Tinkering Cis Motifs Jigsaw Puzzle Led to Root-Specific Drought-Inducible Novel Synthetic Promoters


 ,
,
Abstract
:1. Introduction
2. Results
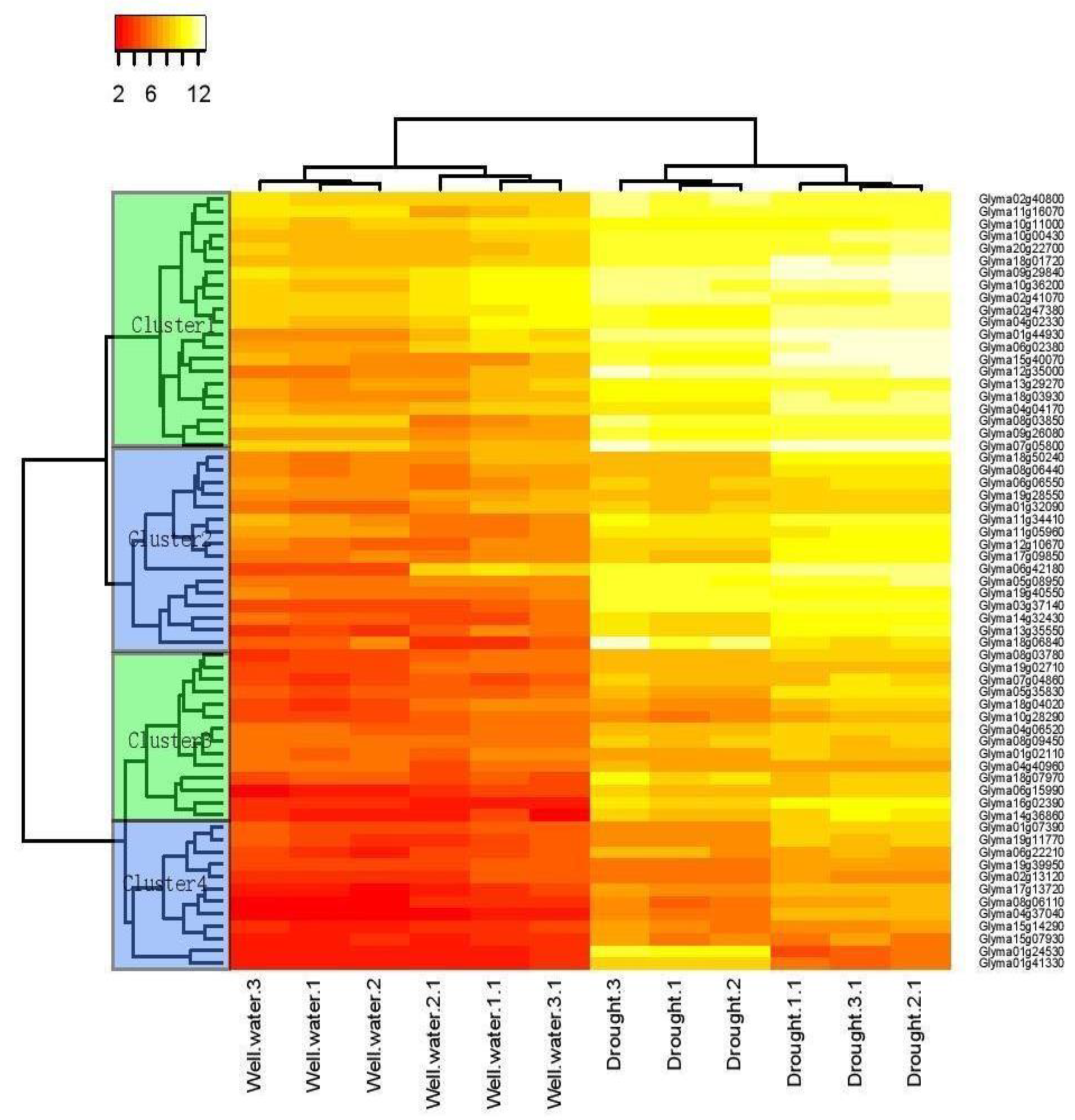
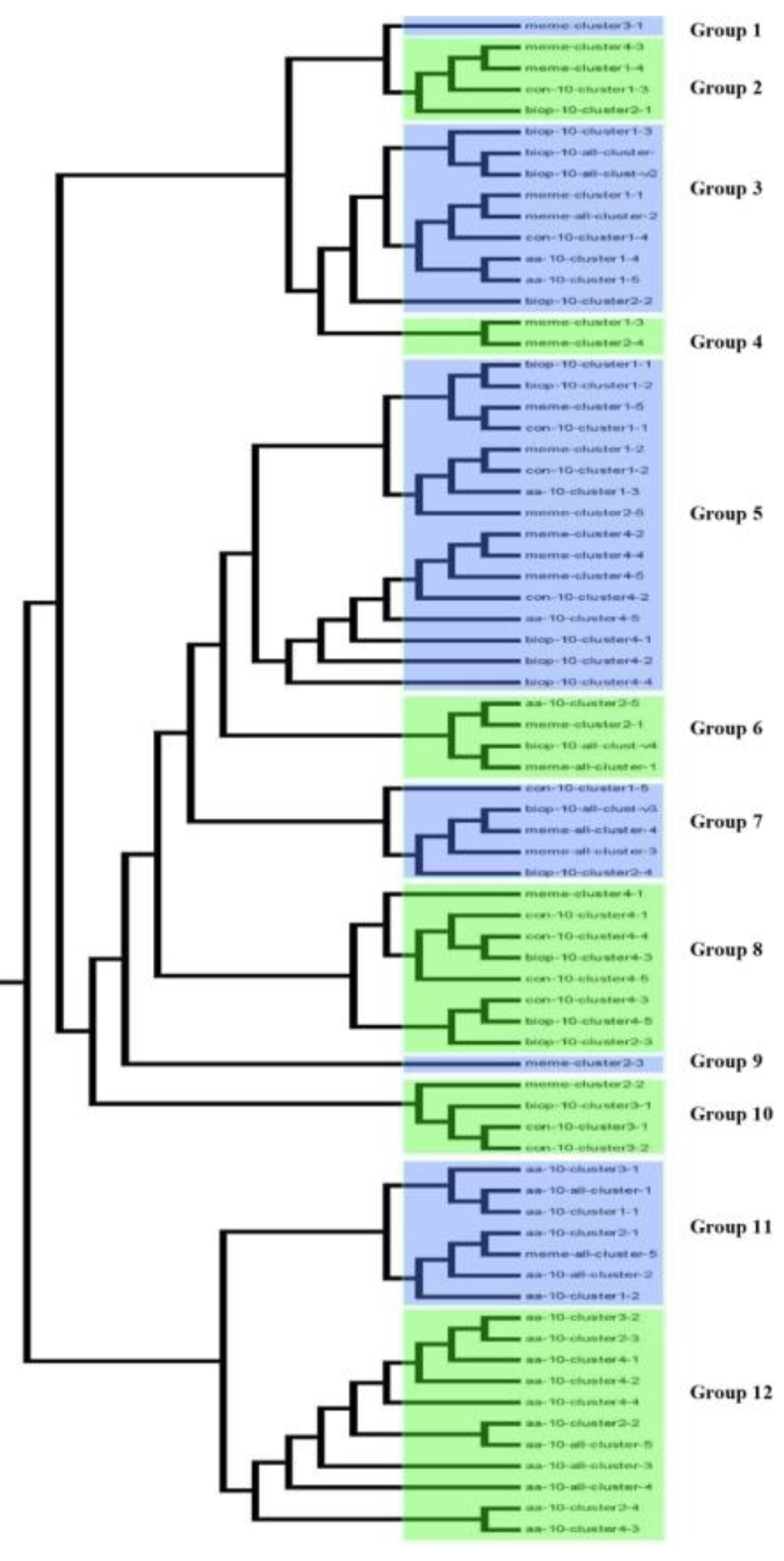
2.1. Transcriptome Analyses Determined Drought-Inducible Cis Motifs
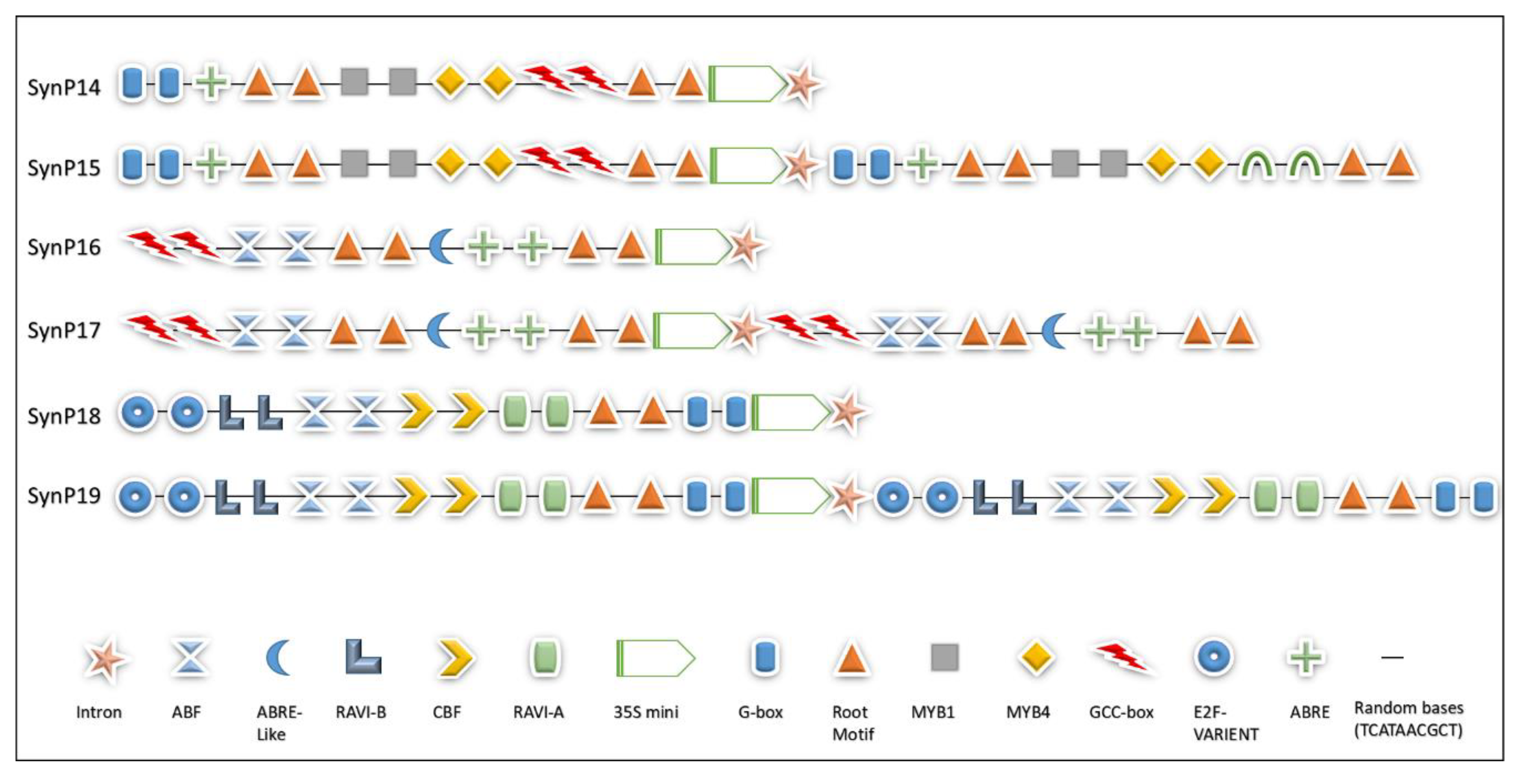
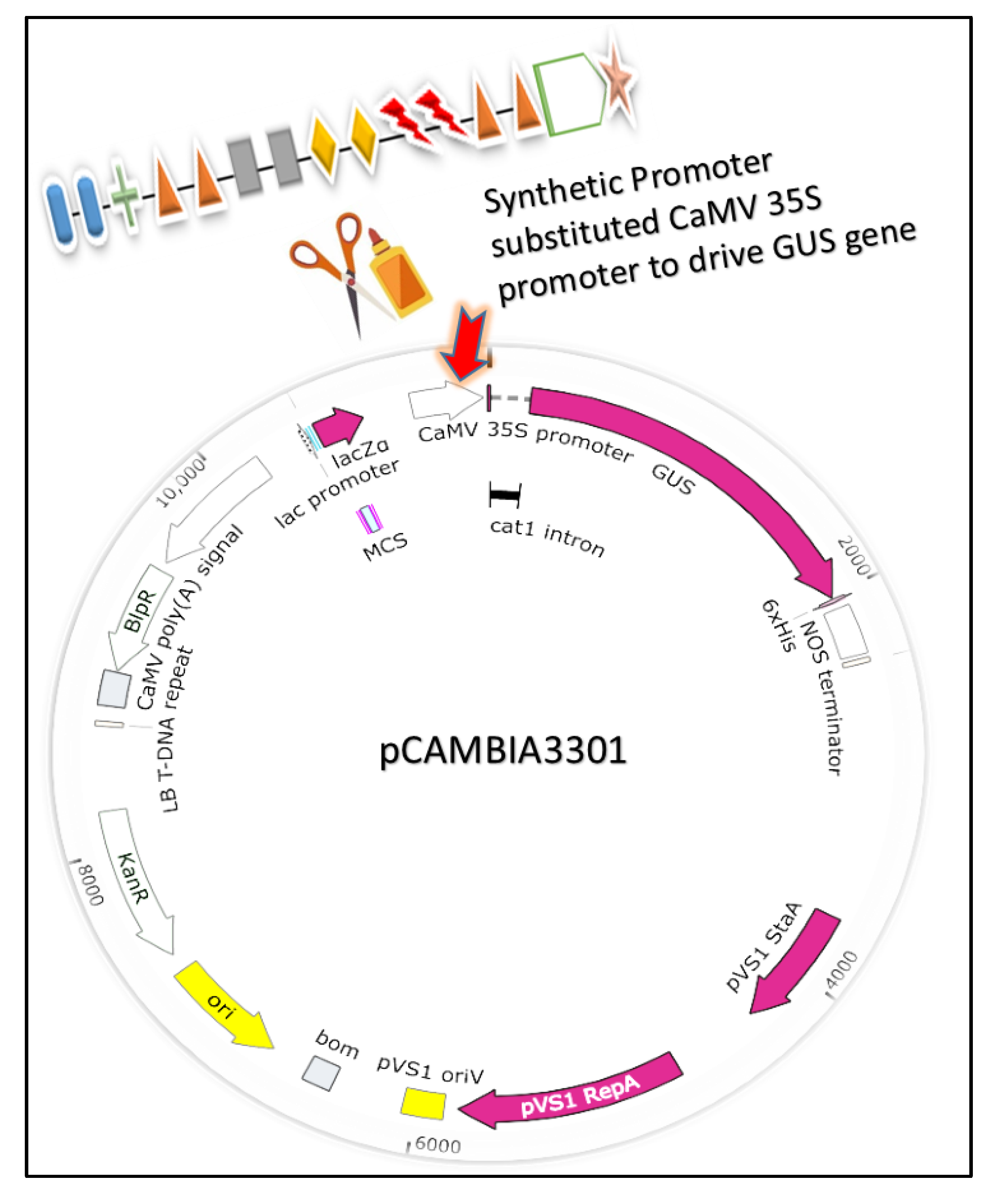
2.2. Motifs Selection and Design of Synthetic Modules
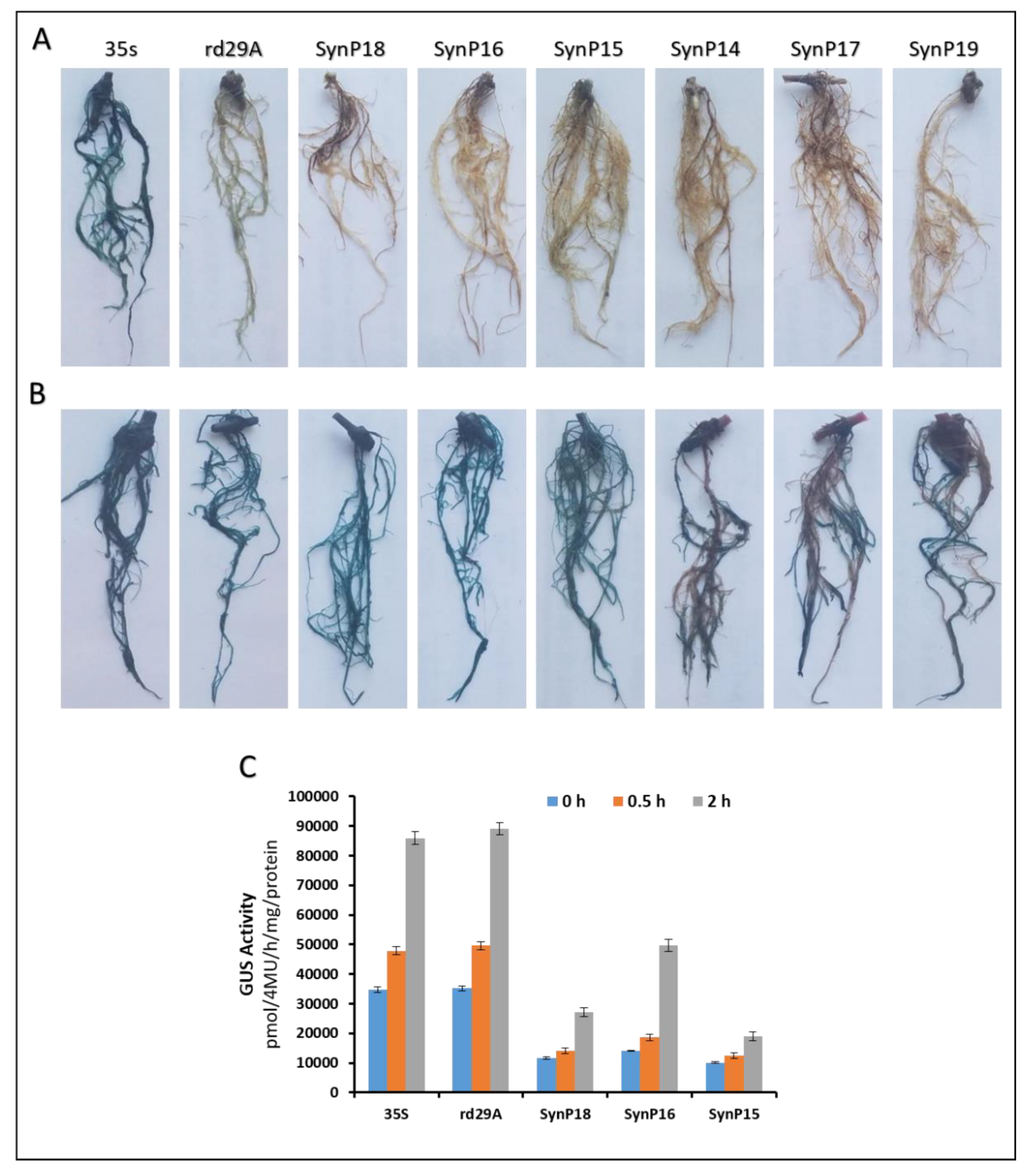
2.3. Agrobacterium-Mediated Transformation of Soybean Hairy Roots and GUS Analyses
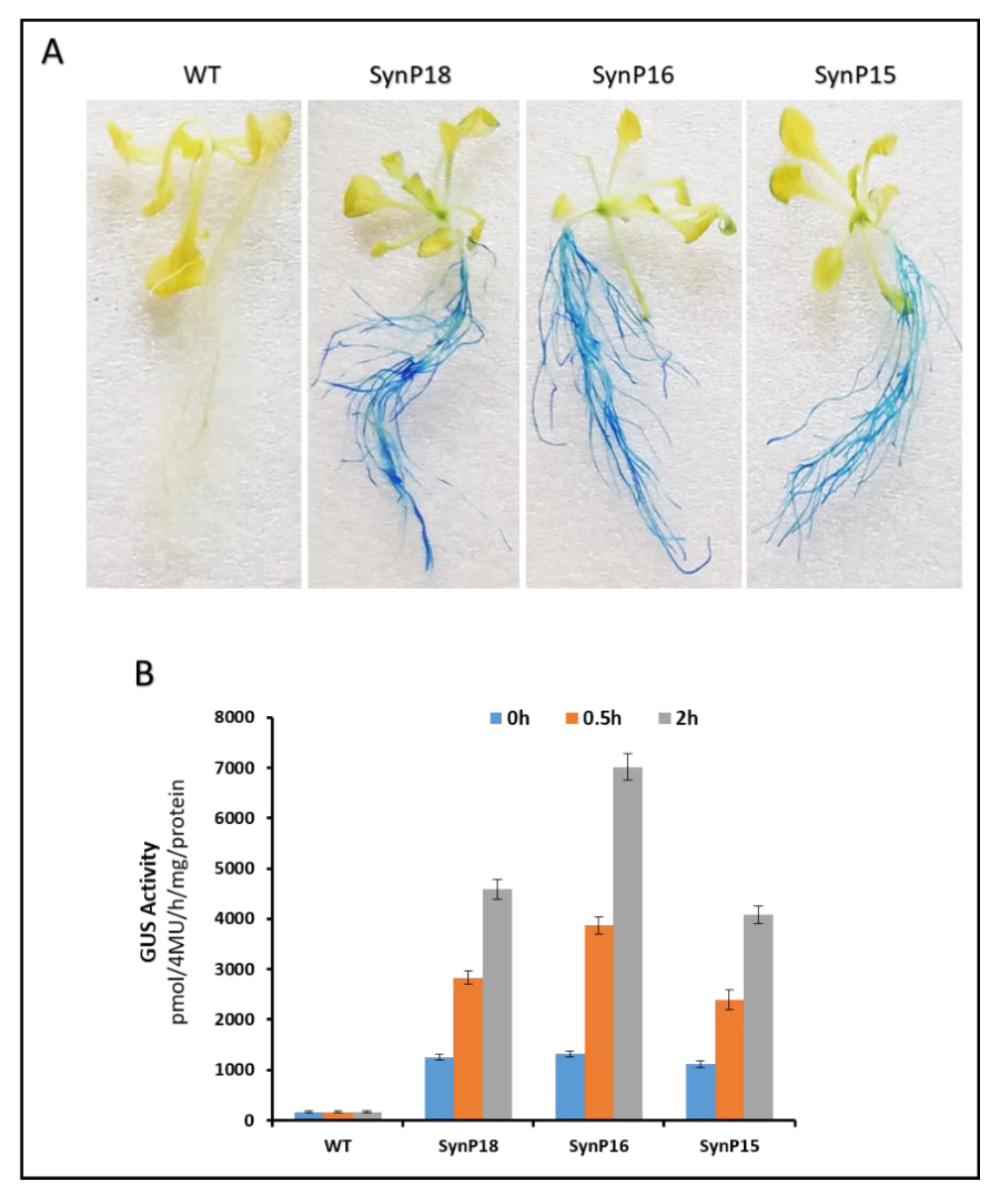
2.4. Functional Validation of Synthetic Promoters in Arabidopsis Thaliana
3. Discussion
4. Materials and Methods
4.1. Selection of Cis Motifs and Designing of Synthetic Modules
4.2. Agrobacterium Rhizogenes-Mediated Transformation of Soybean Hairy Roots
4.3. PEG Stress Treatment of Chimeric Soybean Plants
4.4. GUS Histochemical Staining and Fluorometric Analysis of Transgenic Soybean
4.5. Agrobacterium Tumefacians-Mediated Transformation of Arabidopsis
4.6. Agrobacterium Tumefaciens Strain EHA105 was Used for Stable Transformation in Arabidopsis Thaliana
4.7. Analysis of Transgenic Arabidopsis
4.8. Gus Assays of Drought-Treated Transgenic Arabidopsis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| ABRE | Abscisic Acid Responsive Element |
| ABF | ABRE-Binding Factor |
| BEST | Binding Site Estimation Suite of Tools |
| CaMV | Cauliflower Mosaic Virus |
| CBF | C-repeat (CRT) Binding Factor |
| HDGS | Homology-Dependent Gene Silencing |
| MEME | Multiple EM for Motif Elicitation |
| PEG | Polyethylene Glycol |
| PLACE | Plant Cis-Acting Regulatory Elements |
| SynP | Synthetic Promoter |
| TF | Transcription Factor |
References
- Bailey-Serres, J.; Parker, J.E.; Ainsworth, E.A.; Oldroyd, G.E.D.; Schroeder, J.I. Genetic strategies for improving crop yields. Nature 2019, 575, 109–118. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, M.; Cao, L.; Mwimba, M.; Zhou, Y.; Li, L.; Zhou, M.; Schnable, P.S.; O’Rourke, J.A.; Dong, X.; Wang, W. Comprehensive mapping of abiotic stress inputs into the soybean circadian clock. Proc. Natl. Acad. Sci. USA 2019, 116, 23840–23849. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bechtold, U.; Field, B. Molecular mechanisms controlling plant growth during abiotic stress. J. Exp. Bot. 2018, 69, 2753–2758. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xie, Z.; Nolan, T.; Jiang, H.; Tang, B.; Zhang, M.; Li, Z.; Yin, Y. The AP2/ERF Transcription Factor TINY Modulates Brassinosteroid-Regulated Plant Growth and Drought Responses in Arabidopsis. Plant Cell 2019, 31, 1788–1806. [Google Scholar] [CrossRef] [Green Version]
- Sauter, A.; Davies, W.J.; Hartung, W. The long-distance abscisic acid signal in the droughted plant: The fate of the hormone on its way from root to shoot. J. Exp. Bot. 2001, 52, 1991–1997. [Google Scholar] [CrossRef] [Green Version]
