Altered Cerebral Blood Flow and Potential Neuroprotective Effect of Human Relaxin-2 (Serelaxin) During Hypoxia or Severe Hypovolemia in a Sheep Model

 , , ,
, , , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
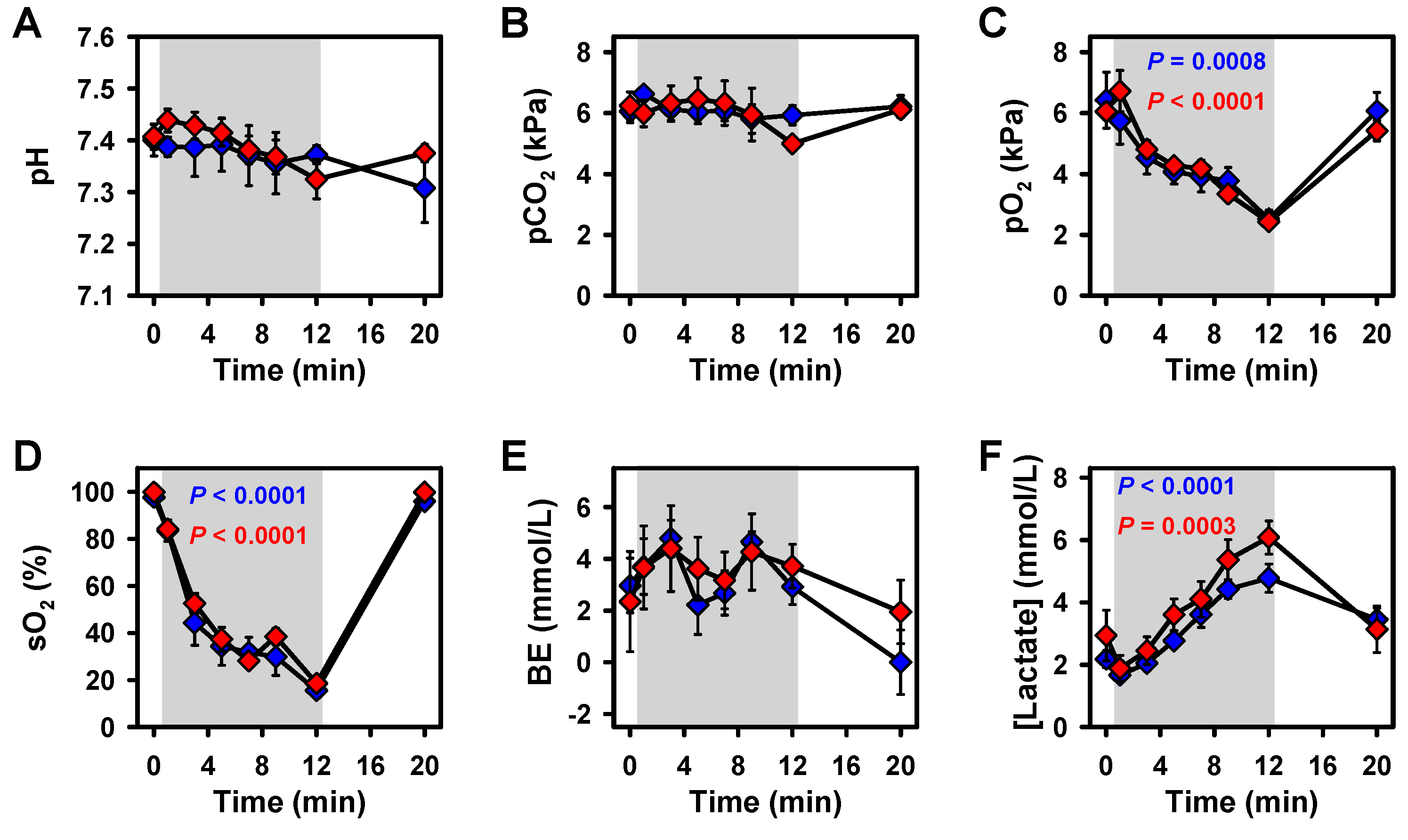
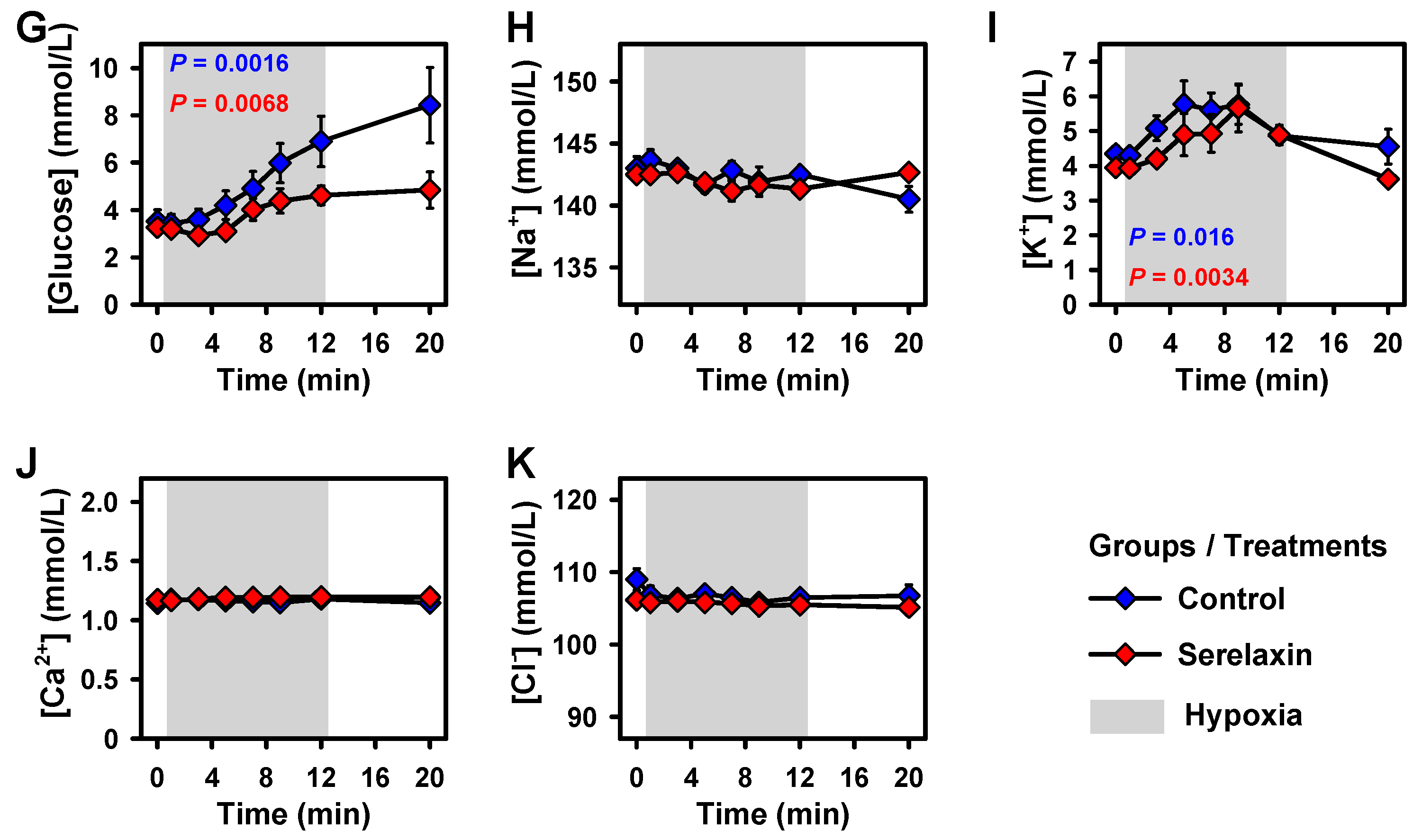
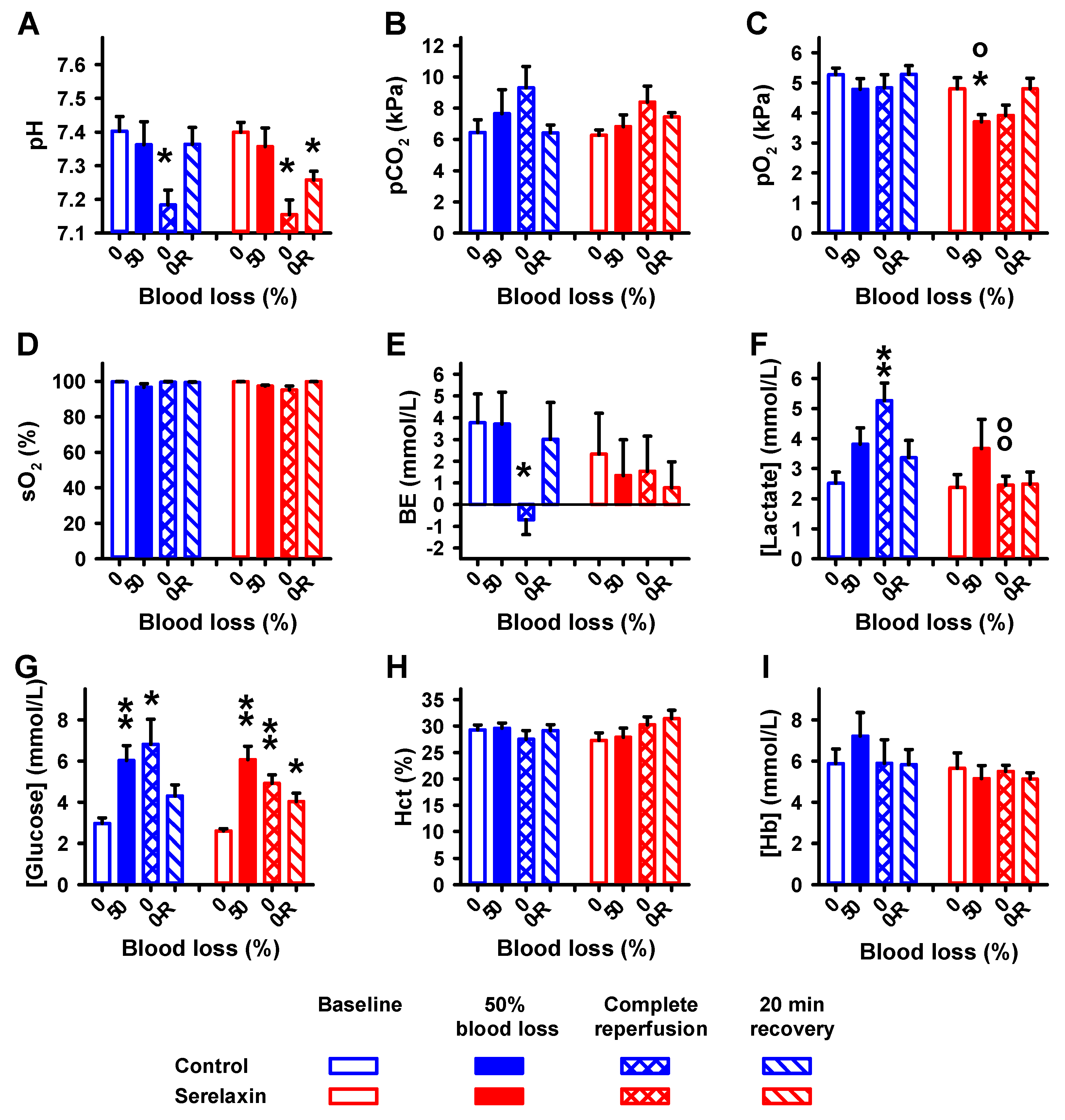
2.1. Blood Parameters
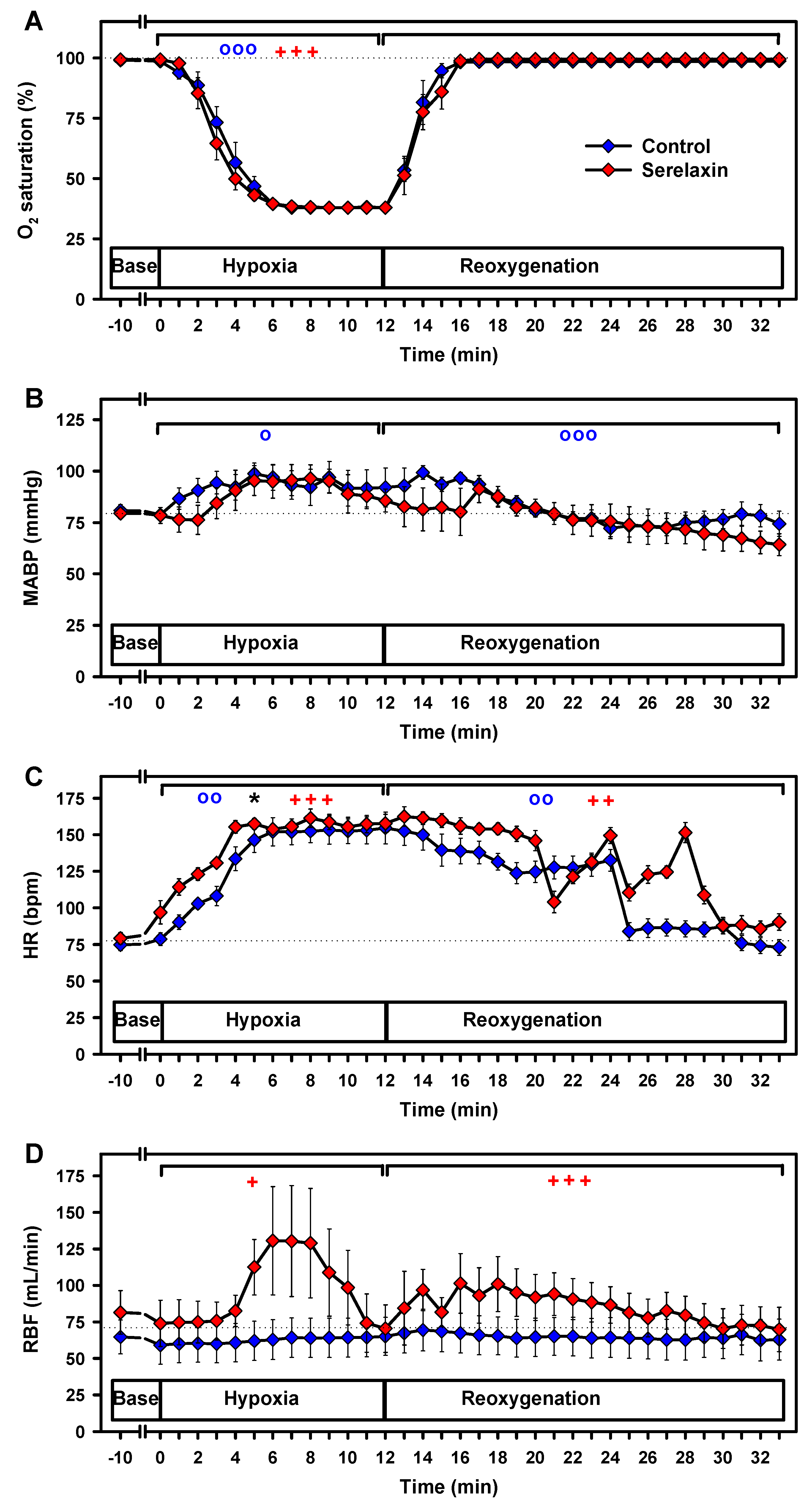
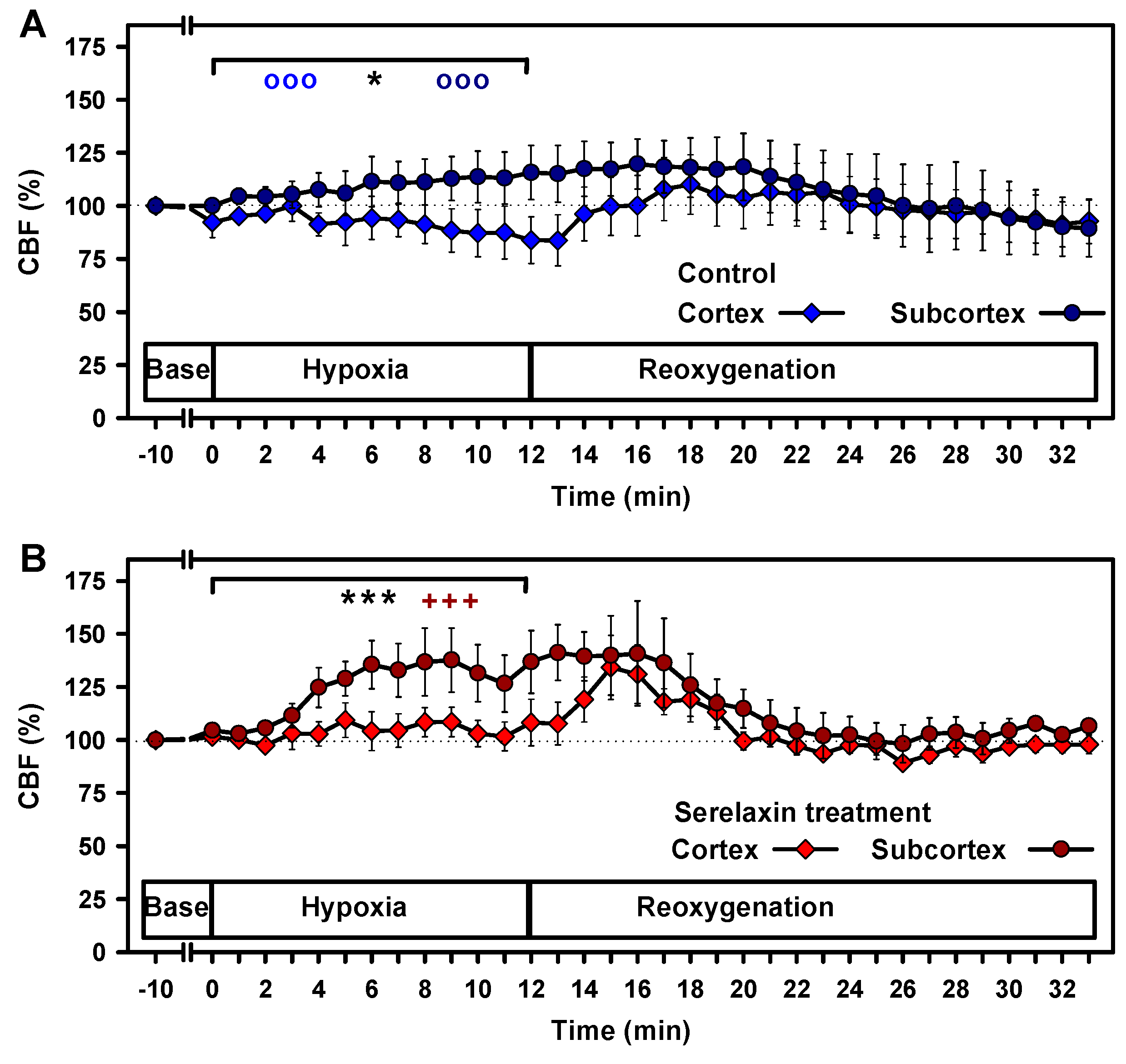
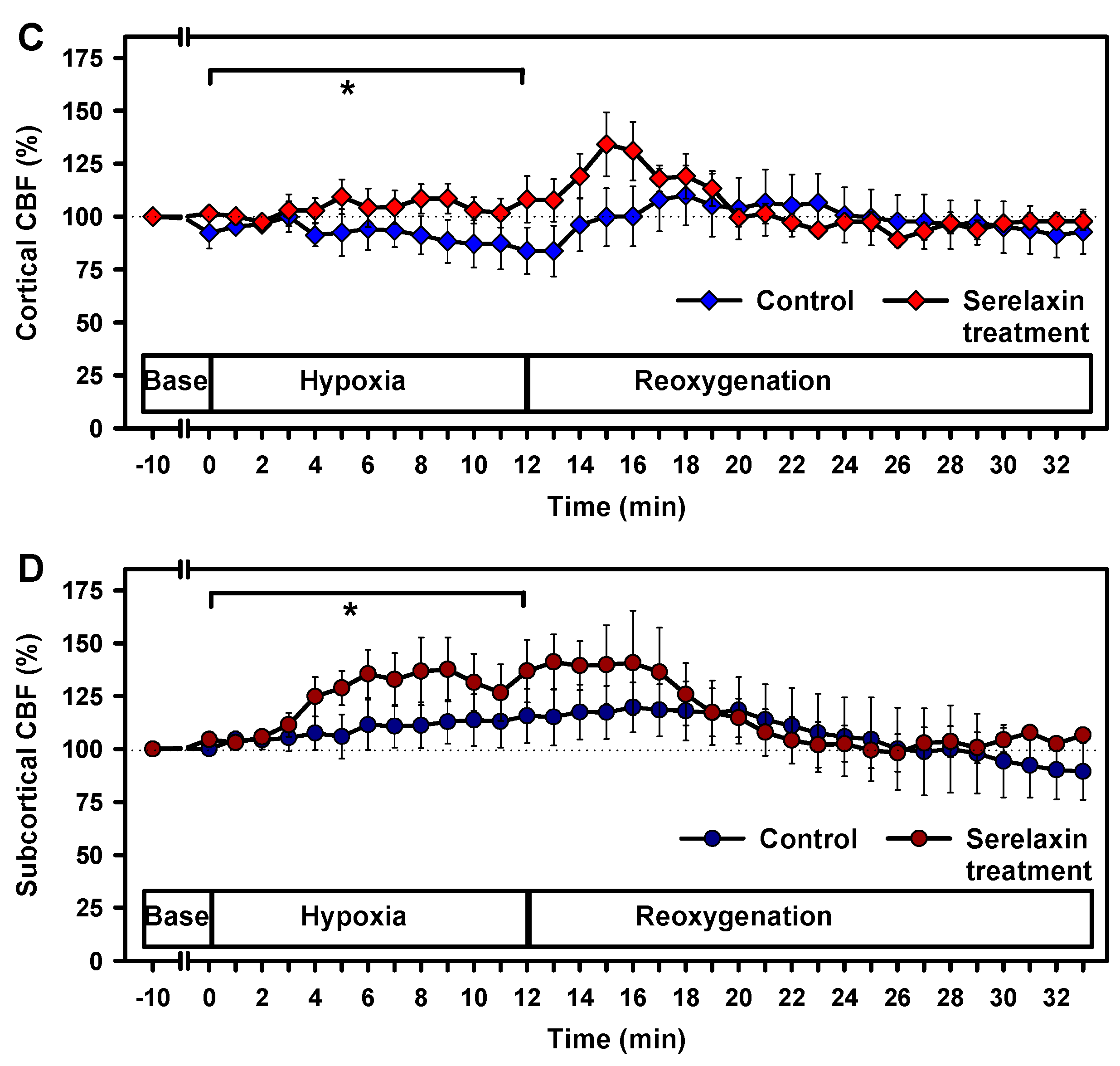
2.2. Effects of Hypoxia on Vital Parameters and CBF
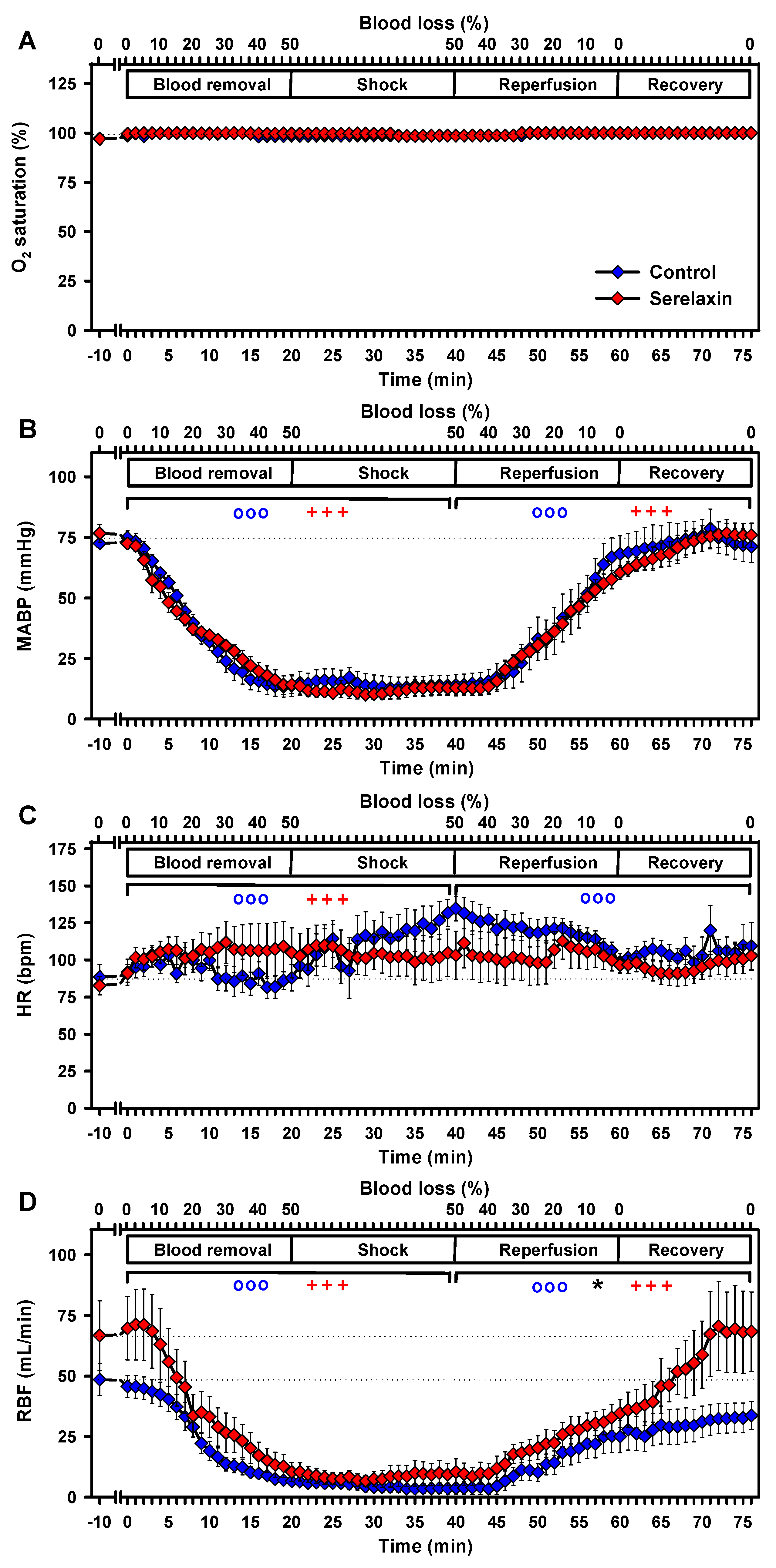
2.3. Effects of Hypovolemia on CBF and Vital Parameters
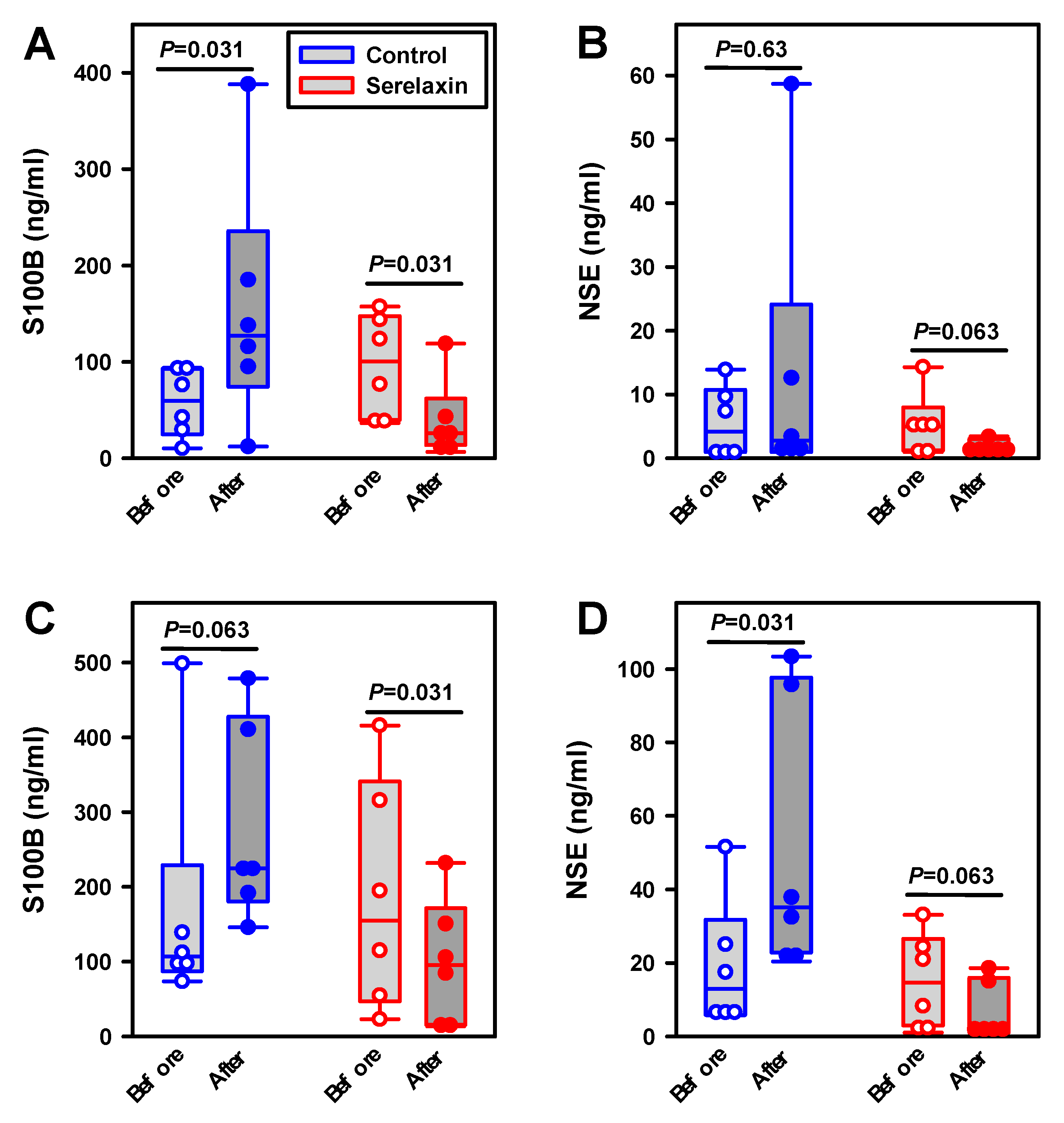
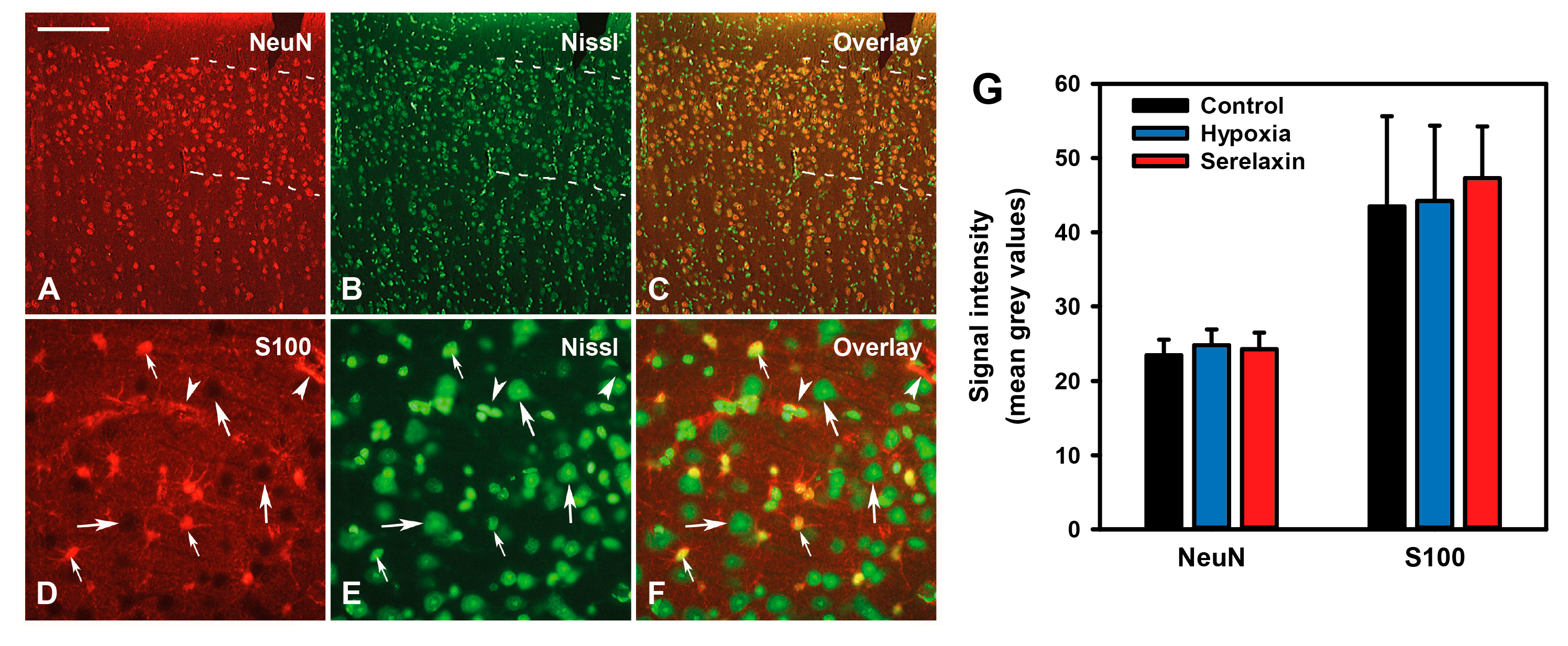
2.4. Effects of Hypoxia and Hypovolemia on Neuroproteins
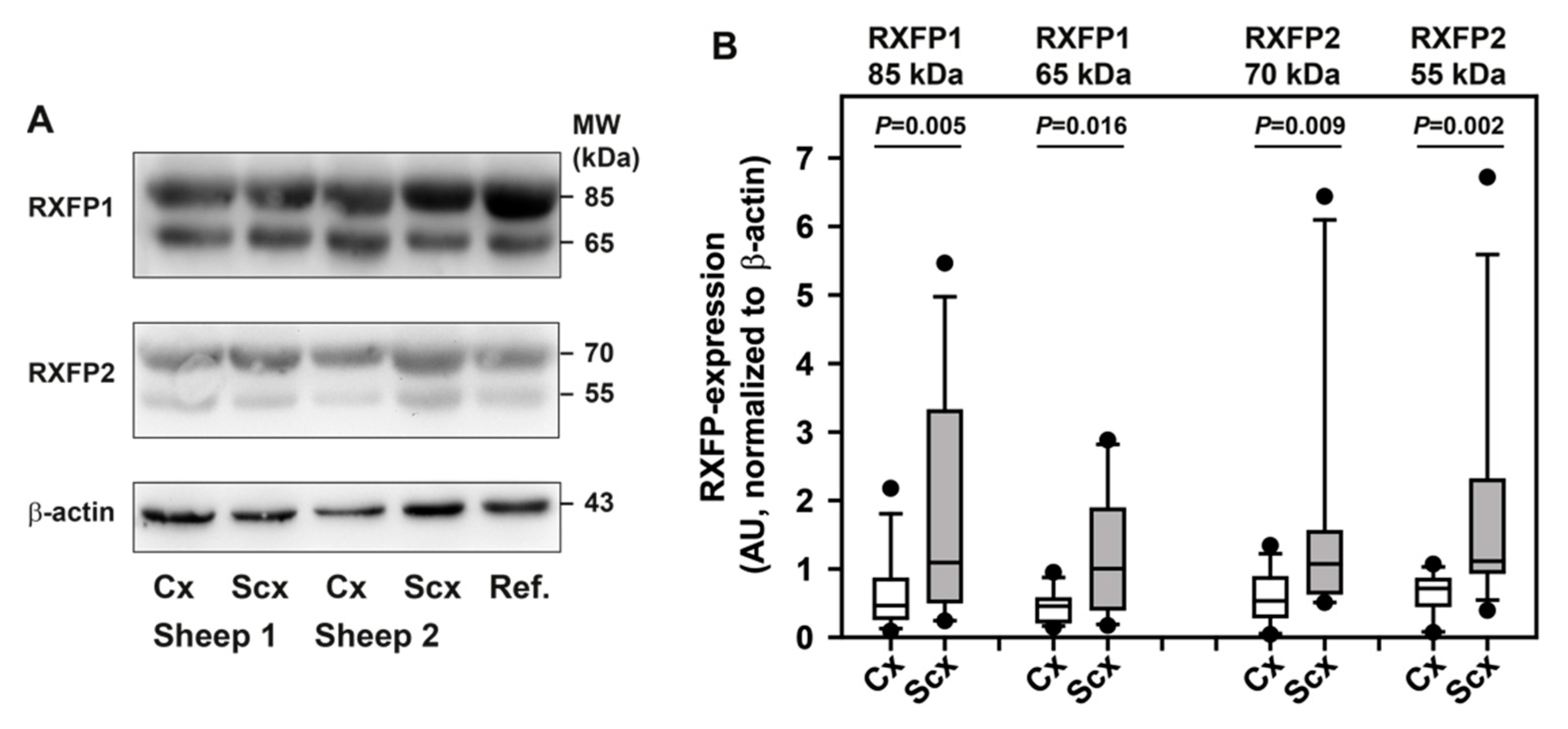
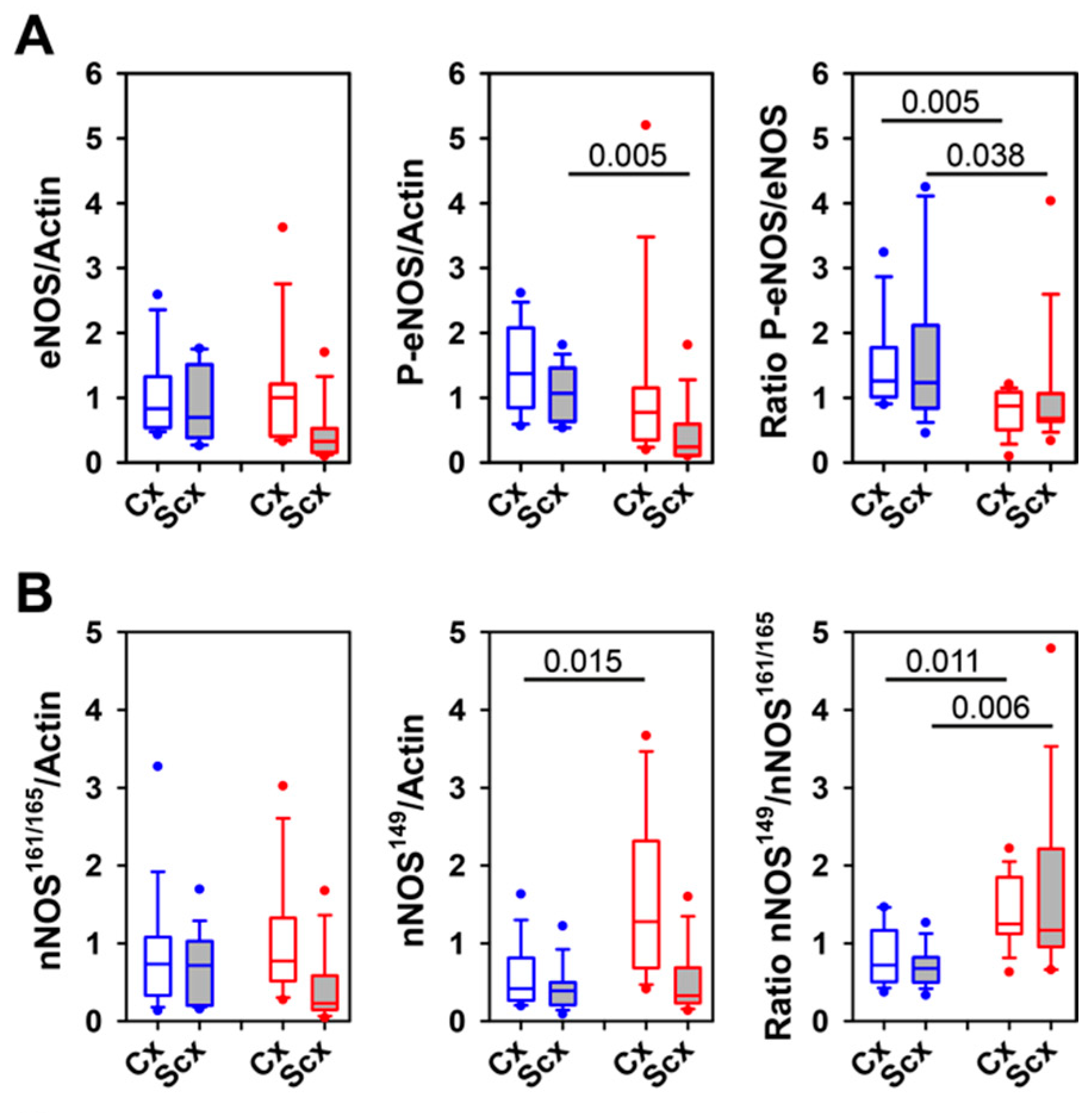
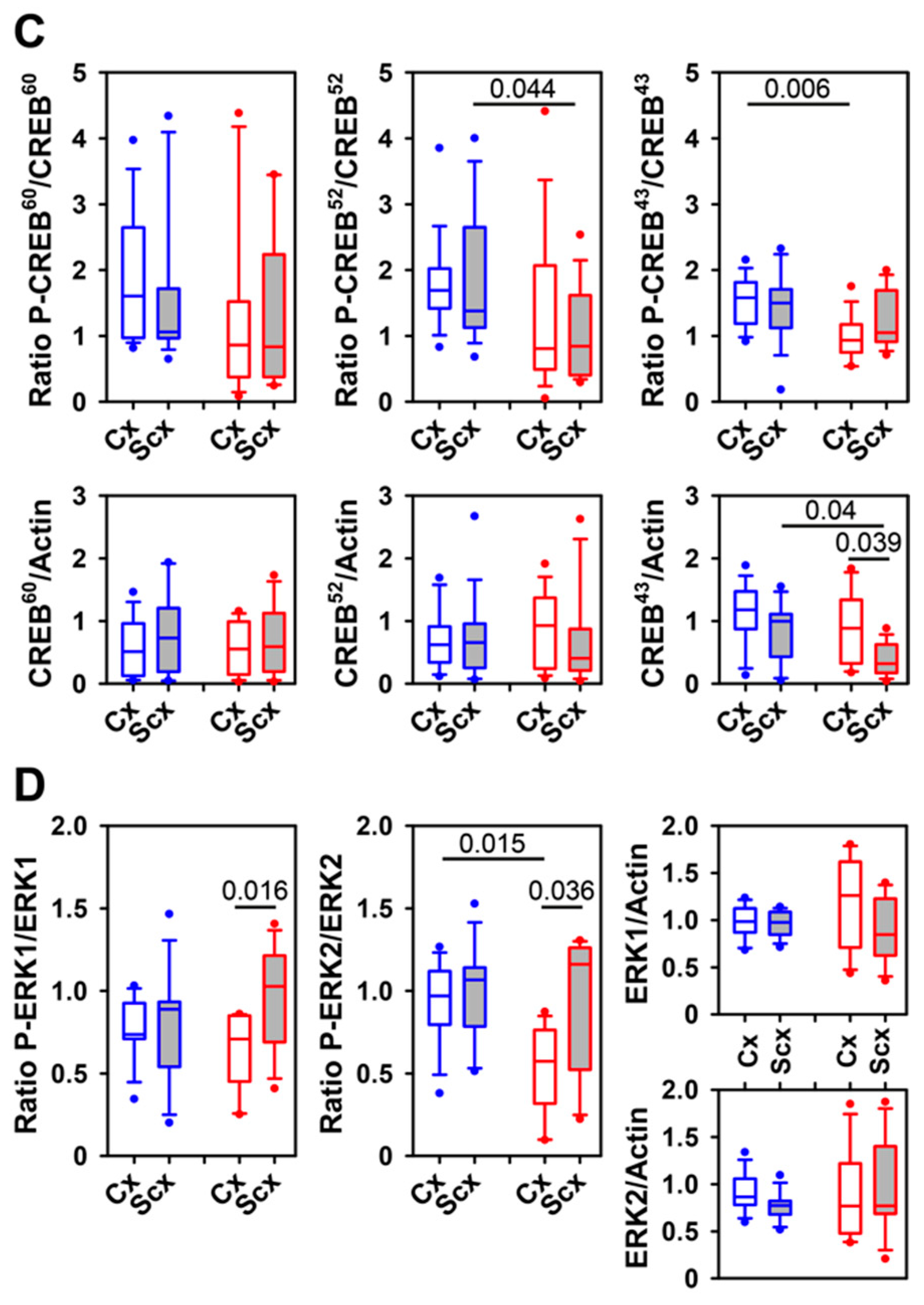
2.5. Expression of Relaxin Receptors and Potential Downstream Signaling Proteins
3. Discussion
4. Materials and Methods
4.1. Animal Care and Surgical Instrumentation
4.2. Cerebral Blood Flow and Microcirculation
4.3. Experimental Protocol
4.4. Induction and Resolution of Hypoxia
4.5. Induction and Resolution of Hypovolemia
4.6. Analysis of Blood Gases
4.7. Quantitation of S100B and NSE
4.8. Sample Preparation and Western Blotting
4.9. Immunofluorescence
4.10. Randomisation and Blinding
4.11. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| BE | base excess |
| CBF | cerebral blood flow |
| CREB | cAMP-responsive element-binding protein |
| cx | cortex |
| ECG | electrocardiogram |
| e/nNOS | endothelial/neuronal NO synthase |
| ERK1/2 | extracellular signal-regulated kinases 1/2 |
| HR | heart rate |
| MABP | mean arterial blood pressure |
| NSE | neuron-specific enolase |
| pCO2 | carbon dioxide partial pressure |
| pO2 | oxygen partial pressure |
| RBF | renal blood flow |
| RXFP1/2 | relaxin receptor 1/2 |
| scx | subcortex |
| sO2 | oxygen saturation |
| S100B | neuroprotein S100B |
References
