Comparative Proteomics Profiling Illuminates the Fruitlet Abscission Mechanism of Sweet Cherry as Induced by Embryo Abortion

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
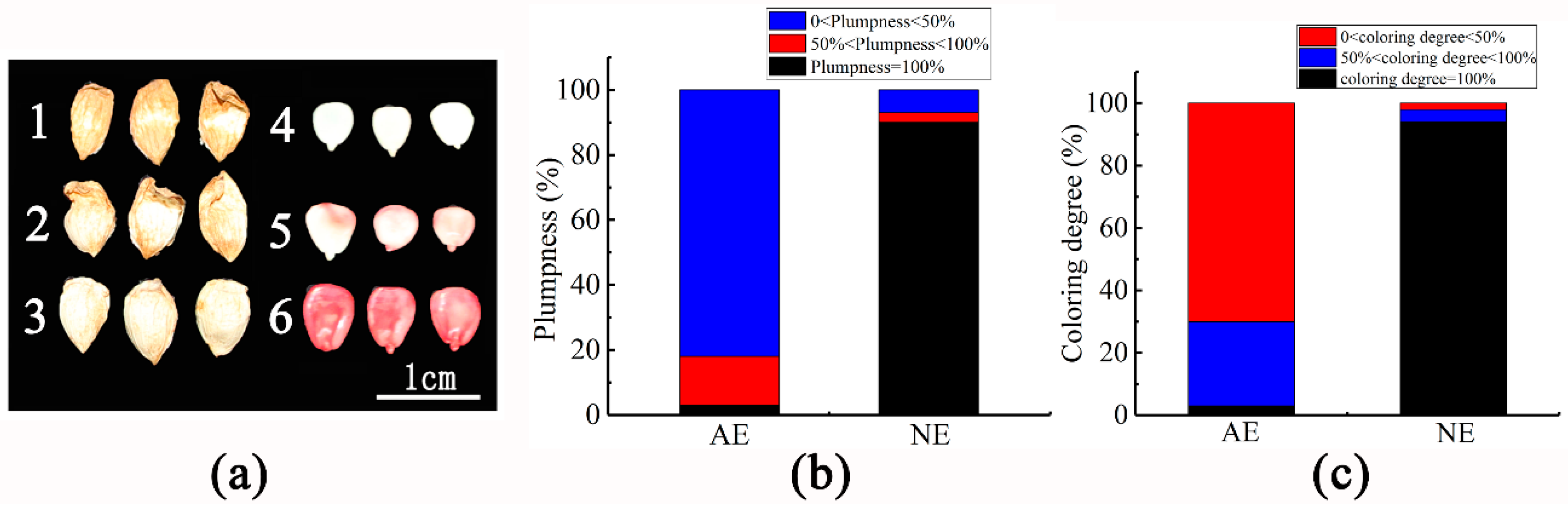
2.1. Embryo Vigor of Shedding Fruit
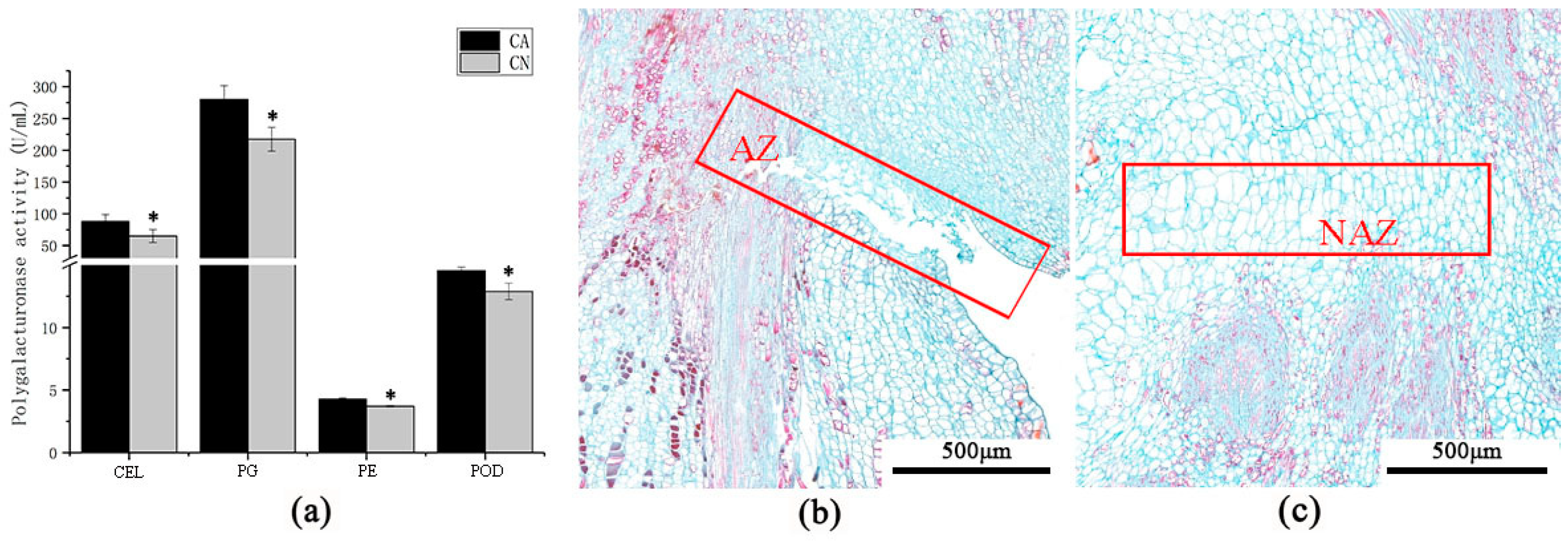
2.2. Enzyme Activity and Anatomical Structure of Carpopodium Abscission Zone
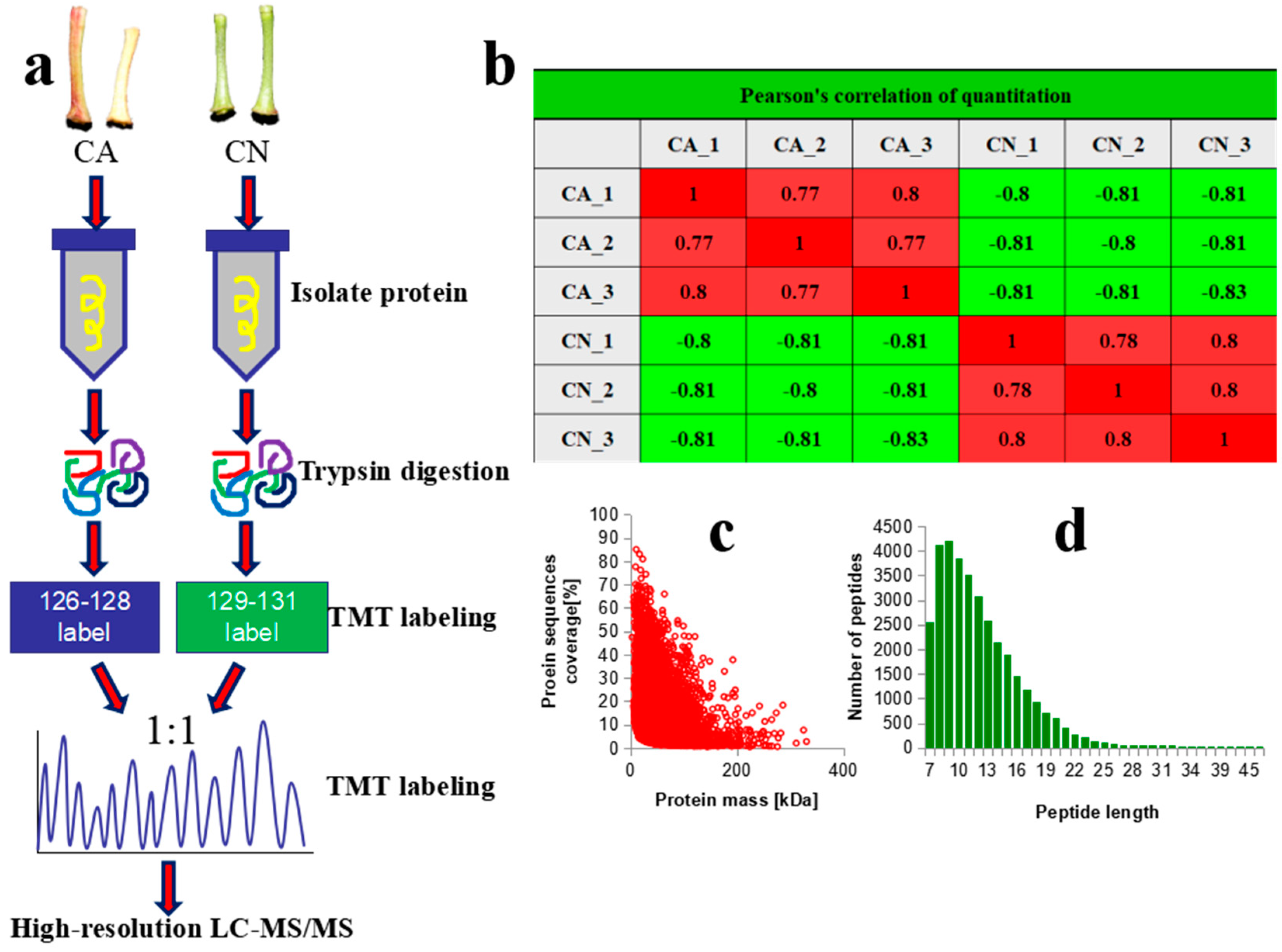
2.3. Quality Control and Quantitative Proteomic Analysis
2.4. Identification of DAPs During Carpopodium Abscission
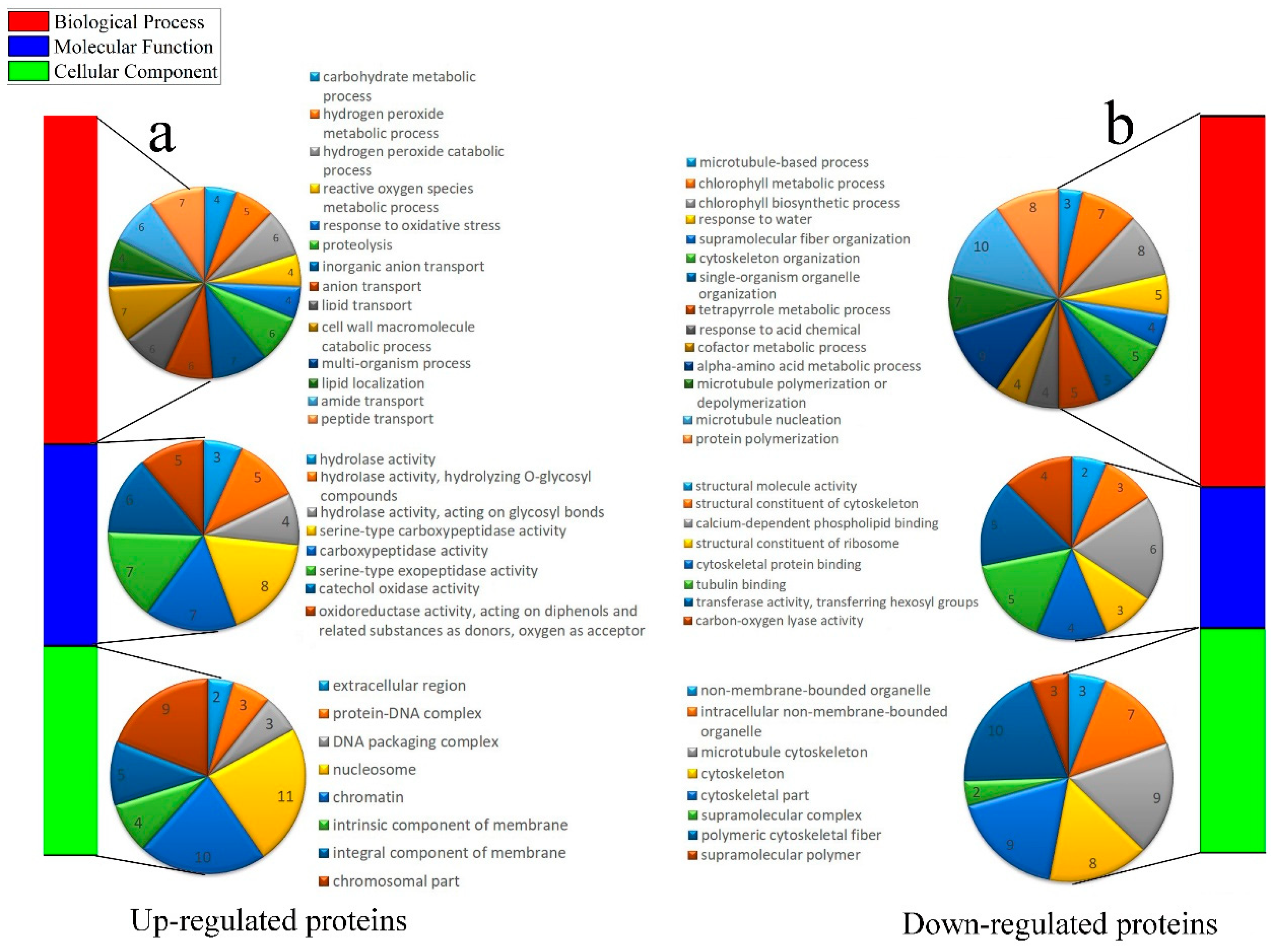
2.5. Enrichment Analysis of DAPs During Carpopodium Abscission
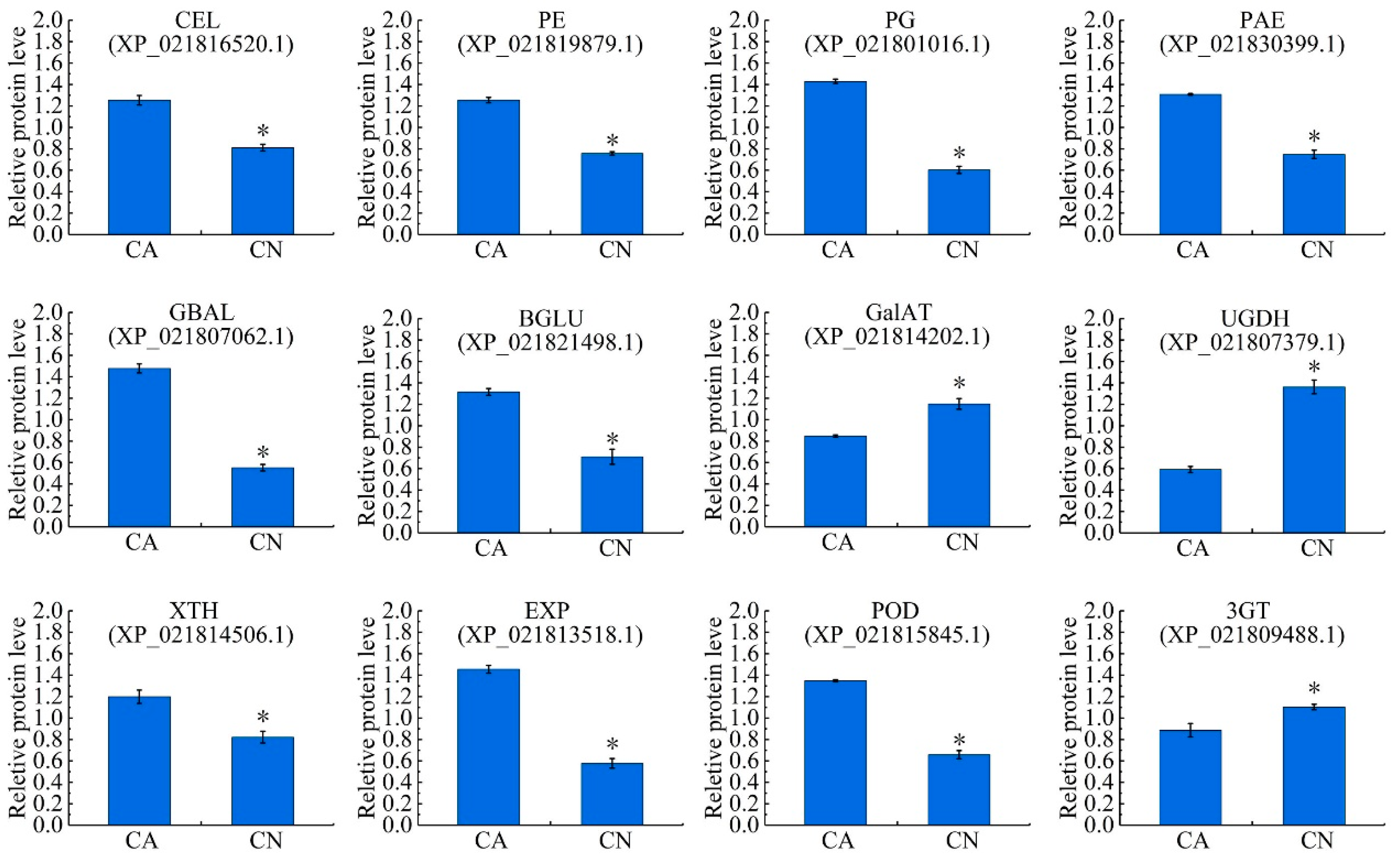
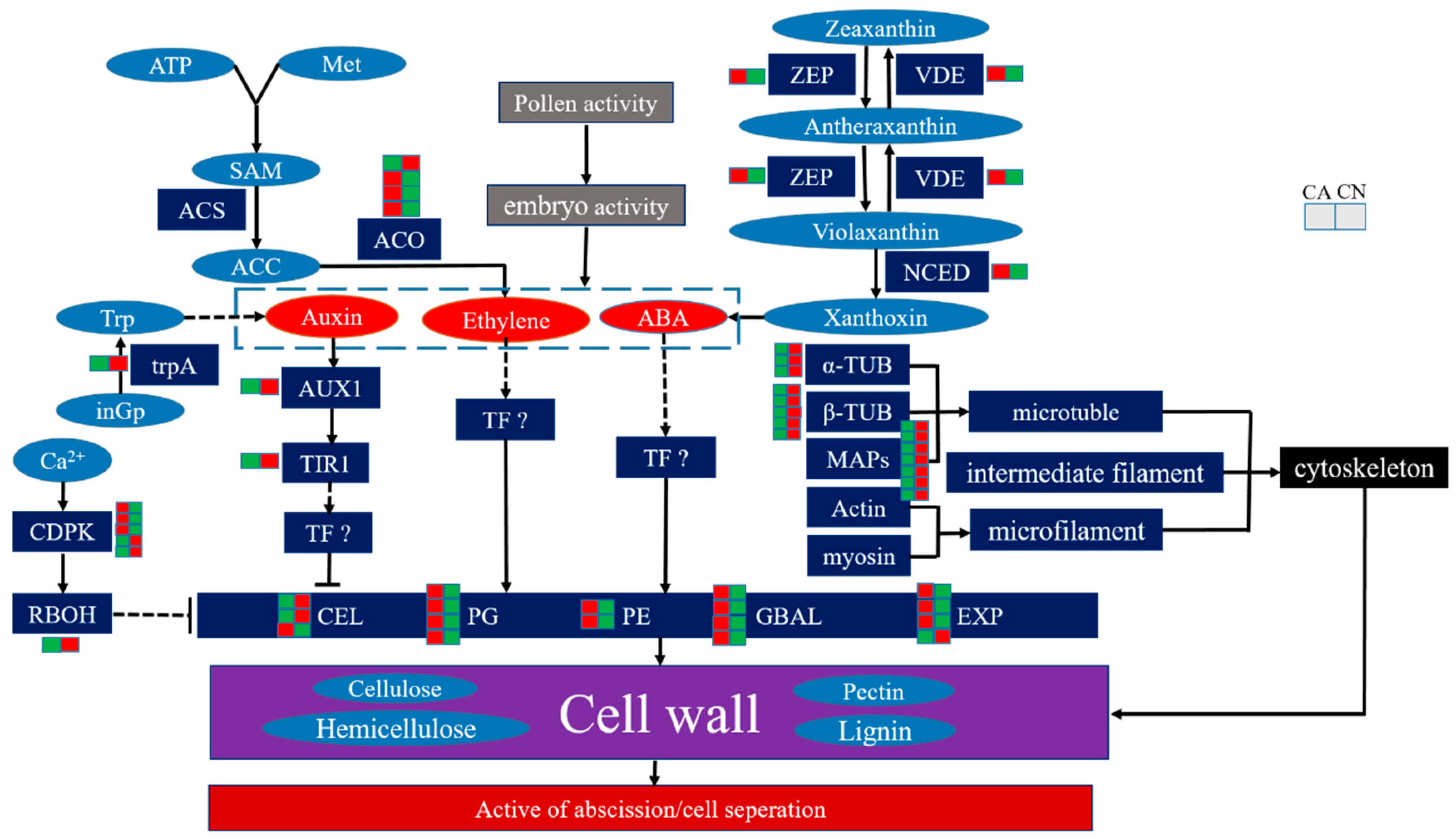
2.6. Biosynthesis of Cell Wall Modifying Proteins and Lignin
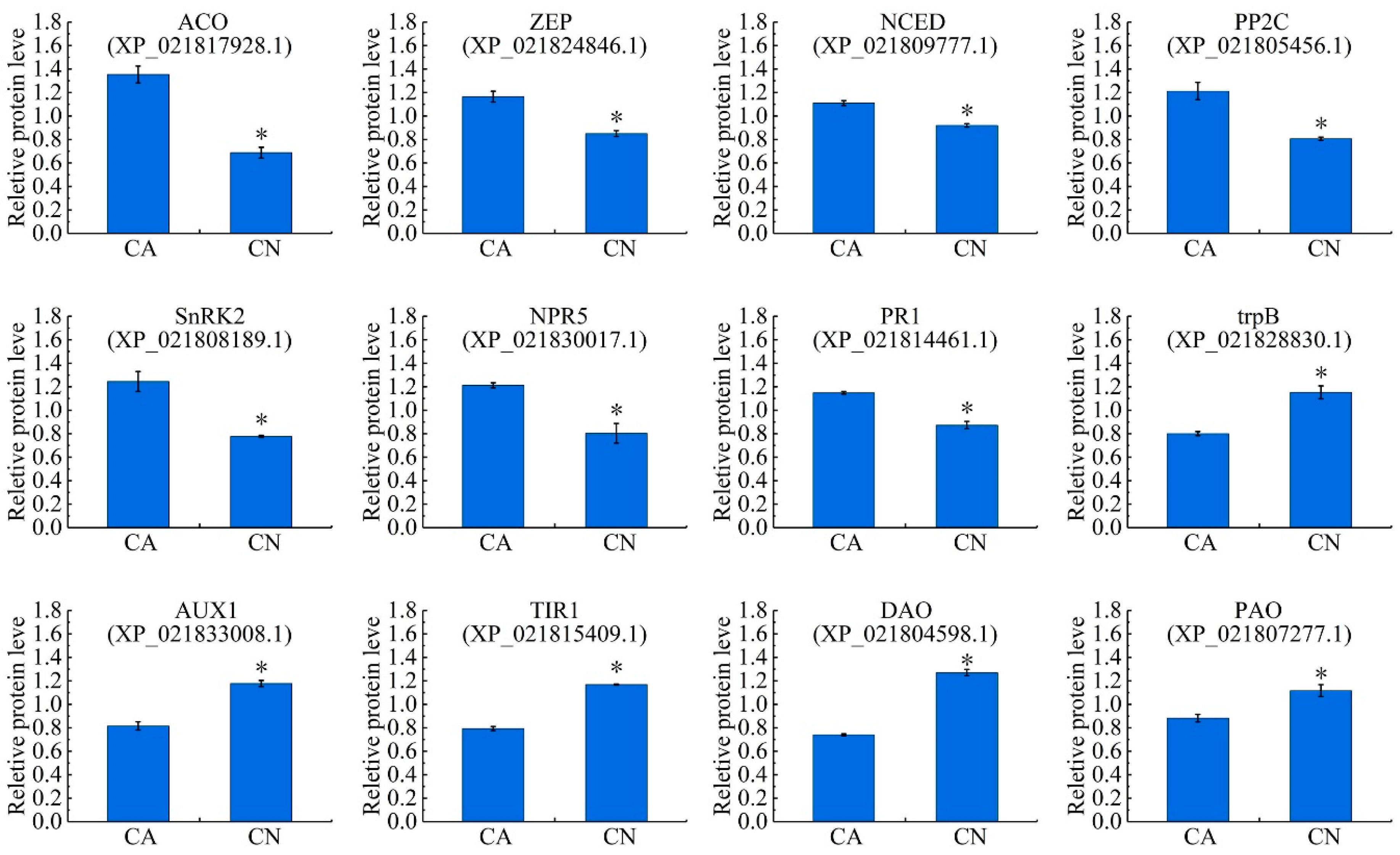
2.7. Plant Hormone Biosynthesis and Signal Transduction
2.8. Cytoskeleton and Transport Proteins
2.9. Transcription Factor
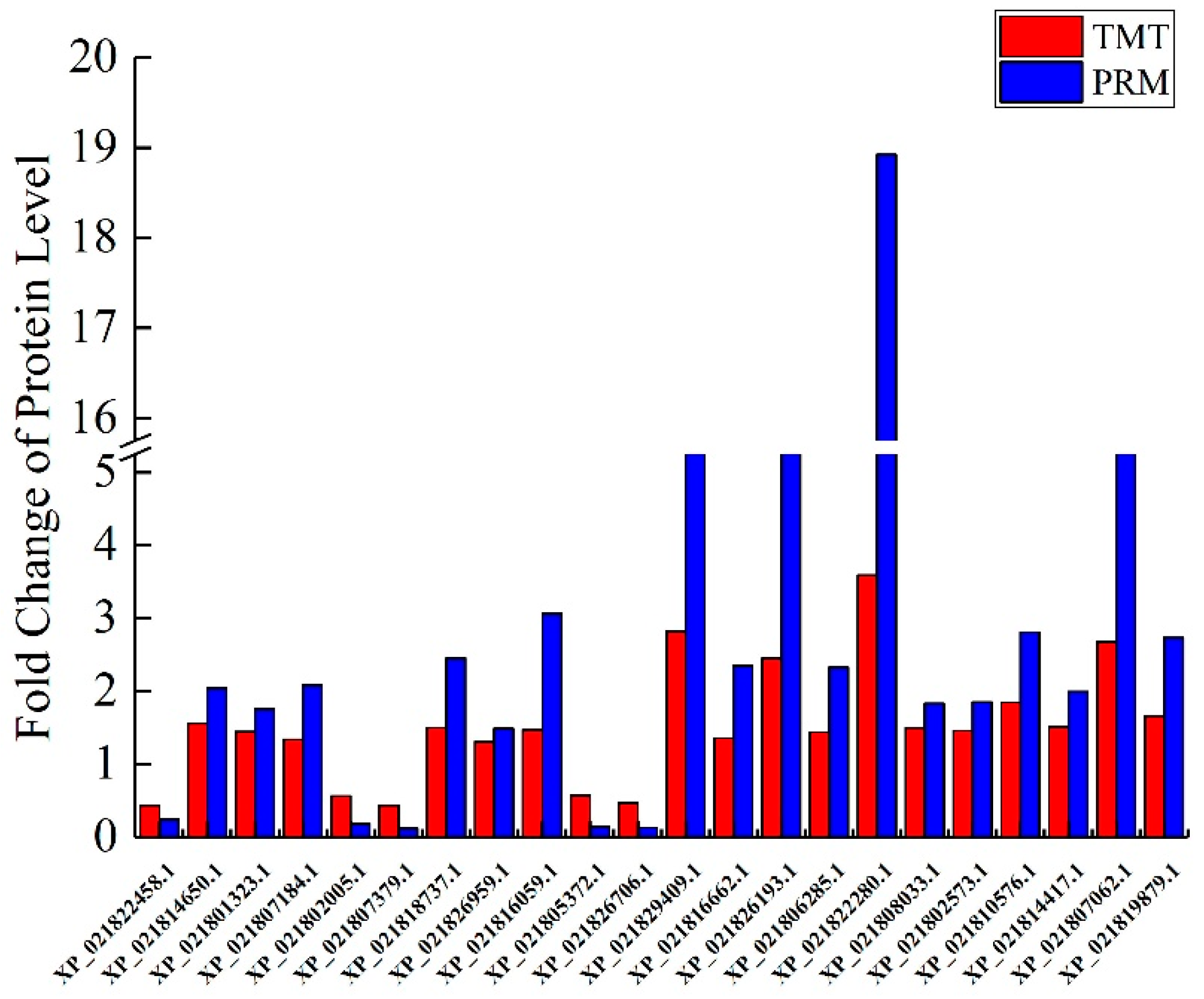
2.10. Validation of DAPs by PRM
3. Discussion
3.1. Embryo Abortion Leads to Fruit Abscission
3.2. Cell Wall Metabolism and Abscission
3.3. Lignin Biosynthesis and Abscission
3.4. Plant Hormone-Related Abscission
3.5. Cytoskeleton and Abscission
3.6. Transcription Factor Raleted Abscission
4. Materials and Methods
4.1. Plant Materials
4.2. Detection of Embryo Activity
4.3. Measurement of Enzyme Activity
4.4. Anatomical Observation of Carpopodium Abscission Zones.
4.5. Protein Extraction and Trypsin Digestion
4.6. TMT Labeling and HPLC Fractionation
4.7. LC-MS/MS Analysis
4.8. Database Search
4.9. Bioinformatics Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| 3GT | anthocyanidin 3-O-glucosyltransferase 5-like |
| ABA | abscisic acid |
| ACO | 1-aminocyclopropane-1-carboxylate oxidase |
| AE | Abscission embryo |
| AGC | Automatic gain control |
| ARF | Auxin response factor |
| Arp2/3 | Actin-related protein2/3 complex |
| AUX1 | auxin transporter-like protein 2 |
| AZ | Abscission Zone |
| BGLU | beta-glucosidase |
| bHLH | basic-helix-loop-helix |
| BP | biological process |
| bZIP | basic region/leucine zipper motif |
| CA | abscising carpopodium |
| CC | cellular component |
| CEL | cellulase |
| CN | non-abscising carpopodium |
| DAO | 2-oxoglutarate-dependent dioxygenase DAO-like |
| DAPs | Differential accumulated proteins |
| EXP | Expansin |
| FAA | Formalin-acetic acid-alcohol |
| FDR | False discovery rate |
| GalAT | galacturonosyltransferase |
| GBAL | β-galactosidase |
| GO | gene ontology |
| HPLC | High-Performance Liquid Chromatography |
| KEGG | The Kyoto Encyclopedia of Genes and Genomes |
| MF | molecular function |
| NE | Non-abscission embryo |
