiérrez, R.; García, M.P.; Sáez, F.J.; Aparicio, F.; Díaz-Flores, L., Jr.; Madrid, J.F. Uptake and intracytoplasmic storage of pigmented particles by human CD34+ stromal cells/telocytes: Endocytic property of telocytes. J. Cell. Mol. Med. 2014, 18, 2478–2487. [Google Scholar] [CrossRef]
- Marini, M.; Ibba-Manneschi, L.; Rosa, I.; Sgambati, E.; Manetti, M. Changes in the telocyte/CD34+ stromal cell and α-+ myoid cell networks in human testicular seminoma. Acta Histochem. 2019, 121, 151442. [Google Scholar] [CrossRef] [PubMed]
- Nicolescu, M.I.; Rusu, M.C.; Voinea, L.M.; Vrapciu, A.D.; Bâră, R.I. Lymphatic lacunae of the human eye conjunctiva embedded within a stroma containing CD34+ telocytes. J. Cell. Mol. Med. 2020, 24, 8871–8875. [Google Scholar] [CrossRef] [PubMed]
- Romano, E.; Rosa, I.; Fioretto, B.S.; Lucattelli, E.; Innocenti, M.; Ibba-Manneschi, L.; Matucci-Cerinic, M.; Manetti, M. A Two-Step immunomagnetic microbead-based method for the isolation of human primary skin telocytes/CD34+ stromal cells. Int. J. Mol. Sci. 2020, 21, 5877. [Google Scholar] [CrossRef] [PubMed]
- Rosa, I.; Marini, M.; Guasti, D.; Ibba-Manneschi, L.; Manetti, M. Morphological evidence of telocytes in human synovium. Sci. Rep. 2018, 8, 3581. [Google Scholar] [CrossRef]
- Rusu, M.C.; Mănoiu, V.S.; Creţoiu, D.; Creţoiu, S.M.; Vrapciu, A.D. Stromal cells/telocytes and endothelial progenitors in the perivascular niches of the trigeminal ganglion. Ann. Anat. 2018, 218, 141–155. [Google Scholar] [CrossRef]
- Zhou, Q.; Wei, L.; Zhong, C.; Fu, S.; Bei, Y.; Huică, R.I.; Wang, F.; Xiao, J. Cardiac telocytes are double positive for CD34/PDGFR-α. J. Cell. Mol. Med. 2015, 19, 2036–2042. [Google Scholar] [CrossRef]
- Zhang, H.; Yu, P.; Zhong, S.; Ge, T.; Peng, S.; Guo, X.; Zhou, Z. Telocytes in pancreas of the Chinese giant salamander (Andrias davidianus). J. Cell. Mol. Med. 2016, 20, 2215–2219. [Google Scholar] [CrossRef]
- Estève, D.; Boulet, N.; Belles, C.; Zakaroff-Girard, A.; Decaunes, P.; Briot, A.; Veeranagouda, Y.; Didier, M.; Remaury, A.; Guillemot, J.C.; et al. Lobular architecture of human adipose tissue defines the niche and fate of progenitor cells. Nat. Commun. 2019, 10, 2549. [Google Scholar] [CrossRef] [Green Version]
- Cretoiu, D.; Xu, J.; Xiao, J.; Cretoiu, S.M. Telocytes and their extracellular vesicles-evidence and hypotheses. Int. J. Mol. Sci. 2016, 17, 1322. [Google Scholar] [CrossRef]
- Cretoiu, D.; Roatesi, S.; Bica, I.; Plesca, C.; Stefan, A.; Bajenaru, O.; Condrat, C.E.; Cretoiu, S.M. Simulation and modeling of telocytes behavior in signaling and intercellular communication processes. Int. J. Mol. Sci. 2020, 21, 2615. [Google Scholar] [CrossRef] [Green Version]
- Ibba-Manneschi, L.; Rosa, I.; Manetti, M. Telocyte implications in human pathology: An overview. Semin. Cell. Dev. Biol. 2016, 55, 62–69. [Google Scholar] [CrossRef] [PubMed]
- Nicolescu, M.I.; Bucur, A.; Dinca, O.; Rusu, M.C.; Popescu, L.M. Telocytes in parotid glands. Anat Rec. 2012, 295, 378–385. [Google Scholar] [CrossRef] [PubMed]
- Veress, B.; Ohlsson, B. Spatial relationship between telocytes, interstitial cells of Cajal and the enteric nervous system in the human ileum and colon. J. Cell. Mol. Med. 2020, 24, 3399–3406. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Díaz-Flores, L.; Gutiérrez, R.; García, M.P.; González, M.; Sáez, F.J.; Aparicio, F.; Díaz-Flores, L., Jr.; Madrid, J.F. Human resident CD34+ stromal cells/telocytes have progenitor capacity and are a source of αSMA+ cells during repair. Histol. Histopathol. 2015, 30, 615–627. [Google Scholar]
- Díaz-Flores, L.; Gutiérrez, R.; Litzarza, K.; González-Gómez, M.; García, M.P.; Saez, F.J.; Díaz-Flores, L., Jr.; Madrid, J.F. Behavior of in situ human native adipose tissue CD34+ stromal/progenitor cells during different stages of repair. Tissue-resident CD34+ stromal cells as a source of myofibroblasts. Anat. Rec. 2015, 298, 917–930. [Google Scholar] [CrossRef]
