Characteristic Dissection of Xanthomonas oryzae pv. oryzae Responsive MicroRNAs in Rice

 , and
, and
Abstract
:1. Introduction
2. Results
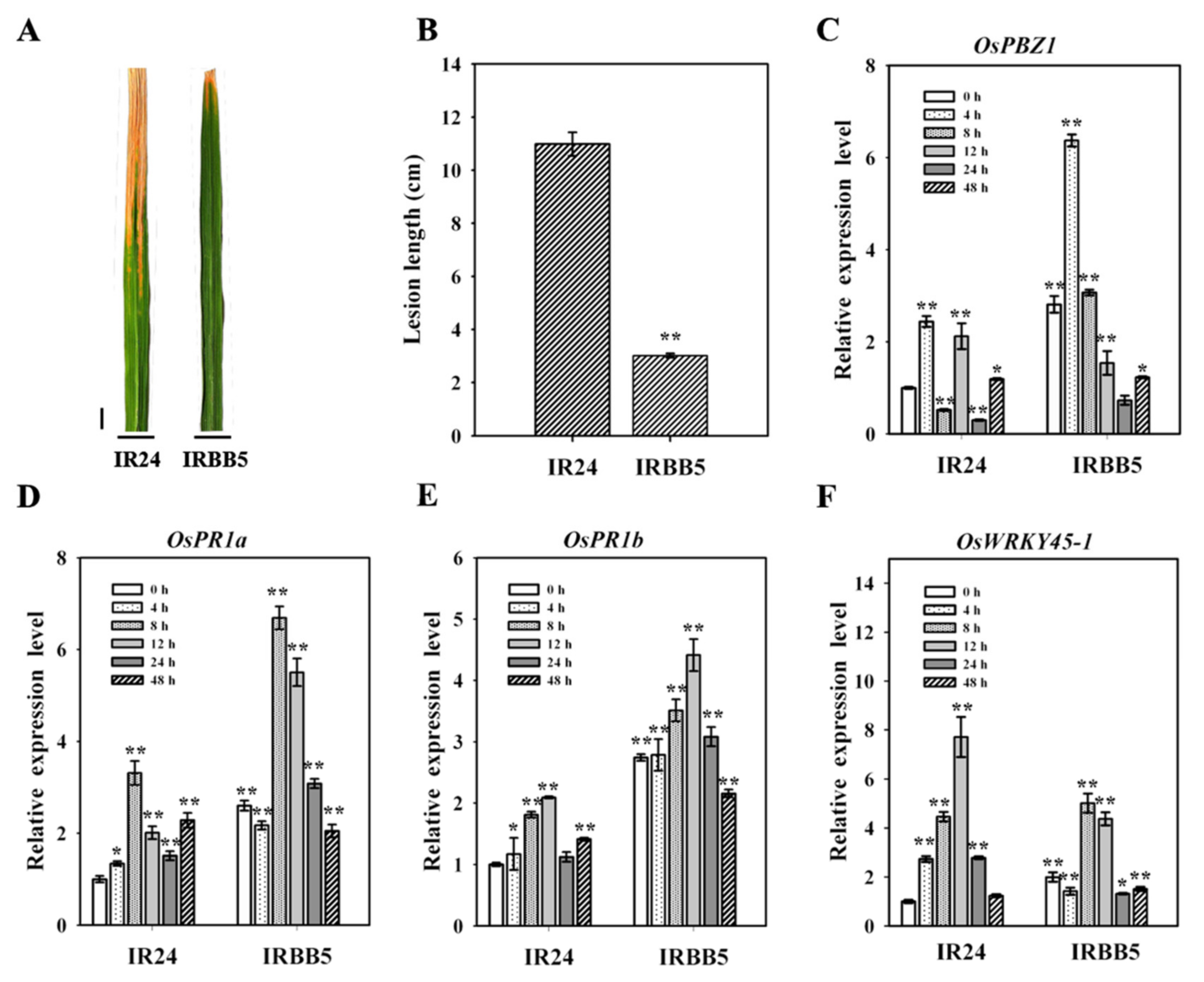
2.1. Diverse Bacterial Blight Response in IR24 and IRBB5 Genotypes
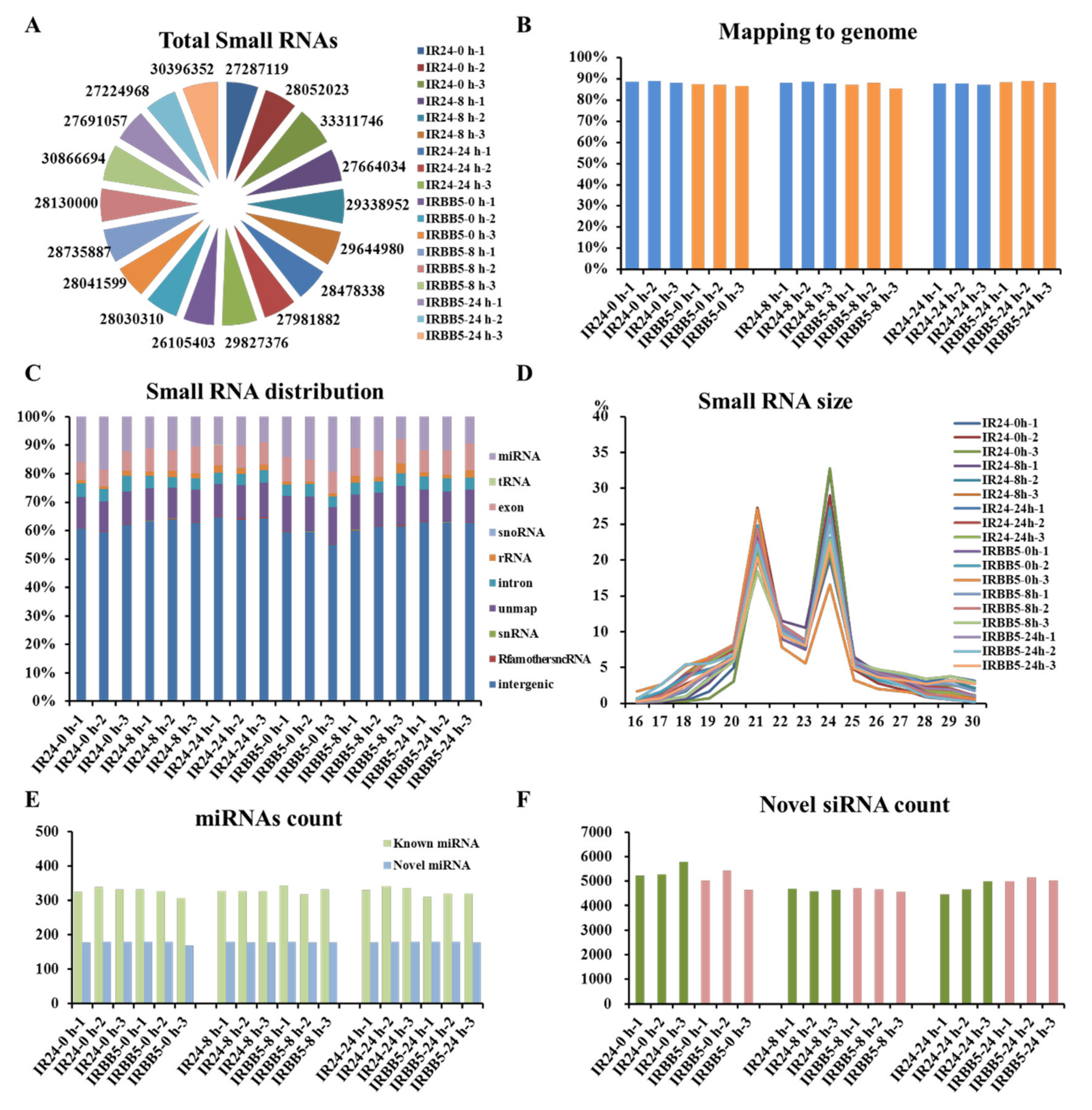
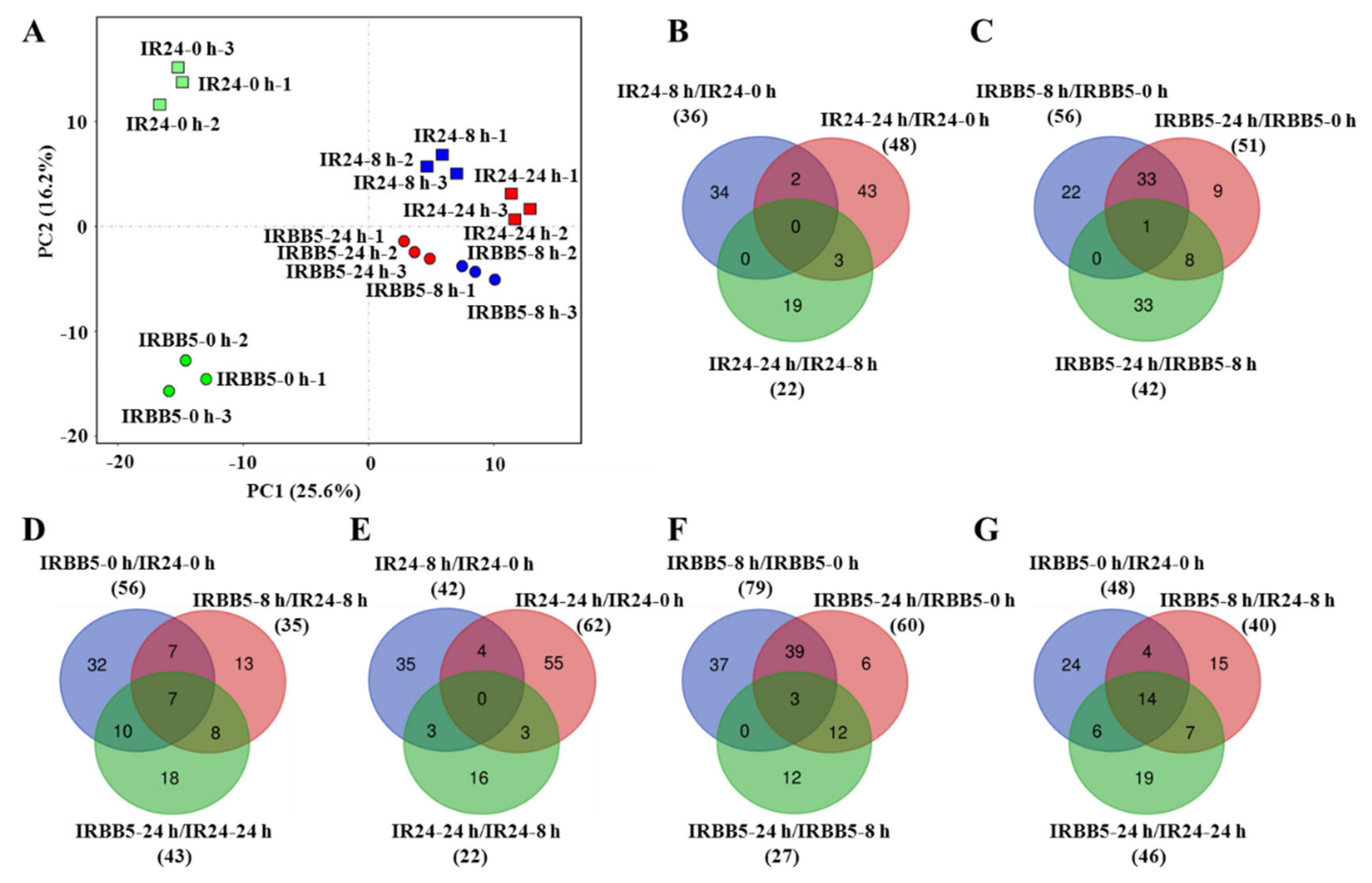
2.2. Overview of Rice Small RNA Sequencing Data under Xoo Strain PXO86 Treatment
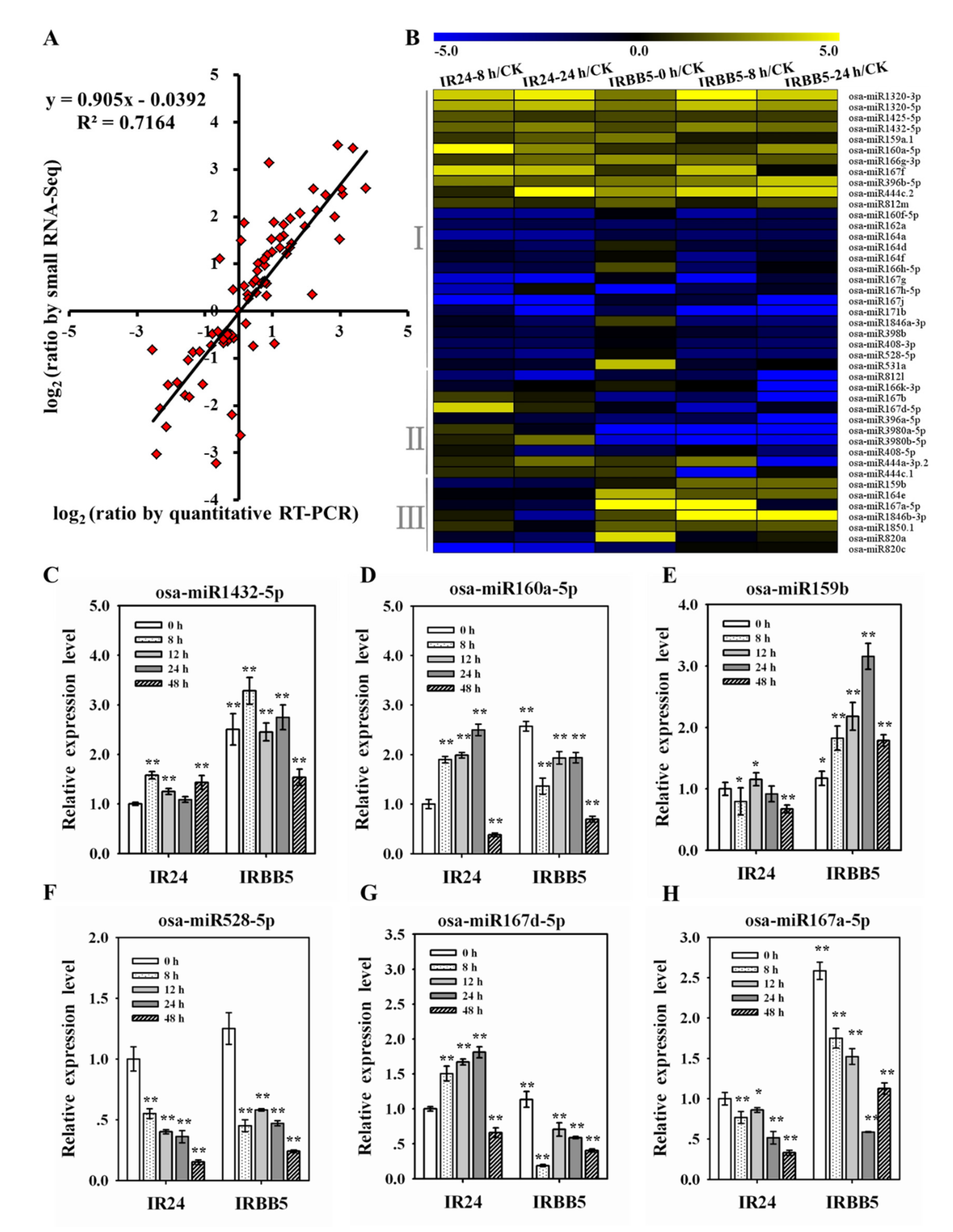
2.3. The Expression Analysis of Xoo-Responsive miRNAs in Infected Rice Leaves
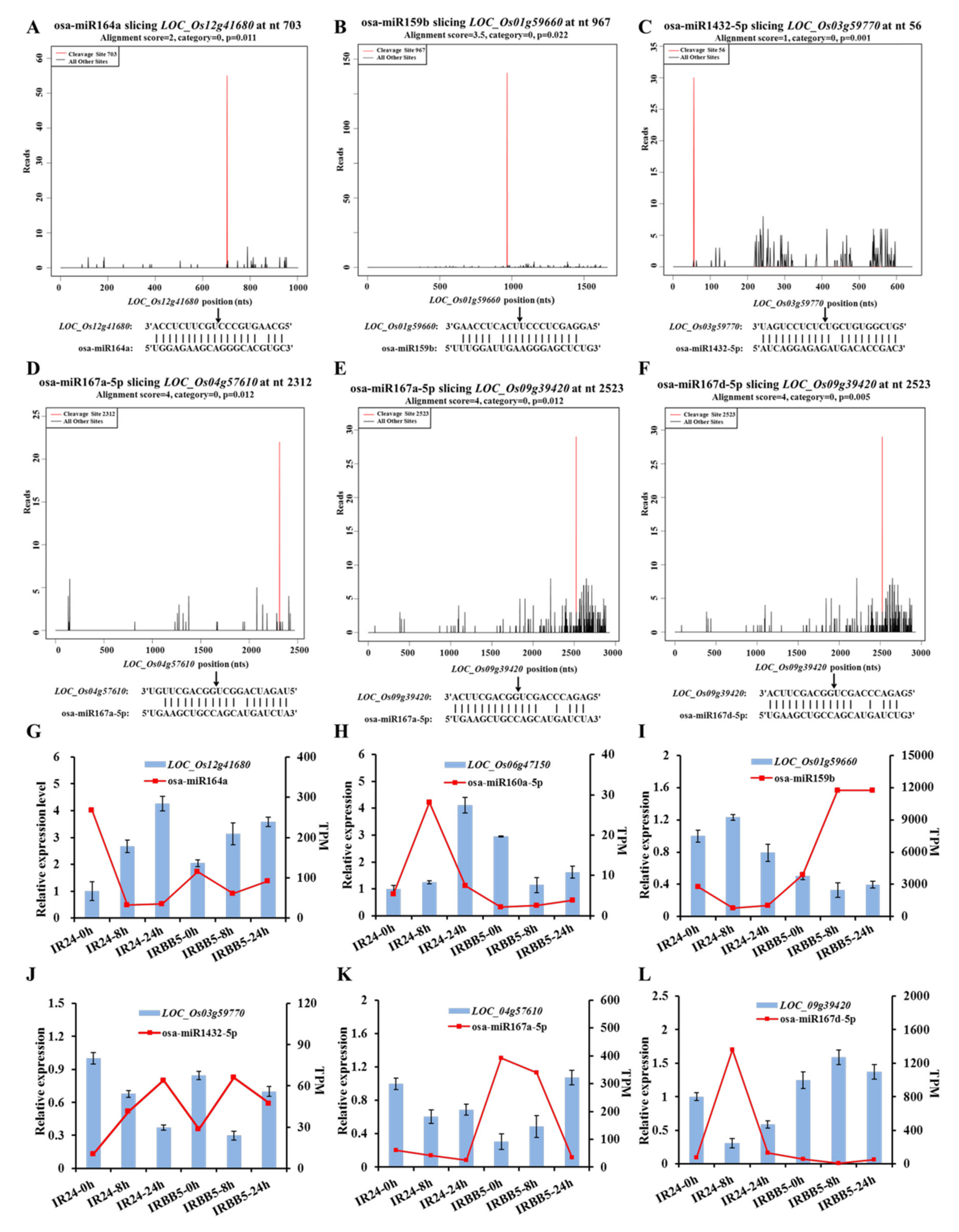
2.4. Target Gene Identification and Function Analysis of miRNAs by Degradome Sequencing
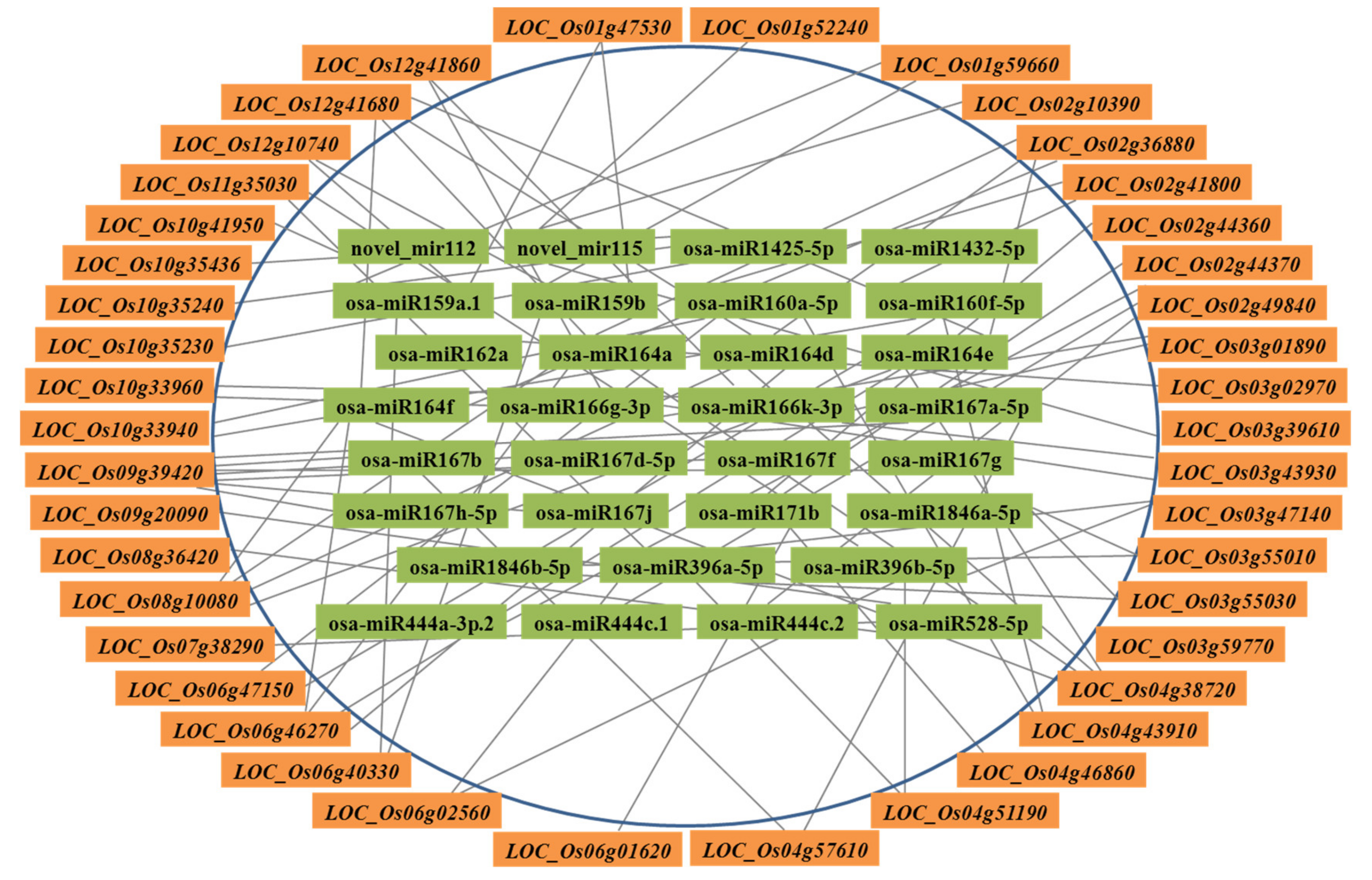
2.5. Multiple Xoo-Responsive miRNA/Target Regulatory Units Detected in Small RNA and Degradome Sequencing Data
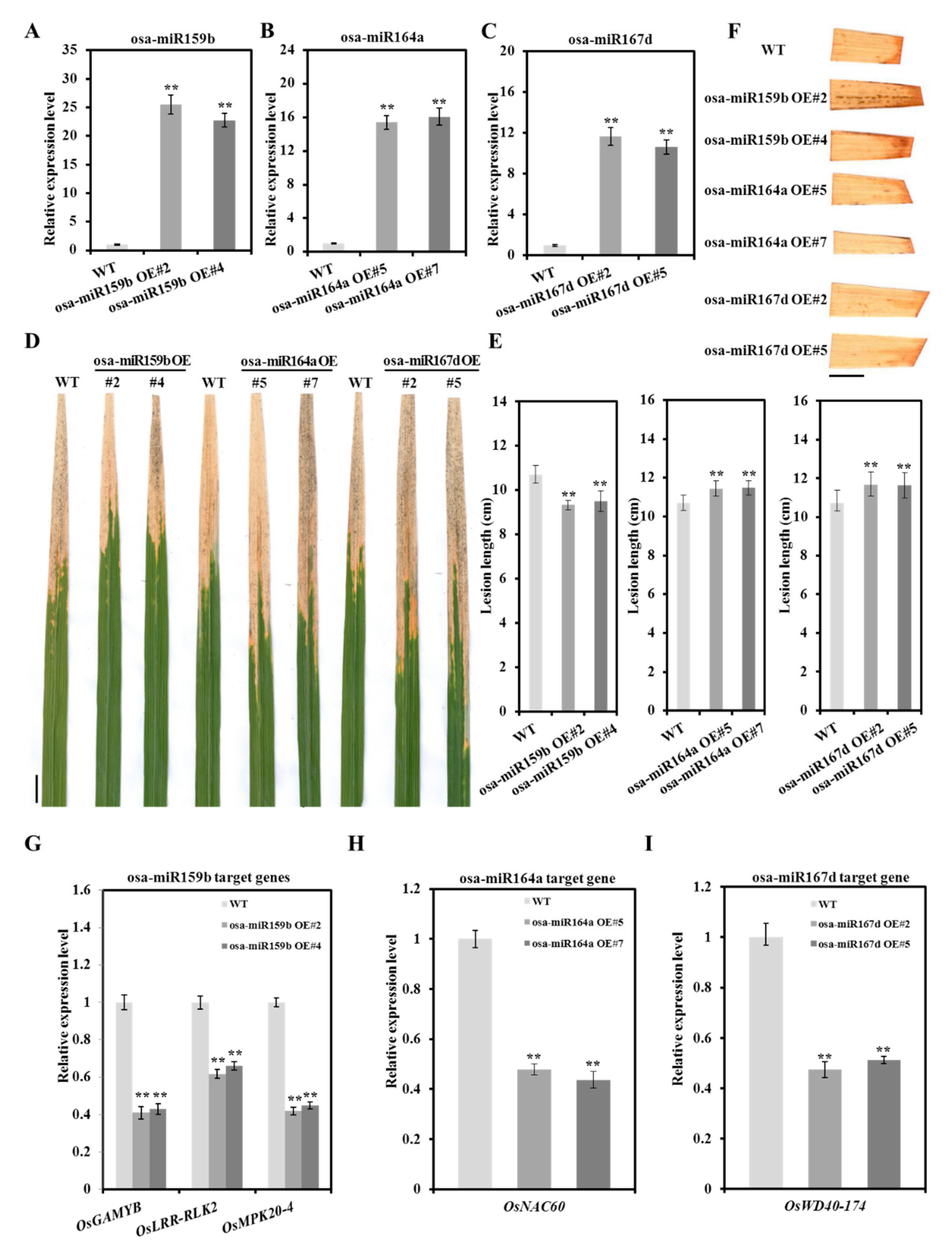
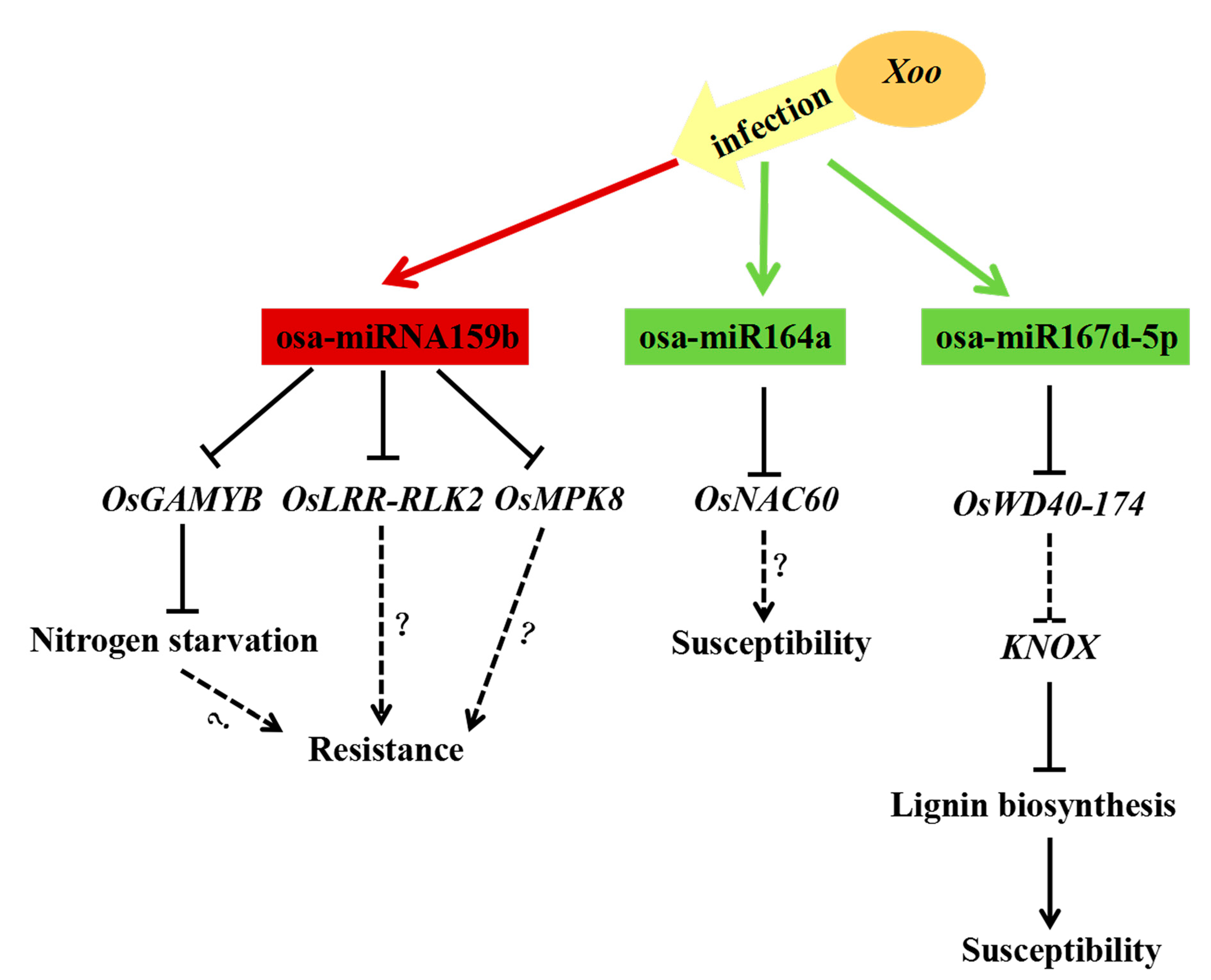
2.6. Diversified Defense Response Caused by Osa-miR159b, Osa-miR164a and Osa-miR167d
3. Discussion
3.1. IR24 and IRBB5 Were Unbiased to Unmask the miRNAs Function in Rice-Xoo Interaction
3.2. Small RNA Sequencing and Degradome Data Jointly Reveal the Post-Transcriptional Regulatory Events upon Xoo Strain Attack
3.3. Multiple miRNA/Targets Regulatory Units Drives Flexible Defender Strategies to BB in Rice
4. Materials and Methods
4.1. Plant Materials and Growth Conditions
4.2. Pathogen Inoculation and Disease Evaluation
4.3. Small RNA Libraries Construction and Analysis
4.4. Degradome Libraries Preparation and Analysis
4.5. Vector Construction and Plant Transformation
4.6. Quantitative Reverse Transcription-PCR Analysis
4.7. 3,3′-Diaminobenzidine (DAB) Staining Assays
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| ABA | Abscisic acid |
| ARF | AUXIN RESPONSE FACTOR |
| BB | Bacterial blight |
| CTK | Cytokinin |
| DAB | 3,3′-Diaminobenzidine |
| DE | Differentially expressed |
| dpi | days post inoculation |
| ET | ethylene |
| GA | gibberellin |
| GRF | GROWTH-REGULATING |
| HB | Homeobox |
| hpi | hours post infection |
| JA | Jasmonic acid |
| miRNAs | microRNAs |
| M. oryzae | Magnaporthe oryzae |
| NIL | Near-isogenic line |
| OE | Overexpressing |
| OsARF8 | LOC_Os04g57610 |
| OsAGO1a | LOC_Os02g45070 |
| OsAGO1b | LOC_Os04g47870 |
| OsDCL1 | LOC_Os03g02970 |
| OsGAMYB | LOC_Os01g59660 |
| OsLRR-RLK2 | LOC_Os12g10740 |
| OsMPK20-4 | LOC_Os01g47530 |
| OsNAC60 | LOC_Os12g41680 |
| OsPBZ1 | Oryza sativa pathogenesis-related protein 10a |
| OsPR1a | Pathogenesis-related protein 1a |
| OsPR1b | Pathogenesis-related protein 1b |
| OsWD40-174 | LOC_Os09g39420 |
| PC | Principle component |
| RDV | Rice dwarf virus |
| Rice | Oryza sativa L. |
| R. solani | Rhizoctonia solani |
| RSV | Rice stripe virus |
| SA | Salicylic acid |
| SPL | SQUAMOSA PROMOTER BINDING-LIKE FACTOR |
| TP309 | Taipei 309 |
| TPM | Transcripts per million |
| WT | Wild-type |
| Xoo | Xanthomonas oryzae pv. oryzae |
References
- Nino-Liu, D.O.; Ronald, P.C.; Bogdanove, A.J. Xanthomonas oryzae pathovars: Model pathogens of a model crop. Mol. Plant. Pathol. 2006, 7, 303–324. [Google Scholar] [CrossRef]