| NPR5 | regulatory protein NPR5-like |
| PAE | pectin acetylesterase |
| PE | pectinesterase |
| PG | polygalacturonase |
| POD | peroxidase |
| PP2C | protein phosphatase 2C |
| PR1 | pathogenesis-related protein 1 |
| PRM | Parallel Reaction Monitoring |
| PRM | Parallel Reaction Monitoring |
| SAMS | S-adenosylmethionine synthetase |
| SnRK2 | SNF1-related protein kinase 2 |
| TIR1 | transport inhibitor response 1 |
| TMT | Tandem Mass Tag |
| TMT-LC/MS | Tandem Mass Tag-liquid chromatograph-mass spectrometer |
| TTC | Triphenyte-trazoliumchloride |
| UGDH | UDP-glucose 6-dehydrogenase |
| XTH | xyloglucan endotransglucosylase/hydrolase protein |
| ZEF | zeaxanthin epoxidase |
References
- Wei, H.; Chen, X.; Zong, X.; Shu, H.; Gao, D.; Liu, Q. Comparative transcriptome analysis of genes involved in anthocyanin biosynthesis in the red and yellow fruits of sweet cherry (Prunus avium L.). PLoS ONE 2015, 10, 1–20. [Google Scholar] [CrossRef] [PubMed]
- Kühn, N.; Serrano, A.; Abello, C.; Arce, A.; Espinoza, C.; Gouthu, S.; Deluc, L.; Arce-Johnson, P. Regulation of polar auxin transport in grapevine fruitlets (Vitis vinifera L.) and the proposed role of auxin homeostasis during fruit abscission. BMC Plant Biol. 2016, 16, 1–17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sawicki, M.; Aït Barka, E.; Clément, C.; Vaillant-Gaveau, N.; Jacquard, C. Cross-talk between environmental stresses and plant metabolism during reproductive organ abscission. J. Exp. Bot. 2015, 66, 1707–1719. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Estornell, L.H.; Agustí, J.; Merelo, P.; Talón, M.; Tadeo, F.R. Elucidating mechanisms underlying organ abscission. Plant Sci. 2013, 199–200, 48–60. [Google Scholar] [CrossRef]
- Glazinska, P.; Wojciechowski, W.; Kulasek, M.; Glinkowski, W.; Marciniak, K.; Klajn, N.; Kesy, J.; Kopcewicz, J. De novo transcriptome profiling of flowers, flower pedicels and pods of lupinus luteus (Yellow lupine) reveals complex expression changes during organ abscission. Front. Plant Sci. 2017, 8. [Google Scholar] [CrossRef] [Green Version]
- Merelo, P.; Agustí, J.; Arbona, V.; Costa, M.L.; Estornell, L.H.; Gómez-Cadenas, A.; Coimbra, S.; Gómez, M.D.; Pérez-Amador, M.A.; Domingo, C.; et al. Cell wall remodeling in abscission zone cells during ethylene-promoted fruit abscission in citrus. Front. Plant Sci. 2017, 8. [Google Scholar] [CrossRef] [Green Version]
- Parra-Lobato, M.C.; Gomez-Jimenez, M.C. Polyamine-induced modulation of genes involved in ethylene biosynthesis and signalling pathways and nitric oxide production during olive mature fruit abscission. J. Exp. Bot. 2011, 62, 4447–4465. [Google Scholar] [CrossRef] [Green Version]
