Claudin-12 Knockout Mice Demonstrate Reduced Proximal Tubule Calcium Permeability

 , , ,
, , ,
Abstract
:1. Introduction
2. Results
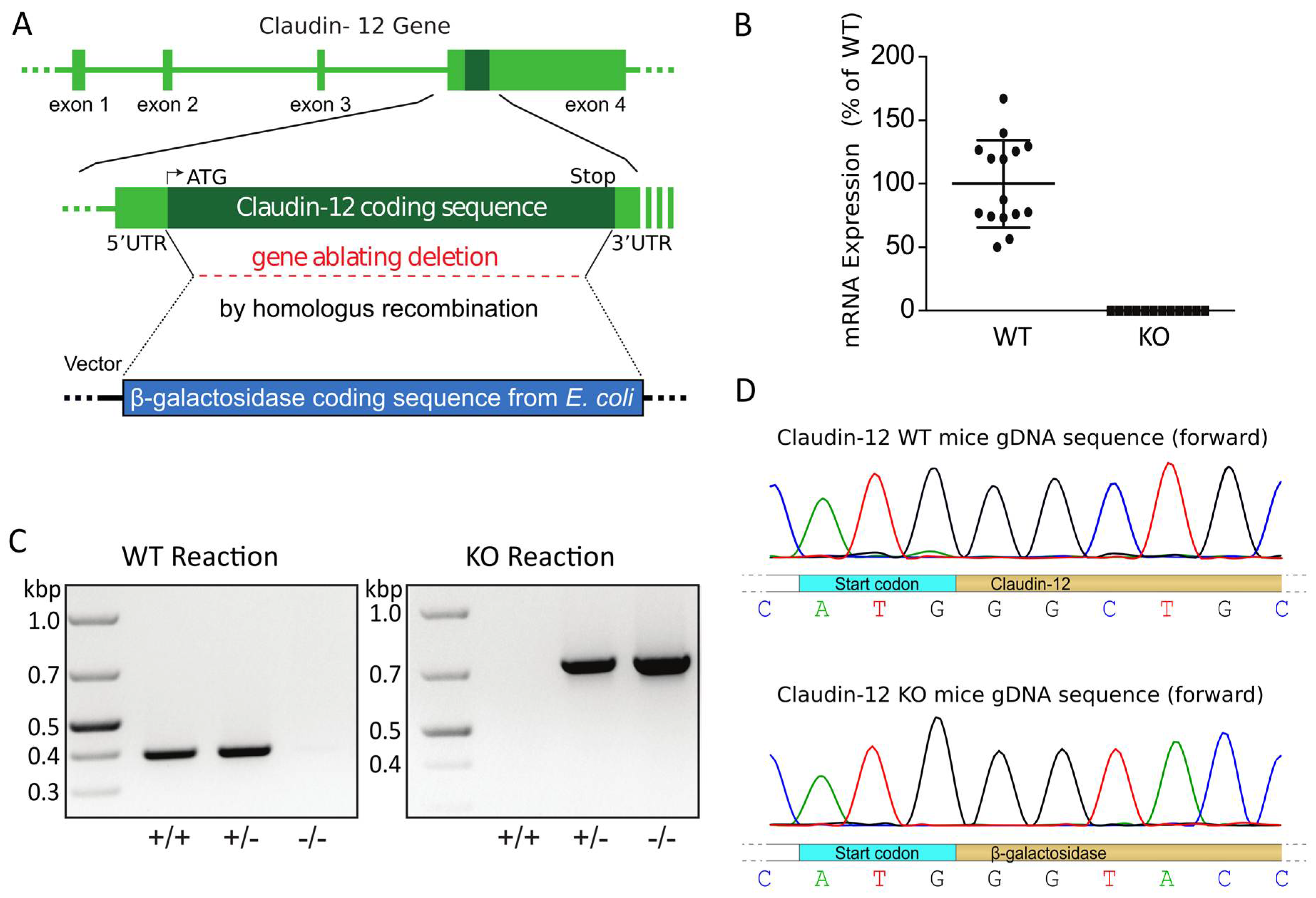
2.1. Generation of a Global Claudin-12 Knockout Model
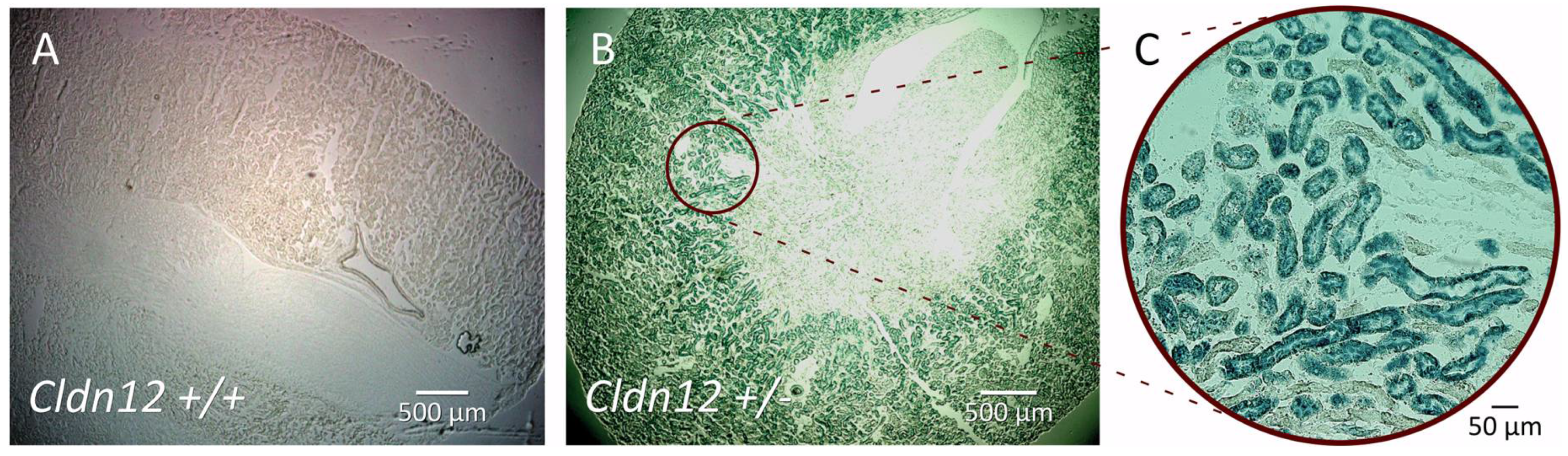
2.2. Claudin-12 is Expressed in the Renal Proximal Tubule
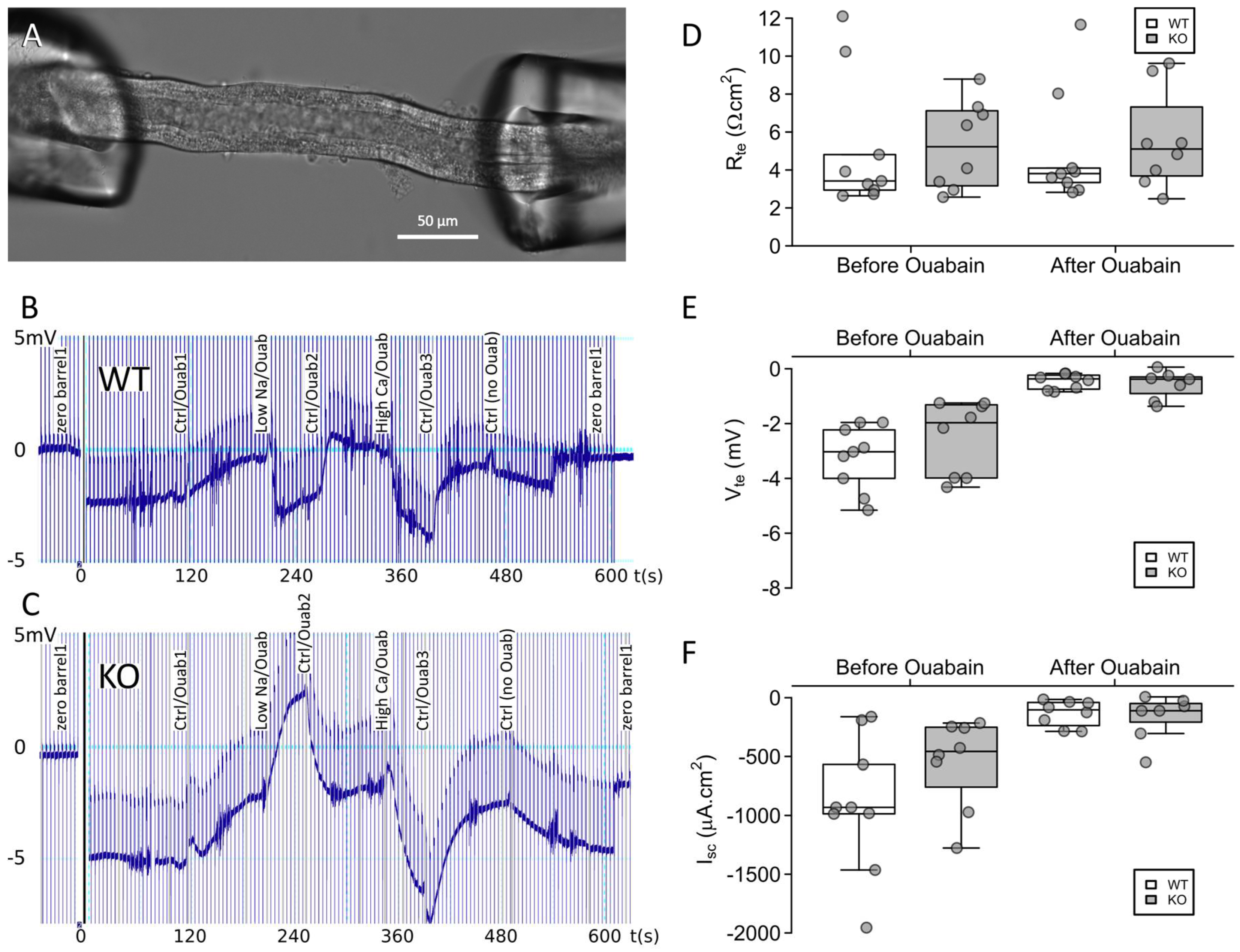
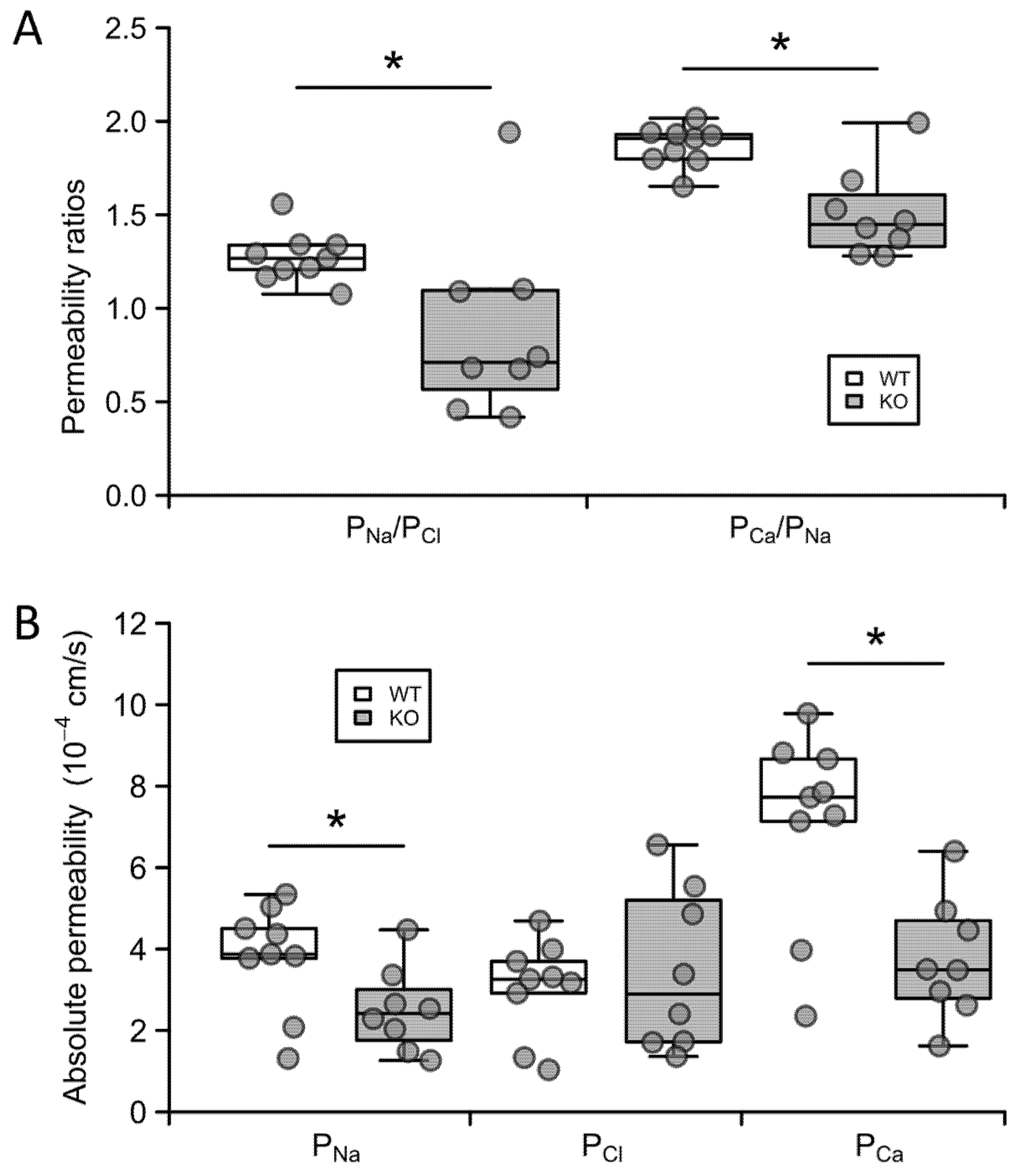
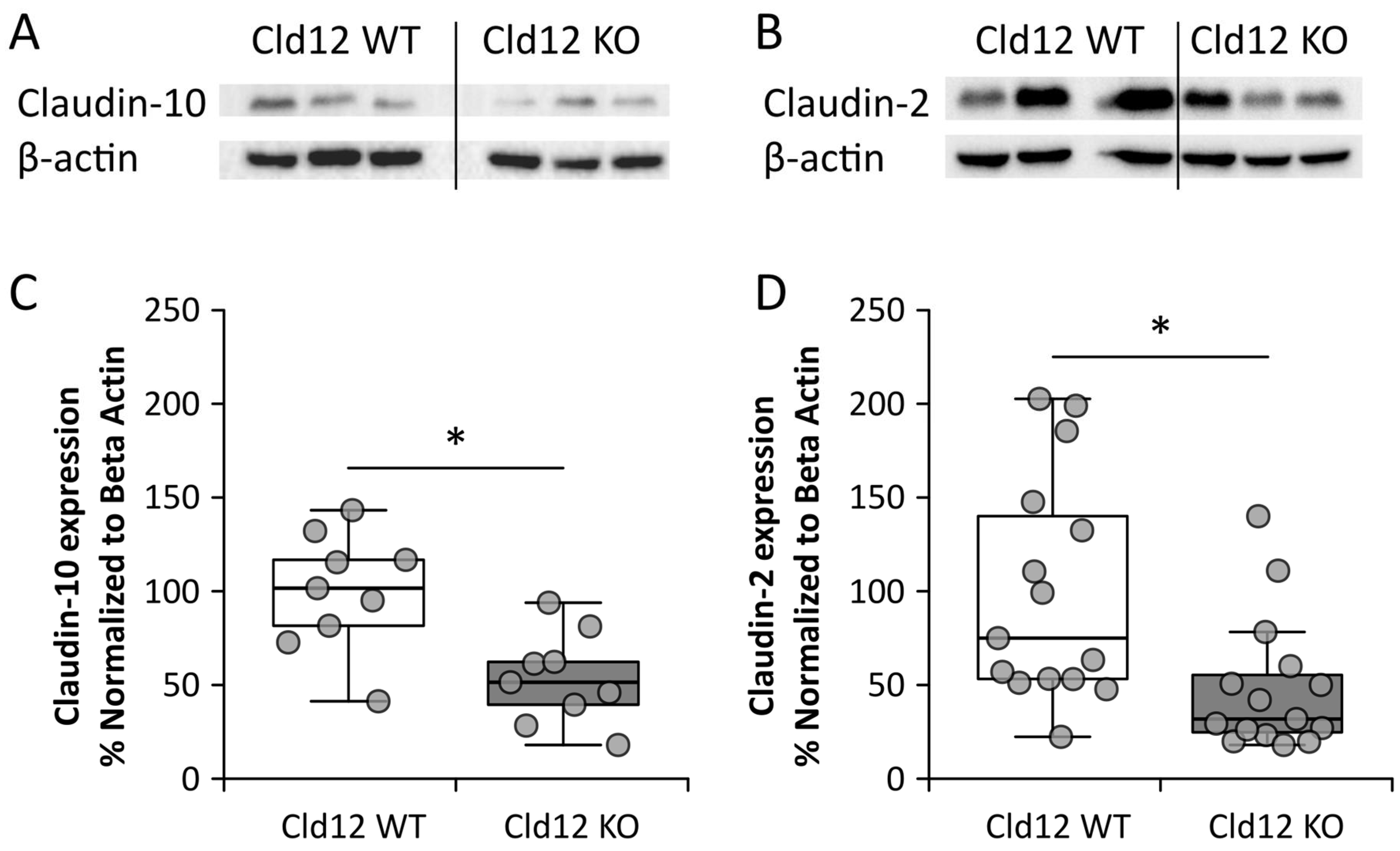
2.3. Claudin-12 Deletion Decreases Sodium and Calcium Permeability of the Proximal Tubule
2.4. Claudin-12 Null Mice Do not Have Hypercalciuria
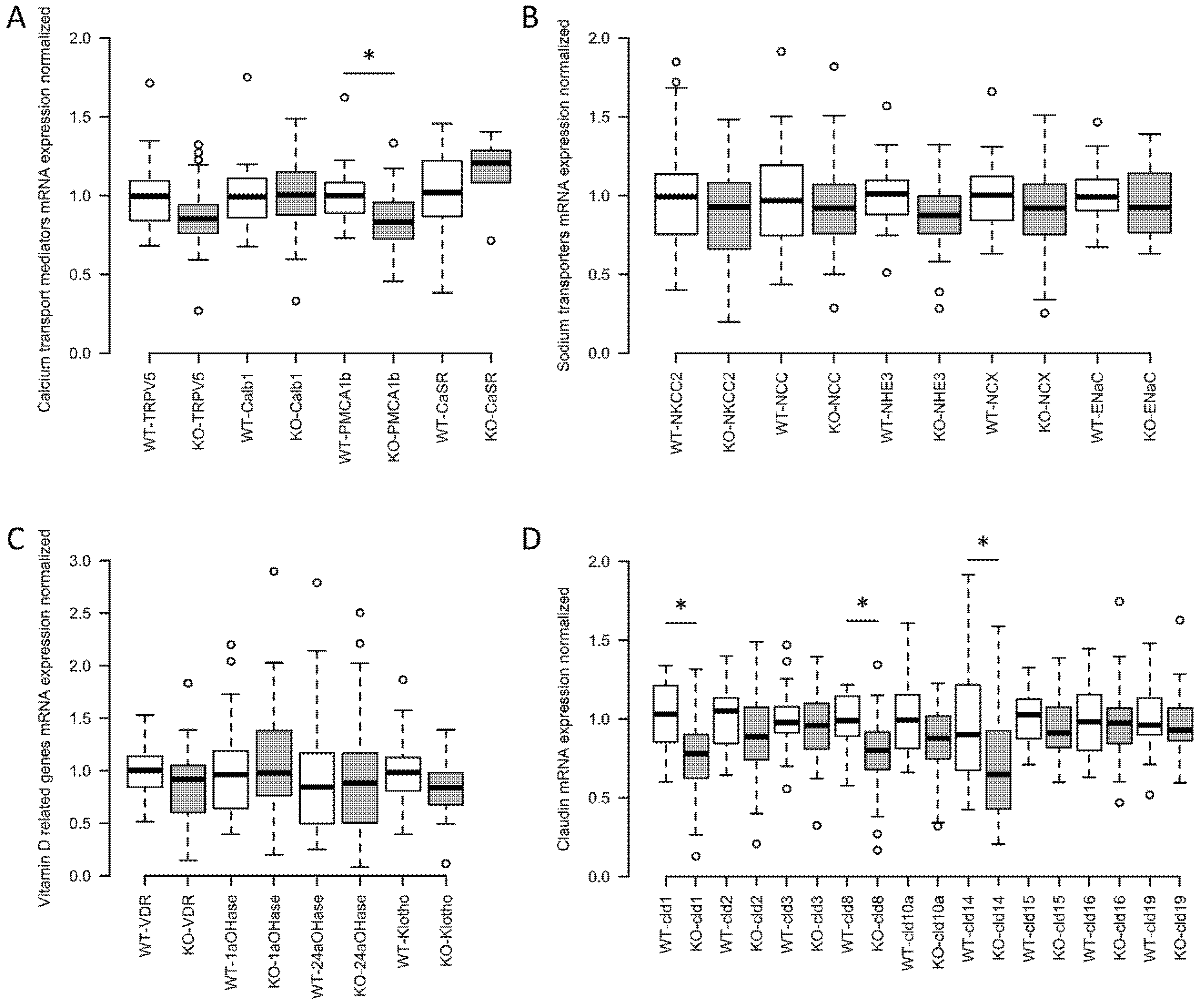
2.5. Renal Compensation in Claudin-12 KO Mice
3. Discussion
4. Materials and Methods
4.1. Generation of Cld12 KO Mice
4.2. Metabolic Cage Studies
4.3. Measurement of Urinary, Fecal, and Plasma Electrolytes
4.4. Measurement of Hormone Levels
4.5. X-Gal Staining and Immunofluorescence Microscopy on Renal Sections
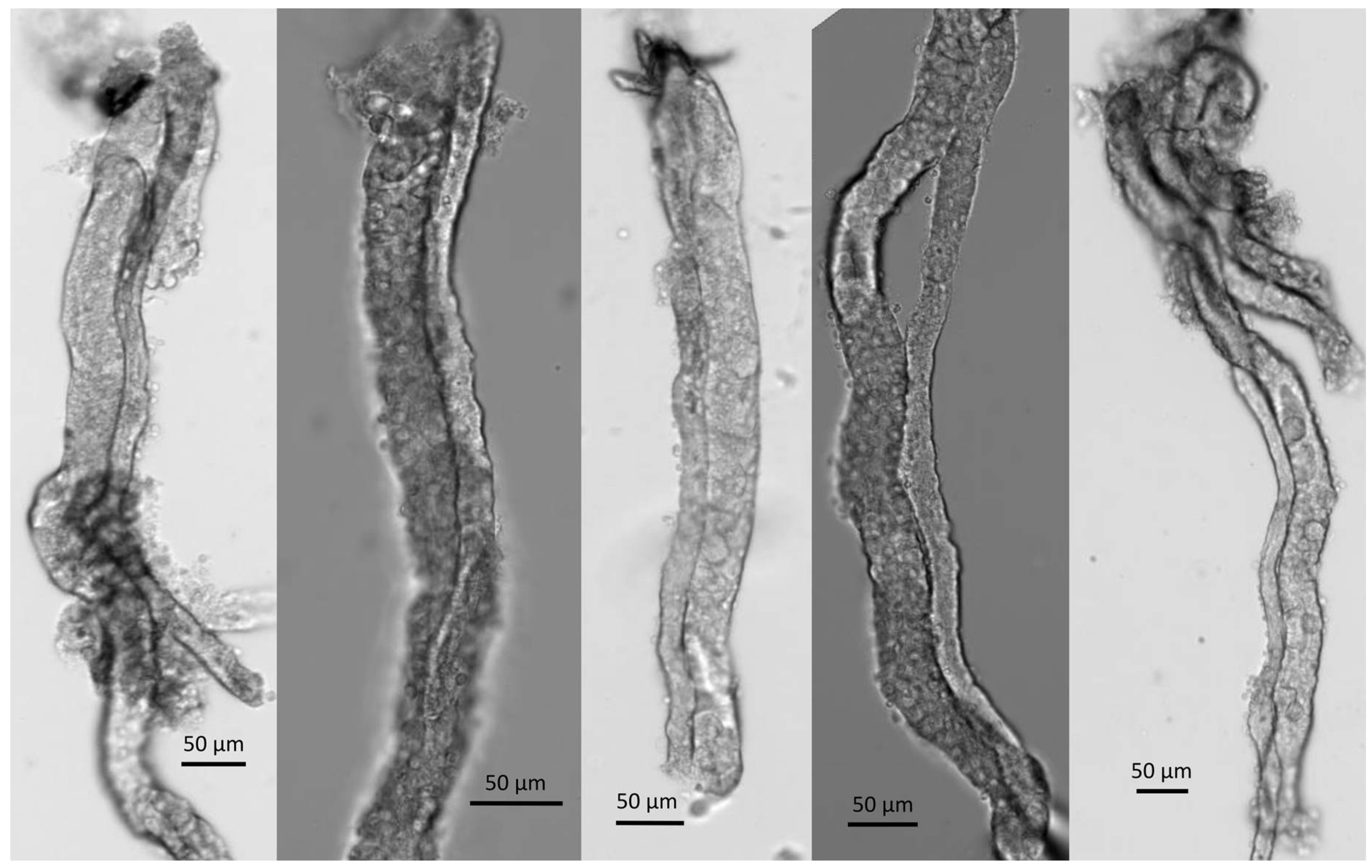
4.6. Proximal Tubular Perfusion and Electrophysiology
4.7. Quantitative Real-Time PCR and Immunoblotting
4.8. Statistical Analysis
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| PT | Renal proximal tubule |
| KO | Null (knock-out) mice |
| WT | Wild-type mice |
| PX | Paracellular permeability to ion X |
| (c)TAL | (cortical) Thick ascending limb of the Henle’s loop |
| PTH | Parathyroid hormone |
| FGF23 | Fibroblast growth factor 23 |
| ES | Embryonic stem cells |
| HET | Heterozygotes |
| AQP1 | Aquaporin I |
| NKCC2 | Sodium–potassium–chloride cotransporter II |
| NCC | Sodium–chloride cotransporter |
| CAII | Carbonic anhydrase II |
| BUN | Blood urea nitrogen |
| PMCA1b | Plasma membrane Ca2+ ATPase, isoform 1b |
| NHE3 | Sodium–hydrogen exchanger type 3 |
| CaSR | Calcium sensing receptor |
