Potential Clinical Implications of miR-1 and miR-21 in Heart Disease and Cardioprotection †



Abstract
:1. Introduction
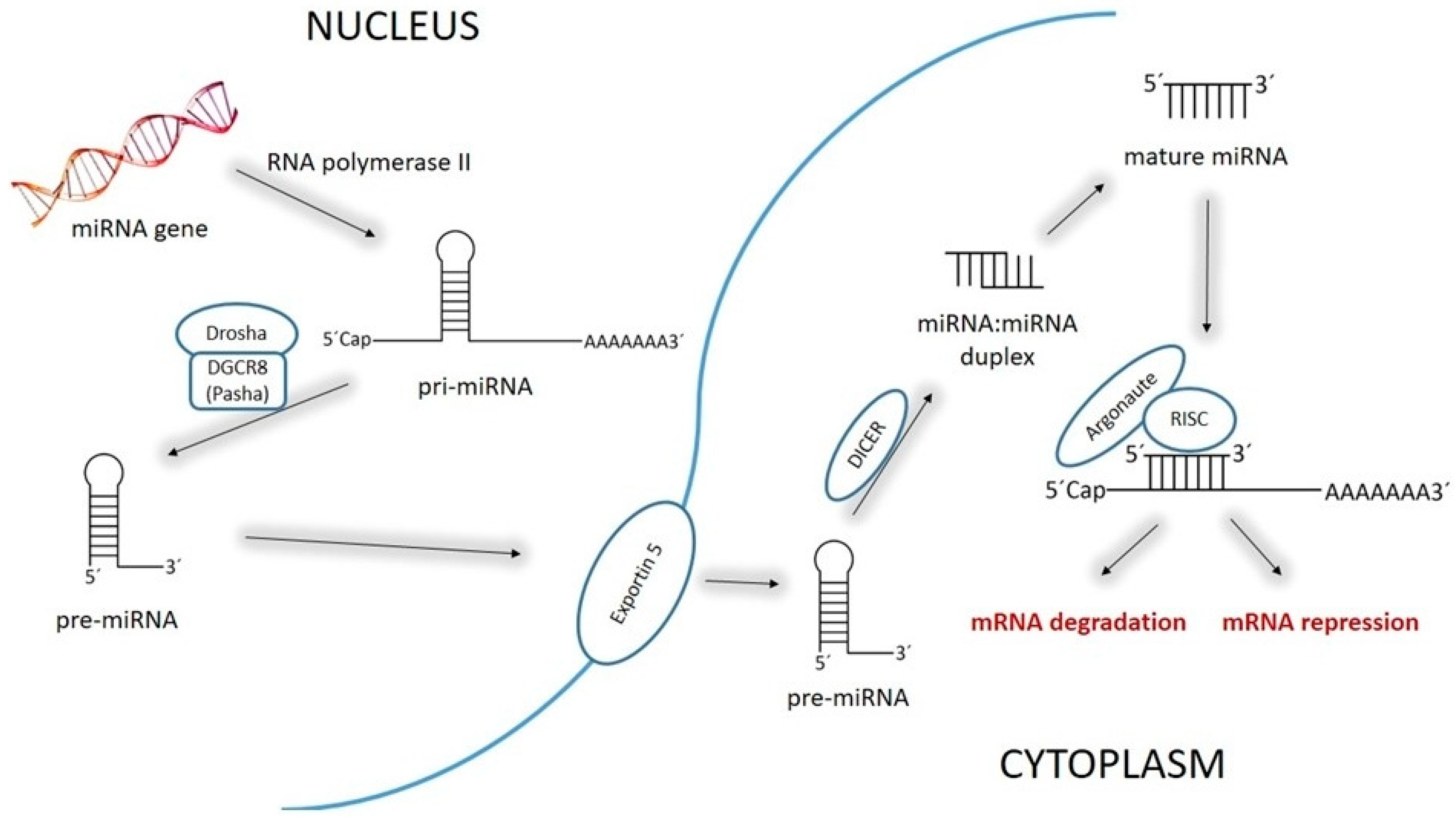
2. MiR-1 and miR-21: Discovery, Structure, Biology, and Physiological functions
3. Role of miR-1 and miR-21 in Ischemic Injury of the Heart
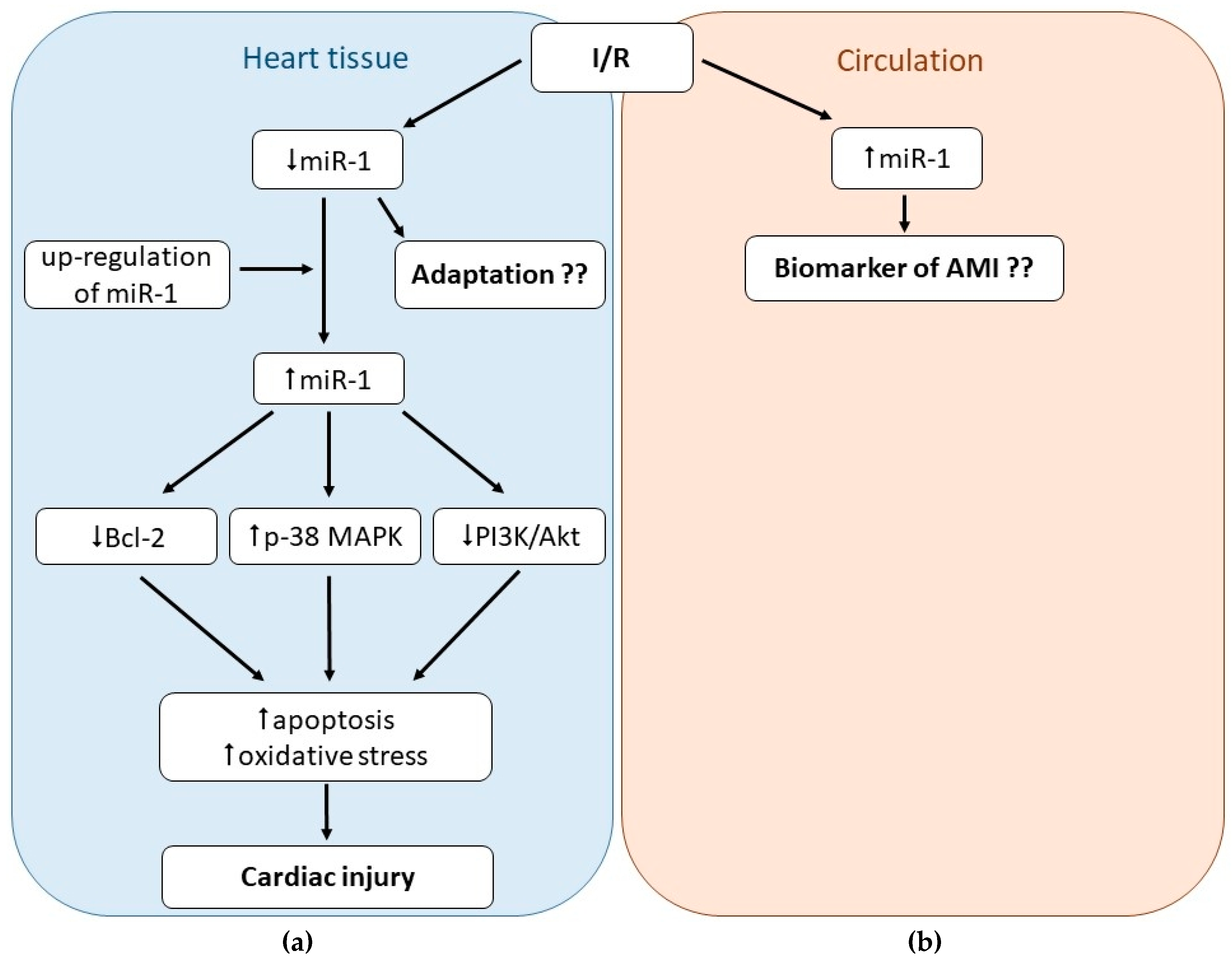
3.1. Role of miR-1 in Cardiac I/R Injury
3.2. Role of miR-21 in Cardiac I/R Injury
4. Role of miR-1 and miR-21 in Cardiac Arrhythmias
4.1. Role of miR-1 in Cardiac Arrhythmias
4.2. Role of miR-21 in Cardiac Arrhythmias
5. Role of miR-1 and miR-21 in Cardiomyopathies of Different Origin
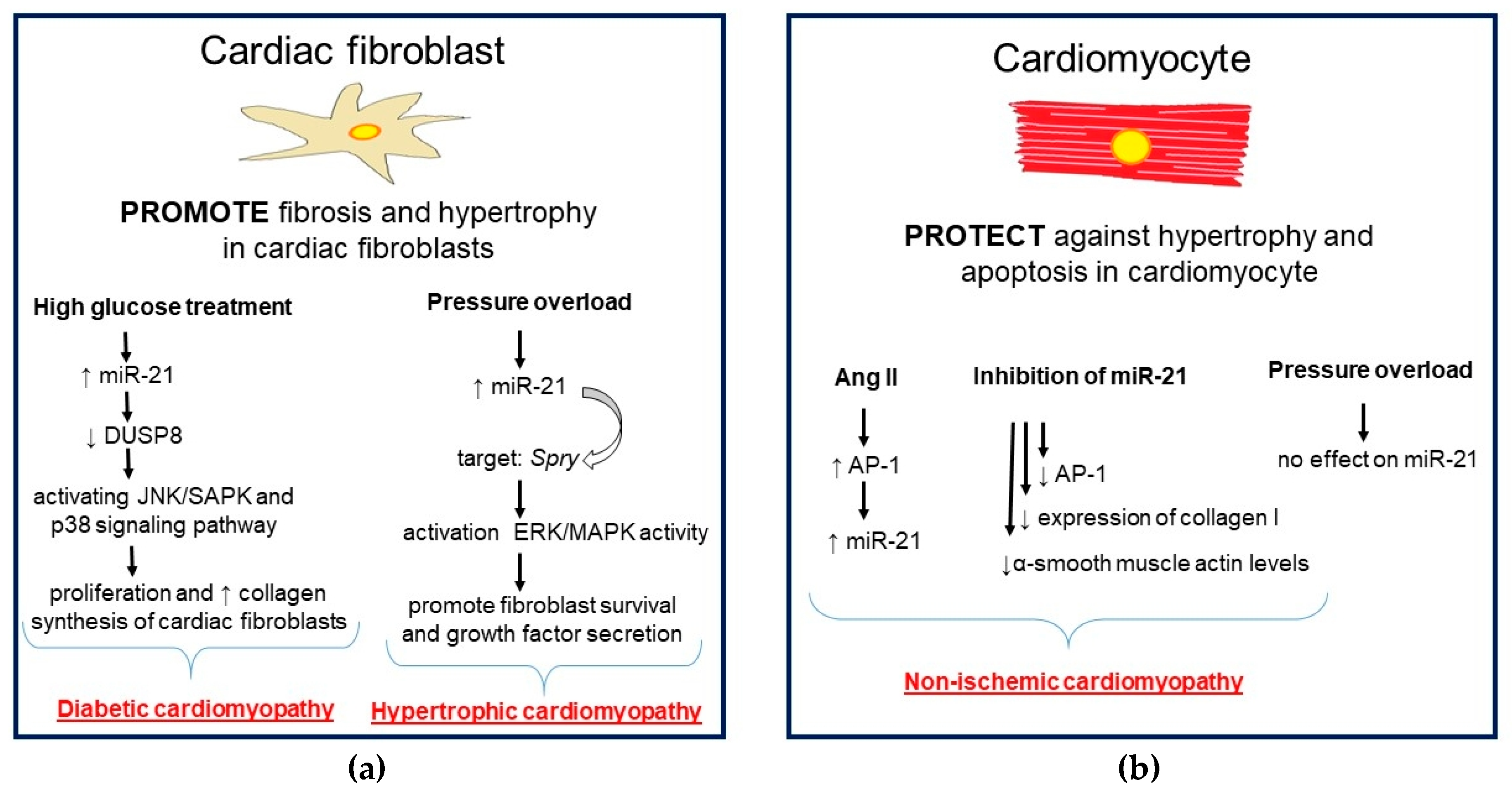
5.1. Role of miR-1 and miR-21 in Hypertrophic and Dilated Cardiomyopathy
5.2. Role of miR-1 and miR-21 in Diabetic Cardiomyopathy
5.3. Role of miR-1 and miR-21 in Viral Myocarditis
6. Role of miR-1 and miR-21 in Cardiotoxicity Induced by Cancer Treatment
6.1. Role of miR-1 and miR-21 in the Heart Injury Caused by Anthracyclines
6.2. Role of miR-1 and miR-21 in the Heart Injury Caused by Radiotherapy
7. Role of miR-1 and miR-21 in Cardioprotection
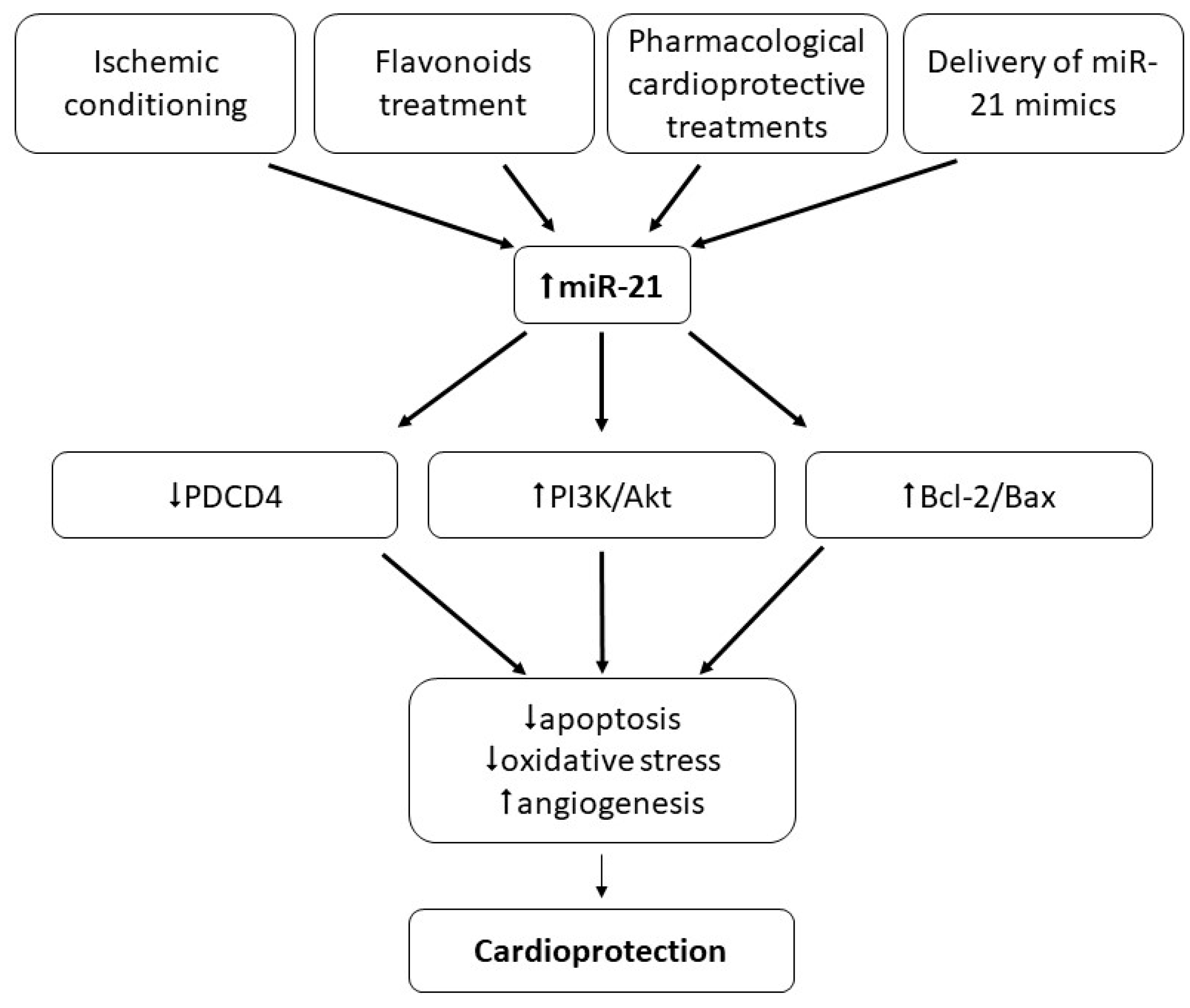
7.1. Role of miR-1 and miR-21 in Ischemic Conditioning
7.2. Role of miR-1 and miR-21 in Cardioprotection Other Than Ischemic Conditioning
8. Clinical implications of miR-1 and miR-21
9. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Devaux, Y.; Zangrando, J.; Schroen, B.; Creemers, E.E.; Pedrazzini, T.; Chang, C.P.; Dorn, G.W.; Thum, T.; Heymans, S. Long noncoding RNAs in cardiac development and ageing. Nat. Rev. Cardiol. 2015, 12, 415–425. [Google Scholar]
- Greco, S.; Somoza, A.S.; Devaux, Y.; Martelli, F. Long Noncoding RNAs and Cardiac Disease. Antioxid. Redox Signal. 2018, 29, 880–901. [Google Scholar] [CrossRef]
- Bei, Y.; Das, S.; Rodosthenous, R.S.; Holvoet, P.; Vanhaverbeke, M.; Monteiro, M.C.; Monteiro, V.V.S.; Radosinska, J.; Bartekova, M.; Jansen, F.; et al. Extracellular vesicles in cardiovascular theranostics. Theranostics 2017, 7, 4168–4182. [Google Scholar] [CrossRef]
- Bartekova, M.; Jelemensky, M.; Dhalla, N.S. Emerging role of non-coding RNAs and extracellular vesicles in cardioprotection by remote ischemic conditioning of the heart. Rev. Cardiovasc. Med. 2019, 20, 59–71. [Google Scholar] [PubMed] [Green Version]
- Gomes, C.P.C.; Spencer, H.; Ford, K.L.; Michel, L.Y.M.; Baker, A.H.; Emanueli, C.; Balligand, J.L.; Devaux, Y. The Function and Therapeutic Potential of Long Non-coding RNAs in Cardiovascular Development and Disease. Mol. Ther. Nucleic Acids 2017, 8, 494–507. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- MacFarlane, L.-A.; Murphy, P.R. MicroRNA: Biogenesis, Function and Role in Cancer. Curr. Genom. 2010, 11, 537–561. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, T.; Dong, Y.H.; Du, W.; Shi, C.Y.; Wang, K.; Tariq, M.A.; Wang, J.X.; Li, P.F. The role of microRNAs in myocardial infarction: From molecular mechanism to clinical application. Int. J. Mol. Sci. 2017, 18, 745. [Google Scholar] [CrossRef] [Green Version]
- Ruggeri, C.; Gioffré, S.; Achilli, F.; Colombo, G.I.; D’Alessandra, Y. Role of microRNAs in doxorubicin-induced cardiotoxicity: An overview of preclinical models and cancer patients. Heart Fail. Rev. 2018, 23, 109–122. [Google Scholar] [CrossRef]
- Ferreira, L.R.P.; Frade, A.F.; Santos, R.H.B.; Teixeira, P.C.; Baron, M.A.; Navarro, I.C.; Benvenuti, L.A.; Fiorelli, A.I.; Bocchi, E.A.; Stolf, N.A.; et al. MicroRNAs miR-1, miR-133a, miR-133b, miR-208a and miR-208b are dysregulated in Chronic Chagas disease Cardiomyopathy. Int. J. Cardiol. 2014, 175, 409–417. [Google Scholar] [CrossRef]
- Li, M.; Chen, X.; Chen, L.; Chen, K.; Zhou, J.; Song, J. MiR-1-3p that correlates with left ventricular function of HCM can serve as a potential target and differentiate HCM from DCM. J. Transl. Med. 2018, 16, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Huntzinger, E.; Izaurralde, E. Gene silencing by microRNAs: Contributions of translational repression and mRNA decay. Nat. Rev. Genet. 2011, 12, 99–110. [Google Scholar] [CrossRef] [PubMed]
- Bartel, D.P. MicroRNAs. Cell 2004, 116, 281–297. [Google Scholar] [CrossRef] [Green Version]
- Bartel, D.P. MicroRNAs: Target Recognition and Regulatory Functions. Cell 2009, 136, 215–233. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fabian, M.R.; Sonenberg, N.; Filipowicz, W. Regulation of mRNA Translation and Stability by microRNAs. Annu. Rev. Biochem. 2010, 79, 351–379. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Almeida, M.I.; Reis, R.M.; Calin, G.A. MicroRNA history: Discovery, recent applications, and next frontiers. Mutat. Res. Fundam. Mol. Mech. Mutagen. 2011, 717, 1–8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Filipowicz, W.; Bhattacharyya, S.N.; Sonenberg, N. Mechanisms of post-transcriptional regulation by microRNAs: Are the answers in sight? Nat. Rev. Genet. 2008, 9, 102–114. [Google Scholar] [CrossRef]
- Mathieu, J.; Ruohola-Baker, H. Regulation of Stem Cell Populations by microRNAs. Adv. Exp. Med. Biol. 2013, 786, 329–351. [Google Scholar]
- Ross, S.A.; Davis, C.D. The Emerging Role of microRNAs and Nutrition in Modulating Health and Disease. Annu. Rev. Nutr. 2014, 34, 305–336. [Google Scholar] [CrossRef]
- Lee, R.C.; Feinbaum, R.L.; Ambros, V. The C. elegans heterochronic gene lin-4 encodes small RNAs with antisense complementarity to lin-14. Cell 1993, 75, 843–854. [Google Scholar] [CrossRef]
- Condorelli, G.; Latronico, M.V.G.; Dorn, G.W. MicroRNAs in heart disease: Putative novel therapeutic targets? Eur. Heart J. 2010, 31, 649–658. [Google Scholar] [CrossRef] [Green Version]
- Siomi, H.; Siomi, M.C. Posttranscriptional Regulation of MicroRNA Biogenesis in Animals. Mol. Cell 2010, 38, 323–332. [Google Scholar] [CrossRef] [PubMed]
- Lin, S.; Gregory, R.I. MicroRNA biogenesis pathways in cancer HHS Public Access. Nat. Rev. Cancer 2015, 15, 321–333. [Google Scholar] [CrossRef] [PubMed]
- Han, C.; Yu, Z.; Duan, Z.; Kan, Q. Role of MicroRNA-1 in Human Cancer and Its Therapeutic Potentials. Biomed. Res. Int. 2014, 2014, 1–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rao, P.K.; Toyama, Y.; Chiang, H.R.; Gupta, S.; Bauer, M.; Medvid, R.; Reinhardt, F.; Liao, R.; Krieger, M.; Jaenisch, R.; et al. Loss of Cardiac microRNA-Mediated Regulation Leads to Dilated Cardiomyopathy and Heart Failure. Circ. Res. 2009, 105, 585–594. [Google Scholar] [CrossRef] [Green Version]
- Lu, T.Y.; Lin, B.; Li, Y.; Arora, A.; Han, L.; Cui, C.; Coronnello, C.; Sheng, Y.; Benos, P.V.; Yang, L. Overexpression of microRNA-1 promotes cardiomyocyte commitment from human cardiovascular progenitors via suppressing WNT and FGF signaling pathways. J. Mol. Cell. Cardiol. 2013, 63, 146–154. [Google Scholar] [CrossRef] [Green Version]
- Sluijter, J.P.G.; Van Mil, A.; Van Vliet, P.; Metz, C.H.G.; Liu, J.; Doevendans, P.A.; Goumans, M.J. MicroRNA-1 and-499 regulate differentiation and proliferation in human-derived cardiomyocyte progenitor cells. Arterioscler. Thromb. Vasc. Biol. 2010, 30, 859–868. [Google Scholar] [CrossRef] [Green Version]