- Campalans, A.; Messeguer, R.; Goday, A.; Pagès, M. Plant responses to drought, from ABA signal transduction events to the action of the induced proteins. Plant Physiol. Biochem. 1999, 37, 327–340. [Google Scholar] [CrossRef]
- Boursiac, Y.; Léran, S.; Corratgé-Faillie, C.; Gojon, A.; Krouk, G.; Lacombe, B. ABA transport and transporters. Trends Plant Sci. 2013, 18, 325–333. [Google Scholar] [CrossRef]
- Wilkinson, S.; Davies, W.J. ABA-based chemical signalling: The co-ordination of responses to stress in plants. Plant. Cell Environ. 2002, 25, 195–210. [Google Scholar] [CrossRef]
- Fujita, Y.; Fujita, M.; Shinozaki, K.; Yamaguchi-Shinozaki, K. ABA-mediated transcriptional regulation in response to osmotic stress in plants. J. Plant Res. 2011, 124, 509–525. [Google Scholar] [CrossRef]
- Ranieri, A.; Bernardi, R.; Lanese, P.; Soldatini, G.F. Changes in free amino acid content and protein pattern of maize seedlings under water stress. Environ. Exp. Bot. 1989, 29, 351–357. [Google Scholar] [CrossRef]
- Bianchi, G.; Gamba, A.; Limiroli, R.; Pozzi, N.; Elster, R.; Salamini, F.; Bartels, D. The unusual sugar composition in leaves of the resurrection plant Myrothamnus flabellifolia. Physiol. Plant. 1993, 87, 223–226. [Google Scholar] [CrossRef]
- Wang, S.; Wan, C.; Wang, Y.; Chen, H.; Zhou, Z.; Fu, H.; Sosebee, R.E. The characteristics of Na+, K+ and free proline distribution in several drought-resistant plants of the Alxa Desert, China. J. Arid Environ. 2004, 56, 525–539. [Google Scholar] [CrossRef]
- Thomas, H.; James, A.R. Freezing Tolerance and Solute Changes in Contrasting Genotypes of Lolium perenne L. Acclimated to Cold and Drought. Ann. Bot. 1993, 72, 249–254. [Google Scholar] [CrossRef]
- Fang, Y.; Xiong, L. General mechanisms of drought response and their application in drought resistance improvement in plants. Cell. Mol. Life Sci. 2015, 72, 673–689. [Google Scholar] [CrossRef]
- Cruz de Carvalho, M.H. Drought stress and reactive oxygen species: Production, scavenging and signaling. Plant Signal. Behav. 2008, 3, 156–165. [Google Scholar] [CrossRef] [Green Version]
- Li, Z.; Zhang, Y.; Peng, D.; Wang, X.; Peng, Y.; He, X.; Zhang, X.; Ma, X.; Huang, L.; Yan, Y. Polyamine regulates tolerance to water stress in leaves of white clover associated with antioxidant defense and dehydrin genes via involvement in calcium messenger system and hydrogen peroxide signaling. Front. Physiol. 2015, 6, 280. [Google Scholar] [CrossRef] [Green Version]
- Rushton, P.J.; Reinstädler, A.; Lipka, V.; Lippok, B.; Somssich, I.E. Synthetic Plant Promoters Containing Defined Regulatory Elements Provide Novel Insights into Pathogen- and Wound-Induced Signaling. Plant Cell 2002, 14, 749–762. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.Z.; Creelman, R.A.; Zhu, J.-K. From laboratory to field. Using information from Arabidopsis to engineer salt, cold, and drought tolerance in crops. Plant Physiol. 2004, 135, 615–621. [Google Scholar] [CrossRef] [Green Version]
