Deciphering the Role of Multiple Thioredoxin Fold Proteins of Leptospirillum sp. in Oxidative Stress Tolerance

Abstract
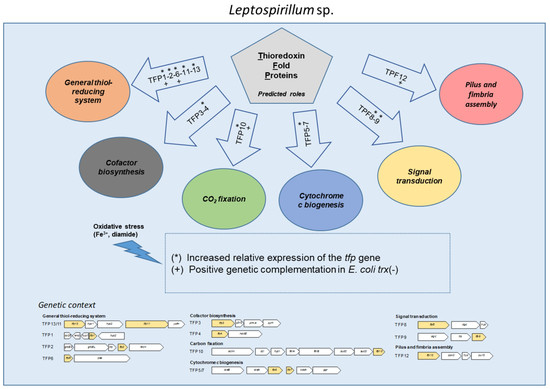
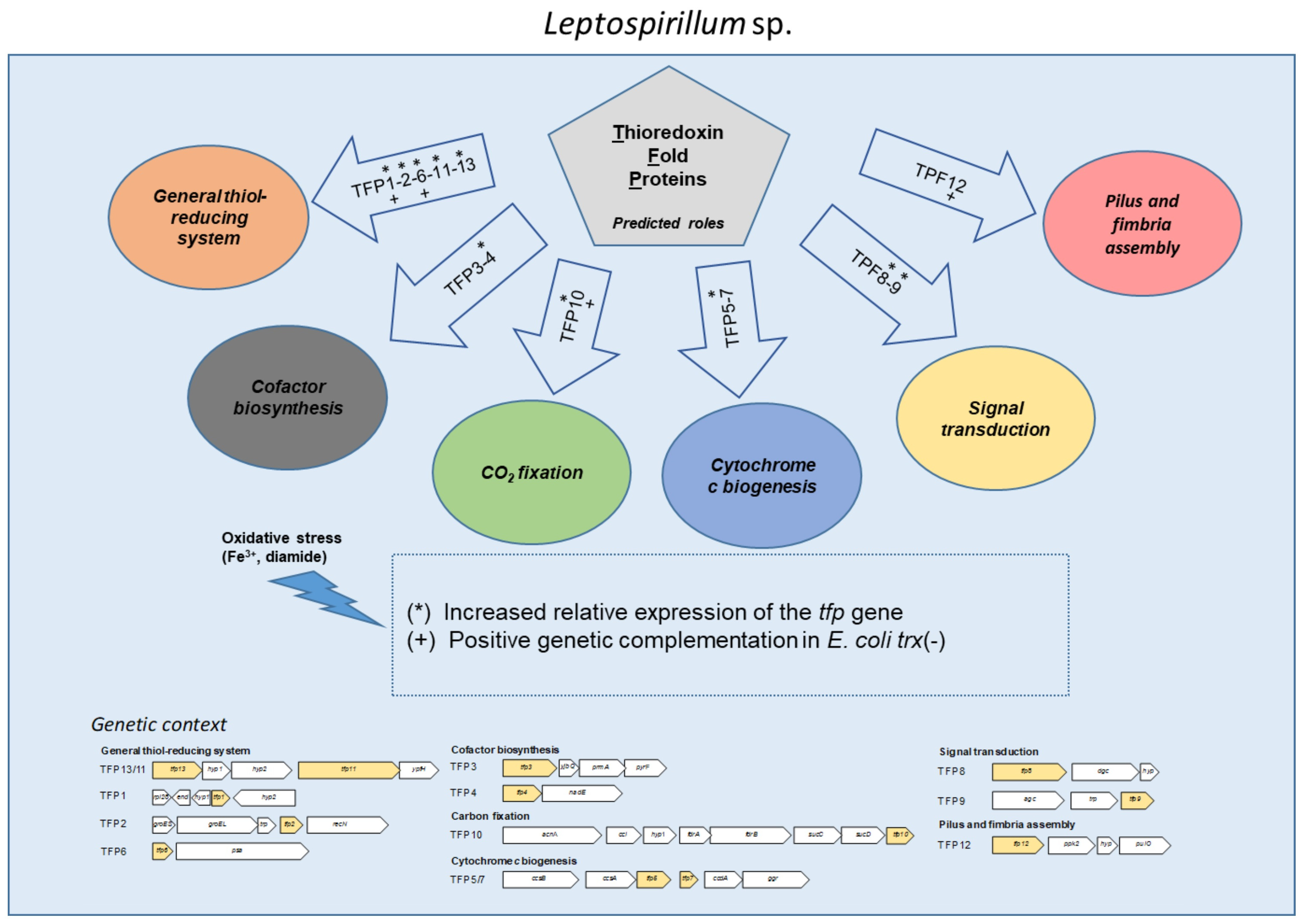
:
1. Introduction
2. Results
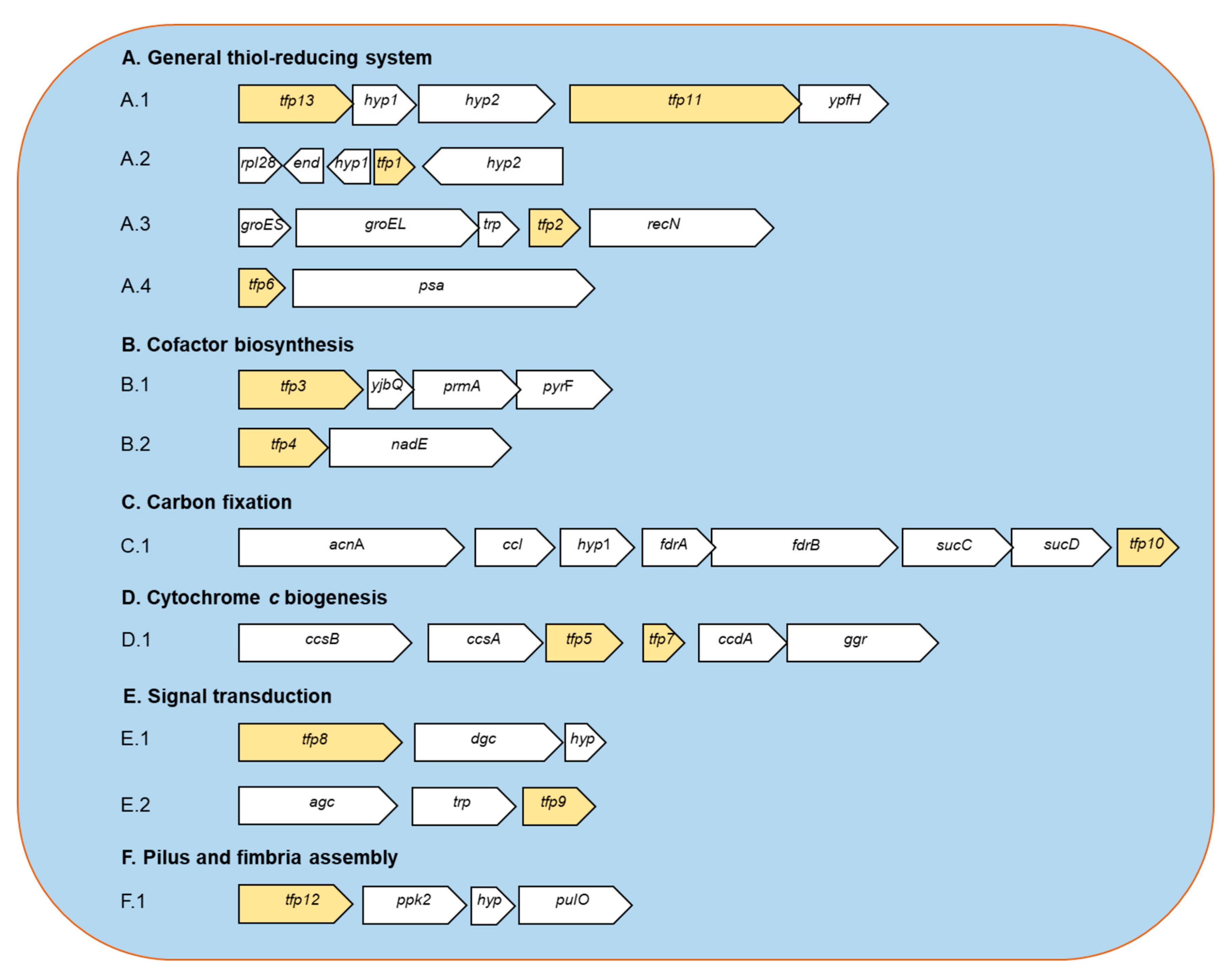
2.1. In Silico Analysis of the Genomic Context and Prediction of the Roles of the tfp Genes
2.1.1. General Thiol-Reducing System (TFP1-TFP2-TFP6-TFP11-TFP13)
2.1.2. Cofactor Biosynthesis (TFP3 and TFP4)
2.1.3. CO2 Fixation (TFP10)
2.1.4. Cytochrome c Biogenesis (TFP5 and TFP7)
2.1.5. Signal Transduction (TFP8 and TFP9)
2.1.6. Pilus and Fimbria Assembly (TFP12)
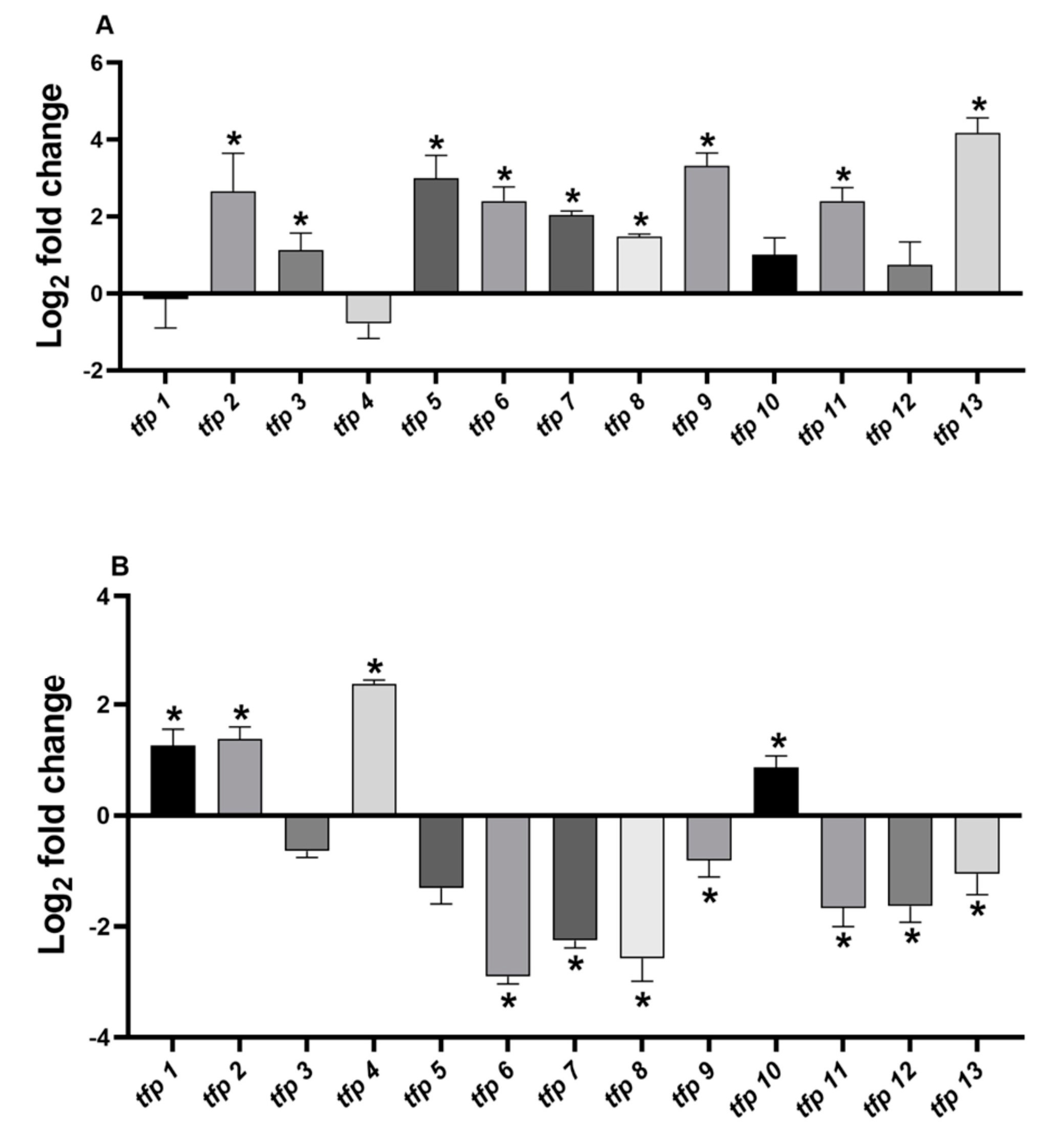
2.2. The Expression of tfp Genes from Leptospirillum sp. CF-1 Is Differentially Regulated under Oxidative Stress Conditions
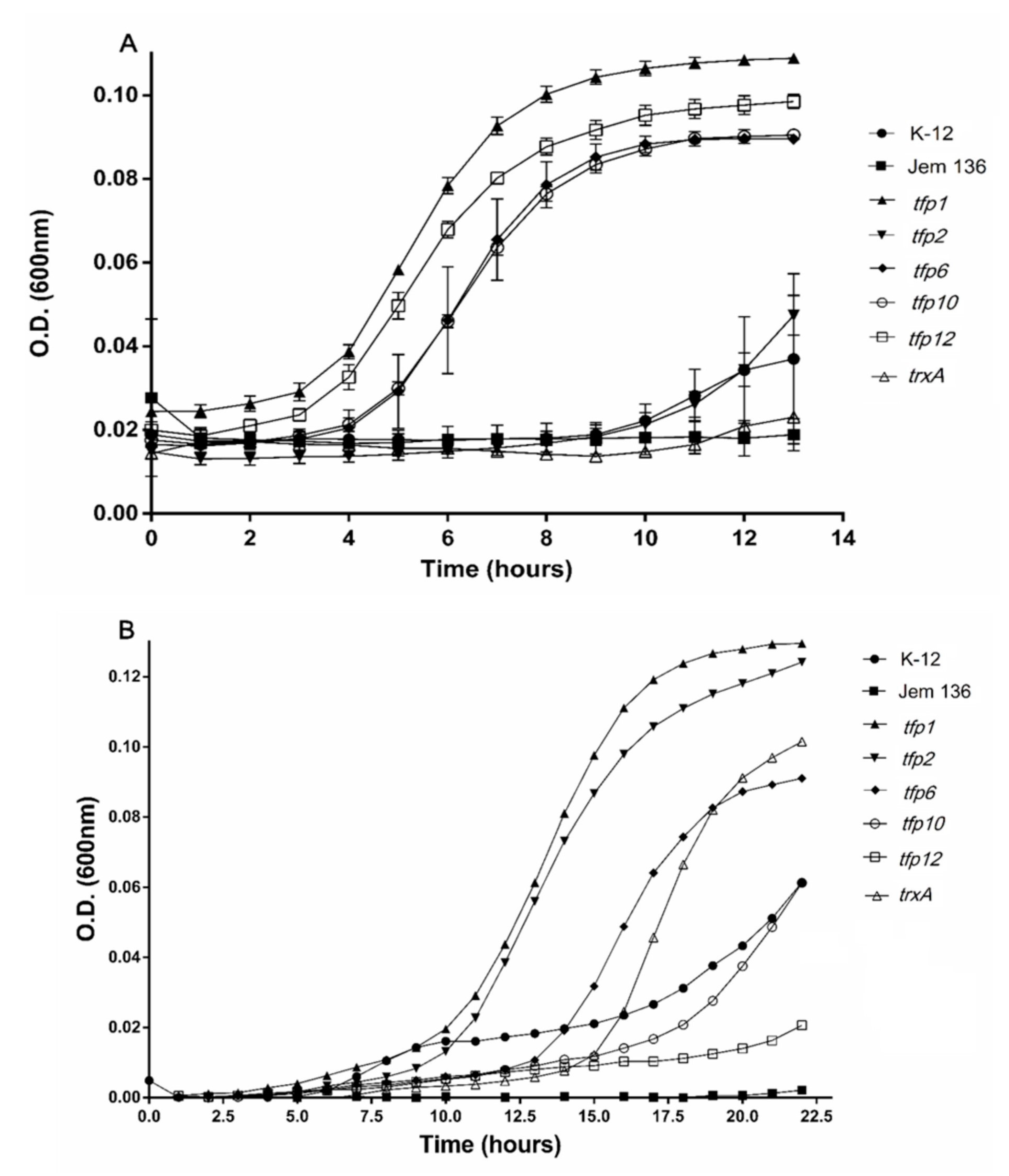
2.3. Tfp Genes from Leptospirillum sp. CF-1 Restored the Response to Oxidative Stress of a Thioredoxin-Deficient Strain of E. coli
3. Discussion
4. Materials and Methods
4.1. Cell Cultures
4.2. Induction of Stress in Leptospirillum sp. CF-1
4.3. RNA Extraction and cDNA Synthesis
4.4. RT-qPCR and PCR
4.5. Cloning of tfp Genes in an Expression Vector
4.6. Determination of Minimal Inhibitory Concentration
4.7. Functional Assays of tfp Genes from the Strain CF-1 in E. coli
4.8. Sequence Alignments and Motif Search
4.9. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Martin, J. Thioredoxin-a fold for all reasons. Structure 1995, 3, 245–250. [Google Scholar] [CrossRef]
- Meyer, Y.; Buchanan, B.; Vignols, F.; Reichheld, J. Thioredoxins and Glutaredoxins: Unifying elements in redox biology. Ann. Rev. Gen. 2009, 43, 335–367. [Google Scholar] [CrossRef]
- Atkinson, H.; Babbitt, P. An atlas of the thioredoxin fold class reveals the complexity of function-enabling adaptations. PLoS Comput. Biol. 2009, 5, e1000541. [Google Scholar] [CrossRef] [Green Version]
- Lu, J.; Holmgren, A. The thioredoxin superfamily in oxidative protein folding. Antioxid. Redox Signal. 2014, 21, 457–470. [Google Scholar] [CrossRef]
- Ren, G.; Stephan, D.; Xu, Z.; Zheng, Y.; Tang, D.; Harrison, R.S.; Kurz, M.; Jarrott, R.; Shouldice, S.R.; Hiniker, A.; et al. Properties of the thioredoxin fold superfamily are modulated by a single amino acid residue. J. Biol. Chem. 2009, 284, 10150–10159. [Google Scholar] [CrossRef] [Green Version]