- Duan, L.; Xiong, X.; Liu, Y.; Wang, J. MiRNA-1: Functional roles and dysregulation in heart disease. Mol. Biosyst. 2014, 10, 2775–2782. [Google Scholar] [CrossRef]
- Cheng, Y.; Zhang, C. MicroRNA-21 in cardiovascular disease. J. Cardiovasc. Transl. Res. 2010, 3, 251–255. [Google Scholar] [CrossRef] [Green Version]
- Kumarswamy, R.; Volkmann, I.; Thum, T. Regulation and function of miRNA-21 in health and disease. RNA Biol. 2011, 8, 706–713. [Google Scholar] [CrossRef] [Green Version]
- Chan, J.A.; Krichevsky, A.M.; Kosik, K.S. MicroRNA-21 is an antiapoptotic factor in human glioblastoma cells. Cancer Res. 2005, 65, 6029–6033. [Google Scholar] [CrossRef] [Green Version]
- Roldo, C.; Missiaglia, E.; Hagan, J.P.; Falconi, M.; Capelli, P.; Bersani, S.; Calin, G.A.; Volinia, S.; Liu, C.G.; Scarpa, A.; et al. MicroRNA expression abnormalities in pancreatic endocrine and acinar tumors are associated with distinctive pathologic features and clinical behavior. J. Clin. Oncol. 2006, 24, 4677–4684. [Google Scholar] [CrossRef] [PubMed]
- Feng, Y.-H.; Tsao, C.-J. Emerging role of microRNA-21 in cancer. Biomed. Rep. 2016, 5, 395–402. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tatsuguchi, M.; Seok, H.Y.; Callis, T.E.; Thomson, J.M.; Chen, J.F.; Newman, M.; Rojas, M.; Hammond, S.M.; Wang, D.Z. Expression of microRNAs is dynamically regulated during cardiomyocyte hypertrophy. J. Mol. Cell. Cardiol. 2007, 42, 1137–1141. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pan, X.; Wang, Z.X.; Wang, R. MicroRNA-21: A novel therapeutic target in human cancer. Cancer Biol. Ther. 2010, 10, 1224–1232. [Google Scholar] [CrossRef] [Green Version]
- Yang, H.S.; Knies, J.L.; Stark, C.; Colburn, N.H. Pdcd4 suppresses tumor phenotype in JB6 cells by inhibiting AP-1 transactivation. Oncogene 2003, 22, 3712–3720. [Google Scholar] [CrossRef] [Green Version]
- Bartekova, M.; Radosinska, J.; Pancza, D.; Barancik, M.; Ravingerova, T. Cardioprotective effects of quercetin against ischemia-reperfusion injury are age-dependent. Physiol. Res. 2016, 65 (Suppl. 1), S101–S107. [Google Scholar]
- Bartekova, M.; Radosinska, J.; Jelemensky, M.; Dhalla, N.S. Role of cytokines and inflammation in heart function during health and disease. Heart Fail. Rev. 2018, 23, 733–758. [Google Scholar] [CrossRef]
- Hughes, B.G.; Schulz, R. Targeting MMP-2 to treat ischemic heart injury. Basic Res. Cardiol. 2014, 109, 424. [Google Scholar] [CrossRef]
- Rodríguez, M.; Lucchesi, B.R.; Schaper, J. Apoptosis in myocardial infarction. Ann. Med. 2002, 34, 470–479. [Google Scholar] [CrossRef]
- Das, D.K.; Maulik, N.; Moraru, I.I. Gene expression on acute myocardial stress. Induction by hypoxia, ischemia, reperfusion, hyperthermia and oxidative stress. J. Mol. Cell. Cardiol. 1995, 27, 181–193. [Google Scholar] [CrossRef]
- Fiedler, J.; Thum, T. MicroRNAs in myocardial infarction. Arterioscler. Thromb. Vasc. Biol. 2013, 33, 201–205. [Google Scholar] [CrossRef] [Green Version]
- Sala, V.; Bergerone, S.; Gatti, S.; Gallo, S.; Ponzetto, A.; Ponzetto, C.; Crepaldi, T. MicroRNAs in myocardial ischemia: Identifying new targets and tools for treating heart disease. New frontiers for miR-medicine. Cell. Mol. Life Sci. 2014, 71, 1439–1452. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Uchida, S.; Dimmeler, S. Long noncoding RNAs in cardiovascular diseases. Circ. Res. 2015, 116, 737–750. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vausort, M.; Wagner, D.R.; Devaux, Y. Long noncoding RNAs in patients with acute myocardial infarction. Circ. Res. 2014, 115, 668–677. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Geng, H.H.; Li, R.; Su, Y.M.; Xiao, J.; Pan, M.; Cai, X.X.; Ji, X.P. The circular RNA Cdr1as promotes myocardial infarction by mediating the regulation of miR-7α on Its Target Genes Expression. PLoS ONE 2016, 11, e0151753. [Google Scholar] [CrossRef] [PubMed]
- Vausort, M.; Salgado-Somoza, A.; Zhang, L.; Leszek, P.; Scholz, M.; Teren, A.; Burkhardt, R.; Thiery, J.; Wagner, D.R.; Devaux, Y. Myocardial Infarction-Associated Circular RNA Predicting Left Ventricular Dysfunction. J. Am. Coll. Cardiol. 2016, 68, 1247–1248. [Google Scholar] [CrossRef]
- Devaux, Y.; Creemers, E.E.; Boon, R.A.; Werfel, S.; Thum, T.; Engelhardt, S.; Dimmeler, S.; Squire, I. Circular RNAs in heart failure. Eur. J. Heart Fail. 2017, 19, 701–709. [Google Scholar] [CrossRef] [Green Version]
- Zhai, C.; Tang, G.; Peng, L.; Hu, H.; Qian, G.; Wang, S.; Yao, J.; Zhang, X.; Fang, Y.; Yang, S.; et al. Inhibition of microRNA-1 attenuates hypoxia/re-oxygenation-induced apoptosis of cardiomyocytes by directly targeting Bcl-2 but not GADD45Beta. Am. J. Transl. Res. 2015, 7, 1952–1962. [Google Scholar]
- Trotta, M.C.; Ferraro, B.; Messina, A.; Panarese, I.; Gulotta, E.; Nicoletti, G.F.; D’Amico, M.; Pieretti, G. Telmisartan cardioprotects from the ischaemic/hypoxic damage through a miR-1-dependent pathway. J. Cell. Mol. Med. 2019, 23, 6635–6645. [Google Scholar] [CrossRef] [Green Version]
- Gui, Y.J.; Yang, T.; Liu, Q.; Liao, C.X.; Chen, J.Y.; Wang, Y.T.; Hu, J.H.; Xu, D.Y. Soluble epoxide hydrolase inhibitors, t-AUCB, regulated microRNA-1 and its target genes in myocardial infarction mice. Oncotarget 2017, 8, 94635–94649. [Google Scholar] [CrossRef] [Green Version]
- Kang, B.; Hong, J.; Xiao, J.; Zhu, X.; Ni, X.; Zhang, Y.; He, B.; Wang, Z. Involvement of miR-1 in the protective effect of hydrogen sulfide against cardiomyocyte apoptosis induced by ischemia/reperfusion. Mol. Biol. Rep. 2014, 41, 6845–6853. [Google Scholar] [CrossRef] [PubMed]
- Boštjančič, E.; Zidar, N.; Štajer, D.; Glavač, D. MicroRNAs miR-1, miR-133a, miR-133b and miR-208 are dysregulated in human myocardial infarction. Cardiology 2010, 115, 163–169. [Google Scholar] [CrossRef]
- Pinchi, E.; Frati, P.; Aromatario, M.; Cipolloni, L.; Fabbri, M.; La Russa, R.; Maiese, A.; Neri, M.; Santurro, A.; Scopetti, M.; et al. miR-1, miR-499 and miR-208 are sensitive markers to diagnose sudden death due to early acute myocardial infarction. J. Cell. Mol. Med. 2019, 23, 6005–6016. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boštjančič, E.; Zidar, N.; Štajer, D.; Glavač, D. MicroRNA miR-1 is up-regulated in remote myocardium in patients with myocardial infarction. Folia Biol. (Praha) 2010, 56, 27–31. [Google Scholar] [PubMed]
- Pan, Z.; Sun, X.; Ren, J.; Li, X.; Gao, X.; Lu, C.; Zhang, Y.; Sun, H.; Wang, Y.; Wang, H.; et al. miR-1 Exacerbates Cardiac Ischemia-Reperfusion Injury in Mouse Models. PLoS ONE 2012, 7, e50515. [Google Scholar] [CrossRef] [PubMed]
- Hao, Y.L.; Fang, H.C.; Zhao, H.L.; Li, X.L.; Luo, Y.; Wu, B.Q.; Fu, M.J.; Liu, W.; Liang, J.J.; Chen, X.H. The role of microRNA-1 targeting of MAPK3 in myocardial ischemia-reperfusion injury in rats undergoing sevoflurane preconditioning via the PI3K/Akt pathway. Am. J. Physiol. Cell Physiol. 2018, 315, C380–C388. [Google Scholar] [CrossRef]
- Qipshidze Kelm, N.; Piell, K.M.; Wang, E.; Cole, M.P. MicroRNAs as predictive biomarkers for myocardial injury in aged mice following myocardial infarction. J. Cell. Physiol. 2018, 233, 5214–5221. [Google Scholar] [CrossRef]