References
- Suki, W.N. Calcium transport in the nephron. Am. J. Physiol. 1979, 237, F1–F6. [Google Scholar] [CrossRef] [PubMed]
- Pan, W.; Borovac, J.; Spicer, Z.; Hoenderop, J.G.; Bindels, R.J.; Shull, G.E.; Doschak, M.R.; Cordat, E.; Alexander, R.T. The epithelial sodium/proton exchanger, NHE3, is necessary for renal and intestinal calcium (re)absorption. Am. J. Physiol. 2012, 302, F943–F956. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brunette, M.; Aras, M. A microinjection study of nephron permeability to calcium and magnesium. Am. J. Physiol. 1971, 221, 1442–1448. [Google Scholar] [CrossRef] [PubMed]
- Frick, A.; Rumrich, G.; Ullrich, K.J.; Lassiter, W.E. Microperfusion study of calcium transport in the proximal tubule of the rat kidney. Pflug. Arch. Gesamte Physiol. Menschen Tiere 1965, 286, 109–117. [Google Scholar] [CrossRef] [PubMed]
- Murayama, Y.; Morel, F.; Le Grimellec, C. Phosphate, calcium and magnesium transfers in proximal tubules and loops of Henle, as measured by single nephron microperfusion experiments in the rat. Pflug. Arch. 1972, 333, 1–16. [Google Scholar] [CrossRef]
- Edwards, A.; Bonny, O. A model of calcium transport and regulation in the proximal tubule. Am. J. Physiol. 2018, 315, F942–F953. [Google Scholar] [CrossRef]
- Worcester, E.M.; Coe, F.L.; Evan, A.P.; Bergsland, K.J.; Parks, J.H.; Willis, L.R.; Clark, D.L.; Gillen, D.L. Evidence for increased postprandial distal nephron calcium delivery in hypercalciuric stone-forming patients. Am. J. Physiol. 2008, 295, F1286–F1294. [Google Scholar] [CrossRef] [Green Version]
- Yu, A.S. Claudins and the kidney. J. Am. Soc. Nephrol. 2015, 26, 11–19. [Google Scholar] [CrossRef] [Green Version]