- Song, W.Y.; Wang, G.L.; Chen, L.L.; Kim, H.S.; Pi, L.Y.; Holsten, T.; Gardner, J.; Wang, B.; Zhai, W.X.; Zhu, L.H.; et al. A receptor kinase-like protein encoded by the rice disease resistance gene, Xa21. Science 1995, 270, 1804–1806. [Google Scholar] [CrossRef] [Green Version]
- Yoshimura, S.; Yamanouchi, U.; Katayose, Y.; Toki, S.; Wang, Z.X.; Kono, I.; Kurata, N.; Yano, M.; Iwata, N.; Sasaki, T.; et al. Expression of Xa1, a bacterial blight-resistance gene in rice, is induced by bacterial inoculation. Proc. Natl. Acad. Sci. USA 1998, 95, 1663–1668. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, Z.; Sun, X.; Wang, S.; Zhang, Q. Genetic and physical mapping of a new gene for bacterial blight resistance in rice. Theor. Appl. Genet. 2003, 106, 1467–1472. [Google Scholar] [CrossRef] [PubMed]
- Iyer, A.S.; McCouch, S.R. The rice bacterial blight resistance gene xa5 encodes a novel form of disease resistance. Mol. Plant. Microbe. Int. 2004, 17, 1348–1354. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gu, K.Y.; Yang, B.; Tian, D.S.; Wu, L.F.; Wang, D.J.; Sreekala, C.; Yang, F.; Chu, Z.Q.; Wang, G.L.; White, F.F.; et al. R gene expression induced by a type-III effector triggers disease resistance in rice. Nature 2005, 435, 1122–1125. [Google Scholar] [CrossRef]
- Chu, Z.H.; Fu, B.Y.; Yang, H.; Xu, C.G.; Li, Z.K.; Sanchez, A.; Park, Y.J.; Bennetzen, J.L.; Zhang, Q.F.; Wang, S.P. Targeting xa13, a recessive gene for bacterial blight resistance in rice. Theor. Appl. Genet. 2006, 112, 455–461. [Google Scholar] [CrossRef]
- Xiang, Y.; Cao, Y.L.; Xu, C.G.; Li, X.H.; Wang, S.Q. Xa3, conferring resistance for rice bacterial blight and encoding a receptor kinase-like protein, is the same as Xa26. Theor. Appl. Genet. 2006, 113, 1347–1355. [Google Scholar] [CrossRef]
- Liu, Q.S.; Yuan, M.; Zhou, Y.; Li, X.H.; Xiao, J.H.; Wang, S.P. A paralog of the MtN3/saliva family recessively confers race-specific resistance to Xanthomonas oryzae in rice. Plant. Cell Environ. 2011, 34, 1958–1969. [Google Scholar] [CrossRef]
- Tian, D.S.; Wang, J.X.; Zeng, X.; Gu, K.; Qiu, C.X.; Yang, X.B.; Zhou, Z.Y.; Goh, M.; Luo, Y.C.; Murata-Hori, M.; et al. The rice TAL effector-dependent resistance protein XA10 triggers cell death and calcium depletion in the endoplasmic reticulum. Plant Cell. 2014, 26, 497–515. [Google Scholar] [CrossRef] [Green Version]
- Wang, C.L.; Zhang, X.P.; Fan, Y.L.; Gao, Y.; Zhu, Q.L.; Zheng, C.K.; Qin, T.F.; Li, Y.Q.; Che, J.Y.; Zhang, M.W.; et al. XA23 is an executor R protein and confers broad-spectrum disease resistance in rice. Mol. Plant. 2015, 8, 290–302. [Google Scholar] [CrossRef] [Green Version]
- Hu, K.M.; Cao, J.B.; Zhang, J.; Xia, F.; Ke, Y.G.; Zhang, H.T.; Xie, W.Y.; Liu, H.B.; Cui, H.; Cao, Y.L.; et al. Improvement of multiple agronomic traits by a disease resistance gene via cell wall reinforcement. Nat. Plants. 2017, 3, 17009–17017. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Lu, Y.G.; Shi, Y.; Wu, L.; Xu, Y.J.; Huang, F.; Guo, X.Y.; Zhang, Y.; Fan, J.; Zhao, J.Q.; et al. Multiple rice microRNAs are involved in immunity against the blast fungus Magnaporthe oryzae. Plant Physiol. 2014, 164, 1077–10920. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Salvador-Guirao, R.; Hsing, Y.I.; Segundo, B.S. The polycistronic miR166k-166h positively regulates rice immunity via post-transcriptional control of EIN2. Front. Plant Sci. 2018, 9, 337–352. [Google Scholar] [CrossRef] [PubMed]