- Netto, L.E.; Antunes, F. The Roles of peroxiredoxin and thioredoxin in hydrogen peroxide sensing and in signal transduction. Mol. Cells 2016, 39, 65–71. [Google Scholar] [PubMed] [Green Version]
- Feng, X.; Sun, W.; Kong, L.; Gao, H. Distinct roles of Shewanella oneidensis thioredoxin in regulation of cellular responses to hydrogen and organic peroxides. App. Environ. Microbiol. 2019, 85, e01700-19. [Google Scholar] [CrossRef] [PubMed]
- Da Silva Neto, J.F.; Negretto, C.C.; Netto, L.E. Analysis of the organic hydroperoxide response of Chromobacterium violaceum reveals that OhrR is a cys-based redox sensor regulated by thioredoxin. PLoS ONE 2012, 7, e47090. [Google Scholar] [CrossRef] [PubMed]
- Kadowaki, T.; Nishiyama, Y.; Hisabori, T.; Hihara, Y. Correction: Identification of OmpR-family response regulators interacting with thioredoxin in the cyanobacterium Synechocystis sp. PCC 6803. PLoS ONE 2015, 10, e0124571. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khairnar, N.P.; Joe, M.H.; Misra, H.S.; Lim, S.Y.; Kim, D.H. FrnE, a cadmium-inducible protein in Deinococcus radiodurans, is characterized as a disulfide isomerase chaperone in vitro and for its role in oxidative stress tolerance in vivo. J. Bacteriol. 2013, 195, 2880–2886. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harrison, A.; Baker, B.D.; Munson, R.S., Jr. Overlapping and complementary oxidative stress defense mechanisms in nontypeable Haemophilus influenzae. J. Bacteriol. 2015, 197, 277–285. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, L.B.; Tang, L.; Ying, S.H.; Feng, M.G. Regulative roles of glutathione reductase and four glutaredoxins in glutathione redox, antioxidant activity, and iron homeostasis of Beauveria bassiana. Appl. Microbiol. Biotechnol. 2016, 100, 5907–5917. [Google Scholar] [CrossRef] [PubMed]
- Hamed, S.M.; Selim, S.; Klöck, G.; AbdElgawad, H. Sensitivity of two green microalgae to copper stress: Growth, oxidative and antioxidants analyses. Ecotoxicol. Environ. Saf. 2017, 144, 19–25. [Google Scholar] [CrossRef] [PubMed]
- Atichartpongkul, S.; Vattanaviboon, P.; Wisitkamol, R.; Jaroensuk, J.; Mongkolsuk, S.; Fuangthong, M. Regulation of organic hydroperoxide stress response by two OhrR homologs in Pseudomonas aeruginosa. PLoS ONE 2016, 11, e0161982. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goemans, C.V.; Vertommen, D.; Agrebi, R.; Collet, J.F. CnoX is a chaperedoxin: A holdase that protects its substrates from irreversible oxidation. Mol. Cell 2018, 70, 614–627. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cárdenas, J.P.; Moya, F.; Covarrubias, P.; Shmaryahu, A.; Levicán, G.; Holmes, D.; Quatrini, R. Comparative genomics of the oxidative stress response in bioleaching microorganisms. Hydrometallurgy 2012, 127–128, 162–167. [Google Scholar] [CrossRef]