- Liu, W.; Stewart, C.N. Plant synthetic promoters and transcription factors. Curr. Opin. Biotechnol. 2016, 37, 36–44. [Google Scholar] [CrossRef] [Green Version]
- Aysha, J.; Noman, M.; Wang, F.; Liu, W.; Zhou, Y.; Li, H.; Li, X. Synthetic Promoters: Designing the cis Regulatory Modules for Controlled Gene Expression. Mol. Biotechnol. 2018, 60, 608–620. [Google Scholar] [CrossRef]
- Ali, S.; Kim, W.-C. A Fruitful Decade Using Synthetic Promoters in the Improvement of Transgenic Plants. Front. Plant Sci. 2019, 10, 1433. [Google Scholar] [CrossRef] [PubMed]
- Hernandez-Garcia, C.M.; Finer, J.J. Identification and validation of promoters and cis-acting regulatory elements. Plant Sci. 2014, 217–218, 109–119. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Potenza, C.; Aleman, L.; Sengupta-Gopalan, C. Targeting transgene expression in research, agricultural, and environmental applications: Promoters used in plant transformation. Vitr. Cell. Dev. Biol. Plant 2004, 40, 1–22. [Google Scholar] [CrossRef]
- Kumar, S.; AlAbed, D.; Whitteck, J.T.; Chen, W.; Bennett, S.; Asberry, A.; Wang, X.; DeSloover, D.; Rangasamy, M.; Wright, T.R.; et al. A combinatorial bidirectional and bicistronic approach for coordinated multi-gene expression in corn. Plant Mol. Biol. 2015, 87, 341–353. [Google Scholar] [CrossRef]
- Bhullar, S.; Datta, S.; Advani, S.; Chakravarthy, S.; Gautam, T.; Pental, D.; Burma, P.K. Functional analysis of cauliflower mosaic virus 35S promoter: Re-evaluation of the role of subdomains B5, B4 and B2 in promoter activity. Plant Biotechnol. J. 2007, 5, 696–708. [Google Scholar] [CrossRef]
- Cazzonelli, C.I.; Velten, J. In vivo characterization of plant promoter element interaction using synthetic promoters. Transgenic Res. 2008, 17, 437–457. [Google Scholar] [CrossRef]
- Mogno, I.; Vallania, F.; Mitra, R.D.; Cohen, B.A. TATA is a modular component of synthetic promoters. Genome Res. 2010, 20, 1391–1397. [Google Scholar] [CrossRef] [Green Version]
- Patro, S.; Kumar, D.; Ranjan, R.; Maiti, I.B.; Dey, N. The development of efficient plant promoters for transgene expression employing plant virus promoters. Mol. Plant 2012, 5, 941–944. [Google Scholar] [CrossRef] [Green Version]
- Acharya, S.; Ranjan, R.; Pattanaik, S.; Maiti, I.B.; Dey, N. Efficient chimeric plant promoters derived from plant infecting viral promoter sequences. Planta 2014, 239, 381–396. [Google Scholar] [CrossRef]
- Bhullar, S.; Chakravarthy, S.; Advani, S.; Datta, S.; Pental, D.; Burma, P.K. Strategies for development of functionally equivalent promoters with minimum sequence homology for transgene expression in plants: Cis-elements in a novel DNA context versus domain swapping. Plant Physiol. 2003, 132, 988–998. [Google Scholar] [CrossRef] [Green Version]
- Moshelion, M.; Altman, A. Current challenges and future perspectives of plant and agricultural biotechnology. Trends Biotechnol. 2015, 33, 337–342. [Google Scholar] [CrossRef] [PubMed]