- Kuwabara, Y.; Ono, K.; Horie, T.; Nishi, H.; Nagao, K.; Kinoshita, M.; Watanabe, S.; Baba, O.; Kojima, Y.; Shizuta, S.; et al. Increased microRNA-1 and microRNA-133a levels in serum of patients with cardiovascular disease indicate myocardial damage. Circ. Cardiovasc. Genet. 2011, 4, 446–454. [Google Scholar] [CrossRef]
- Gidlöf, O.; Andersson, P.; Van Der Pals, J.; Götberg, M.; Erlinge, D. Cardiospecific microRNA plasma levels correlate with troponin and cardiac function in patients with ST elevation myocardial infarction, are selectively dependent on renal elimination, and can be detected in urine samples. Cardiology 2011, 118, 217–226. [Google Scholar] [CrossRef] [Green Version]
- Zhou, X.; Mao, A.; Wang, X.; Duan, X.; Yao, Y.; Zhang, C. Urine and Serum MicroRNA-1 as Novel Biomarkers for Myocardial Injury in Open-Heart Surgeries with Cardiopulmonary Bypass. PLoS ONE 2013, 8, e62245. [Google Scholar] [CrossRef] [Green Version]
- Goldbergova, M.P.; Ipkova, J.; Fedorko, J.; Sevcikova, J.; Parenica, J.; Spinar, J.; Masarik, M.; Vasku, A. MicroRNAs in pathophysiology of acute myocardial infarction and cardiogenic shock. Bratisl. Med. J. 2018, 119, 341–347. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dong, S.; Cheng, Y.; Yang, J.; Li, J.; Liu, X.; Wang, X.; Wang, D.; Krall, T.J.; Delphin, E.S.; Zhang, C. MicroRNA Expression Signature and the Role of MicroRNA-21 in the Early Phase of Acute Myocardial Infarction. J. Biol. Chem. 2009, 284, 29514–29525. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roy, S.; Khanna, S.; Hussain, S.R.A.; Biswas, S.; Azad, A.; Rink, C.; Gnyawali, S.; Shilo, S.; Nuovo, G.J.; Sen, C.K. MicroRNA expression in response to murine myocardial infarction: MiR-21 regulates fibroblast metalloprotease-2 via phosphatase and tensin homologue. Cardiovasc. Res. 2009, 82, 21–29. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gu, G.-L.; Xu, X.-L.; Sun, X.-T.; Zhang, J.; Guo, C.-F.; Wang, C.-S.; Sun, B.; Guo, G.-L.; Ma, K.; Huang, Y.-Y.; et al. Cardioprotective Effect of MicroRNA-21 in Murine Myocardial Infarction. Cardiovasc. Ther. 2015, 33, 109–117. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Y.; Nie, H.; Zhang, K.; Ma, D.; Yang, G.; Zheng, Z.; Liu, K.; Yu, B.; Zhai, C.; Yang, S. A feedback regulatory loop between HIF-1α and miR-21 in response to hypoxia in cardiomyocytes. FEBS Lett. 2014, 588, 3137–3146. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, C.; Li, B.; Liu, Y.; Xing, Y. Ginsenoside Rb1 protects cardiomyocytes from oxygen-glucose deprivation injuries by targeting microRNA-21. Exp. Ther. Med. 2019, 3709–3716. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qin, Y.; Yu, Y.; Dong, H.; Bian, X.; Guo, X.; Dong, S. MicroRNA 21 inhibits left ventricular remodeling in the early phase of rat model with ischemia-reperfusion injury by suppressing cell apoptosis. Int. J. Med. Sci. 2012, 9, 413–423. [Google Scholar] [CrossRef] [Green Version]
- Wang, Z.H.; Sun, X.Y.; Li, C.L.; Sun, Y.M.; Li, J.; Wang, L.F.; Li, Z.Q. miRNA-21 expression in the serum of elderly patients with acute myocardial infarction. Med. Sci. Monit. 2017, 23, 5728–5734. [Google Scholar] [CrossRef] [Green Version]
- Chen, C.-H.; Hsu, S.-Y.; Chiu, C.-C.; Leu, S. MicroRNA-21 Mediates the Protective Effect of Cardiomyocyte-Derived Conditioned Medium on Ameliorating Myocardial Infarction in Rats. Cells 2019, 8, 935. [Google Scholar] [CrossRef] [Green Version]
- Xiao, J.; Pan, Y.; Li, X.H.; Yang, X.Y.; Feng, Y.L.; Tan, H.H.; Jiang, L.; Feng, J.; Yu, X.Y. Cardiac progenitor cell-derived exosomes prevent cardiomyocytes apoptosis through exosomal miR-21 by targeting PDCD4. Cell Death Dis. 2016, 7, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Yuan, J.; Chen, H.; Ge, D.; Xu, Y.; Xu, H.; Yang, Y.; Gu, M.; Zhou, Y.; Zhu, J.; Ge, T.; et al. Mir-21 Promotes Cardiac Fibrosis after Myocardial Infarction Via Targeting Smad7. Cell. Physiol. Biochem. 2017, 42, 2207–2219. [Google Scholar] [CrossRef] [PubMed]
- Adam, O.; Löhfelm, B.; Thum, T.; Gupta, S.K.; Puhl, S.L.; Schäfers, H.J.; Böhm, M.; Laufs, U. Role of miR-21 in the pathogenesis of atrial fibrosis. Basic Res. Cardiol. 2012, 107, 278. [Google Scholar] [CrossRef] [PubMed]
- Cardin, S.; Guasch, E.; Luo, X.; Naud, P.; Le Quang, K.; Shi, Y.F.; Tardif, J.C.; Comtois, P.; Nattel, S. Role for MicroRNA-21 in atrial profibrillatory fibrotic remodeling associated with experimental postinfarction heart failure. Circ. Arrhythm. Electrophysiol. 2012, 5, 1027–1035. [Google Scholar] [CrossRef] [Green Version]
- Zhou, X.; Xu, H.; Liu, Z.; Wu, Q.; Zhu, R.; Liu, J. miR-21 promotes cardiac fibroblast-to-myofibroblast transformation and myocardial fibrosis by targeting Jagged1. J. Cell. Mol. Med. 2018, 22, 3816–3824. [Google Scholar] [CrossRef]
- Bejerano, T.; Etzion, S.; Elyagon, S.; Etzion, Y.; Cohen, S. Nanoparticle Delivery of miRNA-21 Mimic to Cardiac Macrophages Improves Myocardial Remodeling after Myocardial Infarction. Nano Lett. 2018, 18, 5885–5891. [Google Scholar] [CrossRef] [PubMed]
- Zile, M.R.; Mehurg, S.M.; Arroyo, J.E.; Stroud, R.E.; DeSantis, S.M.; Spinale, F.G. Relationship between the temporal profile of plasma microRNA and left ventricular remodeling in patients after myocardial infarction. Circ. Cardiovasc. Genet. 2011, 4, 614–619. [Google Scholar] [CrossRef] [Green Version]
- Jansen, F.; Schäfer, L.; Wang, H.; Schmitz, T.; Flender, A.; Schueler, R.; Hammerstingl, C.; Nickenig, G.; Sinning, J.M.; Werner, N. Kinetics of circulating microRNAs in response to cardiac stress in patients with coronary artery disease. J. Am. Heart Assoc. 2017, 6, 1–11. [Google Scholar] [CrossRef]
- Liebetrau, C.; Möllmann, H.; Dörr, O.; Szardien, S.; Troidl, C.; Willmer, M.; Voss, S.; Gaede, L.; Rixe, J.; Rolf, A.; et al. Release kinetics of circulating muscle-enriched microRNAs in patients undergoing transcoronary ablation of septal hypertrophy. J. Am. Coll. Cardiol. 2013, 62, 992–998. [Google Scholar] [CrossRef] [Green Version]
- Robinson, S.; Follo, M.; Haenel, D.; Mauler, M.; Stallmann, D.; Heger, L.A.; Helbing, T.; Duerschmied, D.; Peter, K.; Bode, C.; et al. Chip-based digital PCR as a novel detection method for quantifying microRNAs in acute myocardial infarction patients article. Acta Pharmacol. Sin. 2018, 39, 1217–1227. [Google Scholar] [CrossRef] [Green Version]
- Robinson, S.; Follo, M.; Haenel, D.; Mauler, M.; Stallmann, D.; Tewari, M.; Duerschmied, D.; Peter, K.; Bode, C.; Ahrens, I.; et al. Droplet digital PCR as a novel detection method for quantifying microRNAs in acute myocardial infarction. Int. J. Cardiol. 2018, 257, 247–254. [Google Scholar] [CrossRef]
- Hortmann, M.; Walter, J.E.; Benning, L.; Follo, M.; Mayr, R.M.; Honegger, U.; Robinson, S.; Stallmann, D.; Duerschmied, D.; Twerenbold, R.; et al. Droplet digital PCR of serum miR-499, miR-21 and miR-208a for the detection of functionally relevant coronary artery disease. Int. J. Cardiol. 2019, 275, 129–135. [Google Scholar] [CrossRef] [PubMed]