- Muto, S.; Hata, M.; Taniguchi, J.; Tsuruoka, S.; Moriwaki, K.; Saitou, M.; Furuse, K.; Sasaki, H.; Fujimura, A.; Imai, M.; et al. Claudin-2-deficient mice are defective in the leaky and cation-selective paracellular permeability properties of renal proximal tubules. Proc. Natl. Acad. Sci. USA 2010, 107, 8011–8016. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beggs, M.R.; Appel, I.; Svenningsen, P.; Skjodt, K.; Alexander, R.T.; Dimke, H. Expression of transcellular and paracellular calcium and magnesium transport proteins in renal and intestinal epithelia during lactation. Am. J. Physiol. 2017, 313, F629–F640. [Google Scholar] [CrossRef] [Green Version]
- Fujita, H.; Sugimoto, K.; Inatomi, S.; Maeda, T.; Osanai, M.; Uchiyama, Y.; Yamamoto, Y.; Wada, T.; Kojima, T.; Yokozaki, H.; et al. Tight junction proteins claudin-2 and -12 are critical for vitamin D-dependent Ca2+ absorption between enterocytes. Mol. Biol. Cell 2008, 19, 1912–1921. [Google Scholar] [CrossRef] [Green Version]
- Kiuchi-Saishin, Y.; Gotoh, S.; Furuse, M.; Takasuga, A.; Tano, Y.; Tsukita, S. Differential expression patterns of claudins, tight junction membrane proteins, in mouse nephron segments. J. Am. Soc. Nephrol. 2002, 13, 875–886. [Google Scholar] [PubMed]
- Borovac, J.; Barker, R.S.; Rievaj, J.; Rasmussen, A.; Pan, W.; Wevrick, R.; Alexander, R.T. Claudin-4 forms a paracellular barrier, revealing the interdependence of claudin expression in the loose epithelial cell culture model opossum kidney cells. Am. J. Physiol. Cell Physiol. 2012, 303, C1278–C1291. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abuazza, G.; Becker, A.; Williams, S.S.; Chakravarty, S.; Truong, H.T.; Lin, F.; Baum, M. Claudins 6, 9 and 13 are developmentally expressed renal tight junction proteins. Am. J. Physiol. Renal Physiol. 2006, 291, F1132–F1141. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dimke, H.; Desai, P.; Borovac, J.; Lau, A.; Pan, W.; Alexander, R.T. Activation of the Ca2+-sensing receptor increases renal claudin-14 expression and urinary Ca2+ excretion. Am. J. Physiol. Renal Physiol. 2013, 304, F761–F769. [Google Scholar] [CrossRef] [Green Version]
- Krishnan, D.; Pan, W.; Beggs, M.R.; Trepiccione, F.; Chambrey, R.; Eladari, D.; Cordat, E.; Dimke, H.; Alexander, R.T. Deficiency of Carbonic Anhydrase II Results in a Urinary Concentrating Defect. Front. Physiol. 2017, 8, 1108. [Google Scholar] [CrossRef] [Green Version]
- Castro Dias, M.; Coisne, C.; Baden, P.; Enzmann, G.; Garrett, L.; Becker, L.; Holter, S.M.; German Mouse Clinic, C.; Hrabe de Angelis, M.; Deutsch, U.; et al. Claudin-12 is not required for blood-brain barrier tight junction function. Fluids Barriers CNS 2019, 16, 30. [Google Scholar] [CrossRef]
- Sabolic, I.; Valenti, G.; Verbavatz, J.M.; Van Hoek, A.N.; Verkman, A.S.; Ausiello, D.A.; Brown, D. Localization of the CHIP28 water channel in rat kidney. Am. J. Physiol. 1992, 263, C1225–C1233. [Google Scholar] [CrossRef]
- Vilas, G.; Krishnan, D.; Loganathan, S.K.; Malhotra, D.; Liu, L.; Beggs, M.R.; Gena, P.; Calamita, G.; Jung, M.; Zimmermann, R.; et al. Increased water flux induced by an aquaporin-1/carbonic anhydrase II interaction. Mol. Biol. Cell 2015, 26, 1106–1118. [Google Scholar] [CrossRef]
- Gong, Y.; Renigunta, V.; Himmerkus, N.; Zhang, J.; Renigunta, A.; Bleich, M.; Hou, J. Claudin-14 regulates renal Ca++ transport in response to CaSR signalling via a novel microRNA pathway. EMBO J. 2012, 31, 1999–2012. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rievaj, J.; Pan, W.; Cordat, E.; Alexander, R.T. The Na+/H+ exchanger isoform 3 is required for active paracellular and transcellular Ca2+ transport across murine cecum. Am. J. Physiol. Gastrointest. Liver Physiol. 