- Ferrer, A.; Orellana, O.; Levicán, G. Oxidative stress and metal tolerance. In Acidophiles: Life in Extremely Acidic Environments, 1st ed.; Johnson, D.B., Quatrini, R., Eds.; Horizon Scientific Press: Norfolk, UK, 2016; pp. 63–76. [Google Scholar]
- Norambuena, J.; Flores, R.; Cárdenas, J.P.; Quatrini, R.; Chávez, R.; Levicán, G. Thiol/disulfide system plays a crucial role in redox protection in the acidophilic iron-oxidizing bacterium Leptospirillum ferriphilum. PLoS ONE 2012, 7, e44576. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zapata, C.; Paillavil, B.; Chávez, R.; Álamos, P.; Levicán, G. Cytochrome c peroxidase (CcP) is a molecular determinant of the oxidative stress response in the extreme acidophilic Leptospirillum sp. CF-1. FEMS Microbiol. Ecol. 2017, 93. [Google Scholar] [CrossRef] [Green Version]
- Ferrer, A.; Bunk, B.; Spröer, C.; Biedendieck, R.; Valdés, N.; Jahn, M.; Jahn, D.; Orellana, O.; Levicán, G. Complete genome sequence of the bioleaching bacterium Leptospirillum sp. group II strain CF-1. J. Biotechnol. 2016, 222, 21–22. [Google Scholar] [CrossRef]
- Lemaire, S.D.; Collin, V.; Keryer, E.; Quesada, A.; Miginiac-Maslow, M. Characterization of thioredoxin y, a new type of thioredoxin identified in the genome of Chlamydomonas reinhardtii. FEBS Lett. 2003, 543, 87–92. [Google Scholar] [CrossRef] [Green Version]
- Mora-García, S.; Rodríguez-Suárez, R.; Wolosiuk, R.A. Role of electrostatic interactions on the affinity of thioredoxin for target proteins. Recognition of chloroplast fructose-1, 6-bisphosphatase by mutant Escherichia coli thioredoxins. J. Biol. Chem. 1998, 273, 16273–16280. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Castanié-Cornet, M.P.; Bruel, N.; Genevaux, P. Chaperone networking facilitates protein targeting to the bacterial cytoplasmic membrane. Biochim. Biophys. Acta 2014, 1843, 1442–1456. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, J.; Wilson, M.A. Escherichia coli thioredoxin-like protein YbbN contains an atypical tetratricopeptide repeat motif and is a negative regulator of GroEL. J. Biol. Chem. 2011, 286, 19459–19469. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Uranga, L.A.; Reyes, E.D.; Patidar, P.L.; Redman, L.N.; Lusetti, S.L. The cohesin-like RecN protein stimulates RecA-mediated recombinational repair of DNA double-strand breaks. Nat. Commun. 2017, 8, 15282. [Google Scholar] [CrossRef]