- Byrum, S.D.; Burdine, M.S.; Orr, L.; Mackintosh, S.G.; Authier, S.; Pouliot, M.; Hauer-Jensen, M.; Tackett, A.J. Time-and radiation-dose dependent changes in the plasma proteome after total body irradiation of non-human primates: Implications for biomarker selection. PLoS ONE 2017, 12, e0174771. [Google Scholar] [CrossRef] [PubMed]
- Cheng, Y.; Zhu, P.; Yang, J.; Liu, X.; Dong, S.; Wang, X.; Chun, B.; Zhuang, J.; Zhang, C. Ischaemic preconditioning-regulated miR-21 protects heart against ischaemia/reperfusion injury via anti-apoptosis through its target PDCD4. Cardiovasc. Res. 2010, 87, 431–439. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, B.; Lin, H.; Xiao, J.; Lu, Y.; Luo, X.; Li, B.; Zhang, Y.; Xu, C.; Bai, Y.; Wang, H.; et al. The muscle-specific microRNA miR-1 regulates cardiac arrhythmogenic potential by targeting GJA1 and KCNJ2. Nat. Med. 2007, 13, 486–491. [Google Scholar] [CrossRef]
- Belevych, A.E.; Sansom, S.E.; Terentyeva, R.; Ho, H.-T.; Nishijima, Y.; Martin, M.M.; Jindal, H.K.; Rochira, J.A.; Kunitomo, Y.; Abdellatif, M.; et al. MicroRNA-1 and -133 Increase Arrhythmogenesis in Heart Failure by Dissociating Phosphatase Activity from RyR2 Complex. PLoS ONE 2011, 6, e28324. [Google Scholar] [CrossRef]
- Zhang, Y.; Sun, L.; Zhang, Y.; Liang, H.; Li, X.; Cai, R.; Wang, L.; Du, W.; Zhang, R.; Li, J.; et al. Overexpression of microRNA-1 Causes Atrioventricular Block in Rodents. Int. J. Biol. Sci. 2013, 9, 455–462. [Google Scholar] [CrossRef] [Green Version]
- Tsoporis, J.N.; Fazio, A.; Rizos, I.K.; Izhar, S.; Proteau, G.; Salpeas, V.; Rigopoulos, A.; Sakadakis, E.; Toumpoulis, I.K.; Parker, T.G. Increased right atrial appendage apoptosis is associated with differential regulation of candidate MicroRNAs 1 and 133A in patients who developed atrial fibrillation after cardiac surgery. J. Mol. Cell. Cardiol. 2018, 121, 25–32. [Google Scholar] [CrossRef]
- Li, Y.-D.; Hong, Y.-F.; Yusufuaji, Y.; Tang, B.-P.; Zhou, X.-H.; Xu, G.-J.; Li, J.-X.; Sun, L.; Zhang, J.-H.; Xin, Q.; et al. Altered expression of hyperpolarization-activated cyclic nucleotide-gated channels and microRNA-1 and -133 in patients with age-associated atrial fibrillation. Mol. Med. Rep. 2015, 12, 3243–3248. [Google Scholar] [CrossRef]
- Girmatsion, Z.; Biliczki, P.; Bonauer, A.; Wimmer-Greinecker, G.; Scherer, M.; Moritz, A.; Bukowska, A.; Goette, A.; Nattel, S.; Hohnloser, S.H.; et al. Changes in microRNA-1 expression and IK1 up-regulation in human atrial fibrillation. Hear. Rhythm 2009, 6, 1802–1809. [Google Scholar] [CrossRef]
- Barana, A.; Matamoros, M.; Dolz-Gaitón, P.; Pérez-Hernández, M.; Amorós, I.; Núñez, M.; Sacristán, S.; Pedraz, Á.; Pinto, Á.; Fernández-Avilés, F.; et al. Chronic Atrial Fibrillation Increases MicroRNA-21 in Human Atrial Myocytes Decreasing L-Type Calcium Current. Circ. Arrhythm. Electrophysiol. 2014, 7, 861–868. [Google Scholar] [CrossRef] [Green Version]
- McManus, D.D.; Tanriverdi, K.; Lin, H.; Esa, N.; Kinno, M.; Mandapati, D.; Tam, S.; Okike, O.N.; Ellinor, P.T.; Keaney, J.F.; et al. Plasma microRNAs are associated with atrial fibrillation and change after catheter ablation (the miRhythm study). Hear. Rhythm 2015, 12, 3–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Adam, O.; Zimmer, C.; Hanke, N.; Hartmann, R.W.; Klemmer, B.; Böhm, M.; Laufs, U. Inhibition of aldosterone synthase (CYP11B2) by torasemide prevents atrial fibrosis and atrial fibrillation in mice. J. Mol. Cell. Cardiol. 2015, 85, 140–150. [Google Scholar] [CrossRef] [PubMed]
- He, X.; Zhang, K.; Gao, X.; Li, L.; Tan, H.; Chen, J.; Zhou, Y. Rapid atrial pacing induces myocardial fibrosis by down-regulating Smad7 via microRNA-21 in rabbit. Heart Vessel. 2016, 31, 1696–1708. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, Z.; Chen, X.; Qian, C.; Dong, Q.; Ding, D.; Wu, Q.; Li, J.; Wang, H.; Li, W.; Xie, Q.; et al. Signal Transducer and Activator of Transcription 3/MicroRNA-21 Feedback Loop Contributes to Atrial Fibrillation by Promoting Atrial Fibrosis in a Rat Sterile Pericarditis Model. Circ. Arrhythm. Electrophysiol. 2016, 9, e003396. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, K.; Narumi, T.; Watanabe, T.; Aono, T.; Goto, J.; Sugai, T.; Toshima, T.; Kato, S.; Tamura, H.; Nishiyama, S.; et al. P1626MicroRNA-21 deteriorates left ventricular reverse remodeling by promoting cardiac fibrosis in non-ischemic cardiomyopathy. Eur. Heart J. 2019, 40, 9–11. [Google Scholar] [CrossRef]
- Wei, Y.; Peng, S.; Wu, M.; Sachidanandam, R.; Tu, Z.; Zhang, S.; Falce, C.; Sobie, E.A.; Lebeche, D.; Zhao, Y. Multifaceted roles of miR-1s in repressing the fetal gene program in the heart. Cell Res. 2014, 24, 278–292. [Google Scholar] [CrossRef] [Green Version]
- Thum, T.; Gross, C.; Fiedler, J.; Fischer, T.; Kissler, S.; Bussen, M.; Galuppo, P.; Just, S.; Rottbauer, W.; Frantz, S.; et al. MicroRNA-21 contributes to myocardial disease by stimulating MAP kinase signalling in fibroblasts. Nature 2008, 456, 980–984. [Google Scholar] [CrossRef]
- Shan, Z.-X.; Lin, Q.-X.; Deng, C.-Y.; Zhu, J.-N.; Mai, L.-P.; Liu, J.-L.; Fu, Y.-H.; Liu, X.-Y.; Li, Y.-X.; Zhang, Y.-Y.; et al. miR-1/miR-206 regulate Hsp60 expression contributing to glucose-mediated apoptosis in cardiomyocytes. FEBS Lett. 2010, 584, 3592–3600. [Google Scholar] [CrossRef] [Green Version]
- Yildirim, S.S.; Akman, D.; Catalucci, D.; Turan, B. Relationship Between Downregulation of miRNAs and Increase of Oxidative Stress in the Development of Diabetic Cardiac Dysfunction: Junctin as a Target Protein of miR-1. Cell Biochem. Biophys. 2013, 67, 1397–1408. [Google Scholar] [CrossRef]
- Liu, S.; Li, W.; Xu, M.; Huang, H.; Wang, J.; Chen, X. Micro-RNA 21Targets DUSP 8 to Promote Collagen Synthesis in High Glucose-Treated Primary Cardiac Fibroblasts. Can. J. Cardiol. 2014, 30, 1689–1699. [Google Scholar] [CrossRef]
- Corsten, M.F.; Papageorgiou, A.; Verhesen, W.; Carai, P.; Lindow, M.; Obad, S.; Summer, G.; Coort, S.L.M.; Hazebroek, M.; van Leeuwen, R.; et al. MicroRNA Profiling Identifies MicroRNA-155 as an Adverse Mediator of Cardiac Injury and Dysfunction During Acute Viral Myocarditis. Circ. Res. 2012, 111, 415–425. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Y.L.; Wu, W.; Xue, Y.; Gao, M.; Yan, Y.; Kong, Q.; Pang, Y.; Yang, F. MicroRNA-21 and -146b are involved in the pathogenesis of murine viral myocarditis by regulating TH-17 differentiation. Arch. Virol. 2013, 158, 1953–1963. [Google Scholar] [CrossRef] [PubMed]
- Kura, B.; Yin, C.; Frimmel, K.; Krizak, J.; Okruhlicova, L.; Kukreja, R.C.; Slezak, J. Changes of microRNA-1, -15b and -21 levels in irradiated rat hearts after treatment with potentially radioprotective drugs. Physiol. Res. 2016, 65 (Suppl. 1), S129–S137. [Google Scholar]
- Kura, B.; Kalocayova, B.; LeBaron, T.W.; Frimmel, K.; Buday, J.; Surovy, J.; Slezak, J. Regulation of microRNAs by molecular hydrogen contributes to the prevention of radiation-induced damage in the rat myocardium. Mol. Cell. Biochem. 2019, 457, 61–72. [Google Scholar] [CrossRef] [PubMed]