- Campo, S.; Peris, C.; Sire, C.; Moreno, A.B.; Donaire, L.; Zytnicki, M.; Notredame, C.; Llave, C.; Segundo, B.S. Identification of a novel microRNA (miRNA) from rice that targets an alternatively spliced transcript of the Nramp6 (Natural resistance-associated macrophage protein 6) gene involved in pathogen resistance. New Phytol. 2013, 199, 212–227. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.Y.; Xia, Y.Q.; Lin, S.Y.; Wang, Y.R.; Guo, B.H.; Song, X.N.; Ding, S.C.; Zheng, L.Y.; Feng, R.Y.; Chen, S.L.; et al. Osa-miR164a targets OsNAC60 and negatively regulates rice immunity against the blast fungus Magnaporthe oryzae. Plant J. 2018, 95, 584–597. [Google Scholar] [CrossRef] [Green Version]
- Zhao, Z.X.; Feng, Q.; Cao, X.L.; Zhu, Y.; Wang, H.; Chandran, V.; Fan, J.; Zhao, J.Q.; Pu, M.; Li, Y.; et al. Osa-miR167d facilitates infection of Magnaporthe oryzae in rice. J. Integ. Plant Biol. 2019. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Zhao, S.L.; Li, J.L.; Hu, X.H.; Wang, H.; Cao, X.L.; Xu, Y.J.; Zhao, Z.X.; Xiao, Z.Y.; Yang, N.; et al. Osa-miR169 negatively regulates rice immunity against the blast fungus magnaporthe oryzae. Front. Plant Sci. 2017, 8. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.; Bao, Y.L.; Shan, D.Q.; Wang, Z.H.; Song, X.N.; Wang, Z.Y.; Wang, J.S.; He, L.Q.; Wu, L.; Zhang, Z.G.; et al. Magnaporthe oryzae induces the expression of a microRNA to suppress the immune response in rice. Plant Physiol. 2018, 177, 352–368. [Google Scholar] [CrossRef] [Green Version]
- Chandran, V.; Wang, H.; Gao, F.; Cao, X.L.; Chen, Y.P.; Li, G.B.; Zhu, Y.; Yang, X.M.; Zhang, L.L.; Zhao, Z.X.; et al. MiR396-OsGRFs Module Balances Growth and Rice Blast Disease-Resistance. Front. Plant Sci. 2018, 9, 1999–2015. [Google Scholar] [CrossRef] [Green Version]
- Lin, R.; He, L.; He, J.Y.; Qin, P.G.; Wang, Y.R.; Deng, Q.M.; Yang, X.T.; Li, S.C.; Wang, S.Q.; Wang, W.M.; et al. Comprehensive analysis of microRNA-Seq and target mRNAs of rice sheath blight pathogen provides new insights into pathogenic regulatory mechanisms. DNA Res. 2016, 23, 415–425. [Google Scholar] [CrossRef] [PubMed]
- Lian, S.; Cho, W.K.; Kim, S.M.; Choi, H.; Kim, K.H. Time-course small RNA profiling reveals rice miRNAs and their target genes in response torice stripe virus infection. PLoS ONE 2016, 11, e0162319. [Google Scholar] [CrossRef] [PubMed]
- Zarreen, F.; Kumar, G.; Johnson, A.M.; Dasgupta, I. Small RNA-based interactions between rice and the viruses which cause the tungro disease. Virology 2018, 523, 64–73. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.G.; Yang, R.X.; Yang, Z.R.; Yao, S.Z.; Zhao, S.S.; Wang, Y.; Li, P.C.; Song, X.W.; Jin, L.; Zhou, T.; et al. Ros accumulation and antiviral defence control by microRNA528 in rice. Nat. Plants 2017, 3, 16203–16209. [Google Scholar] [CrossRef]