2013, 305, G303–G313. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alexander, R.T.; Dimke, H.; Cordat, E. Proximal tubular NHEs: Sodium, protons and calcium? Am. J. Physiol. Renal Physiol. 2013, 305, F229–F236. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schultheis, P.J.; Clarke, L.L.; Meneton, P.; Miller, M.L.; Soleimani, M.; Gawenis, L.R.; Riddle, T.M.; Duffy, J.J.; Doetschman, T.; Wang, T.; et al. Renal and intestinal absorptive defects in mice lacking the NHE3 Na+/H+ exchanger. Nat. Genet. 1998, 19, 282–285. [Google Scholar] [CrossRef] [PubMed]
- Leong, P.K.; Devillez, A.; Sandberg, M.B.; Yang, L.E.; Yip, D.K.; Klein, J.B.; McDonough, A.A. Effects of ACE inhibition on proximal tubule sodium transport. Am. J. Physiol. Renal Physiol. 2006, 290, F854–F863. [Google Scholar] [CrossRef] [Green Version]
- Riquier-Brison, A.D.; Leong, P.K.; Pihakaski-Maunsbach, K.; McDonough, A.A. Angiotensin II stimulates trafficking of NHE3, NaPi2, and associated proteins into the proximal tubule microvilli. Am. J. Physiol. Renal Physiol. 2010, 298, F177–F186. [Google Scholar] [CrossRef] [Green Version]
- Plain, A.; Wulfmeyer, V.C.; Milatz, S.; Klietz, A.; Hou, J.; Bleich, M.; Himmerkus, N. Corticomedullary difference in the effects of dietary Ca2+ on tight junction properties in thick ascending limbs of Henle’s loop. Pflug. Arch. 2016, 468, 293–303. [Google Scholar] [CrossRef]
- Enck, A.H.; Berger, U.V.; Yu, A.S. Claudin-2 is selectively expressed in proximal nephron in mouse kidney. Am. J. Physiol. Renal Physiol. 2001, 281, F966–F974. [Google Scholar] [CrossRef]
- Pei, L.; Solis, G.; Nguyen, M.T.; Kamat, N.; Magenheimer, L.; Zhuo, M.; Li, J.; Curry, J.; McDonough, A.A.; Fields, T.A.; et al. Paracellular epithelial sodium transport maximizes energy efficiency in the kidney. J. Clin. Investig. 2016, 126, 2509–2518. [Google Scholar] [CrossRef] [Green Version]