- Patterson, S.E. Cutting loose. Abscission and dehiscence in Arabidopsis. Plant Physiol. 2001, 126, 494–500. [Google Scholar] [CrossRef] [Green Version]
- Agustí, J.; Merelo, P.; Cercós, M.; Tadeo, F.R.; Talón, M. Ethylene-induced differential gene expression during abscission of citrus leaves. J. Exp. Bot. 2008, 59, 2717–2733. [Google Scholar] [CrossRef] [Green Version]
- Nakano, T.; Fujisawa, M.; Shima, Y.; Ito, Y. Expression profiling of tomato pre-abscission pedicels provides insights into abscission zone properties including competence to respond to abscission signals. BMC Plant Biol. 2013, 13, 1. [Google Scholar] [CrossRef] [Green Version]
- Nakano, T.; Kato, H.; Shima, Y.; Ito, Y. Apple SVP family MADS-box proteins and the tomato pedicel abscission zone regulator JOINTLESS have similar molecular activities. Plant Cell Physiol. 2015, 56, 1097–1106. [Google Scholar] [CrossRef] [Green Version]
- Fu, X.; Shi, Z.; Jiang, Y.; Jiang, L.; Qi, M.; Xu, T.; Li, T. A family of auxin conjugate hydrolases from Solanum lycopersicum and analysis of their roles in flower pedicel abscission. BMC Plant Biol. 2019, 19, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Ellis, C.M.; Nagpal, P.; Young, J.C.; Hagen, G.; Guilfoyle, T.J.; Reed, J.W. AUXIN RESPONSE FACTOR1 and AUXIN RESPONSE FACTOR2 regulate senescence and floral organ abscission in Arabidopsis thaliana. Development 2005, 132, 4563–4574. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, X.L.; Qi, M.F.; Xu, T.; Lu, X.J.; Li, T.L. Proteomics profiling of ethylene-induced tomato flower pedicel abscission. J. Proteom. 2015, 121, 67–87. [Google Scholar] [CrossRef] [PubMed]
- Tsuchiya, M.; Satoh, S.; Iwai, H. Distribution of XTH, expansin, and secondary-wall-related CesA in floral and fruit abscission zones during fruit development in tomato (Solanum lycopersicum). Front. Plant Sci. 2015, 6, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Li, C.; Ma, X.; Huang, X.; Wang, H.; Wu, H.; Zhao, M.; Li, J. Involvement of HD-ZIP I transcription factors LcHB2 and LcHB3 in fruitlet abscission by promoting transcription of genes related to the biosynthesis of ethylene and ABA in litchi. Tree Physiol. 2019, 510642. [Google Scholar] [CrossRef]
- Qi, M.F.; Xu, T.; Chen, W.Z.; Li, T.L. Ultrastructural localization of polygalacturonase in ethylene-stimulated abscission of tomato pedicel explants. Sci. World J. 2014, 2014. [Google Scholar] [CrossRef] [Green Version]
- Taylor, J.E.; Whitelaw, C.A. Signals in abscission. New Phytol. 2001, 151, 323–340. [Google Scholar] [CrossRef]
