Dystonia: Sparse Synapses for D2 Receptors in Striatum of a DYT1 Knock-out Mouse Model

 ,
, 
Abstract
:1. Introduction
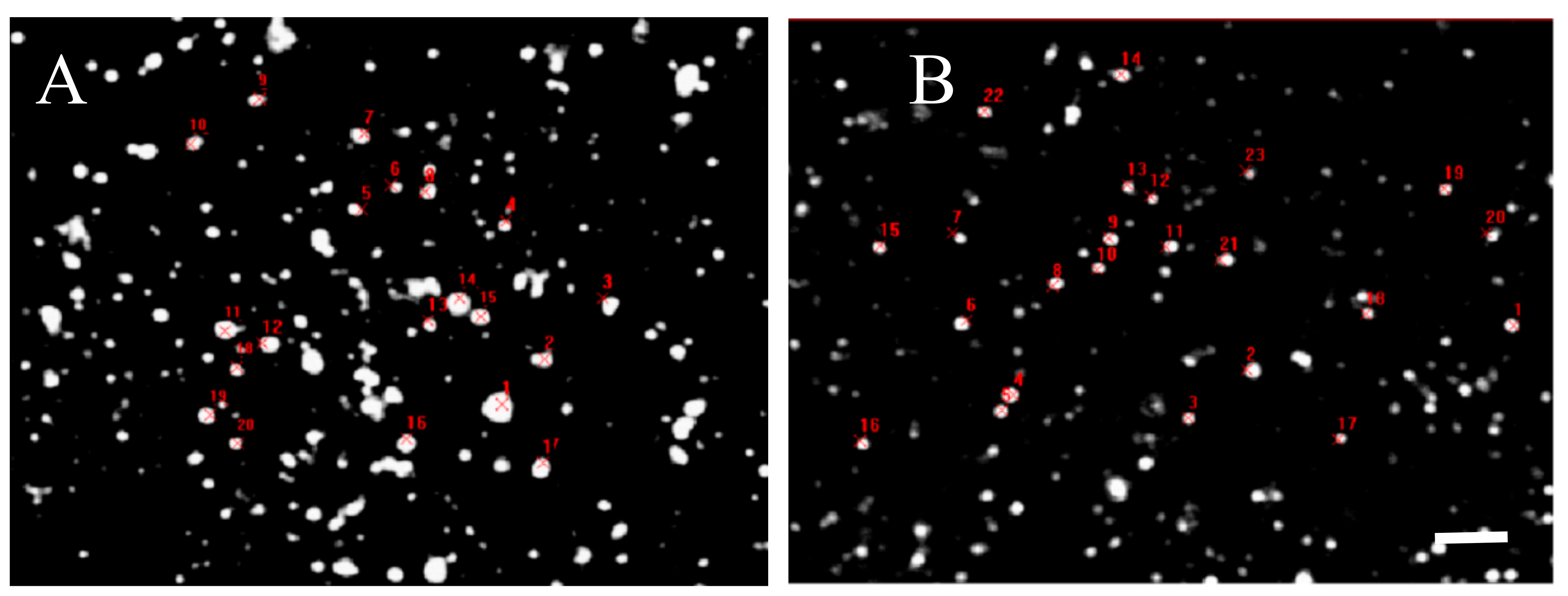
2. Results
3. Discussion
4. Materials and Methods
4.1. Animals and Methods
4.2. Quantitative Analysis of D2 Protein
4.3. Immune-Histochemistry
4.4. Immune-Fluorescence Techniques and Confocal Microscopy
4.5. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Marsden, C.D.; Sheehy, M.P. Writer’s cramp. Trends Neurosci. 1990, 13, 148–153. [Google Scholar] [CrossRef]
- Berardelli, A.; Rothwell, J.C.; Hallett, M.; Thompson, P.D.; Manfredi, M.; Marsden, C.D. The pathophysiology of primary dystonia. Brain 1998, 121, 1195–1212. [Google Scholar] [CrossRef] [PubMed]
- Lanska, D.J. Chapter 33: The History of Movement Disorders. In Handbook of Clinical Neurology. The History of Neurology; Elsevier: Amsterdam, The Netherlands, 2010; Volume 95, pp. 501–546. [Google Scholar]
- Albanese, A.; Bhatia, K.; Bressman, S.B.; Delong, M.R.; Fahn, S.; Fung, V.S.; Hallett, M.; Jankovic, J.; Jinnah, H.A.; Klein, C.; et al. Phenomenology and classification of dystonia: A consensus update. Mov. Disord. 2013, 28, 863–873. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Munts, A.G.; Koehler, P.J. How psychogenic is dystonia? Views from past to present. Brain 2010, 133 Pt 5, 1552–1564. [Google Scholar] [CrossRef] [Green Version]
- Schwalbe, W. Eine Eigentumliche Tonische Krampfform mit hysterischen Symptomen; Inaugural-Dissertation-UniversitätsBuch-druckerei, G. Schade:: Berlin, Germany, 1908. [Google Scholar]
- Oppenheim, H. Uber eine eigenartige Krampfkrankheit des kindlichen und jugendlichen Alters (Dysbasia lordotica progressiva, Dystonia musculorum deformans). Neurol. Centrabl. 1911, 30, 1090–1107. [Google Scholar]
- Ozelius, L.J.; Hewett, J.W.; Page, C.E.; Bressman, S.B.; Kramer, P.L.; Shalish, C.; de Leon, D.; Brin, M.F.; Raymond, D.; Corey, D.P.; et al. The early-onset torsion dystonia gene (DYT1) encodes an ATP-binding protein. Nat. Genet. 1997, 17, 40–48. [Google Scholar] [CrossRef]
- Fahn, S.; Bressman, S.B.; Marsden, C.D. Classification of dystonia. Adv. Neurol. 1998, 78, 1–10. [Google Scholar] [CrossRef]
- Standaert, D.G. Update on the pathology of dystonia. Neurobiol. Dis. 2011, 42, 148–151. [Google Scholar] [CrossRef] [Green Version]
- Rondot, P. The shadow of movement. J. Neurol. 1991, 238, 411–419. [Google Scholar] [CrossRef]
- Kanowsky, P. Dystonia: A disorder of motor programming or motor execution? Mov. Disord. 2002, 17, 1143–1147. [Google Scholar] [CrossRef]
- DeFazio, G.; Berardelli, A.; Hallett, M. Do primary adult-onset focal dystonias share aetiological factors? Brain 2007, 130, 1183–1193. [Google Scholar] [CrossRef] [PubMed]
- Ulug, A.M.; Vo, A.; Argyelan, M.; Tanabe, L.; Schiffer, W.K.; Dewey, S.; Dauer, W.T.; Eidelberg, D. Cerebellothalamocortical pathway abnormalities in torsinA DYT1 knock-in mice. Proc. Natl. Acad. Sci. USA 2011, 108, 6638–6643. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goodchild, R.E.; Grundmann, K.; Pisani, A. New genetic insights highlight ‘old’ ideas on motor dysfunction in dystonia. Trends Neurosci. 2013, 36, 717–725. [Google Scholar] [CrossRef] [PubMed]