- Greger, R.; Lang, F.; Oberleithner, H. Distal site of calcium reabsorption in the rat nephron. Pflug. Arch. 1978, 374, 153–157. [Google Scholar] [CrossRef]
- Gattineni, J.; Bates, C.; Twombley, K.; Dwarakanath, V.; Robinson, M.L.; Goetz, R.; Mohammadi, M.; Baum, M. FGF23 decreases renal NaPi-2a and NaPi-2c expression and induces hypophosphatemia in vivo predominantly via FGF receptor 1. Am. J. Physiol. Renal Physiol. 2009, 297, F282–F291. [Google Scholar] [CrossRef] [Green Version]
- Olauson, H.; Lindberg, K.; Amin, R.; Jia, T.; Wernerson, A.; Andersson, G.; Larsson, T.E. Targeted deletion of Klotho in kidney distal tubule disrupts mineral metabolism. J. Am. Soc. Nephrol. 2012, 23, 1641–1651. [Google Scholar] [CrossRef] [Green Version]
- Ure, M.E.; Heydari, E.; Pan, W.; Ramesh, A.; Rehman, S.; Morgan, C.; Pinsk, M.; Erickson, R.; Herrmann, J.M.; Dimke, H.; et al. A variant in a cis-regulatory element enhances claudin-14 expression and is associated with pediatric-onset hypercalciuria and kidney stones. Hum. Mutat. 2017, 38, 649–657. [Google Scholar] [CrossRef] [PubMed]
- Wiebe, S.A.; Plain, A.; Pan, W.; O’Neill, D.; Braam, B.; Alexander, R.T. NHE8 attenuates Ca2+ influx into NRK cells and the proximal tubule epithelium. Am. J. Physiol. Renal Physiol. 2019, 317, F240–F253. [Google Scholar] [CrossRef] [PubMed]
- Greger, R. Cation selectivity of the isolated perfused cortical thick ascending limb of Henle’s loop of rabbit kidney. Pflug. Arch. 1981, 390, 30–37. [Google Scholar] [CrossRef] [PubMed]