- Conrad, K.P.; Shroff, S.G. Effects of relaxin on arterial dilation, remodeling, and mechanical properties. Curr. Hypertens. Rep. 2011, 13, 409–420. [Google Scholar] [CrossRef] [PubMed]
- Hernandez, A.V. Serelaxin: Insights into its haemodynamic, biochemical, and clinical effects in acute heart failure. Eur. Heart J. 2014, 35, 410–412. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ennezat, P.V.; Stewart, M.; Samson, R.; Bouabdallaoui, N.; Marechaux, S.; Banfi, C.; Bouvaist, H.; Le Jemtel, T.H. Recent therapeutic trials on fluid removal and vasodilation in acute heart failure. Eur. Heart J. Acute Cardiovasc. Care 2016, 5, 86–95. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sasser, J.M.; Cunningham, M.W., Jr.; Baylis, C. Serelaxin reduces oxidative stress and asymmetric dimethylarginine in angiotensin II-induced hypertension. Am. J. Physiol. Ren. Physiol. 2014, 307, F1355–F1362. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leo, C.H.; Jelinic, M.; Parkington, H.C.; Tare, M.; Parry, L.J. Acute intravenous injection of serelaxin (recombinant human relaxin-2) causes rapid and sustained bradykinin-mediated vasorelaxation. J. Am. Heart Assoc. 2014, 3, e000493. [Google Scholar] [CrossRef] [Green Version]
- Sarwar, M.; Samuel, C.S.; Bathgate, R.A.; Stewart, D.R.; Summers, R.J. Serelaxin-mediated signal transduction in human vascular cells: Bell-shaped concentration-response curves reflect differential coupling to G proteins. Br. J. Pharmacol. 2015, 172, 1005–1019. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Segal, M.S.; Sautina, L.; Li, S.; Diao, Y.; Agoulnik, A.I.; Kielczewski, J.; McGuane, J.T.; Grant, M.B.; Conrad, K.P. Relaxin increases human endothelial progenitor cell NO and migration and vasculogenesis in mice. Blood 2012, 119, 629–636. [Google Scholar] [CrossRef] [Green Version]
- Bergeron, L.H.; Willcox, J.M.; Alibhai, F.J.; Connell, B.J.; Saleh, T.M.; Wilson, B.C.; Summerlee, A.J. Relaxin peptide hormones are protective during the early stages of ischemic stroke in male rats. Endocrinology 2015, 156, 638–646. [Google Scholar] [CrossRef] [Green Version]