- Nishimura, Y.; Kondo, C.; Morikawa, Y.; Tonomura, Y.; Torii, M.; Yamate, J.; Uehara, T. Plasma miR-208 as a useful biomarker for drug-induced cardiotoxicity in rats. J. Appl. Toxicol. 2015, 35, 173–180. [Google Scholar] [CrossRef] [PubMed]
- Rigaud, V.O.C.; Ferreira, L.R.P.; Ayub-Ferreira, S.M.; ávila, M.S.; Brandão, S.M.G.; Cruz, F.D.; Santos, M.H.H.; Cruz, C.B.B.V.; Alves, M.S.L.; Issa, V.S.; et al. Circulating miR-1 as a potential biomarker of doxorubicininduced cardiotoxicity in breast cancer patients. Oncotarget 2017, 8, 6994–7002. [Google Scholar] [CrossRef] [Green Version]
- Ruggeri, C.; Gioffré, S.; Chiesa, M.; Buzzetti, M.; Milano, G.; Scopece, A.; Castiglioni, L.; Pontremoli, M.; Sironi, L.; Pompilio, G.; et al. A specific circulating MicroRNA cluster is associated to late differential cardiac response to doxorubicin-induced cardiotoxicity in vivo. Dis. Markers 2018, 2018. [Google Scholar] [CrossRef] [Green Version]
- Razavi-Azarkhiavi, K.; Jaafari, M.R.; Abnous, K.; Razavi, B.M.; Jafarian, A.H.; Hassani, F.V.; Shirani, K.; Karimi, G. The Cardiotoxic Mechanism of Doxorubicin (DOX) and Pegylated Liposomal DOX in Mice Bearing C-26 Colon Carcinoma: A Study Focused on microRNA Role for Toxicity Assessment of New Formulations. Pharm. Res. 2017, 34, 1849–1856. [Google Scholar] [CrossRef]
- Leger, K.J.; Leonard, D.; Nielson, D.; de Lemos, J.A.; Mammen, P.P.A.; Winick, N.J. Circulating microRNAs: Potential Markers of Cardiotoxicity in Children and Young Adults Treated With Anthracycline Chemotherapy. J. Am. Heart Assoc. 2017, 6, e004653. [Google Scholar] [CrossRef]
- Todorova, V.K.; Makhoul, I.; Wei, J.; Klimberg, V.S. Circulating miRNA profiles of doxorubicin-induced cardiotoxicity in breast cancer patients. Ann. Clin. Lab. Sci. 2017, 47, 115–119. [Google Scholar]
- Desai, V.G.; Kwekel, J.C.; Vijay, V.; Moland, C.L.; Herman, E.H.; Lee, T.; Han, T.; Lewis, S.M.; Davis, K.J.; Muskhelishvili, L.; et al. Early biomarkers of doxorubicin-induced heart injury in a mouse model. Toxicol. Appl. Pharmacol. 2014, 281, 221–229. [Google Scholar] [CrossRef] [PubMed]
- Tong, Z.; Jiang, B.; Wu, Y.; Liu, Y.; Li, Y.; Gao, M.; Jiang, Y.; Lv, Q.; Xiao, X. MiR-21 protected cardiomyocytes against doxorubicin-induced apoptosis by targeting BTG2. Int. J. Mol. Sci. 2015, 16, 14511–14525. [Google Scholar] [CrossRef] [PubMed]
- Fu, D. Cardiac Arrhythmias: Diagnosis, Symptoms, and Treatments. Cell Biochem. Biophys. 2015, 73, 291–296. [Google Scholar] [CrossRef] [PubMed]
- Terentyev, D.; Belevych, A.E.; Terentyeva, R.; Martin, M.M.; Malana, G.E.; Kuhn, D.E.; Abdellatif, M.; Feldman, D.S.; Elton, T.S.; Györke, S. miR-1 Overexpression Enhances Ca2+ Release and Promotes Cardiac Arrhythmogenesis by Targeting PP2A Regulatory Subunit B56α and Causing CaMKII-Dependent Hyperphosphorylation of RyR2. Circ. Res. 2009, 104, 514–521. [Google Scholar] [CrossRef] [Green Version]
- Jia, X.; Zheng, S.; Xie, X.; Zhang, Y.; Wang, W.; Wang, Z.; Zhang, Y.; Wang, J.; Gao, M.; Hou, Y. MicroRNA-1 Accelerates the Shortening of Atrial Effective Refractory Period by Regulating KCNE1 and KCNB2 Expression: An Atrial Tachypacing Rabbit Model. PLoS ONE 2013, 8, e85639. [Google Scholar] [CrossRef] [Green Version]
- Su, X.; Liang, H.; Wang, H.; Chen, G.; Jiang, H.; Wu, Q.; Liu, T.; Liu, Q.; Yu, T.; Gu, Y.; et al. Over-expression of microRNA-1 causes arrhythmia by disturbing intracellular trafficking system. Sci. Rep. 2017, 7, 46259. [Google Scholar] [CrossRef] [Green Version]
- Yu, H.D.; Xia, S.; Zha, C.Q.; Deng, S.B.; Du, J.L.; She, Q. Spironolactone Regulates HCN Protein Expression Through Micro-RNA-1 in Rats with Myocardial Infarction. J. Cardiovasc. Pharmacol. 2015, 65, 587–592. [Google Scholar] [CrossRef] [Green Version]
- Torrado, M.; Franco, D.; Lozano-Velasco, E.; Hernández-Torres, F.; Calviño, R.; Aldama, G.; Centeno, A.; Castro-Beiras, A.; Mikhailov, A. A MicroRNA-Transcription Factor Blueprint for Early Atrial Arrhythmogenic Remodeling. Biomed. Res. Int. 2015, 2015, 1–13. [Google Scholar] [CrossRef]
- Tao, H.; Zhang, M.; Yang, J.-J.; Shi, K.-H. MicroRNA-21 via Dysregulation of WW Domain-Containing Protein 1 Regulate Atrial Fibrosis in Atrial Fibrillation. Hear. Lung Circ. 2018, 27, 104–113. [Google Scholar] [CrossRef]
- Szemraj-Rogucka, Z.M.; Szemraj, J.; Masiarek, K.; Majos, A. Circulating microRNAs as biomarkers for myocardial fibrosis in patients with left ventricular non-compaction cardiomyopathy. Arch. Med. Sci. 2019, 15, 376–384. [Google Scholar] [CrossRef]
- Wexler, R.K.; Elton, T.; Pleister, A.; Feldman, D. Cardiomyopathy: An overview. Am. Fam. Physician 2009, 79, 778–784. [Google Scholar] [PubMed]
- Li, H.; Chen, C.; Fan, J.; Yin, Z.; Ni, L.; Cianflone, K.; Wang, Y.; Wang, D.W. Identification of cardiac long non-coding RNA profile in human dilated cardiomyopathy. Cardiovasc. Res. 2018, 114, 747–758. [Google Scholar] [CrossRef] [PubMed]
- Narula, J.; Haider, N.; Virmani, R.; DiSalvo, T.G.; Kolodgie, F.D.; Hajjar, R.J.; Schmidt, U.; Semigran, M.J.; Dec, G.W.; Khaw, B.-A. Apoptosis in Myocytes in End-Stage Heart Failure. N. Engl. J. Med. 1996, 335, 1182–1189. [Google Scholar] [CrossRef] [PubMed]
- Lakdawala, N.K.; Winterfield, J.R.; Funke, B.H. Dilated Cardiomyopathy. Circ. Arrhythm. Electrophysiol. 2013, 6, 228–237. [Google Scholar] [CrossRef] [Green Version]
- Morkin, E. Control of cardiac myosin heavy chain gene expression. Microsc. Res. Tech. 2000, 50, 522–531. [Google Scholar] [CrossRef]
- Guo, R.; Nair, S. Role of microRNA in diabetic cardiomyopathy: From mechanism to intervention. Biochim. Biophys. Acta Mol. Basis Dis. 2017, 1863, 2070–2077. [Google Scholar] [CrossRef]
- Tabák, A.G.; Herder, C.; Rathmann, W.; Brunner, E.J.; Kivimäki, M. Prediabetes: A high-risk state for diabetes development. Lancet 2012, 379, 2279–2290. [Google Scholar] [CrossRef] [Green Version]
- Shen, E.; Diao, X.; Wang, X.; Chen, R.; Hu, B. MicroRNAs Involved in the Mitogen-Activated Protein Kinase Cascades Pathway During Glucose-Induced Cardiomyocyte Hypertrophy. Am. J. Pathol. 2011, 179, 639–650. [Google Scholar] [CrossRef]
- Cheng, Y.; Zhao, W.; Zhang, X.; Sun, L.; Yang, H.; Wang, Y.; Cao, Y.; Chu, Y.; Liu, G. Downregulation of microRNA-1 attenuates glucose-induced apoptosis by regulating the liver X receptor α in cardiomyocytes. Exp. Ther. Med. 2018, 16, 1814–1824. [Google Scholar] [CrossRef]
- Lopes, M.B.; Freitas, R.C.C.; Hirata, M.H.; Hirata, R.D.C.; Rezende, A.A.; Silbiger, V.N.; Bortolin, R.H.; Luchessi, A.D. MRNA-miRNA integrative analysis of diabetes-induced cardiomyopathy in rats. Front. Biosci. Sch. 2017, 9, 194–229. [Google Scholar]
- Wong, L.L.; Wang, J.; Liew, O.W.; Richards, A.M.; Chen, Y.T. MicroRNA and heart failure. Int. J. Mol. Sci. 2016, 17, 502. [Google Scholar] [CrossRef] [PubMed]
- Dai, B.; Li, H.; Fan, J.; Zhao, Y.; Yin, Z.; Nie, X.; Wang, D.W.; Chen, C. MiR-21 protected against diabetic cardiomyopathy induced diastolic dysfunction by targeting gelsolin. Cardiovasc. Diabetol. 2018, 17, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Fung, G.; Luo, H.; Qiu, Y.; Yang, D.; McManus, B. Myocarditis. Circ. Res. 2016, 118, 496–514. [Google Scholar] [CrossRef] [PubMed]