- Meir, S.; Hunter, D.A.; Chen, J.; Halaly, V.; Reid, M.S. Molecular Changes Occurring during Acquisition of Abscission Competence following Auxin Depletion in Mirabilis jalapa. Plant Physiol. 2006, 141, 1604–1616. [Google Scholar] [CrossRef] [Green Version]
- Sundaresan, S.; Philosoph-Hadas, S.; Riov, J.; Belausov, E.; Kochanek, B.; Tucker, M.L.; Meir, S. Abscission of flowers and floral organs is closely associated with alkalization of the cytosol in abscission zone cells. J. Exp. Bot. 2015, 66, 1355–1368. [Google Scholar] [CrossRef] [Green Version]
- Mishra, A.; Khare, S.; Trivedi, P.K.; Nath, P. Effect of ethylene, 1-MCP, ABA and IAA on break strength, cellulase and polygalacturonase activities during cotton leaf abscission. South African J. Bot. 2008, 74, 282–287. [Google Scholar] [CrossRef] [Green Version]
- Gao, Y.; Liu, C.; Li, X.; Xu, H.; Liang, Y.; Ma, N.; Fei, Z.; Gao, J.; Jiang, C.Z.; Ma, C. Transcriptome profiling of petal abscission zone and functional analysis of an Aux/IAA family gene RhiAA16 involved in petal shedding in rose. Front. Plant Sci. 2016, 7, 1–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sundaresan, S.; Philosoph-Hadas, S.; Riov, J.; Mugasimangalam, R.; Kuravadi, N.A.; Kochanek, B.; Salim, S.; Tucker, M.L.; Meir, S. De novo transcriptome sequencing and development of abscission zone-specific microarray as a new molecular tool for analysis of tomato organ abscission. Front. Plant Sci. 2016, 6. [Google Scholar] [CrossRef] [Green Version]
- Roberts, J.A.; Gonzalez-Carranza, Z.H. Pectinase functions in abscission. Stewart Postharvest Rev. 2009, 5, 1–4. [Google Scholar] [CrossRef]
- Poovaiah, B.W.; Rasmussen, H.P. Calcium Distribution in the Abscission Zone of Bean Leaves. Plant Physiol. 1973, 52, 683–684. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Patterson, S.E.; Bleecker, A.B. Ethylene-Dependent and -Independent Processes Associated with Floral Organ Abscission in Arabidopsis. Plant Physiol. 2004, 134, 194–203. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, J.H.; Liu, J.J.; Chen, K.L.; Li, H.W.; He, J.; Guan, B.; He, L. Comparative transcriptome and proteome profiling of two Citrus sinensis cultivars during fruit development and ripening. BMC Genom. 2017, 18, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Zhang, S.; Zhang, D.; Fan, S.; Du, L.; Shen, Y.; Xing, L.; Li, Y.; Ma, J.; Han, M. Effect of exogenous GA3 and its inhibitor paclobutrazol on floral formation, endogenous hormones, and flowering-associated genes in ‘Fuji’ apple (Malus domestica Borkh.). Plant Physiol. Biochem. 2016, 107, 178–186. [Google Scholar] [CrossRef]
- Li, J.M.; Huang, X.S.; Li, L.T.; Zheng, D.M.; Xue, C.; Zhang, S.L.; Wu, J. Proteome analysis of pear reveals key genes associated with fruit development and quality. Planta 2015, 241, 1363–1379. [Google Scholar] [CrossRef]