- Tong, A.; Yuan, Q.; Wang, S.; Peng, J.J.; Lu, Y.W.; Zheng, H.Y.; Lin, L.; Chen, H.; Gong, Y.F.; Chen, J.P.; et al. Altered accumulation of osa-miR171b contributes to rice stripe virus infection by regulating disease symptoms. J. Exp. Bot. 2017, 68, 4357–4367. [Google Scholar] [CrossRef]
- Wang, H.C.; Jiao, X.M.; Kong, X.Y.; Hamera, S.; Wu, Y.; Chen, X.Y.; Fang, R.X.; Yan, Y.S. A signaling cascade from miR444 to RDR1 in Rice antiviral RNA silencing pathway. Plant Physiol. 2016, 170, 2365–2377. [Google Scholar] [CrossRef] [Green Version]
- Zhang, C.; Ding, Z.M.; Wu, K.C.; Yang, L.; Li, Y.; Yang, Z.; Shi, S.; Liu, X.J.; Zhao, S.S.; Yang, Z.R.; et al. Suppression of jasmonic acid-mediated defense by viral-inducible microRNA319 facilitates virus infection in rice. Mol. Plant 2016, 9, 1302–1314. [Google Scholar] [CrossRef] [Green Version]
- Zhao, Y.T.; Wang, M.; Wang, Z.M.; Fang, R.X.; Wang, X.J.; Jia, Y.T. Dynamic and coordinated expression changes of rice small RNAs in response to Xanthomonas oryzae pv. oryzae. J. Genet. Genomics 2015, 42, 625–637. [Google Scholar] [CrossRef]
- Hong, H.M.; Liu, Y.Y.; Zhang, H.T.; Xiao, J.H.; Li, X.H.; Wang, S.P. Small RNAs and gene network in a durable disease resistance gene-mediated defense responses in rice. PLoS ONE 2015, 10, e0137360. [Google Scholar] [CrossRef] [Green Version]
- Zhang, S.; Mei, J.; Wang, T.; Wang, C.C.; Zhang, W.L.; Yang, L. Identification and expression analysis of OsmiR1861k in rice leaves in response to Xanthomonas oryzae pv. oryzae. J. Gen. Plant Pathol. 2015, 81, 108–117. [Google Scholar] [CrossRef]
- Liu, M.M.; Shi, Z.Y.; Zhang, X.H.; Wang, M.X.; Zhang, L.; Zheng, K.Z.; Liu, J.Y.; Hu, X.M.; Di, C.R.; Qian, Q.; et al. Inducible overexpression of Ideal Plant. Architecture1 improves both yield and disease resistance in rice. Nat. Plants 2019, 5, 389–400. [Google Scholar] [CrossRef] [PubMed]
- Yu, C.; Chen, Y.T.; Cao, Y.Q.; Chem, H.M.; Wang, J.C.; Bi, Y.M.; Tian, F.; Yang, F.H.; Rothstein, S.J.; Zhou, X.P.; et al. Overexpression of miR169o, an Overlapping microRNA in response to both nitrogen limitation and bacterial infection, promotes nitrogen use efficiency and susceptibility to bacterial blight in rice. Plant. Cell Physiol. 2018, 59, 1234–1247. [Google Scholar] [CrossRef] [PubMed]
- Blair, M.W.; Mccouch, S.R. Microsatellite and sequence-tagged site markers diagnostic for the rice bacterial leaf blight resistance gene xa-5. Theor. Appl. Genet. 1997, 95, 174–184. [Google Scholar] [CrossRef]
- Kim, S.G.; Kim, S.T.; Wang, Y.; Yu, S.; Choi, I.S.; Kim, Y.C.; Kim, W.T.; Agrawal, G.K.; Rakwal, R.; Kang, K.Y. The RNase activity of rice probenazole-induced protein1 (PBZ1) plays a key role in cell death in plants. Mol. Cells 2011, 31, 25–31. [Google Scholar] [CrossRef] [PubMed]
- Mitsuhara, I.; Iwai, T.; Seo, S.; Yanagawa, Y.; Kawahigasi, H.; Hirose, S.; Ohkawa, Y.; Ohashi, Y. Characteristic expression of twelve rice PR1 family genes in response to pathogen infection, wounding, and defense-related signal compounds (121/180). Mol. Genet. Genomics 2008, 279, 415–427. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tao, Z.; Liu, H.B.; Qiu, D.Y.; Zhou, Y.Z.; Li, X.H.; Xu, C.G.; Wang, S.P. A Pair of allelic WRKY genes play opposite roles in rice-bacteria interactions. Plant Physiol. 2009, 151, 936–948. [Google Scholar] [CrossRef] [Green Version]