- Dey, N.; Sarkar, S.; Acharya, S.; Maiti, I.B. Synthetic promoters in planta. Planta 2015, 242, 1077–1094. [Google Scholar] [CrossRef] [PubMed]
- Venter, M. Synthetic promoters: Genetic control through cis engineering. Trends Plant Sci. 2007, 12, 118–124. [Google Scholar] [CrossRef] [PubMed]
- Deb, D.; Shrestha, A.; Maiti, I.B.; Dey, N. Recombinant Promoter (MUASCsV8CP) Driven Totiviral Killer Protein 4 (KP4) Imparts Resistance Against Fungal Pathogens in Transgenic Tobacco. Front. Plant Sci. 2018, 9, 278. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, J.-S.; Mizoi, J.; Yoshida, T.; Fujita, Y.; Nakajima, J.; Ohori, T.; Todaka, D.; Nakashima, K.; Hirayama, T.; Shinozaki, K.; et al. An ABRE promoter sequence is involved in osmotic stress-responsive expression of the DREB2A gene, which encodes a transcription factor regulating drought-inducible genes in Arabidopsis. Plant Cell Physiol. 2011, 52, 2136–2146. [Google Scholar] [CrossRef]
- Zheng, H.; Lei, Y.; Lin, S.; Zhang, Q.; Zhang, Z. Bidirectionalization of a methyl jasmonate-inducible plant promoter. Biotechnol. Lett. 2011, 33, 387–393. [Google Scholar] [CrossRef] [PubMed]
- Shokouhifar, F.; Zamani, M.R.; Motallebi, M.; Mousavi, A.; Malboobi, M.A. Construction and functional analysis of pathogen-inducible synthetic promoters in Brassica napus. Biol. Plant. 2011, 55, 689. [Google Scholar] [CrossRef]
- Porto, M.S.; Pinheiro, M.P.N.; Batista, V.G.L.; dos Santos, R.C.; Filho, P.d.A.M.; de Lima, L.M. Plant promoters: An approach of structure and function. Mol. Biotechnol. 2014, 56, 38–49. [Google Scholar] [CrossRef] [Green Version]
- Kumari, S.; Ware, D. Genome-wide computational prediction and analysis of core promoter elements across plant monocots and dicots. PLoS ONE 2013, 8, e79011. [Google Scholar] [CrossRef] [Green Version]
- Sawant, S.; Singh, P.K.; Madanala, R.; Tuli, R. Designing of an artificial expression cassette for the high-level expression of transgenes in plants. Theor. Appl. Genet. 2001, 102, 635–644. [Google Scholar] [CrossRef]
- Rodrigues, F.A.; Fuganti-Pagliarini, R.; Marcolino-Gomes, J.; Nakayama, T.J.; Molinari, H.B.C.; Lobo, F.P.; Harmon, F.G.; Nepomuceno, A.L. Daytime soybean transcriptome fluctuations during water deficit stress. BMC Genomics 2015, 16, 505. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Le, D.T.; Nishiyama, R.; Watanabe, Y.; Tanaka, M.; Seki, M.; Ham, L.H.; Yamaguchi-Shinozaki, K.; Shinozaki, K.; Tran, L.-S.P. Differential gene expression in soybean leaf tissues at late developmental stages under drought stress revealed by genome-wide transcriptome analysis. PLoS ONE 2012, 7, e49522. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jensen, S.T.; Liu, J.S. BioOptimizer: A Bayesian scoring function approach to motif discovery. Bioinformatics 2004, 20, 1557–1564. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Che, D.; Jensen, S.; Cai, L.; Liu, J.S. BEST: Binding-site Estimation Suite of Tools. Bioinformatics 2005, 21, 2909–2911. [Google Scholar] [CrossRef] [Green Version]