- Gumpena, R.; Kishor, C.; Ganji, R.J.; Jain, N.; Addlagatta, A. Glu121-Lys319 salt bridge between catalytic and N-terminal domains is pivotal for the activity and stability of Escherichia coli aminopeptidase N. Protein Sci. 2012, 21, 727–736. [Google Scholar] [CrossRef] [Green Version]
- Chandu, D.; Nandi, D. PepN is the major aminopeptidase in Escherichia coli: Insights on substrate specificity and role during sodium-salicylate-induced stress. Microbiology 2003, 149, 3437–3447. [Google Scholar] [CrossRef] [Green Version]
- Foglino, M.; Gharbi, S.; Lazdunski, A. Nucleotide sequence of the pepN gene encoding aminopeptidase N of Escherichia coli. Gene 1986, 49, 303–309. [Google Scholar] [CrossRef]
- Ito, K.; Nakajima, Y.; Onohara, Y.; Takeo, M.; Nakashima, K.; Matsubara, F.; Ito, T.; Yoshimoto, T. Crystal structure of aminopeptidase N (proteobacteria alanyl aminopeptidase) from Escherichia coli and conformational change of methionine 260 involved in substrate recognition. J. Biol. Chem. 2006, 281, 33664–33676. [Google Scholar] [CrossRef] [Green Version]
- Lockwood, T.D. Redox control of protein degradation. Antioxid. Redox Signal. 2000, 2, 851–878. [Google Scholar] [CrossRef] [Green Version]
- Morett, E.; Saab-Rincón, G.; Olvera, L.; Olvera, M.; Flores, H.; Grande, R. Sensitive genome-wide screen for low secondary enzymatic activities: The YjbQ family shows thiamin phosphate synthase activity. J. Mol. Biol. 2008, 376, 839–853. [Google Scholar] [CrossRef]
- Mohorko, E.; Abicht, H.K.; Bühler, D.; Glockshuber, R.; Hennecke, H.; Fischer, H.M. Thioredoxin-like protein TlpA from Bradyrhizobium japonicum is a reductant for the copper metallochaperone ScoI. FEBS Lett. 2012, 586, 4094–4099. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xiao, W.; Wang, R.S.; Handy, D.E.; Loscalzo, J. NAD(H) and NADP(H) Redox couples and cellular energy metabolism. Antioxid. Redox Signal. 2018, 28, 251–272. [Google Scholar] [CrossRef] [PubMed]