- Pollack, A.; Kontorovich, A.R.; Fuster, V.; Dec, G.W. Viral myocarditis-diagnosis, treatment options, and current controversies. Nat. Rev. Cardiol. 2015, 12, 670–680. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Li, J.; Xuan, L.; Liu, Y.; Shao, L.; Ge, H.; Gu, J.; Wei, C.; Zhao, M. Astragalus Root dry extract restores connexin43 expression by targeting miR-1 in viral myocarditis. Phytomedicine 2018, 46, 32–38. [Google Scholar] [CrossRef] [PubMed]
- Kura, B.; Babal, P.; Slezak, J. Implication of microRNAs in the development and potential treatment of radiation-induced heart disease. Can. J. Physiol. Pharmacol. 2017, 95, 1236–1244. [Google Scholar] [CrossRef]
- Binaschi, M.; Capranico, G.; Dal Bo, L.; Zunino, F. Relationship between lethal effects and topoisomerase II-mediated double-stranded DNA breaks produced by anthracyclines with different sequence specificity. Mol. Pharmacol. 1997, 51, 1053–1059. [Google Scholar] [CrossRef]
- Jensen, B.V.; Skovsgaard, T.; Nielsen, S.L. Functional monitoring of anthracycline cardiotoxicity: A prospective, blinded, long-term observational study of outcome in 120 patients. Ann. Oncol. 2002, 13, 699–709. [Google Scholar] [CrossRef]
- Koleini, N.; Kardami, E. Autophagy and mitophagy in the context of doxorubicin-induced cardiotoxicity. Oncotarget 2017, 8, 46663–46680. [Google Scholar] [CrossRef] [Green Version]
- Slezak, J.; Kura, B.; Ravingerová, T.; Tribulova, N.; Okruhlicova, L.; Barancik, M. Mechanisms of cardiac radiation injury and potential preventive approaches. Can. J. Physiol. Pharmacol. 2015, 93, 737–753. [Google Scholar] [CrossRef]
- Slezak, J.; Kura, B.; Babal, P.; Barancik, M.; Ferko, M.; Frimmel, K.; Kalocayova, B.; Kukreja, R.C.; Lazou, A.; Mezesova, L.; et al. Potential markers and metabolic processes involved in the mechanism of radiation-induced heart injury. Can. J. Physiol. Pharmacol. 2017, 95, 1190–1203. [Google Scholar] [CrossRef] [PubMed]
- Michalowski, A.S. On radiation damage to normal tissues and its treatment: II. Anti-inflammatory drugs. Acta Oncol. 1994, 33, 139–157. [Google Scholar] [CrossRef] [PubMed]
- McDonald, K. Diastolic heart failure in the elderly: Underlying mechanisms and clinical relevance. Int. J. Cardiol. 2008, 125, 197–202. [Google Scholar] [CrossRef] [PubMed]
- Chello, M.; Mastroroberto, P.; Romano, R.; Zofrea, S.; Bevacqua, I.; Marchese, A.R. Changes in the proportion of types I and III collagen in the left ventricular wall of patients with post-irradiative pericarditis. Vascular 1996, 4, 222–226. [Google Scholar] [CrossRef]
- Constine, L.S.; Schwartz, R.G.; Savage, D.E.; King, V.; Muhs, A. Cardiac function, perfusion, and morbidity in irradiated long-term survivors of Hodgkin’s disease. Int. J. Radiat. Oncol. Biol. Phys. 1997, 39, 897–906. [Google Scholar] [CrossRef]
- Robert Stewart, J.; Fajardo, L.F. Radiation-induced heart disease: An update. Prog. Cardiovasc. Dis. 1984, 27, 173–194. [Google Scholar] [CrossRef]
- Barancik, M.; Okruhlicova, L.; Fogarassyova, M.; Bartekova, M.; Slezak, J. Mediastinal irradiation modulates myocardial and circulating matrix metalloproteinases. Exp. Clin. Cardiol. 2013, 18, 37–40. [Google Scholar]
- Viczenczova, C.; Bacova, B.S.; Benova, T.E.; Kura, B.; Yin, C.; Weismann, P.; Kukreja, R.; Slezak, J.; Tribulova, N. Myocardial connexin-43 and PKC signalling are involved in adaptation of the heart to irradiation-induced injury: Implication of miR-1 and miR-21. Gen. Physiol. Biophys. 2016, 35, 215–222. [Google Scholar] [CrossRef] [Green Version]
- Frey, N.; Katus, H.A.; Olson, E.N.; Hill, J.A. Hypertrophy of the Heart: A New Therapeutic Target? Circulation 2004, 109, 1580–1589. [Google Scholar] [CrossRef] [Green Version]
- Surova, O.; Akbar, N.S.; Zhivotovsky, B. Knock-down of core proteins regulating microrna biogenesis has no effect on sensitivity of lung cancer cells to ionizing radiation. PLoS ONE 2012, 7, e33134. [Google Scholar] [CrossRef] [Green Version]
- Kraemer, A.; Anastasov, N.; Angermeier, M.; Winkler, K.; Atkinson, M.J.; Moertl, S. MicroRNA-Mediated Processes are Essential for the Cellular Radiation Response. Radiat. Res. 2011, 176, 575. [Google Scholar] [CrossRef] [PubMed]
- Simone, N.L.; Soule, B.P.; Ly, D.; Saleh, A.D.; Savage, J.E.; DeGraff, W.; Cook, J.; Harris, C.C.; Gius, D.; Mitchell, J.B. Ionizing radiation-induced oxidative stress alters miRNA expression. PLoS ONE 2009, 4, e6377. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ikeda, S.; He, A.; Kong, S.W.; Lu, J.; Bejar, R.; Bodyak, N.; Lee, K.-H.; Ma, Q.; Kang, P.M.; Golub, T.R.; et al. MicroRNA-1 Negatively Regulates Expression of the Hypertrophy-Associated Calmodulin and Mef2a Genes. Mol. Cell. Biol. 2009, 29, 2193–2204. [Google Scholar] [CrossRef] [Green Version]
- Sayed, D.; Hong, C.; Chen, I.Y.; Lypowy, J.; Abdellatif, M. MicroRNAs play an essential role in the development of cardiac hypertrophy. Circ. Res. 2007, 100, 416–424. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Salloum, F.N.; Yin, C.; Kukreja, R.C. Role of miRs in Cardiac Preconditioning. J. Cardiovasc. Pharmacol. 2010, 56, 581–588. [Google Scholar] [CrossRef] [Green Version]
- Yin, C.; Salloum, F.N.; Kukreja, R.C. A novel role of microRNA in late preconditioning: Upregulation of endothelial nitric oxide synthase and heat shock protein 70. Circ. Res. 2009, 104, 572–575. [Google Scholar] [CrossRef]
- Duan, X.; Ji, B.; Wang, X.; Liu, J.; Zheng, Z.; Long, C.; Tang, Y.; Hu, S. Expression of MicroRNA-1 and MicroRNA-21 in different protocols of ischemic conditioning in an isolated rat heart model. Cardiology 2012, 122, 36–43. [Google Scholar] [CrossRef]
- He, B.; Xiao, J.; Ren, A.J.; Zhang, Y.F.; Zhang, H.; Chen, M.; Xie, B.; Gao, X.G.; Wang, Y.W. Role of miR-1 and miR-133a in myocardial ischemic postconditioning. J. Biomed. Sci. 2011, 18, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Spannbauer, A.; Traxler, D.; Lukovic, D.; Zlabinger, K.; Winkler, J.; Gugerell, A.; Ferdinandy, P.; Hausenloy, D.J.; Pavo, N.; Emmert, M.Y.; et al. Effect of ischemic preconditioning and postconditioning on exosome-rich fraction microrna levels, in relation with electrophysiological parameters and ventricular arrhythmia in experimental closed-chest reperfused myocardial infarction. Int. J. Mol. Sci. 2019, 20, 2140. [Google Scholar] [CrossRef] [Green Version]
- Gao, Y.; Huang, R.; Chen, R.; Li, J.; Luo, W. Ischemic postconditioning altered microRNAs in human valve replacement. J. Surg. Res. 2016, 200, 28–35. [Google Scholar] [CrossRef]
- Tu, Y.; Wan, L.; Fan, Y.; Wang, K.; Bu, L.; Huang, T.; Cheng, Z.; Shen, B. Ischemic Postconditioning-Mediated miRNA-21 Protects against Cardiac ischemia/reperfusion Injury via PTEN/Akt Pathway. PLoS ONE 2013, 8, e75872. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Slagsvold, K.H.; Rognmo, Ø.; Høydal, M.; Wisløff, U.; Wahba, A. Remote ischemic preconditioning preserves mitochondrial function and influences myocardial MicroRNA expression in atrial myocardium during coronary bypass surgery. Circ. Res. 2014, 114, 851–859. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brandenburger, T.; Grievink, H.; Heinen, N.; Barthel, F.; Huhn, R.; Stachuletz, F.; Kohns, M.; Pannen, B.; Bauer, I. Effects of remote ischemic preconditioning and myocardial ischemia on microRNA-1 expression in the rat heart in vivo. Shock 2014, 42, 234–238. [Google Scholar] [CrossRef] [PubMed]