- Asanuma, K.; Ma, Y.; Okulski, J.; Dhawan, V.; Chaly, T.; Carbon, M.; Bressman, S.B.; Eidelberg, D. Decreased striatal D2 receptor binding in non-manifesting carriers of the DYT1 dystonia mutation. Neurology 2005, 64, 347–349. [Google Scholar] [CrossRef] [PubMed]
- Carbon, M.; Niethammer, M.; Peng, S.; Raymond, D.; Dhawan, V.; Chaly, T.; Ma, Y.; Bressman, S.; Eidelberg, D. Abnormal striatal and thalamic dopamine neurotransmission: Genotype-related features of dystonia. Neurology 2009, 72, 2097–2103. [Google Scholar] [CrossRef] [Green Version]
- Napolitano, F.; Pasqualetti, M.; Usiello, A.; Santini, E.; Pacini, G.; Sciamanna, G.; Errico, F.; Tassone, A.; Di Dato, V.; Martella, G.; et al. Dopamine D2 receptor dysfunction is rescued by adenosine A2A receptor antagonism in a model of DYT1 dystonia. Neurobiol. Dis. 2010, 38, 434–445. [Google Scholar] [CrossRef] [Green Version]
- Yokoi, F.; Dang, M.T.; Li, J.; Standaert, D.G.; Li, Y. Motor deficits and decreased striatal dopamine receptor 2 binding activity in the striatum-specific Dyt1 conditional knock-out mice. PLoS ONE 2011, 6, e24539. [Google Scholar] [CrossRef] [Green Version]
- Dang, M.T.; Yokoi, F.; Cheetham, C.C.; Lu, J.; Vo, V.; Lovinger, D.M.; Li, Y. An anticholinergic reverses motor control and corticostriatal LTD deficits in Dyt1 DGAG knock-in mice. Behav. Brain Res. 2012, 226, 465–472. [Google Scholar] [CrossRef] [Green Version]
- Pisani, A.; Martella, G.; Tscherter, A.; Bonsi, P.; Sharma, N.; Bernardi, G.; Standaert, D. Altered responses to dopaminergic D2 receptor activation and N-type calcium currents in striatal cholinergic interneurons in a mouse model of DYT1 dystonia. Neurobiol. Dis. 2006, 24, 318–325. [Google Scholar] [CrossRef]
- Sciamanna, G.; Tassone, A.; Martella, G.; Mandolesi, G.; Puglisi, F.; Cuomo, D.; Madeo, G.; Ponterio, G.; Standaert, D.G.; Bonsi, P.; et al. Developmental profile of the aberrant dopamine D2 receptor response in striatal cholinergic interneurons in DYT1 dystonia. PLoS ONE 2011, 6, e24261. [Google Scholar] [CrossRef] [Green Version]
- Sciamanna, G.; Hollis, R.; Ball, C.; Martella, G.; Tassone, A.; Marshall, A.; Parsons, D.; Li, X.; Yokoi, F.; Zhang, L.; et al. Cholinergic dysregulation produced by selective inactivation of the dystonia-associated protein torsinA. Neurobiol. Dis. 2012, 47, 416–427. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martella, G.; Maltese, M.; Nistico, R.; Schirinzi, T.; Madeo, G.; Sciamanna, G.; Ponterio, G.; Tassone, A.; Mandolesi, G.; Vanni, V.; et al. Regional specificity of synaptic plasticity deficits in a knock-in mouse model of DYT1 dystonia. Neurobiol. Dis. 2014, 65, 124–132. [Google Scholar] [CrossRef] [PubMed]
- Scarduzio, M.; Zimmerman, C.N.; Jaunarajs, K.L.; Wang, Q.; Standaert, D.G.; McMahon, L.L. Strength of cholinergic tone dictates the polarity of dopamine D2 receptor modulation of striatal cholinergic interneuron excitability in DYT1 dystonia. Exp. Neurol. 2017, 295, 162–175. [Google Scholar] [CrossRef] [PubMed]