- Bhat, S.A.; Iqbal, I.K.; Kumar, A. Imaging the NADH:NAD+ homeostasis for understanding the metabolic response of Mycobacterium to physiologically relevant stresses. Front. Cell Infect. Microbiol. 2016, 6, 145. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lindahl, M.; Florencio, F.J. Thioredoxin-linked processes in cyanobacteria are as numerous as in chloroplasts, but targets are different. Proc. Natl. Acad. Sci. USA 2003, 100, 16107–16112. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Levicán, G.; Ugalde, J.A.; Ehrenfeld, N.; Maass, A.; Parada, P. Comparative genomic analysis of carbon and nitrogen assimilation mechanisms in three indigenous bioleaching bacteria: Predictions and validations. BMC Genom. 2008, 9, 581–599. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- König, J.; Baier, M.; Horling, F.; Kahmann, U.; Harris, G.; Schürmann, P.; Dietz, K.J. The plant-specific function of 2-Cys peroxiredoxin-mediated detoxification of peroxides in the redox-hierarchy of photosynthetic electron flux. Proc. Natl. Acad. Sci. USA 2002, 99, 5738–5743. [Google Scholar] [CrossRef] [Green Version]
- Tretter, L.; Adam-Vizi, V. Inhibition of Krebs cycle enzymes by hydrogen peroxide: A key role of [alpha]-ketoglutarate dehydrogenase in limiting NADH production under oxidative stress. J. Neurosci. 2000, 20, 8972–8979. [Google Scholar] [CrossRef]
- Kranz, R.G.; Richard-Fogal, C.; Taylor, J.S.; Frawley, E.R. Cytochrome c biogenesis: Mechanisms for covalent modifications and trafficking of heme and for heme-iron redox control. Microbiol. Mol. Biol. Rev. 2009, 73, 510–528. [Google Scholar] [CrossRef] [Green Version]
- Levicán, G.; Gómez, M.J.; Chávez, R.; Orellana, O.; Moreno-Paz, M.; Parro, V. Comparative genomic analysis reveals novel facts about Leptospirillum spp. cytochromes. J. Mol. Microbiol. Biotechnol. 2012, 22, 94–104. [Google Scholar] [CrossRef]
- Lübben, M.; Morand, K. Novel prenylated hemes as cofactors of cytochrome oxidases. Archaea have modified hemes A and O. J. Biol. Chem. 1994, 269, 21473–21479. [Google Scholar]
- Opoku-Temeng, C.; Sintim, H.O. Targeting c-di-GMP signaling, biofilm formation, and bacterial motility with small molecules. Methods Mol. Biol. 2017, 1657, 419–430. [Google Scholar] [PubMed]
- Díaz, M.; Castro, M.; Copaja, S.; Guiliani, N. Biofilm formation by the acidophile bacterium Acidithiobacillus thiooxidans involves c-di-GMP pathway and pel exopolysaccharide. Genes (Basel) 2018, 9, E113. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cruz, D.P.; Huertas, M.G.; Lozano, M.; Zárate, L.; Zambrano, M.M. Comparative analysis of diguanylate cyclase and phosphodiesterase genes in Klebsiella pneumoniae. BMC Microbiol. 2012, 12, 139. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Green, J.; Stapleton, M.R.; Smith, L.J.; Artymiuk, P.J.; Kahramanoglou, C.; Hunt, D.M.; Buxton, R.S. Cyclic-AMP and bacterial cyclic-AMP receptor proteins revisited: Adaptation for different ecological niches. Curr. Opin. Microbiol. 2014, 18, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Chi, A.; Valenzuela, L.; Beard, S.; Mackey, A.J.; Shabanowitz, J.; Hunt, D.F.; Jerez, C.A. Periplasmic proteins of the extremophile Acidithiobacillus ferrooxidans: A high throughput proteomics analysis. Mol. Cell Proteom. 2007, 6, 2239–2251. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Guo, J.; Zhang, N.; Yuan, W.; Lin, Z.; Huang, W. Characterization and analysis of a novel diguanylate cyclase PA0847 from Pseudomonas aeruginosa PAO1. Infect. Drug Resist. 2019, 12, 655–665. [Google Scholar] [CrossRef] [Green Version]
- Huang, C.J.; Wang, Z.C.; Huang, H.Y.; Huang, H.D.; Peng, H.L. YjcC, a c-di-GMP phosphodiesterase protein, regulates the oxidative stress response and virulence of Klebsiella pneumoniae CG43. PLoS ONE 2013, 8, e66740. [Google Scholar] [CrossRef] [Green Version]
- Buensuceso, R.N.; Nguyen, Y.; Zhang, K.; Daniel-Ivad, M.; Sugiman-Marangos, S.N.; Fleetwood, A.D.; Zhulin, I.B.; Junop, M.S.; Howell, P.L.; Burrows, L.L. The conserved tetratricopeptide repeat-containing c-terminal domain of Pseudomonas aeruginosa FimV is required for its cyclic amp-dependent and -independent functions. J. Bacteriol. 2016, 198, 2263–2274. [Google Scholar] [CrossRef] [Green Version]
- Kornberg, A.; Rao, N.N.; Ault-Riché, D. Inorganic polyphosphate: A molecule of many functions. Annu. Rev. Biochem. 1999, 68, 89–125. [Google Scholar] [CrossRef]
- Orell, A.; Navarro, C.A.; Rivero, M.; Aguilar, J.S.; Jerez, C.A. Inorganic polyphosphates in extremophiles and their possible functions. Extremophiles 2012, 16, 573–583. [Google Scholar] [CrossRef]
- Ishige, K.; Zhang, H.; Kornberg, A. Polyphosphate kinase (PPK2), a potent polyphosphate-driven generator of GTP. Proc. Natl. Acad. Sci. USA 2002, 99, 16684–16688. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- LaPointe, C.F.; Taylor, R.K. The type 4 prepilin peptidases comprise a novel family of aspartic acid proteases. J. Biol. Chem. 2000, 275, 1502–1510. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Korotkov, K.V.; Sandkvist, M.; Hol, W.G. The type II secretion system: Biogenesis, molecular architecture and mechanism. Nat. Rev. Microbiol. 2012, 10, 336–351. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lewenza, S.; Gardy, J.L.; Brinkman, F.S.; Hancock, R.E. Genome-wide identification of Pseudomonas aeruginosa exported proteins using a consensus computational strategy combined with a laboratory-based PhoA fusion screen. Genome Res. 2005, 15, 321–329. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, H.Z.; Donnenberg, M.S. DsbA is required for stability of the type IV pilin of enteropathogenic Escherichia coli. Mol. Microbiol. 1996, 21, 787–797. [Google Scholar] [CrossRef] [Green Version]
- Prieto-Alamo, M.J.; Jurado, J.; Gallardo-Madueno, R.; Monje-Casas, F.; Holmgren, A.; Pueyo, C. Transcriptional regulation of glutaredoxin and thioredoxin pathways and related enzymes in response to oxidative stress. J. Biol. Chem. 2000, 275, 13398–13405. [Google Scholar] [CrossRef] [Green Version]
- Cha, J.Y.; Kim, J.Y.; Jung, I.J.; Kim, M.R.; Melencion, A.; Alam, S.S.; Yun, D.J.; Lee, S.Y.; Kim, M.G.; Kim, W.Y. NADPH-dependent thioredoxin reductase A (NTRA) confers elevated tolerance to oxidative stress and drought. Plant Physiol. Biochem. 2014, 80, 184–191. [Google Scholar] [CrossRef] [PubMed]
- McCarver, A.C.; Lessner, F.H.; Soroeta, J.M.; Lessner, D.J. Methanosarcina acetivorans utilizes a single NADPH-dependent thioredoxin system and contains additional thioredoxin homologues with distinct functions. Microbiology 2017, 163, 62–74. [Google Scholar] [CrossRef]
- Kosower, N.S.; Kosower, E.M. Diamide: An oxidant probe for thiols. Methods Enzymol. 1995, 251, 123–133. [Google Scholar]
- Reott, M.A.; Parker, A.C.; Rocha, E.R.; Smith, C.J. Thioredoxins in redox maintenance and survival during oxidative stress of Bacteroides fragilis. J. Bacteriol. 2009, 191, 3384–3391. [Google Scholar] [CrossRef] [Green Version]
- McCarver, A.C.; Lessner, D.J. Molecular characterization of the thioredoxin system from Methanosarcina acetivorans. FEBS J. 2014, 281, 4598–4611. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Berndt, C.; Lillig, C.H. Glutathione, glutaredoxins, and iron. Antioxid. Redox Signal. 2017, 27, 1235–1251. [Google Scholar] [CrossRef] [PubMed]
- Ezraty, B.; Gennaris, A.; Barras, F.; Collet, J.F. Oxidative stress, protein damage and repair in bacteria. Nat. Rev. Microbiol. 2017, 15, 385–396. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| TFP | Accession Number | pI/MW | Cellular Location | Protein | Genomic Context-Derived Function |
|---|---|---|---|---|---|