- Wilson, B.C.; Connell, B.; Saleh, T.M. Relaxin-induced reduction of infarct size in male rats receiving MCAO is dependent on nitric oxide synthesis and not estrogenic mechanisms. Neurosci. Lett. 2006, 393, 160–164. [Google Scholar] [CrossRef]
- Wilson, B.C.; Milne, P.; Saleh, T.M. Relaxin pretreatment decreases infarct size in male rats after middle cerebral artery occlusion. Ann. NY Acad. Sci. 2005, 1041, 223–228. [Google Scholar] [CrossRef]
- Bischoff, S.; Schmidt, M.; Lehmann, T.; Irintchev, A.; Schubert, H.; Jung, C.; Schwab, M.; Huber, O.; Matziolis, G.; Schiffner, R. Increase of cortical cerebral blood flow and further cerebral microcirculatory effects of Serelaxin in a sheep model. Am. J. Physiol. Heart Circ. Physiol. 2016, 311, H613–H620. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Teerlink, J.R.; Metra, M.; Felker, G.M.; Ponikowski, P.; Voors, A.A.; Weatherley, B.D.; Marmor, A.; Katz, A.; Grzybowski, J.; Unemori, E.; et al. Relaxin for the treatment of patients with acute heart failure (Pre-RELAX-AHF): A multicentre, randomised, placebo-controlled, parallel-group, dose-finding phase IIb study. Lancet 2009, 373, 1429–1439. [Google Scholar] [CrossRef]
- Metra, M.; Teerlink, J.R.; Cotter, G.; Davison, B.A.; Felker, G.M.; Filippatos, G.; Greenberg, B.H.; Pang, P.S.; Ponikowski, P.; Voors, A.A.; et al. Effects of Serelaxin in Patients with Acute Heart Failure. N. Engl. J. Med. 2019, 381, 716–726. [Google Scholar] [CrossRef] [PubMed]
- Schiffner, R.; Reiche, J.; Schmidt, M.; Jung, C.; Walther, S.; Irintchev, A.; Bischoff, S.J. Pulmonary arterial compliance and pulmonary hemodynamic effects of Serelaxin in a sheep model. Clin. Hemorheol. Microcirc. 2017, 66, 219–229. [Google Scholar] [CrossRef]
- Schiffner, R.; Nistor, M.; Bischoff, S.J.; Matziolis, G.; Schmidt, M.; Lehmann, T. Effects of human relaxin-2 (serelaxin) on hypoxic pulmonary vasoconstriction during acute hypoxia in a sheep model. Hypoxia 2018, 6, 11–22. [Google Scholar] [CrossRef] [Green Version]