- Hu, Q.; Luo, W.; Huang, L.; Huang, R.; Chen, R. Apoptosis-related microRNA changes in the right atrium induced by remote ischemic perconditioning during valve replacement surgery. Sci. Rep. 2016, 6, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Yin, C.; Wang, X.; Kukreja, R.C. Endogenous microRNAs induced by heat-shock reduce myocardial infarction following ischemia-reperfusion in mice. FEBS Lett. 2008, 582, 4137–4142. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Olson, J.M.; Yan, Y.; Bai, X.; Ge, Z.; Liang, M.; Kriegel, A.J.; Twaroski, D.M.; Bosnjak, Z.J. Up-regulation of MicroRNA-21 Mediates Isoflurane-induced Protection of Cardiomyocytes. Anesthesiology 2015, 122, 795–805. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qiao, S.; Olson, J.M.; Paterson, M.; Yan, Y.; Zaja, I.; Liu, Y.; Riess, M.L.; Kersten, J.R.; Liang, M.; Warltier, D.C.; et al. MicroRNA-21 mediates isoflurane-induced cardioprotection against ischemia-reperfusion injury via Akt/Nitric oxide synthase/mitochondrial permeability transition pore pathway. Anesthesiology 2015, 123, 786–798. [Google Scholar] [CrossRef] [Green Version]
- Mukhopadhyay, P.; Mukherjee, S.; Ahsan, K.; Bagchi, A.; Pacher, P.; Das, D.K. Restoration of altered MicroRNA expression in the ischemic heart with resveratrol. PLoS ONE 2010, 5, e15705. [Google Scholar] [CrossRef] [Green Version]
- Ma, N.; Bai, J.; Zhang, W.; Luo, H.; Zhang, X.; Liu, D.; Qiao, C. Trimetazidine protects against cardiac ischemia/reperfusion injury via effects on cardiac miRNA21 expression, Akt and the Bcl2/Bax pathway. Mol. Med. Rep. 2016, 14, 4216–4222. [Google Scholar] [CrossRef]
- Shen, H.; Yao, Z.; Zhao, W.; Zhang, Y.; Yao, C.; Tong, C. miR-21 enhances the protective effect of loperamide on rat cardiomyocytes against hypoxia/reoxygenation, reactive oxygen species production and apoptosis via regulating Akap8 and Bard1 expression. Exp. Ther. Med. 2019, 17, 1312–1320. [Google Scholar] [CrossRef] [Green Version]
- Pan, Q.; Qin, X.; Ma, S.; Wang, H.; Cheng, K.; Song, X.; Gao, H.; Wang, Q.; Tao, R.; Wang, Y.; et al. Myocardial protective effect of extracellular superoxide dismutase gene modified bone marrow mesenchymal stromal cells on infarcted mice hearts. Theranostics 2014, 4, 475–486. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- He, F.; Xiao, Z.; Yao, H.; Li, S.; Feng, M.; Liu, Z.; Wang, W.; Liu, Z.; Wu, J. The protective role of microRNA-21 against coxsackievirus B3 infection through targeting the MAP2K3/P38 MAPK signaling pathway. J. Transl. Med. 2019, 17, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Zhang, L.; Chu, W.; Wang, B.; Zhao, M.; Li, X.; Li, B.; Lu, Y.; Shan, H. Cellular Physiology Biochemistry and Biochemistr y Tanshinone IIA Inhibits miR-1 Expression through p38 MAPK Signal Pathway in Post-infarction Rat Cardiomyocytes. Cell. Physiol. Biochem. 2010, 150081, 991–998. [Google Scholar] [CrossRef] [PubMed]
- Yu, L.; Yu, H.; Li, X.; Jin, C.; Zhao, Y.; Xu, S.; Sheng, X. P38 MAPK/miR-1 are involved in the protective effect of EGCG in high glucose-induced Cx43 downregulation in neonatal rat cardiomyocytes. Cell Biol. Int. 2016, 40, 934–942. [Google Scholar] [CrossRef]
- Chen, T.; Ding, G.; Jin, Z.; Wagner, M.B.; Yuan, Z. Insulin ameliorates miR-1-induced injury in H9c2 cells under oxidative stress via Akt activation. Mol. Cell. Biochem. 2012, 369, 167–174. [Google Scholar] [CrossRef]
- Wu, J.; Sun, C.; Wang, R.; Li, J.; Zhou, M.; Yan, M.; Xue, X.; Wang, C. Cardioprotective effect of paeonol against epirubicin-induced heart injury via regulating miR-1 and PI3K/AKT pathway. Chem. Biol. Interact. 2018, 286, 17–25. [Google Scholar] [CrossRef]
- Devaux, Y.; Vausort, M.; Goretti, E.; Nazarov, P.V.; Azuaje, F.; Gilson, G.; Corsten, M.F.; Schroen, B.; Lair, M.L.; Heymans, S.; et al. Use of circulating microRNAs to diagnose acute myocardial infarction. Clin. Chem. 2012, 58, 559–567. [Google Scholar] [CrossRef] [Green Version]
- Devaux, Y.; Dankiewicz, J.; Salgado-Somoza, A.; Stammet, P.; Collignon, O.; Gilje, P.; Gidlöf, O.; Zhang, L.; Vausort, M.; Hassager, C.; et al. Association of circulating MicroRNA-124-3p levels with outcomes after out-of-hospital cardiac arrest: A substudy of a randomized clinical trial. JAMA Cardiol. 2016, 1, 305–313. [Google Scholar] [CrossRef] [Green Version]
- Devaux, Y.; Mueller, M.; Haaf, P.; Goretti, E.; Twerenbold, R.; Zangrando, J.; Vausort, M.; Reichlin, T.; Wildi, K.; Moehring, B.; et al. Diagnostic and prognostic value of circulating microRNAs in patients with acute chest pain. J. Intern. Med. 2015, 277, 260–271. [Google Scholar] [CrossRef] [Green Version]
- Devaux, Y.; Vausort, M.; McCann, G.P.; Zangrando, J.; Kelly, D.; Razvi, N.; Zhang, L.; Ng, L.L.; Wagner, D.R.; Squire, I.B. MicroRNA-150: A novel marker of left ventricular remodeling after acute myocardial infarction. Circ. Cardiovasc. Genet. 2013, 6, 290–298. [Google Scholar] [CrossRef] [Green Version]
- Scherrer, N.; Fays, F.; Mueller, B.; Luft, A.; Fluri, F.; Christ-Crain, M.; Devaux, Y.; Katan, M. Microrna 150-5p improves risk classification for mortality within 90 days after acute ischemic stroke. J. Stroke 2017, 19, 323–332. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schulte, C.; Barwari, T.; Joshi, A.; Theofilatos, K.; Zampetaki, A.; Barallobre-Barreiro, J.; Singh, B.; Sörensen, N.A.; Neumann, J.T.; Zeller, T.; et al. Comparative analysis of circulating noncoding rnas versus protein biomarkers in the detection of myocardial injury. Circ. Res. 2019, 125, 328–340. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Type of CVD | miR | Findings | Reference |
|---|---|---|---|
| Ischemia-reperfusion (I/R) injury | miR-1 | ↓miR-1 in heart tissue in response to I/R in rats, mice, and infarcted human hearts | [48,49,50,52,53] |
| ↓miR-1 in H9c2 cells and neonatal cardiac myocytes in response to H/R | [48,51] | ||
| ↑miR-1 in remote myocardium compared to infarcted zone or healthy hearts in infarcted human hearts | [54] | ||
| ↑levels of circulating miR-1 after AMI in pigs and humans | [58,59] | ||
| miR-1 overexpression exacerbated cardiac I/R injury in transgenic mice | [55] | ||
| miR-1 inhibition protects against I/R (H/R) injury in rats, mice, and H9c2 cells | [48,55,56] | ||
| miR-21 | ↓miR-21 in in infarct areas, ↑miR-21 in borderline areas in I/R model in rats | [62] | |
| ↑miR-21 in infarct zone of mouse hearts on days two and seven post-MI | [63] | ||
| ↓miR-21 in heart tissue in the first and second week but unchanged in the fourth week post-AMI in mice | [64] | ||
| ↑miR-21 in mouse neonatal cardiomyocytes, H9c2, HL-1, and HCM cells exposed to H/R | [65] | ||
| diverse time-dependent changes in circulating miR-21 in post-MI patients | [76] | ||
| ↑circulating miR-21 in patients with CAD undergoing dobutamine stress echocardiography | [82] | ||