- Wilderman, P.R.; Xu, M.; Jin, Y.; Coates, R.M.; Peters, R.J. Identification of syn-Pimara-7,15-diene synthase reveals functional clustering of terpene synthases involved in rice phytoalexin/allelochemical biosynthesis. Plant Physiol. 2004, 135, 2098–2105. [Google Scholar] [CrossRef] [Green Version]
- Spoel, S.H.; Dong, X. Making sense of hormone crosstalk during plant immune responses. Cell Host Microbe. 2008, 3, 348–351. [Google Scholar] [CrossRef] [Green Version]
- Pieterse, C.M.; Leon-Reyes, A.; Sjoerd, V.D.; Wees, S.C. Networking by small-molecule hormones in plant immunity. Nat. Chem. Bio. 2009, 5, 308–316. [Google Scholar] [CrossRef] [Green Version]
- Huang, L.F.; Lin, K.H.; He, S.L.; Chen, J.L.; Jiang, J.Z.; Chen, B.H.; Hou, Y.S.; Chen, R.S.; Hong, C.Y.; Ho, S.L. Multiple patterns of regulation and overexpression of a ribonuclease-like pathogenesis-related protein gene, OsPR10a, conferring disease resistance in rice and Arabidopsis. PLoS ONE 2016, 11, e0156414. [Google Scholar] [CrossRef]
- Sun, Z.T.; He, Y.Q.; Li, J.M.; Wang, X.; Chen, J.Q. Genome-wide characterization of rice black streaked dwarf virus-responsive microRNAs in rice leaves and roots by small RNA and degradome sequencing. Plant. Cell Physiol. 2015, 56, 688–699. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Du, P.; Wu, J.G.; Zhang, J.Y.; Zhao, S.Q.; Zheng, H.; Gao, G.; Wei, L.P.; Li, Y. Viral infection induces expression of novel phased microRNAs from conserved cellular microRNA precursors. PLoS Pathog. 2011, 7, e1002176. [Google Scholar] [CrossRef] [PubMed]
- Katz, A.; Oliva, M.; Mosquna, A.; Hakim, O.; Ohad, N. FIE and CURLY LEAF polycomb proteins interact in the regulation of homeobox gene expression during sporophyte development. Plant J. 2004, 37, 707–719. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mele, G.; Ori, N.; Sato, Y.; Hake, S. The knotted1-like homeobox gene BREVIPEDICELLUS regulates cell differentiation by modulating metabolic pathways. Gene Dev. 2003, 17, 2852. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hong, Y.F.; Ho, T.H.; Wu, C.F.; Ho, S.L.; Yeh, R.H.; Lu, C.A.; Chen, P.W.; Yu, L.C.; Chao, A.; Yu, S.M. Convergent starvation signals and hormone crosstalk in regulating nutrient mobilization upon germination in cereals. Plant Cell. 2012, 24, 2857–2873. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ham, Y.; Kim, T.J. Nitrogen sources inhibit biofilm formation of Xanthomonas oryzae pv. oryzae. J. Microbiol. Biotechn. 2018, 28, 2071–2078. [Google Scholar] [CrossRef]
- Reyna, N.S.; Yang, Y.N. Molecular analysis of the rice MAP kinase gene family in relation to Magnaporthe grisea infection. Mol. Plant Microbe. Int. 2006, 19, 530–540. [Google Scholar] [CrossRef] [Green Version]