- Mahony, S.; Benos, P. V STAMP: A web tool for exploring DNA-binding motif similarities. Nucleic Acids Res. 2007, 35, W253–W258. [Google Scholar] [CrossRef] [Green Version]
- Yilmaz, A.; Mejia-Guerra, M.K.; Kurz, K.; Liang, X.; Welch, L.; Grotewold, E. AGRIS: The Arabidopsis Gene Regulatory Information Server, an update. Nucleic Acids Res. 2011, 39, D1118–D1122. [Google Scholar] [CrossRef] [Green Version]
- Bülow, L.; Brill, Y.; Hehl, R. AthaMap-assisted transcription factor target gene identification in Arabidopsis thaliana. Database (Oxford). 2010, 2010, baq034. [Google Scholar] [CrossRef]
- Palaniswamy, S.K.; James, S.; Sun, H.; Lamb, R.S.; Davuluri, R.V.; Grotewold, E. AGRIS and AtRegNet. a platform to link cis-regulatory elements and transcription factors into regulatory networks. Plant Physiol. 2006, 140, 818–829. [Google Scholar] [CrossRef] [Green Version]
- Steffens, N.O.; Galuschka, C.; Schindler, M.; Bülow, L.; Hehl, R. AthaMap: An online resource for in silico transcription factor binding sites in the Arabidopsis thaliana genome. Nucleic Acids Res. 2004, 32, D368–D372. [Google Scholar] [CrossRef] [Green Version]
- Davuluri, R.V.; Sun, H.; Palaniswamy, S.K.; Matthews, N.; Molina, C.; Kurtz, M.; Grotewold, E. AGRIS: Arabidopsis gene regulatory information server, an information resource of Arabidopsis cis-regulatory elements and transcription factors. BMC Bioinform. 2003, 4, 25. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Higo, K.; Ugawa, Y.; Iwamoto, M.; Korenaga, T. Plant cis-acting regulatory DNA elements (PLACE) database: 1999. Nucleic Acids Res. 1999, 27, 297–300. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, L.; Song, Z.; Li, F.; Li, X.; Ji, H.; Yang, S. The specific MYB binding sites bound by TaMYB in the GAPCp2/3 promoters are involved in the drought stress response in wheat. BMC Plant Biol. 2019, 19, 366. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sheshadri, S.A.; Nishanth, M.J.; Simon, B. Stress-Mediated cis-Element Transcription Factor Interactions Interconnecting Primary and Specialized Metabolism in planta. Front. Plant Sci. 2016, 7, 1725. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tripathi, P.; Rabara, R.C.; Reese, R.N.; Miller, M.A.; Rohila, J.S.; Subramanian, S.; Shen, Q.J.; Morandi, D.; Bücking, H.; Shulaev, V.; et al. A toolbox of genes, proteins, metabolites and promoters for improving drought tolerance in soybean includes the metabolite coumestrol and stomatal development genes. BMC Genomics 2016, 17, 102. [Google Scholar] [CrossRef] [Green Version]
- Kim, H.-J.; Kim, Y.-K.; Park, J.-Y.; Kim, J. Light signalling mediated by phytochrome plays an important role in cold-induced gene expression through the C-repeat/dehydration responsive element (C/DRE) in Arabidopsis thaliana. Plant J. 2002, 29, 693–704. [Google Scholar] [CrossRef]