| ↑miR-21 in serum of elderly patients with AMI correlated with levels of CK-MB and cTnI | [68] | ||
| ↓miR-21 in rat neonatal cardiomyocytes exposed to OGD and ↑PDCD4, ↑apoptosis, ↑ROS | [66] | ||
| miR-21 protected cultured cardiac myocytes against H/R-induced apoptosis via ↓PDCD4 | [83] | ||
| ↑miR-21 in the infarct zone promoted myocardial fibrosis post-MI in mice | [71] | ||
| ↑miR-21 in heart atria was associated with ↑atrial collagen content in patients with AF | [72] | ||
| Cardiac arrhythmias | miR-1 | ↑miR-1 expression in the heart associated with ↑arrhythmogenesis in rodents, dogs, and humans | [84,85,86,87] |
| ↓miR-1 expression in heart tissue in patients with age-associated AF and in patients with permanent AF undergoing heart surgery | [88,89] | ||
| miR-21 | ↑miR-21 in heart tissue in patients with AF | [90] | |
| ↓plasma levels of miR-1 in patients with AF | [91] | ||
| ↑miR-21 promote fibrosis in AF in rodents | [92,93,94] | ||
| Non-ischemic cardiomyopathy | miR-21 | ↑miR-21 expression in mice heart with transverse aortic constriction, ↓PDCD4 | [95] |
| ↑miR-21 expression in NRCM with Ang II | [95] | ||
| Dilated cardiomyopathy | miR-1 | unchanged miR-1-3p expression in left ventricles in humans with dilated cardiomyopathy | [10] |
| ↓miR-1 in heart of (miR-1 dKO) mice | [96] | ||
| miR-21 | ↑miR-21 in left ventricles in patients with dilated cardiomyopathy | [10] | |
| Hypertrophic cardiomyopathy | miR-1 | ↓miR-1-3p in human left ventricles | [10] |
| miR-21 | unchanged miR-21 in left ventricles in patients with hypertrophic cardiomyopathy | [10] | |
| ↑miR-21 in fibroblasts of the pressure-overloaded heart | [97] | ||
| unchanged miR-21 in cardiomyocytes of the pressure-overloaded heart | [97] | ||
| Diabetic cardiomyopathy | miR-1 | ↑miR-1 regulation in H9C2 in high glucose | [98] |
| ↓miR-1 in cardiomyocytes treated with high glucose | [99] | ||
| miR-21 | ↑miR-21 in high glucose-treated cardiac fibroblasts | [100] | |
| Viral myocarditis | miR-21 | ↑miR-21 in human and murine coxsakcie B3 myocarditis | [101,102] |
| Radiation-induced heart disease | miR-1 | ↓miR-1 in left ventricle six weeks after 25 Gy and 10 Gy irradiation of mediastinum area | [103,104] |
| miR-21 | ↑miR-21 in left ventricle six weeks after 25 Gy and 10 Gy irradiation of mediastinum area | [103,104] | |
| Anthracyclines-induced cardiomyopathy | miR-1 | ↑miR-1 in blood plasma of rats after doxorubicin treatment | [105] |
| ↑miR-1 in blood plasma of cancer patients after doxorubicin treatment | [106] | ||
| ↓miR-1 in blood plasma of rats after doxorubicin-induced injury | [107] | ||
| ↑miR-1 in heart tissue of rats after doxorubicin treatment | [108] | ||
| ↓miR-1 in blood plasma of cancer child and young adult patients after anthracycline treatment | [109] | ||
| ↓miR-1 in blood plasma of breast cancer patients after doxorubicin treatment | [110] | ||
| miR-21 | ↑miR-21 in mice hearts after doxorubicin treatment | [108,111,112] |
| Type of Intervention | miR | Findings | Reference |
|---|---|---|---|
| Ischemic conditioning | miR-1 | ↑miR-1 in heart tissue after IPostC in rats | [158] |
| ↓miR-1 in plasma after IPostC in pigs | [159] | ||
| ↓miR-1 in right atria after IPostC in human patients undergoing cardiac surgery | [160] | ||
| ↓miR-1 in heart tissue after RIPC in rats | [157] | ||
| ↓miR-1 by RIPC in in vivo cardiac I/R model in rats | [163] | ||
| RIPC prevented up-regulation of miR-1 in right atria and preserved mitochondrial respiration during heart surgery in humans | [162] | ||
| ↓miR-1 by RIPerC during heart surgery in humans | [164] | ||
| miR-21 | ↑miR-21 in heart tissue after IPC and IPostC in animal models and humans | [83,160,161] | |
| knockdown of miR-21 abolished cardioprotective effects of IPost in mice | [161] | ||
| knockdown of cardiac miR-21 abolished IPC-mediated cardioprotection against I/R in rats | [83] | ||
| Pharmacological | miR-1 | insulin protected against miR-1-mediated H2O2-induced injury in H9c2 cells | [175] |
| miR-21 | isoflurane-induced up-regulation of miR-21 associated with ↓PDCD4 protected cardiomyocytes against H2O2 injury | [166] | |
| isoflurane protected mouse hearts exposed to I/R via miR-21 and Akt/NOS/mPTP | [167] | ||
| trimetazidine-induced ↑miR-21 accompanied by cardioprotection against I/R, ↑p-Akt and ↑Bcl-2/Bax in rats. Cardioprotection reversed by anti-miR-21 | [169] | ||
| miR-21 enhanced protective effect of loperamide against H/R injury in rat cardiomyocytes associated with ↓ROS and ↓apoptosis | [170] | ||
| Non-pharmacological | miR-1 | down-regulation of miR-1 by traditional Chinese medicine Tanshinone IIA led to cardioprotection via inhibition of I/R-induced p-38 MAPK in rats | [173] |
| phenolic compound paeonol exerts cardioprotection against epirubicin-induced heart injury via regulation of miR-1, PI3K/AKT/mTOR and NF-κB | [176] | ||
| miR-21 | resveratrol-induced up-regulation of miR-21 associated with protection against I/R in rats | [168] | |
| miR transfection and delivery | miR-21 | adenovirus miR-21 transfection decreased IS via targeting PDCD4/AP-1 | [62] |
| adenovirus miR-21 transfection improved LV remodeling & ↓apoptosis in cardiac I/R in rats | [67] | ||
| lentivirus miR-21 transfection induced cardioprotection against I/R in mice manifested by ↓IS, ↓fibrosis and ↓apoptosis | [64] | ||
| miR-21 transfection to human cardiomyocytes ↓apoptosis via JNK/p38-MAPK/caspase-3 | [68] | ||
| chemically synthesized exogenous miR-21 reduced IS in mice, miR-21-induced protection was abolished with miR-21 inhibitor co-treatment | [165] | ||
| nanoparticle delivery of miR-21 to cardiac macrophages post-MI promoted angiogenesis, reduced hypertrophy, fibrosis, and apoptosis in the remote myocardium | [75] | ||
| miR-21 pretreatment exerted cardioprotection against CVB3 infection via targeting MAP2K3/p38-MAPK in mice | [172] | ||
| Exosomal miR | miR-21 | depletion of exosomal miR-21 reduced protective effect of conditioned medium in H2O2-induced oxidative stress in cardiomyocytes, and in rat hearts exposed to AMI | [69] |
| ↑miR-21 in CPC-derived exosomes prevented apoptosis in H9c2 cells via ↓PDCD4 | [70] |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kura, B.; Kalocayova, B.; Devaux, Y.; Bartekova, M. Potential Clinical Implications of miR-1 and miR-21 in Heart Disease and Cardioprotection. Int. J. Mol. Sci. 2020, 21, 700. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21030700
Kura B, Kalocayova B, Devaux Y, Bartekova M. Potential Clinical Implications of miR-1 and miR-21 in Heart Disease and Cardioprotection. International Journal of Molecular Sciences. 2020; 21(3):700. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21030700
Chicago/Turabian StyleKura, Branislav, Barbora Kalocayova, Yvan Devaux, and Monika Bartekova. 2020. "Potential Clinical Implications of miR-1 and miR-21 in Heart Disease and Cardioprotection" International Journal of Molecular Sciences 21, no. 3: 700. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21030700