- Sheikh, A.H.; Raghuram, B.; Jalmi, S.K.; Wankhede, D.P.; Singh, P.; Sinha, A.K. Interaction between two rice mitogen activated protein kinases and its possible role in plant defense. BMC Plant Biol. 2013, 13, 121–131. [Google Scholar] [CrossRef] [Green Version]
- Chen, H.L.; Wang, S.P.; Zhang, Q.F. New gene for bacterial blight resistance in rice located on chromosome 12 identified from minghui 63, an elite restorer line. Phytopathology 2002, 92, 750–754. [Google Scholar] [CrossRef]
- Meyers, B.C.; Axtell, M.J.; Bartel, B.; Bartel, D.P.; Baulcombe, D.; Bowman, J.L.; Cao, X.; Carrington, J.C.; Chen, X.; Green, P.J.; et al. Criteria for annotation of plant microRNAs. Plant Cell. 2008, 20, 3186–3190. [Google Scholar] [CrossRef]
- Addo-Quaye, C.; Eshoo, T.W.; Bartel, D.P.; Axtell, M.J. Endogenous siRNA and miRNA targets identified by sequencing of the Arabidopsis degradome. Curr. Biol. 2008, 18, 758–762. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Addo-Quaye, C.; Miller, W.; Axtell, M.J. CleaveLand: A pipeline for using degradome data to find cleaved small RNA targets. Bioinformatics 2008, 25, 130–131. [Google Scholar] [CrossRef] [PubMed]
- Asuka, N.; Ikuko, A.; Makoto, M. A protocol for Agrobacterium-mediated transformation in rice. Nat. Protoc. 2006, 1, 2796–2802. [Google Scholar]
- Varkonyi-Gasic, E.; Wu, R.; Wood, M.; Walton, E.F.; Hellens, R.P. Protocol: A highly sensitive RT-PCR method for detection and quantification of microRNAs. Plant Methods 2007, 3, 12–21. [Google Scholar] [CrossRef] [Green Version]
- Daudi, A.; O’Brien, J.A. Detection of hydrogen peroxide by DAB staining in Arabidopsis leaves. Bio. Protoc. 2012, 2, e263–e266. [Google Scholar] [CrossRef] [Green Version]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| miRNA | Target | Cleavage Site (nt) | Category | Target Annotation | Response Type | Raw Reads in IRBB5 | Raw Reads in IR24 |
|---|---|---|---|---|---|---|---|
| osa-miR1425. | LOC_Os10g35436. | 1213 | 0 | Pentatricopeptide repeat protein 1 | Bacterium, fungus, innate immunity/ Jasmonic acid (JA) | 34 | 92 |
| LOC_Os10g35230 | 1192 | 0 | Pentatricopeptide repeat protein 2 | 10 | 13 | ||
| LOC_Os10g35240 | 1219 | 0 | Pentatricopeptide repeat protein 4 | 42 | 57 | ||
| osa-miR1426 | LOC_Os10g06130 | 829 | 0 | RNA recognition motif containing protein | Jasmonic acid (JA) | 13 | 12 |
| osa-miR1428 | LOC_Os03g17980 | 1228 | 0 | Serine/threonine protein kinase | 2 | 10 | |
| osa-miR1432 | LOC_Os03g59770 | 56 | 0 | calcium-binding protein CML21-like | 30 | 48 | |
| osa-miR1437 | LOC_Os01g22490 | 381 | 0 | Ubiquitin 5 | 13 | 9 | |
| osa-miR156 | LOC_Os06g45310 | 759 | 0 | Squamosa promoter binding-like 11 | 10 | 30 | |
| LOC_Os02g04680 | 1158 | 0 | Squamosa promoter binding-like 3 | 21 | 34 | ||
| LOC_Os08g41940 | 1008 | 0 | Squamosa promoter binding-like 16 | 9 | 7 | ||
| LOC_Os06g49010 | 1155 | 0 | Squamosa promoter binding-like 12 | 17 | 11 | ||
| osa-miR159 | LOC_Os01g59660 | 967 | 0 | GAMYB-like protein 1 | Gibberellin (GA) | 140 | 280 |
| LOC_Os12g10740 | 1018 | 0 | Leucine-rich repeat family protein | 2 | 10 | ||
| LOC_Os01g47530 | 1664 | 0 | Mitogen-activated protein kinase 20-4 | Basal defense | 7 | 31 | |
| LOC_Os06g40330 | 1429 | 0 | MYB transcription factor | 12 | 25 | ||