- Schiffner, R.; Lehmann, T.; Bischoff, S.J.; Zippelius, T.; Nistor, M.; Schmidt, M. Pulmonary hemodynamic effects and pulmonary arterial compliance during hypovolemic shock and reinfusion with human relaxin-2 (serelaxin) treatment in a sheep model. Clin. Hemorheol. Microcirc. 2018, 70, 311–325. [Google Scholar] [CrossRef]
- Nistor, M.; Behringer, W.; Schmidt, M.; Schiffner, R. A Systematic Review of Neuroprotective Strategies during Hypovolemia and Hemorrhagic Shock. Int. J. Mol. Sci. 2017, 18, 2247. [Google Scholar] [CrossRef] [Green Version]
- Fabricius, M.; Lauritzen, M. Laser-Doppler evaluation of rat brain microcirculation: Comparison with the [14C]-iodoantipyrine method suggests discordance during cerebral blood flow increases. J. Cereb. Blood Flow Metab. Off. J. Int. Soc. Cereb. Blood Flow Metab. 1996, 16, 156–161. [Google Scholar] [CrossRef] [Green Version]
- Schiffner, R.; Bischoff, S.J.; Lehmann, T.; Rakers, F.; Rupprecht, S.; Matziolis, G.; Schubert, H.; Schwab, M.; Huber, O.; Lemke, C.; et al. Underlying mechanism of subcortical brain protection during hypoxia and reoxygenation in a sheep model-Influence of alpha1-adrenergic signalling. PLoS ONE 2018, 13, e0196363. [Google Scholar] [CrossRef] [Green Version]
- Schiffner, R.; Bischoff, S.J.; Lehmann, T.; Rakers, F.; Rupprecht, S.; Reiche, J.; Matziolis, G.; Schubert, H.; Schwab, M.; Huber, O.; et al. Redistribution of Cerebral Blood Flow during Severe Hypovolemia and Reperfusion in a Sheep Model: Critical Role of alpha1-Adrenergic Signaling. Int. J. Mol. Sci. 2017, 18, 1031. [Google Scholar] [CrossRef] [Green Version]
- Lewis, N.C.; Messinger, L.; Monteleone, B.; Ainslie, P.N. Effect of acute hypoxia on regional cerebral blood flow: Effect of sympathetic nerve activity. J. Appl. Physiol. 2014, 116, 1189–1196. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fisher, C.; MacLean, M.; Morecroft, I.; Seed, A.; Johnston, F.; Hillier, C.; McMurray, J. Is the pregnancy hormone relaxin also a vasodilator peptide secreted by the heart? Circulation 2002, 106, 292–295. [Google Scholar] [CrossRef] [PubMed]