- Díaz-Flores, L.; Gutiérrez, R.; García, M.P.; González-Gómez, M.; Díaz-Flores, L., Jr.; Álvarez-Argüelles, H.; Carrasco, J.L. Presence/Absence and specific location of resident CD34+ stromal cells/telocytes condition stromal cell development in repair and tumors. Front. Cell. Dev. Biol. 2020, 8, 544845. [Google Scholar] [CrossRef]
- Vannucchi, M.G.; Bani, D.; Faussone-Pellegrini, M.S. Telocytes contribute as cell progenitors and differentiation inductors in tissue regeneration. Curr. Stem Cell Res. Ther. 2016, 11, 383–389. [Google Scholar] [CrossRef]
- Bani, D.; Nistri, S. New insights into the morphogenic role of stromal cells and their relevance for regenerative medicine. lessons from the heart. J. Cell. Mol. Med. 2014, 18, 363–370. [Google Scholar] [CrossRef]
- Ceafalan, L.; Gherghiceanu, M.; Popescu, L.M.; Simionescu, O. Telocytes in human skin--are they involved in skin regeneration? J. Cell. Mol. Med. 2012, 16, 1405–1420. [Google Scholar] [CrossRef]
- Gherghiceanu, M.; Popescu, L.M. Cardiomyocyte precursors and telocytes in epicardial stem cell niche: Electron microscope images. J. Cell. Mol. Med. 2010, 14, 871–877. [Google Scholar] [CrossRef] [Green Version]
- Gibbons, S.J.; De Giorgio, R.; Faussone Pellegrini, M.S.; Garrity-Park, M.M.; Miller, S.M.; Schmalz, P.F.; Young-Fadok, T.M.; Larson, D.W.; Dozois, E.J.; Camilleri, M.; et al. Apoptotic cell death of human interstitial cells of Cajal. Neurogastroenterol. Motil. 2009, 2, 85–93. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Popescu, L.M.; Manole, E.; Serboiu, C.S.; Manole, C.G.; Suciu, L.C.; Gherghiceanu, M.; Popescu, B.O. Identification of telocytes in skeletal muscle interstitium: Implication for muscle regeneration. J. Cell. Mol. Med. 2011, 15, 1379–1392. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Popescu, L.M.; Gherghiceanu, M.; Suciu, L.C.; Manole, C.G.; Hinescu, M.E. Telocytes and putative stem cells in the lungs: Electron microscopy, electron tomography and laser scanning microscopy. Cell Tissue Res. 2011, 345, 391–403. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Manetti, M.; Tani, A.; Rosa, I.; Chellini, F.; Squecco, R.; Idrizaj, E.; Zecchi-Orlandini, S.; Ibba-Manneschi, L.; Sassoli, C. Morphological evidence for telocytes as stromal cells supporting satellite cell activation in eccentric contraction-induced skeletal muscle injury. Sci. Rep. 2019, 9, 14515. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Popescu, L.M.; Nicolescu, M.I. Telocytes and stem cells. In Resident Stem Cells and Regenerative Therapy; dos Santos Goldenberg, R.C., de Carvalho, A.C.C., Eds.; Academic Press: Cambridge, MA, USA; Elsevier: Oxford, UK, 2013; pp. 205–231. [Google Scholar]
- Sahoo, S.; Losordo, D.W. Exosomes and cardiac repair after myocardial infarction. Circ. Res. 2014, 114, 333–344. [Google Scholar] [CrossRef]
- Suciu, L.; Popescu, L.M.; Gherghiceanu, M.; Regalia, T.; Nicolescu, M.I.; Hinescu, M.E.; Faussone-Pellegrini, M.S. Telocytes in human term placenta: Morphology and phenotype. Cells Tissues Organs 2010, 192, 325–339. [Google Scholar] [CrossRef]
- Vannucchi, M.G.; Traini, C.; Manetti, M.; Ibba-Manneschi, L.; Faussone-Pellegrini, M.S. Telocytes express PDGFRα in the human gastrointestinal tract. J. Cell. Mol. Med. 2013, 17, 1099–1108. [Google Scholar] [CrossRef]
- Wang, F.; Song, Y.; Bei, Y.; Zhao, Y.; Xiao, J.; Yang, C. Telocytes in liver regeneration: Possible roles. J. Cell. Mol. Med. 2014, 18, 1720–1726. [Google Scholar] [CrossRef]
- Zhou, J.; Wang, Y.; Zhu, P.; Sun, H.; Mou, Y.; Duan, C.; Yao, A.; Lv, S.; Wang, C. Distribution and characteristics of telocytes as nurse cells in the architectural organization of engineered heart tissues. Sci. China Life Sci. 2014, 57, 241–247. [Google Scholar] [CrossRef] [Green Version]