| osa-miR160 | LOC_Os02g41800 | 1340 | 0 | Auxin response factor 8 | 134 | 105 | |
| LOC_Os10g33940 | 1367 | 0 | Auxin response factor 16 | Auxin | 1212 | 1347 | |
| LOC_Os06g47150 | 1379 | 0 | Auxin response factor 18 | Auxin | 2407 | 2987 | |
| LOC_Os04g43910 | 1355 | 0 | Auxin response factor 10 | 48 | 59 | ||
| osa-miR162 | LOC_Os03g02970 | 2988 | 0 | Dicer-like protein 1 | 24 | 42 | |
| osa-miR164 | LOC_Os08g10080 | 697 | 2 | NAC transcription factor 104 | 17 | 2 | |
| LOC_Os04g38720 | 697 | 0 | NAC transcription factor 2 | Gibberellin (GA) | 29 | 7 | |
| LOC_Os12g41680 | 703 | 0 | NAC transcription factor 60 | Magnaporthe oryzae | 55 | 140 | |
| LOC_Os06g46270 | 679 | 0 | NAC transcription factor 11 | Auxin | 28 | 20 | |
| LOC_Os02g36880 | 787 | 0 | NAC transcription factor 1 | 72 | 47 | ||
| osa-miR166 | LOC_Os03g43930 | 622 | 0 | Homeobox-leucine zipper protein HOX32 | Auxin | 66 | 143 |
| LOC_Os12g41860 | 613 | 0 | Homeobox-leucine zipper protein HOX33 | Auxin | 66 | 142 | |
| LOC_Os03g01890 | 574 | 0 | Homeobox-leucine zipper protein HOX10 | Auxin | 266 | 277 | |
| LOC_Os10g33960 | 580 | 0 | Homeobox-leucine zipper protein HOX9 | Auxin | 267 | 278 | |
| osa-miR167 | LOC_Os04g57610 | 2312 | 0 | Auxin response factor 8 | Auxin | 22 | 8 |
| LOC_Os09g39420 | 2523 | 2 | WD40 repeat-like domain containing protein | 3 | 29 | ||
| osa-miR168 | LOC_Os02g45070 | 427 | 2 | Protein argonaute 1A | Virus, innate immunity/Auxin | 31 | 34 |
| LOC_Os04g47870 | 535 | 2 | Protein argonaute 1B | Virus, innate immunity /Auxin | 6 | 19 | |
| osa-miR171 | LOC_Os06g01620 | 452 | 0 | GRAS plant-specific transcription factor | 354 | 367 | |
| LOC_Os04g46860 | 1130 | 0 | GRAS plant-specific transcription factor | 206 | 328 | ||
| LOC_Os02g44370 | 1139 | 0 | GRAS plant-specific transcription factor | 80 | 75 | ||
| LOC_Os02g44360 | 1121 | 0 | GRAS plant-specific transcription factor | 73 | 73 | ||
| osa-miR172 | LOC_Os05g03040 | 1423 | 2 | AP2/EREBP transcription factor | Ethylene (ETH) | 1728 | 1728 |
| LOC_Os07g13170 | 1207 | 0 | AP2 transcription factor | 21 | 43 | ||
| osa-miR1846 | LOC_Os03g55010 | 1219 | 0 | UDP-glucosyltransferase family protein | 6 | 10 | |
| LOC_Os03g55030 | 1213 | 0 | UDP-glucosyltransferase family protein | 6 | 10 | ||
| osa-miR2093 | LOC_Os09g06499 | 1549 | 2 | Sulfate transporter 4.1 | 18 | 10 | |
| osa-miR2094 | LOC_Os08g14440 | 506 | 2 | Uridylyltransferase-related | Cytokinin (CTK) | 26 | 14 |
| osa-miR2100 | LOC_Os05g27940 | 250 | 2 | Ribosomal protein S7 | 7 | 11 | |
| osa-miR2102 | LOC_Os03g46570 | 902 | 2 | SKIPa-interacting protein 31 | Bacterium, fungus/ Gibberellin (GA) | 11 | 3 |
| LOC_Os09g34140 | 183 | 2 | Conserved hypothetical protein | 17 | 12 | ||
| osa-miR2925 | LOC_Os07g25430 | 235 | 2 | Photosystem I reaction center subunit IV, chloroplast precursor | Cytokinin (CTK) | 42 | 21 |
| LOC_Os01g45470 | 82 | 2 | Conserved hypothetical protein | 32 | 34 | ||
| osa-miR2926 | LOC_Os10g35840 | 1175 | 2 | Glutamyl-tRNA reductase 2 | 31 | 22 | |
| osa-miR393 | LOC_Os05g05800 | 1552 | 2 | F-Box auxin receptor protein | Defense response/ Auxin, Ethylene (ETH) | 66 | 102 |