- Dahlke, M.; Halabi, A.; Canadi, J.; Tsubouchi, C.; Machineni, S.; Pang, Y. Pharmacokinetics of serelaxin in patients with severe renal impairment or end-stage renal disease requiring hemodialysis: A single-dose, open-label, parallel-group study. J. Clin. Pharmacol. 2016, 56, 474–483. [Google Scholar] [CrossRef] [PubMed]
- Noziere, P.; Remond, D.; Ferlay, A.; Doreau, M. Differences between blood and plasma concentrations of acetate, beta-hydroxybutyrate, glucose, ammonia and urea: Implications for measurement of portal net fluxes in ewes. Reprod. Nutr. Dev. 1998, 38, 509–518. [Google Scholar] [CrossRef] [PubMed]
- Niwa, K.; Takizawa, S.; Takagi, S.; Shinohara, Y. Mild hypothermia disturbs regional cerebrovascular autoregulation in awake rats. Brain Res. 1998, 789, 68–73. [Google Scholar] [CrossRef]
- Waschke, K.F.; Riedel, M.; Lenz, C.; Albrecht, D.M.; van Ackern, K.; Kuschinsky, W. Regional heterogeneity of cerebral blood flow response to graded pressure-controlled hemorrhage. J. Trauma 2004, 56, 591–603. [Google Scholar] [CrossRef]
- Willie, C.K.; Macleod, D.B.; Shaw, A.D.; Smith, K.J.; Tzeng, Y.C.; Eves, N.D.; Ikeda, K.; Graham, J.; Lewis, N.C.; Day, T.A.; et al. Regional brain blood flow in man during acute changes in arterial blood gases. J. Physiol. 2012, 590, 3261–3275. [Google Scholar] [CrossRef] [Green Version]
- Kick, O.; Van Aken, H.; Wouters, P.F.; Verbesselt, K.; Van Hemelrijck, J. Vital organ blood flow during deliberate hypotension in dogs. Anesth. Analg. 1993, 77, 737–742. [Google Scholar] [CrossRef]
- Verge, D.; Daval, G.; Marcinkiewicz, M.; Patey, A.; el Mestikawy, S.; Gozlan, H.; Hamon, M. Quantitative autoradiography of multiple 5-HT1 receptor subtypes in the brain of control or 5,7-dihydroxytryptamine-treated rats. J Neurosci. 1986, 6, 3474–3482. [Google Scholar] [CrossRef]
- Agoston, D.V.; Shutes-David, A.; Peskind, E.R. Biofluid biomarkers of traumatic brain injury. Brain Inj. 2017, 31, 1195–1203. [Google Scholar] [CrossRef] [Green Version]