- Gunzel, D.; Stuiver, M.; Kausalya, P.J.; Haisch, L.; Krug, S.M.; Rosenthal, R.; Meij, I.C.; Hunziker, W.; Fromm, M.; Muller, D. Claudin-10 exists in six alternatively spliced isoforms that exhibit distinct localization and function. J. Cell Sci. 2009, 122, 1507–1517. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kimizuka, H.; Koketsu, K. Ion transport through cell membrane. J. Theor. Biol. 1964, 6, 290–305. [Google Scholar] [CrossRef]
- Thissen, D.; Steinberg, L.; Kuang, D. Quick and easy implementation of the Benjamini-Hochberg procedure for controlling the false positive rate in multiple comparisons. J. Educ. Behav. Stat. 2002, 27, 77–83. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| WT (n = 12–13) | KO (n = 17–19) | p-Value | |
|---|---|---|---|
| Female Weight, g | 26.8 ± 1.3 (n = 10) | 26.6 ± 0.7 (n = 9) | 0.918 |
| Male Weight, g | 29.4 ± 0.9 (n = 3) | 30.9 ± 0.8 (n = 10) | 0.337 |
| H2O drunk, mL/24 h | 6.0 ± 0.4 | 6.3 ± 0.3 | 0.426 |
| Chow eaten, g | 4.8 ± 0.2 | 5.3 ± 0.2 | 0.068 |
| Ca2+ingested 1, µmol/24 h | 1138 ± 46 | 1266 ± 46 | 0.068 |
| Urine volume, mL/24 h | 1.6 ± 0.2 | 1.7 ± 0.2 | 0.603 |
| Fecal excretion 2, g/24 h | 5.8 ± 0.3 | 6.4 ± 0.3 | 0.162 |
| Fecal calcium excretion | |||
| Ca2+feces, µmol/24 h | 1065 ± 53 | 1116 ± 57 | 0.536 |
| Ca2+feces/Ca2+ingested | 0.94 ± 0.04 | 0.88 ± 0.03 | 0.177 |
| WT (n = 12–13) | KO (n = 17–19) | p-Value | |
|---|---|---|---|
| Na+, mmol/L | 150.7 ± 1.2 | 150.9 ± 0.6 | 0.820 |
| K+, mmol/L | 4.7 ± 0.2 | 4.6 ± 0.2 | 0.784 |
| Cl−, mmol/L 1 | 119 ± 1 | 117 ± 0.7 | 0.180 |
| Ca2+, mmol/L | 2.4 ± 0.2 | 2.5 ± 0.1 | 0.369 |
| Cr, µmol/L | 0.030 ± 0.004 | 0.036 ± 0.004 | 0.315 |
| Glucose, mmol/L | 8.8 ± 0.5 | 9.3 ± 0.4 | 0.389 |
| BUN, mmol/L 1 | 29.2 ± 1.5 | 29.3 ± 1.5 | 0.954 |
| PTH, pg/mL | 421 ± 85 | 332 ± 52 | 0.370 |
| FGF23, pg/mL | 273 ± 16 | 306 ± 14 | 0.173 |
| Vitamin D3, pg/mL | 81 ± 15 | 80 ± 11 | 0.971 |