- Chan, Z.; Wang, Q.; Xu, X.; Meng, X.; Qin, G.; Li, B.; Tian, S. Functions of defense-related proteins and dehydrogenases in resistance response induced by salicylic acid in sweet cherry fruits at different maturity stages. Proteomics 2008, 8, 4791–4807. [Google Scholar] [CrossRef]
- Bargiela, R.; Herbst, F.A.; Martínez-Martínez, M.; Seifert, J.; Rojo, D.; Cappello, S.; Genovese, M.; Crisafi, F.; Denaro, R.; Chernikova, T.N.; et al. Metaproteomics and metabolomics analyses of chronically petroleum-polluted sites reveal the importance of general anaerobic processes uncoupled with degradation. Proteomics 2015, 15, 3508–3520. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsuchiya, H.; Tanaka, K.; Saeki, Y. The parallel reaction monitoring method contributes to a highly sensitive polyubiquitin chain quantification. Biochem. Biophys. Res. Commun. 2013, 436, 223–229. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, Q.; Liu, B.; Ruan, D.; Niu, C.; Shen, J.; Ni, M.; Cong, W.; Lu, X.; Jin, L. A novel targeted proteomics method for identification and relative quantitation of difference in nitration degree of OGDH between healthy and diabetic mouse. Proteomics 2014, 14, 2417–2426. [Google Scholar] [CrossRef] [PubMed]
- Dong, H.; Li, Y.; Fan, H.; Zhou, D.; Li, H. Quantitative proteomics analysis reveals resistance differences of banana cultivar ‘Brazilian’ to Fusarium oxysporum f. sp. cubense races 1 and 4. J. Proteom. 2019, 203, 103376. [Google Scholar] [CrossRef]
- Ferrero, S.; Carretero-Paulet, L.; Mendes, M.A.; Botton, A.; Eccher, G.; Masiero, S.; Colombo, L. Transcriptomic signatures in seeds of apple (Malus domestica L. Borkh) during fruitlet abscission. PLoS ONE 2015, 10, 1–15. [Google Scholar] [CrossRef] [Green Version]
- He, J.H.; Ma, F.W.; Chen, Y.Y.; Shu, H.R. Differentially expressed genes implicated in embryo abortion of mango identified by suppression subtractive hybridization. Genet. Mol. Res. 2012, 11, 3966–3974. [Google Scholar] [CrossRef]
- Blanusa, T.; Else, M.A.; Atkinson, C.J.; Davies, W.J. The regulation of sweet cherry fruit abscission by polar auxin transport. Plant Growth Regul. 2005, 45, 189–198. [Google Scholar] [CrossRef]
- Blanusa, T.; Else, M.A.; Davies, W.J.; Atkinson, C.J. Regulation of sweet cherry fruit abscission: The role of photo-assimilation, sugars and abscisic acid. J. Horticultural Sci. Biotechnol. 2006, 81, 613–620. [Google Scholar] [CrossRef]
- Mesejo, C.; Muñoz-Fambuena, N.; Reig, C.; Martínez-Fuentes, A.; Agustí, M. Cell division interference in newly fertilized ovules induces stenospermocarpy in cross-pollinated citrus fruit. Plant Sci. 2014, 225, 86–94. [Google Scholar] [CrossRef]