- Oris, C.; Pereira, B.; Durif, J.; Simon-Pimmel, J.; Castellani, C.; Manzano, S.; Sapin, V.; Bouvier, D. The Biomarker S100B and Mild Traumatic Brain Injury: A Meta-analysis. Pediatrics 2018, 141. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, K.K.; Yang, Z.; Zhu, T.; Shi, Y.; Rubenstein, R.; Tyndall, J.A.; Manley, G.T. An update on diagnostic and prognostic biomarkers for traumatic brain injury. Expert Rev. Mol. Diagn. 2018, 18, 165–180. [Google Scholar] [CrossRef] [PubMed]
- Christl, J.; Verhulsdonk, S.; Pessanha, F.; Menge, T.; Seitz, R.J.; Kujovic, M.; Hoft, B.; Supprian, T.; Lange-Asschenfeldt, C. Association of Cerebrospinal Fluid S100B Protein with Core Biomarkers and Cognitive Deficits in Prodromal and Mild Alzheimer’s Disease. J. Alzheimers Dis. 2019, 72, 1119–1127. [Google Scholar] [CrossRef] [PubMed]
- Silva, F.P.; Schmidt, A.P.; Valentin, L.S.; Pinto, K.O.; Zeferino, S.P.; Oses, J.P.; Wiener, C.D.; Otsuki, D.A.; Tort, A.B.; Portela, L.V.; et al. S100B protein and neuron-specific enolase as predictors of cognitive dysfunction after coronary artery bypass graft surgery: A prospective observational study. Eur. J. Anaesthesiol. 2016, 33, 681–689. [Google Scholar] [CrossRef] [PubMed]
- Mendes Arent, A.; de Souza, L.F.; Walz, R.; Dafre, A.L. Perspectives on molecular biomarkers of oxidative stress and antioxidant strategies in traumatic brain injury. BioMed Res. Int. 2014, 2014, 723060. [Google Scholar] [CrossRef]
- Thebaud, B.; Husson, B.; Navelet, Y.; Huault, G.; Landrieu, P.; Devictor, D.; Sebire, G. Haemorrhagic shock and encephalopathy syndrome: Neurological course and predictors of outcome. Intensive Care Med. 1999, 25, 293–299. [Google Scholar] [CrossRef]
- Heckbert, S.R.; Vedder, N.B.; Hoffman, W.; Winn, R.K.; Hudson, L.D.; Jurkovich, G.J.; Copass, M.K.; Harlan, J.M.; Rice, C.L.; Maier, R.V. Outcome after hemorrhagic shock in trauma patients. J. Trauma 1998, 45, 545–549. [Google Scholar] [CrossRef]
- Kudo, Y.; Ohtaki, H.; Dohi, K.; Yin, L.; Nakamachi, T.; Endo, S.; Yofu, S.; Hiraizumi, Y.; Miyaoka, H.; Shioda, S. Neuronal damage in rat brain and spinal cord after cardiac arrest and massive hemorrhagic shock. Crit. Care Med. 2006, 34, 2820–2826. [Google Scholar] [CrossRef] [Green Version]