| 1 | WP_023525652.1 | 9.47/9512.98 | ND | Thioredoxin-like | General thiol-reductase system |
| 2 | WP_014960847.1 | 5.42/11712.63 | Cytoplasm | Thioredoxin-like | General thiol-reductase system |
| 3 | WP_023524535.1 | 7.73/38929.40 | ND | Thiol disulfide oxidoreductase | Thiamine biosynthesis |
| 4 | WP_023525480.1 | 9.30/23548.14 | Cytoplasmic membrane | Thioredoxin (Trx2)-like proteins | NAD+ biosynthesis |
| 5 | WP_014962035.1 | 9.08/22475.37 | Cytoplasmic membrane | Thioredoxin fold protein CcsX | Cytochrome c biogenesis |
| 6 | WP_036080148.1 | 5.70/17114.98 | Cytoplasm | Thioredoxin-like | General thiol-reductase system |
| 7 | WP_049713760.1 | 8.60/12904.08 | Cytoplasmic membrane | Thioredoxin-like protein | Cytochrome c biogenesis |
| 8 | WP_023524407.1 | 5.34/4016.03 | Cytoplasm | Thiol/Disulfide oxidoreductase | Signal transduction |
| 9 | WP_042225252.1 | 5.68/24829.29 | Cytoplasm | Thiol/Disulfide oxidase | Signal transduction |
| 10 | WP_014960570.1 | 6.82/17281.80 | Cytoplasm | 2-Cys peroxiredoxin | CO2 fixation |
| 11 | WP_053094110.1 | 5.56/79330.91 | Cytoplasm | Thioredoxin domain-containing protein YyaL | General thiol-reductase system |
| 12 | WP_038505022.1 | 6.18/25842.79 | Cytoplasm | Cytoplasmic thiol disulfide oxidoreductase | Pilus/fimbria assembly |
| 13 | WP_014962072.1 | 5.91/33482.30 | Cytoplasm | Thioredoxin reductase TrxB | General thiol-reductase system |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
González, D.; Álamos, P.; Rivero, M.; Orellana, O.; Norambuena, J.; Chávez, R.; Levicán, G. Deciphering the Role of Multiple Thioredoxin Fold Proteins of Leptospirillum sp. in Oxidative Stress Tolerance. Int. J. Mol. Sci. 2020, 21, 1880. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21051880
González D, Álamos P, Rivero M, Orellana O, Norambuena J, Chávez R, Levicán G. Deciphering the Role of Multiple Thioredoxin Fold Proteins of Leptospirillum sp. in Oxidative Stress Tolerance. International Journal of Molecular Sciences. 2020; 21(5):1880. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21051880
Chicago/Turabian StyleGonzález, Daniela, Pamela Álamos, Matías Rivero, Omar Orellana, Javiera Norambuena, Renato Chávez, and Gloria Levicán. 2020. "Deciphering the Role of Multiple Thioredoxin Fold Proteins of Leptospirillum sp. in Oxidative Stress Tolerance" International Journal of Molecular Sciences 21, no. 5: 1880. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21051880