- Ben-Cheikh, W.; Perez-Botella, J.; Tadeo, F.R.; Talon, M.; Primo-Millo, E. Pollination increases gibberellin levels in developing ovaries of seeded varieties of citrus. Plant Physiol. 1997, 114, 557–564. [Google Scholar] [CrossRef] [Green Version]
- Tokuji, Y.; Kuriyama, K. Involvement of gibberellin and cytokinin in the formation of embryogenic cell clumps in carrot (Daucus carota). J. Plant Physiol. 2003, 160, 133–141. [Google Scholar] [CrossRef] [PubMed]
- Pellan-Delourme, R.; Renard, M. Cytoplasmic male sterility in rapeseed (Brassica napus L.): Female fertility of restored rapeseed with “Ogura” and cybrids cytoplasms. Genome 1988, 30, 234–238. [Google Scholar] [CrossRef]
- Lal, N.; Kumar Gupta, A.; Nath, V. Fruit Retention in Different Litchi Germplasm Influenced by Temperature. Int. J. Current Microbiol. Appl. Sci. 2017, 6, 1189–1194. [Google Scholar] [CrossRef]
- Gunawardena, A.H.L.A.N.; Greenwood, J.S.; Dengler, N.G. Cell wall degradation and modification during programmed cell death in lace plant, Aponogeton madagascariensis (Aponogetonaceae). Amer. J. Bot. 2007, 94, 1116–1128. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Giné-Bordonaba, J.; Echeverria, G.; Ubach, D.; Aguiló-Aguayo, I.; López, M.L.; Larrigaudière, C. Biochemical and physiological changes during fruit development and ripening of two sweet cherry varieties with different levels of cracking tolerance. Plant Physiol. Biochem. 2017, 111, 216–225. [Google Scholar] [CrossRef]
- Kim, J.; Sundaresan, S.; Philosoph-Hadas, S.; Yang, R.; Meir, S.; Tucker, M.L. Examination of the abscission-associated transcriptomes for soybean, tomato, and arabidopsis highlights the conserved biosynthesis of an extensible extracellular matrix and boundary layer. Front. Plant Sci. 2015, 6, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Goldental-Cohen, S.; Burstein, C.; Biton, I.; Ben Sasson, S.; Sadeh, A.; Many, Y.; Doron-Faigenboim, A.; Zemach, H.; Mugira, Y.; Schneider, D.; et al. Ethephon induced oxidative stress in the olive leaf abscission zone enables development of a selective abscission compound. BMC Plant Biol. 2017, 17, 1–17. [Google Scholar] [CrossRef]
- Ke, X.; Wang, H.; Li, Y.; Zhu, B.; Zang, Y.; He, Y.; Cao, J.; Zhu, Z.; Yu, Y. Genome-wide identification and analysis of polygalacturonase genes in solanum lycopersicum. Int. J. Mol. Sci. 2018, 19, 2290. [Google Scholar] [CrossRef] [Green Version]
- Gao, Y.; Liu, Y.; Liang, Y.; Lu, J.; Jiang, C.; Fei, Z.; Jiang, C.Z.; Ma, C.; Gao, J. Rosa hybrida RhERF1 and RhERF4 mediate ethylene- and auxin-regulated petal abscission by influencing pectin degradation. Plant J. 2019, 99, 1159–1171. [Google Scholar] [CrossRef]