- Teerlink, J.R.; Cotter, G.; Davison, B.A.; Felker, G.M.; Filippatos, G.; Greenberg, B.H.; Ponikowski, P.; Unemori, E.; Voors, A.A.; Adams, K.F., Jr.; et al. Serelaxin, recombinant human relaxin-2, for treatment of acute heart failure (RELAX-AHF): A randomised, placebo-controlled trial. Lancet 2013, 381, 29–39. [Google Scholar] [CrossRef]
- Bathgate, R.A.; Halls, M.L.; van der Westhuizen, E.T.; Callander, G.E.; Kocan, M.; Summers, R.J. Relaxin family peptides and their receptors. Physiol. Rev. 2013, 93, 405–480. [Google Scholar] [CrossRef]
- Chan, S.L.; Sweet, J.G.; Cipolla, M.J. Treatment for cerebral small vessel disease: Effect of relaxin on the function and structure of cerebral parenchymal arterioles during hypertension. FASEB J. Off. Publ. Fed. Am. Soc. Exp. Biol. 2013, 27, 3917–3927. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chan, S.L.; Cipolla, M.J. Relaxin causes selective outward remodeling of brain parenchymal arterioles via activation of peroxisome proliferator-activated receptor-gamma. FASEB J. Off. Publ. Fed. Am. Soc. Exp. Biol. 2011, 25, 3229–3239. [Google Scholar] [CrossRef] [Green Version]
- Debrah, D.O.; Novak, J.; Matthews, J.E.; Ramirez, R.J.; Shroff, S.G.; Conrad, K.P. Relaxin is essential for systemic vasodilation and increased global arterial compliance during early pregnancy in conscious rats. Endocrinology 2006, 147, 5126–5131. [Google Scholar] [CrossRef] [Green Version]
- Halls, M.L.; Bathgate, R.A.; Sutton, S.W.; Dschietzig, T.B.; Summers, R.J. International Union of Basic and Clinical Pharmacology. XCV. Recent advances in the understanding of the pharmacology and biological roles of relaxin family peptide receptors 1-4, the receptors for relaxin family peptides. Pharmacol. Rev. 2015, 67, 389. [Google Scholar] [CrossRef] [PubMed]
- Worlein, J.M.; Baker, K.; Bloomsmith, M.; Coleman, K.; Koban, T.L. The Eighth Edition of the Guide for the Care and Use of Laboratory Animals; Implications for Behavioral Management. Am. J. Primatol. 2011, 73, 98. [Google Scholar]