- Bonsi, P.; Ponterio, G.; Vanni, V.; Tassone, A.; Sciamanna, G.; Migliarini, S.; Martella, G.; Meringolo, M.; Dehay, B.; Doudnikoff, E.; et al. RGS 9-2 rescues dopamine D2 receptor levels and signaling in DYT1 dystonia mouse models. EMBO Mol. Med. 2018, 11, e9283. [Google Scholar]
- Peterson, D.A.; Sejnowski, T.J.; Poizner, H. Convergent evidence for abnormal striatal synaptic plasticity in dystonia. Neurobiol. Dis. 2010, 37, 558–573. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Quartarone, A.; Pisani, A. Abnormal plasticity in dystonia: Disruption of synaptic homeostasis. Neurobiol. Dis. 2011, 42, 162–170. [Google Scholar] [CrossRef]
- Yao, W.-D.; Spealman, R.D.; Zhang, J. Dopaminergic signaling in dendritic spines. Biochem. Pharmacol. 2008, 75, 2055–2069. [Google Scholar] [CrossRef] [Green Version]
- Hersch, S.M.; Ciliax, B.J.; Gutekunst, C.A.; Rees, H.D.; Heilman, C.J.; Yung, K.K.L.; Bolam, J.P.; Ince, E.; Yi, H.; Levey, A.I. Electron microscopic analysis of D1 and D2 dopamine receptor proteins in the dorsal striatum and their synaptic relationships with motor corticostriatal afferents. J. Neurosci. 1995, 75, 5222–5237. [Google Scholar] [CrossRef] [Green Version]
- Descarries, L.; Watkins, K.C.; Garcia, S.; Bosler, O.; Doucet, G. Dual character, asynaptic and synaptic, of the dopamine innervation in adult rat neostriatum: A quantitative autoradiographic and immunocytochemical analysis. J. Comp. Neurol. 1996, 375, 167–186. [Google Scholar] [CrossRef]
- DeFelipe, J.; Marco, P.; Busturia, I.; Merchán-Pérez, A. Estimation of the number of synapses in the cerebral cortex: Methodological considerations. Cereb. Cortex 1999, 9, 722–732. [Google Scholar] [CrossRef] [Green Version]
- Santuy, A.; Rodríguez, J.-R.; De Felipe, J.; Merchán-Pérez, A. Study of the size and shape of synapses in the juvenile rat somatosensory cortex with 3D electron microscopy. eNeuro 2018, 5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arbuthnott, G.W.; Wickens, J. Space, time and dopamine. Trends Neurosci. 2007, 30, 62–69. [Google Scholar] [CrossRef] [PubMed]
- Rice, M.E.; Patel, J.C.; Cragg, S.J. Dopamine release in the basal ganglia. Neuroscience 2011, 198, 112–137. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Song, C.-H.; Fan, X.; Exeter, C.J.; Hess, E.J.; Jinnah, H.A. Functional analysis of dopaminergic systems in a DYT1 knock-in mouse model of dystonia. Neurobiol. Dis. 2012, 48, 66–78. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, Y.; De Cuypere, M.; LeDoux, M.S. Abnormal motor function and dopamine neurotransmission in DYT1 ΔGAG transgenic mice. Exp. Neurol. 2008, 210, 719–730. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Augood, S.J.; Hollingsworth, Z.; Albers, D.S.; Yang, L.; Leung, J.-C.; Muller, B.; Klein, C.; Breakefield, X.O.; Standaert, D.G. Dopamine transmission in DYT1 dystonia: A biochemical and autoradiographical study. Neurology 2002, 59, 445–448. [Google Scholar] [CrossRef]
- Simonyan, K.; Berman, B.D.; Herscovitch, P.; Hallett, M. Abnormal striatal dopaminergic neurotransmission during rest and task production in spasmodic dysphonia. J. Neurosci. 2013, 33, 14705–14714. [Google Scholar] [CrossRef] [Green Version]