| LOC_Os04g32460 | 1513 | 2 | Auxin signaling F-box 2 | Defense response/Auxin | 23 | 20 | |
| osa-miR394 | LOC_Os01g69940 | 1093 | 2 | Cyclin-like F-box domain containing protein | Auxin | 165 | 169 |
| osa-miR396 | LOC_Os06g02560 | 410 | 2 | Growth-regulating factor 5 | 62 | 55 | |
| LOC_Os11g35030 | 593 | 0 | Growth regulating factor protein 8 | Gibberellin (GA) | 31 | 27 | |
| LOC_Os04g51190 | 461 | 2 | Growth-regulating factor 3 | Gibberellin (GA) | 18 | 13 | |
| LOC_Os03g47140 | 572 | 2 | Growth regulating factor protein 9 | 70 | 105 | ||
| osa-miR399 | LOC_Os04g55230 | 3898 | 2 | Tetratricopeptide repeat protein | Abscisic Acid (ABA) | 7 | 22 |
| LOC_Os09g39180 | 731 | 2 | Similar to Nucleic acid-binding protein precursor | Innate immunity Abscisic Acid (ABA) | 10 | 17 | |
| osa-miR414 | LOC_Os08g33370 | 725 | 2 | G-box factor 14-3-3 protein | Xoo, Magnaporthe oryzae/ Jasmonic acid (JA), Ethylene (ETH), Abscisic Acid (ABA) | 1 | 11 |
| osa-miR444 | LOC_Os02g49840 | 290 | 2 | MADS-box transcription factor 57 | 147 | 211 | |
| osa-miR528 | LOC_Os09g20090 | 32 | 2 | Ascorbate oxidase | Antiviral defence/ Salicylic Acid (SA) | 3 | 23 |
| LOC_Os07g38290 | 538 | 2 | Cupredoxin domain containing protein | 837 | 1780 | ||
| LOC_Os08g36420 | 80 | 2 | Zinc/iron permease family protein | 8 | 14 | ||
| osa-miR530-5p | LOC_Os04g51400 | 510 | 2 | RING finger protein with microtubule-targeting domain 1 | 605 | 1107 | |
| osa-miR5809 | LOC_Os12g25180 | 403 | 2 | Hypothetical conserved gene | 29 | 35 | |
| novel_mir112 | LOC_Os10g41590 | 1206 | 2 | Flowering-Related RING Protein 1 | Innate immunity | 5 | 16 |
| novel_mir89 | LOC_Os02g04680 | 1156 | 0 | Squamosa promoter binding-like 3 | 77 | 36 | |
| LOC_Os06g45310 | 760 | 0 | Squamosa promoter binding-like 11 | 14 | 41 | ||
| novel_mir115 | LOC_Os03g39610 | 743 | 0 | chlorophyll A-B binding protein | Reactive oxygen species Biosynthesis/ Abscisic Acid (ABA) | 85 | 70 |
| LOC_Os02g10390 | 706 | 0 | chlorophyll A-B binding protein | Cytokinin (CTK) | 61 | 59 | |
| LOC_Os01g52240 | 749 | 2 | chlorophyll A-B binding protein | 3158 | 4197 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jia, Y.; Li, C.; Li, Q.; Liu, P.; Liu, D.; Liu, Z.; Wang, Y.; Jiang, G.; Zhai, W. Characteristic Dissection of Xanthomonas oryzae pv. oryzae Responsive MicroRNAs in Rice. Int. J. Mol. Sci. 2020, 21, 785. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21030785
Jia Y, Li C, Li Q, Liu P, Liu D, Liu Z, Wang Y, Jiang G, Zhai W. Characteristic Dissection of Xanthomonas oryzae pv. oryzae Responsive MicroRNAs in Rice. International Journal of Molecular Sciences. 2020; 21(3):785. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21030785
Chicago/Turabian StyleJia, Yanfeng, Chunrong Li, Quanlin Li, Pengcheng Liu, Dongfeng Liu, Zhenzhen Liu, Yanyan Wang, Guanghuai Jiang, and Wenxue Zhai. 2020. "Characteristic Dissection of Xanthomonas oryzae pv. oryzae Responsive MicroRNAs in Rice" International Journal of Molecular Sciences 21, no. 3: 785. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21030785