- Gutierrez, G.; Reines, H.D.; Wulf-Gutierrez, M.E. Clinical review: Hemorrhagic shock. Crit. Care 2004, 8, 373–381. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Laemmli, U.K. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature 1970, 227, 680–685. [Google Scholar] [CrossRef]
- Kyhse-Andersen, J. Electroblotting of multiple gels: A simple apparatus without buffer tank for rapid transfer of proteins from polyacrylamide to nitrocellulose. J. Biochem. Biophys. Methods 1984, 10, 203–209. [Google Scholar] [CrossRef]
- Irintchev, A.; Rollenhagen, A.; Troncoso, E.; Kiss, J.Z.; Schachner, M. Structural and functional aberrations in the cerebral cortex of tenascin-C deficient mice. Cereb. Cortex 2005, 15, 950–962. [Google Scholar] [CrossRef]













© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Schiffner, R.; Bischoff, S.J.; Lehmann, T.; Irintchev, A.; Nistor, M.; Lemke, C.; Schmidt, M. Altered Cerebral Blood Flow and Potential Neuroprotective Effect of Human Relaxin-2 (Serelaxin) During Hypoxia or Severe Hypovolemia in a Sheep Model. Int. J. Mol. Sci. 2020, 21, 1632. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21051632
Schiffner R, Bischoff SJ, Lehmann T, Irintchev A, Nistor M, Lemke C, Schmidt M. Altered Cerebral Blood Flow and Potential Neuroprotective Effect of Human Relaxin-2 (Serelaxin) During Hypoxia or Severe Hypovolemia in a Sheep Model. International Journal of Molecular Sciences. 2020; 21(5):1632. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21051632
Chicago/Turabian StyleSchiffner, René, Sabine J. Bischoff, Thomas Lehmann, Andrey Irintchev, Marius Nistor, Cornelius Lemke, and Martin Schmidt. 2020. "Altered Cerebral Blood Flow and Potential Neuroprotective Effect of Human Relaxin-2 (Serelaxin) During Hypoxia or Severe Hypovolemia in a Sheep Model" International Journal of Molecular Sciences 21, no. 5: 1632. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21051632