- Cosgrove, D.J. Enzymes and Other Agents That Enhance Cell Wall Extensibility. Annu. Rev. Plant Physiol. Plant Mol. Biol. 1999, 50, 391–417. [Google Scholar] [CrossRef] [Green Version]
- Vissenberg, K.; Fry, S.C.; Verbelen, J.P. Root hair initiation is coupled to a highly localized increase of xyloglucan endotransglycosylase action in arabidopsis roots. Plant Physiol. 2001, 127, 1125–1135. [Google Scholar] [CrossRef] [PubMed]
- Carmen, C.; Jocelyn, R.K.C.; Alan, B.B. Auxin regulation and spatial localization of an endo-1,4-beta-D-glucanase and a xyloglucan endotransglycosylase in expanding tomato hypocotyls. Plant J. 1997, 12, 417–426. [Google Scholar] [CrossRef] [Green Version]
- Van Nocker, S. Development of the abscission zone. Stewart Postharvest Rev. 2009, 5. [Google Scholar] [CrossRef]
- Liljegren, S.J.; Ditta, G.S.; Eshed, Y.; Savidge, B.; Bowmant, J.L.; Yanofsky, M.F. SHATTERPROOF MADS-box genes control dispersal in Arabidopsis. Nature 2000, 404, 766–770. [Google Scholar] [CrossRef]
- Smékalová, V.; Doskočilová, A.; Komis, G.; Šamaj, J. Crosstalk between secondary messengers, hormones and MAPK modules during abiotic stress signalling in plants. Biotechnol. Adv. 2014, 32, 2–11. [Google Scholar] [CrossRef]
- Nakano, T.; Ito, Y. Molecular mechanisms controlling plant organ abscission. Plant Biotechnol. 2013, 30, 209–216. [Google Scholar] [CrossRef] [Green Version]
- Einhorn, T.C.; Arrington, M. ABA and Shading Induce ‘Bartlett’ Pear Abscission and Inhibit Photosynthesis but Are Not Additive. J. Plant Growth Regul. 2018, 37, 300–308. [Google Scholar] [CrossRef]
- Shen, C.J.; Bai, Y.H.; Wang, S.K.; Zhang, S.N.; Wu, Y.R.; Chen, M.; Jiang, D.A.; Qi, Y.H. Expression profile of PIN, AUX/LAX and PGP auxin transporter gene families in Sorghum bicolor under phytohormone and abiotic stress. FEBS J. 2010, 277, 2954–2969. [Google Scholar] [CrossRef]
- Goto, D.B.; Ogi, M.; Kijima, F.; Kumagai, T.; Van Werven, F.; Onouchi, H.; Naito, S. A single-nucleotide mutation in a gene encoding S-adenosylmethionine synthetase is associated with methionine over-accumulation phenotype in Arabidopsis thaliana. Genes Genet. Syst. 2002, 77, 89–95. [Google Scholar] [CrossRef] [Green Version]
- Chersicola, M.; Kladnik, A.; Žnidarič, M.T.; Mrak, T.; Gruden, K.; Dermastia, M. 1-Aminocyclopropane-1-Carboxylate Oxidase Induction in Tomato Flower Pedicel Phloem and Abscission Related Processes Are Differentially Sensitive To Ethylene. Front. Plant Sci. 2017, 8, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Daher, F.B.; Braybrook, S.A. How to let go: Pectin and plant cell adhesion. Front. Plant Sci. 2015, 6, 1–8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Higgs, H.N.; Pollard, T.D. Regulation of Actin Filament Network Formation Through ARP2/3 Complex: Activation by a Diverse Array of Proteins. Annual Rev. Biochem. 