- Wong, D.C.J.; Ariani, P.; Castellarin, S.; Polverari, A.; Vandelle, E. Co-expression network analysis and cis-regulatory element enrichment determine putative functions and regulatory mechanisms of grapevine ATL E3 ubiquitin ligases. Sci. Rep. 2018, 8, 3151. [Google Scholar] [CrossRef] [Green Version]
- Dixit, S.K.; Gupta, A.; Fatima, U.; Senthil-Kumar, M. AtGBF3 confers tolerance to Arabidopsis thaliana against combined drought and Pseudomonas syringae stress. Environ. Exp. Bot. 2019, 168, 103881. [Google Scholar] [CrossRef]
- Müller, M.; Munné-Bosch, S. Ethylene Response Factors: A Key Regulatory Hub in Hormone and Stress Signaling. Plant Physiol. 2015, 169, 32–41. [Google Scholar] [CrossRef] [Green Version]
- Baldoni, E.; Genga, A.; Cominelli, E. Plant MYB Transcription Factors: Their Role in Drought Response Mechanisms. Int. J. Mol. Sci. 2015, 16, 15811–15851. [Google Scholar] [CrossRef] [Green Version]
- Pasquali, G.; Biricolti, S.; Locatelli, F.; Baldoni, E.; Mattana, M. Osmyb4 expression improves adaptive responses to drought and cold stress in transgenic apples. Plant Cell Rep. 2008, 27, 1677–1686. [Google Scholar] [CrossRef]
- Fu, M.; Kang, H.K.; Son, S.-H.; Kim, S.-K.; Nam, K.H. A Subset of Arabidopsis RAV Transcription Factors Modulates Drought and Salt Stress Responses Independent of ABA. Plant Cell Physiol. 2014, 55, 1892–1904. [Google Scholar] [CrossRef] [Green Version]
- Kereszt, A.; Li, D.; Indrasumunar, A.; Nguyen, C.D.T.; Nontachaiyapoom, S.; Kinkema, M.; Gresshoff, P.M. Agrobacterium rhizogenes-mediated transformation of soybean to study root biology. Nat. Protoc. 2007, 2, 948–952. [Google Scholar] [CrossRef]
- Hernandez-Garcia, C.M.; Bouchard, R.A.; Rushton, P.J.; Jones, M.L.; Chen, X.; Timko, M.P.; Finer, J.J. High level transgenic expression of soybean (Glycine max) GmERF and Gmubi gene promoters isolated by a novel promoter analysis pipeline. BMC Plant Biol. 2010, 10, 237. [Google Scholar] [CrossRef] [Green Version]
- Cho, H.-J.; Farrand, S.K.; Noel, G.R.; Widholm, J.M. High-efficiency induction of soybean hairy roots and propagation of the soybean cyst nematode. Planta 2000, 210, 195–204. [Google Scholar] [CrossRef]
- Giri, A.; Narasu, M.L. Transgenic hairy roots: Recent trends and applications. Biotechnol. Adv. 2000, 18, 1–22. [Google Scholar] [CrossRef]
- Ma, J.K.-C.; Drake, P.M.W.; Christou, P. The production of recombinant pharmaceutical proteins in plants. Nat. Rev. Genet. 2003, 4, 794–805. [Google Scholar] [CrossRef]
- Farre, G.; Ramessar, K.; Twyman, R.M.; Capell, T.; Christou, P. The humanitarian impact of plant biotechnology: Recent breakthroughs vs bottlenecks for adoption. Curr. Opin. Plant Biol. 2010, 13, 219–225. [Google Scholar] [CrossRef]
- Zhu, C.; Naqvi, S.; Gomez-Galera, S.; Pelacho, A.M.; Capell, T.; Christou, P. Transgenic strategies for the nutritional enhancement of plants. Trends Plant Sci. 2007, 12, 548–555. [Google Scholar] [CrossRef]