- Mandache, E.; Gherghiceanu, M.; Macarie, C.; Kostin, S.; Popescu, L.M. Telocytes in human isolated atrial amyloidosis: Ultrastructural remodelling. J. Cell. Mol. Med. 2010, 14, 2739–2747. [Google Scholar] [CrossRef] [Green Version]
- Iwasa, S.; Enomoto, A.; Onoue, S.; Nakai, M. Chromatographic analysis of conformationally changed insulin and its cytotoxic effect on PC12 cells. J. Health Sci. 2009, 55, 825–831. [Google Scholar] [CrossRef] [Green Version]
- Díaz-Flores, L.; Gutiérrez, R.; García, M.P.; Gayoso, S.; Gutiérrez, E.; Díaz-Flores, L., Jr.; Carrasco, J.L. Telocytes in the normal and pathological peripheral nervous system. Int. J. Mol. Sci. 2020, 21, 4320. [Google Scholar] [CrossRef] [PubMed]
- Suster, S.; Fisher, C. Immunoreactivity for the human hematopoietic progenitor cell antigen (CD34) in lipomatous tumors. Am. J. Surg. Pathol. 1997, 21, 195–200. [Google Scholar] [CrossRef] [PubMed]
- Lin, T.M.; Chang, H.W.; Wang, K.H.; Kao, A.P.; Chang, C.C.; Wen, C.H.; Lai, C.S.; Lin, S.D. Isolation and identification of mesenchymal stem cells from human lipoma tissue. Biochem. Biophys. Res. Commun. 2007, 361, 883–889. [Google Scholar] [CrossRef]
- Inatani, H.; Yamamoto, N.; Hayashi, K.; Kimura, H.; Takeuchi, A.; Miwa, S.; Higuchi, T.; Abe, K.; Taniguchi, Y.; Yamada, S.; et al. Do mesenchymal stem cells derived from atypical lipomatous tumors have greater differentiation potency than cells from normal adipose tissues? Clin. Orthop. Relat. Res. 2017, 475, 1693–1701. [Google Scholar] [CrossRef] [Green Version]
- Tremp, M.; Menzi, N.; Tchang, L.; di Summa, P.G.; Schaefer, D.J.; Kalbermatten, D.F. Adipose-derived stromal cells from lipomas: Isolation, characterisation and review of the literature. Pathobiology 2016, 83, 258–266. [Google Scholar] [CrossRef]
- Qian, Y.W.; Gao, J.H.; Lu, F.; Zheng, X.D. The differences between adipose tissue derived stem cells and lipoma mesenchymal stem cells in characteristics. Zhonghua Zheng Xing Wai Ke Za Zhi. 2010, 26, 125–132. [Google Scholar]
- Enzinger, F.M.; Harvey, D.A. Spindle cell lipoma. Cancer 1975, 36, 1852–1859. [Google Scholar] [CrossRef]
- Suster, S.; Fisher, C.; Moran, C.A. Dendritic fibromyxolipoma: Clinicopathologic study of a distinctive benign soft tissue lesion that may be mistaken for a sarcoma. Ann. Diagn. Pathol. 1998, 2, 111–120. [Google Scholar] [CrossRef]
- Enzinger, F.M. Benign lipomatous tumors simulating a sarcoma. In Management of Primary Bone and Soft Tissue Tumors; Year Book Medical: Chicago, IL, USA, 1977; pp. 11–24. [Google Scholar]
- Xanthos, T.; Giannakopoulos, N.; Papadimitriou, L. Lipomatous hypertrophy of the interatrial septum: A pathological and clinical approach. Int. J. Cardiol. 2007, 121, 4–8. [Google Scholar] [CrossRef]









Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Díaz-Flores, L.; Gutiérrez, R.; García, M.P.; González-Gómez, M.; Carrasco, J.L.; Alvarez-Argüelles, H.; Díaz-Flores, L., Jr. Telocytes/CD34+ Stromal Cells in Pathologically Affected White Adipose Tissue. Int. J. Mol. Sci. 2020, 21, 9694. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21249694
Díaz-Flores L, Gutiérrez R, García MP, González-Gómez M, Carrasco JL, Alvarez-Argüelles H, Díaz-Flores L Jr. Telocytes/CD34+ Stromal Cells in Pathologically Affected White Adipose Tissue. International Journal of Molecular Sciences. 2020; 21(24):9694. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21249694
Chicago/Turabian StyleDíaz-Flores, Lucio, Ricardo Gutiérrez, Ma Pino García, Miriam González-Gómez, Jose Luís Carrasco, Hugo Alvarez-Argüelles, and Lucio Díaz-Flores, Jr. 2020. "Telocytes/CD34+ Stromal Cells in Pathologically Affected White Adipose Tissue" International Journal of Molecular Sciences 21, no. 24: 9694. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21249694