2001, 70, 649–676. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Du, M.; Tian, X.; Xu, D.; Li, Z. Study on the changes of micr otubule cytoskeleton of abscission zone dur ing leaf abscission in cotton (in chinese). J. Shihezi Univ. 2016, 34, 1–7. [Google Scholar] [CrossRef]
- Rajangam, A.S.; Kumar, M.; Aspeborg, H.; Guerriero, G.; Arvestad, L.; Pansri, P.; Brown, C.J.L.; Hober, S.; Blomqvist, K.; Divne, C.; et al. MAP20, a microtubule-associated protein in the secondary cell walls of hybrid aspen, is a target of the cellulose synthesis inhibitor 2,6-dichlorobenzonitrile. Plant Physiol. 2008, 148, 1283–1294. [Google Scholar] [CrossRef] [Green Version]
- Wing, R.A.; Mao, L.; Begum, D.; Chuang, H.; Budiman, M.A.; Szymkowiak, E.J.; Irish, E.E. JOINTLESS is a MADS-box gene controlling tomato flower abscission zone development. Nature 2000, 406, 910–913. [Google Scholar] [CrossRef]
- Xie, Q.; Hu, Z.; Zhu, Z.; Dong, T.; Zhao, Z.; Cui, B.; Chen, G. Overexpression of a novel MADS-box gene SlFYFL delays senescence, fruit ripening and abscission in tomato. Sci. Rep. 2014, 4, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Li, C.; Zhao, M.; Ma, X.; Wen, Z.; Ying, P.; Peng, M.; Ning, X.; Xia, R.; Wu, H.; Li, J. The HD-Zip transcription factor LcHB2 regulates litchi fruit abscission through the activation of two cellulase genes. J. Exp. Bot. 2019, 70, 5189–5203. [Google Scholar] [CrossRef] [Green Version]








© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Qiu, Z.-L.; Wen, Z.; Yang, K.; Tian, T.; Qiao, G.; Hong, Y.; Wen, X.-P. Comparative Proteomics Profiling Illuminates the Fruitlet Abscission Mechanism of Sweet Cherry as Induced by Embryo Abortion. Int. J. Mol. Sci. 2020, 21, 1200. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21041200
Qiu Z-L, Wen Z, Yang K, Tian T, Qiao G, Hong Y, Wen X-P. Comparative Proteomics Profiling Illuminates the Fruitlet Abscission Mechanism of Sweet Cherry as Induced by Embryo Abortion. International Journal of Molecular Sciences. 2020; 21(4):1200. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21041200
Chicago/Turabian StyleQiu, Zhi-Lang, Zhuang Wen, Kun Yang, Tian Tian, Guang Qiao, Yi Hong, and Xiao-Peng Wen. 2020. "Comparative Proteomics Profiling Illuminates the Fruitlet Abscission Mechanism of Sweet Cherry as Induced by Embryo Abortion" International Journal of Molecular Sciences 21, no. 4: 1200. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21041200