- Bower, R.; Birch, R.G. Transgenic sugarcane plants via microprojectile bombardment. Plant J. 1992, 2, 409–416. [Google Scholar] [CrossRef]
- Arencibia, A.D.; Carmona, E.R.; Tellez, P.; Chan, M.-T.; Yu, S.-M.; Trujillo, L.E.; Oramas, P. An efficient protocol for sugarcane (Saccharum spp. L.) transformation mediated by Agrobacterium tumefaciens. Transgenic Res. 1998, 7, 213–222. [Google Scholar] [CrossRef]
- Christy, L.A.; Arvinth, S.; Saravanakumar, M.; Kanchana, M.; Mukunthan, N.; Srikanth, J.; Thomas, G.; Subramonian, N. Engineering sugarcane cultivars with bovine pancreatic trypsin inhibitor (aprotinin) gene for protection against top borer (Scirpophaga excerptalis Walker). Plant Cell Rep. 2009, 28, 175–184. [Google Scholar] [CrossRef] [PubMed]
- Manickavasagam, M.; Ganapathi, A.; Anbazhagan, V.R.; Sudhakar, B.; Selvaraj, N.; Vasudevan, A.; Kasthurirengan, S. Agrobacterium-mediated genetic transformation and development of herbicide-resistant sugarcane (Saccharum species hybrids) using axillary buds. Plant Cell Rep. 2004, 23, 134–143. [Google Scholar] [CrossRef] [PubMed]
- Arvinth, S.; Arun, S.; Selvakesavan, R.K.; Srikanth, J.; Mukunthan, N.; Ananda Kumar, P.; Premachandran, M.N.; Subramonian, N. Genetic transformation and pyramiding of aprotinin-expressing sugarcane with cry1Ab for shoot borer (Chilo infuscatellus) resistance. Plant Cell Rep. 2010, 29, 383–395. [Google Scholar] [CrossRef] [PubMed]
- Gurr, S.J.; Rushton, P.J. Engineering plants with increased disease resistance: What are we going to express? Trends Biotechnol. 2005, 23, 275–282. [Google Scholar] [CrossRef] [PubMed]
- Ito, M.; Iwase, M.; Kodama, H.; Lavisse, P.; Komamine, A.; Nishihama, R.; Machida, Y.; Watanabe, A. A novel cis-acting element in promoters of plant B-type cyclin genes activates M phase-specific transcription. Plant Cell 1998, 10, 331–341. [Google Scholar]
- Slusarczyk, A.L.; Lin, A.; Weiss, R. Foundations for the design and implementation of synthetic genetic circuits. Nat. Rev. Genet. 2012, 13, 406–420. [Google Scholar] [CrossRef]
- Maiti, I.B.; Gowda, S.; Kiernan, J.; Ghosh, S.K.; Shepherd, R.J. Promoter/leader deletion analysis and plant expression vectors with the figwort mosaic virus (FMV) full length transcript (FLt) promoter containing single or double enhancer domains. Transgenic Res. 1997, 6, 143–156. [Google Scholar] [CrossRef]
- Shen, Q.; Zhang, P.; Ho, T.H. Modular nature of abscisic acid (ABA) response complexes: Composite promoter units that are necessary and sufficient for ABA induction of gene expression in barley. Plant Cell 1996, 8, 1107–1119. [Google Scholar]
- Ishige, F.; Takaichi, M.; Foster, R.; Chua, N.-H.; Oeda, K. A G-box motif (GCCACGTGCC) tetramer confers high-level constitutive expression in dicot and monocot plants. Plant J. 1999, 18, 443–448. [Google Scholar] [CrossRef]
- Liu, W.; Yuan, J.S.; Stewart, C.N.J. Advanced genetic tools for plant biotechnology. Nat. Rev. Genet. 2013, 14, 781–793. [Google Scholar] [CrossRef]