| WT (n = 12) | KO (n = 13) | p-Value | |
|---|---|---|---|
| Na+/Creatinine | 31 ± 8 | 39 ± 9 | 0.495 |
| Cl−/Creatinine | 140 ± 45 | 136 ± 33 | 0.951 |
| K+/Creatinine | 99 ± 25 | 93 ± 21 | 0.855 |
| Ca2+/Creatinine | 0.24 ± 0.05 | 0.22 ± 0.04 | 0.819 |
| PO43-/Creatinine | 5.4 ± 0.8 | 5.0 ± 0.7 | 0.688 |
| Creatinine, µmol/24 h | 12 ± 2 | 16 ± 3 | 0.377 |
| Normal | High Ca | Low Ca | |||||||
|---|---|---|---|---|---|---|---|---|---|
| WT (n = 12) | KO (n = 8) | p-Value | WT (n = 14) | KO (n = 15) | p-Value | WT (n = 15) | KO (n = 11) | p-Value | |
| Urine Ca2 +/Cr | 0.88 ± 0.08 | 0.79 ± 0.07 | 0.456 | 2.65 ± 0.48 | 3.10 ± 0.52 | 0.555 | 0.62 ± 0.03 | 0.60 ± 0.01 | 0.614 |
| FE Ca | 0.63 ± 0.15 | 0.37 ± 0.21 | 0.323 | 1.88 ± 0.46 | 1.67 ± 0.31 | 0.703 | 0.55 ± 0.14 | 0.31 ± 0.07 | 0.178 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Plain, A.; Pan, W.; O’Neill, D.; Ure, M.; Beggs, M.R.; Farhan, M.; Dimke, H.; Cordat, E.; Alexander, R.T. Claudin-12 Knockout Mice Demonstrate Reduced Proximal Tubule Calcium Permeability. Int. J. Mol. Sci. 2020, 21, 2074. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21062074
Plain A, Pan W, O’Neill D, Ure M, Beggs MR, Farhan M, Dimke H, Cordat E, Alexander RT. Claudin-12 Knockout Mice Demonstrate Reduced Proximal Tubule Calcium Permeability. International Journal of Molecular Sciences. 2020; 21(6):2074. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21062074
Chicago/Turabian StylePlain, Allein, Wanling Pan, Deborah O’Neill, Megan Ure, Megan R. Beggs, Maikel Farhan, Henrik Dimke, Emmanuelle Cordat, and R. Todd Alexander. 2020. "Claudin-12 Knockout Mice Demonstrate Reduced Proximal Tubule Calcium Permeability" International Journal of Molecular Sciences 21, no. 6: 2074. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21062074