- Hadipour-Niktarash, A.; Rommelfanger, K.S.; Masilamoni, G.J.; Smith, Y.; Wichmann, T. Extrastriatal D2-like receptors modulate basal ganglia pathways in normal and Parkinsonian monkeys. J. Neurophysiol. 2012, 107, 1500–1512. [Google Scholar] [CrossRef] [Green Version]
- Seeman, P.; Niznik, H.B. Dopamine receptors and transporters in Parkinson’s disease and schizophrenia. FASEB J. 1990, 4, 2737–2744. [Google Scholar] [CrossRef]
- Hisahara, S.; Shimohama, S. Dopamine receptors and Parkinson’s disease. Int. J. Med. Chem. 2011, 2011, 1–16. [Google Scholar] [CrossRef]
- D’Angelo, V.; Castelli, V.; Giorgi, M.; Cardarelli, S.; Saverioni, I.; Palumbo, F.; Bonsi, P.; Pisani, A.; Giampà, C.; Sorge, R.; et al. Phosphodiesterase-10A inverse changes in striatopallidal and striatoentopeduncular pathways of a transgenic mouse model of DYT1 dystonia. J. Neurosci. 2017, 37, 2112–2124. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ferré, S.; Quiroz, C.; Woods, A.S.; Cunha, R.; Popoli, P.; Ciruela, F.; Lluis, C.; Franco, R.; Azdad, K.; Schiffmann, S.N. An update on adenosine A2A-Dopamine D2 receptor interactions. Implications for the function of G Protein-coupled receptors. Curr. Pharm. Des. 2008, 14, 1468–1474. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goodchild, R.E.; Kim, C.E.; Dauer, W.T. Loss of the dystonia-associated protein torsinA selectively disrupts the neuronal nuclear envelope. Neuron 2005, 48, 923–932. [Google Scholar] [CrossRef] [Green Version]
- Giorgi, M.; D’Angelo, V.; Esposito, Z.; Nuccetelli, V.; Sorge, R.; Martorana, A.; Stefani, A.; Bernardi, G.; Sancesario, G. Lowered cAMP and cGMP signalling in the brain during levodopa-induced dyskinesias in hemiparkinsonian rats: New aspects in the pathogenetic mechanisms. Eur. J. Neurosci. 2008, 28, 941–950. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Spots | Area µm2 | Grey level | Area µm2 | Grey Level |
| Mean | 6.73 | 252.00 | 2.4 | 216.18 |
| ± SE | 3.41 | 0.30 | 0.16 | 13.63 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
D’Angelo, V.; Paldino, E.; Cardarelli, S.; Sorge, R.; Fusco, F.R.; Biagioni, S.; Mercuri, N.B.; Giorgi, M.; Sancesario, G. Dystonia: Sparse Synapses for D2 Receptors in Striatum of a DYT1 Knock-out Mouse Model. Int. J. Mol. Sci. 2020, 21, 1073. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21031073
D’Angelo V, Paldino E, Cardarelli S, Sorge R, Fusco FR, Biagioni S, Mercuri NB, Giorgi M, Sancesario G. Dystonia: Sparse Synapses for D2 Receptors in Striatum of a DYT1 Knock-out Mouse Model. International Journal of Molecular Sciences. 2020; 21(3):1073. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21031073
Chicago/Turabian StyleD’Angelo, Vincenza, Emanuela Paldino, Silvia Cardarelli, Roberto Sorge, Francesca Romana Fusco, Stefano Biagioni, Nicola Biagio Mercuri, Mauro Giorgi, and Giuseppe Sancesario. 2020. "Dystonia: Sparse Synapses for D2 Receptors in Striatum of a DYT1 Knock-out Mouse Model" International Journal of Molecular Sciences 21, no. 3: 1073. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21031073