- Wray, G.A. Promoter Logic. Science (80-.). 1998, 279, 1871–1872. [Google Scholar] [CrossRef] [PubMed]
- Zhang, N.; McHale, L.K.; Finer, J.J. A Leader Intron of a Soybean Elongation Factor 1A (eEF1A) Gene Interacts with Proximal Promoter Elements to Regulate Gene Expression in Synthetic Promoters. PLoS ONE 2016, 11, e0166074. [Google Scholar] [CrossRef] [PubMed]
- Lescot, M.; Déhais, P.; Thijs, G.; Marchal, K.; Moreau, Y.; Van de Peer, Y.; Rouzé, P.; Rombauts, S. PlantCARE, a database of plant cis-acting regulatory elements and a portal to tools for in silico analysis of promoter sequences. Nucleic Acids Res. 2002, 30, 325–327. [Google Scholar] [CrossRef] [PubMed]
- Shahmuradov, I.A.; Gammerman, A.J.; Hancock, J.M.; Bramley, P.M.; Solovyev, V. V PlantProm: A database of plant promoter sequences. Nucleic Acids Res. 2003, 31, 114–117. [Google Scholar] [CrossRef] [Green Version]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Jefferson, R.A.; Kavanagh, T.A.; Bevan, M.W. GUS fusions: Beta-glucuronidase as a sensitive and versatile gene fusion marker in higher plants. EMBO J. 1987, 6, 3901–3907. [Google Scholar] [CrossRef]
- Enhanced recovery of transformants of Agrobacterium tumefaciens after freeze-thaw transformation and drug selection. Biotechniques 1994, 16, 664–669.








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | Sequence |
|---|---|
| ABF [21] | CACGTGGC |
| ABRE [35] | TACGTGGC |
| ABRE-LIKE [54] | ACGTGTC |
| CBF [55] | TGGCCGAC |
| E2F-VARIANT [56] | TCTCCCGCC |
| G-BOX [57] | CACGTG |
| GCC-BOX [58] | GCCGCC |
| MYB1 [59] | TCCTACC |
| MYB4 [60] | ACCTACC |
| RAV1-A [61] | CAACA |
| RAV1-B [61] | CACCTG |
| RANDOM BASES | TCATAACGCT |
| ROOT MOTIF | ATATT |
| 35S MINI | TCTCCACTGACGTAAGGGATGACGCACAATCCCACTATCCTTCGCAAGACCCTTCCTCTATATAAGGAAGTTCATTTCATTTGGAGAGGA |
| INTRON | CCAGATCTTATATAAGTTTTTGGTTCAAGAAAGTTTTTGGTTACTGATGAATAGATCTATTAACTGTTACTTTAATCGATTCAAGCTAAAGTTTTTTGGTTACTGATGAATAGATCTATTATCTGTTACTTTTAATCGGTCAAGCTCAAGTTTTTTGGTTACTGATGAATAGATCTATATACGTCACAGTGTGCTAAACATGCCCTTGTTTTATCTCGATC |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jameel, A.; Noman, M.; Liu, W.; Ahmad, N.; Wang, F.; Li, X.; Li, H. Tinkering Cis Motifs Jigsaw Puzzle Led to Root-Specific Drought-Inducible Novel Synthetic Promoters. Int. J. Mol. Sci. 2020, 21, 1357. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21041357
Jameel A, Noman M, Liu W, Ahmad N, Wang F, Li X, Li H. Tinkering Cis Motifs Jigsaw Puzzle Led to Root-Specific Drought-Inducible Novel Synthetic Promoters. International Journal of Molecular Sciences. 2020; 21(4):1357. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21041357
Chicago/Turabian StyleJameel, Aysha, Muhammad Noman, Weican Liu, Naveed Ahmad, Fawei Wang, Xiaowei Li, and Haiyan Li. 2020. "Tinkering Cis Motifs Jigsaw Puzzle Led to Root-Specific Drought-Inducible Novel Synthetic Promoters" International Journal of Molecular Sciences 21, no. 4: 1357. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21041357