Knockdown of Musashi RNA Binding Proteins Decreases Radioresistance but Enhances Cell Motility and Invasion in Triple-Negative Breast Cancer

 ,
,  , ,
, ,  and
and
Abstract
:1. Introduction
2. Results
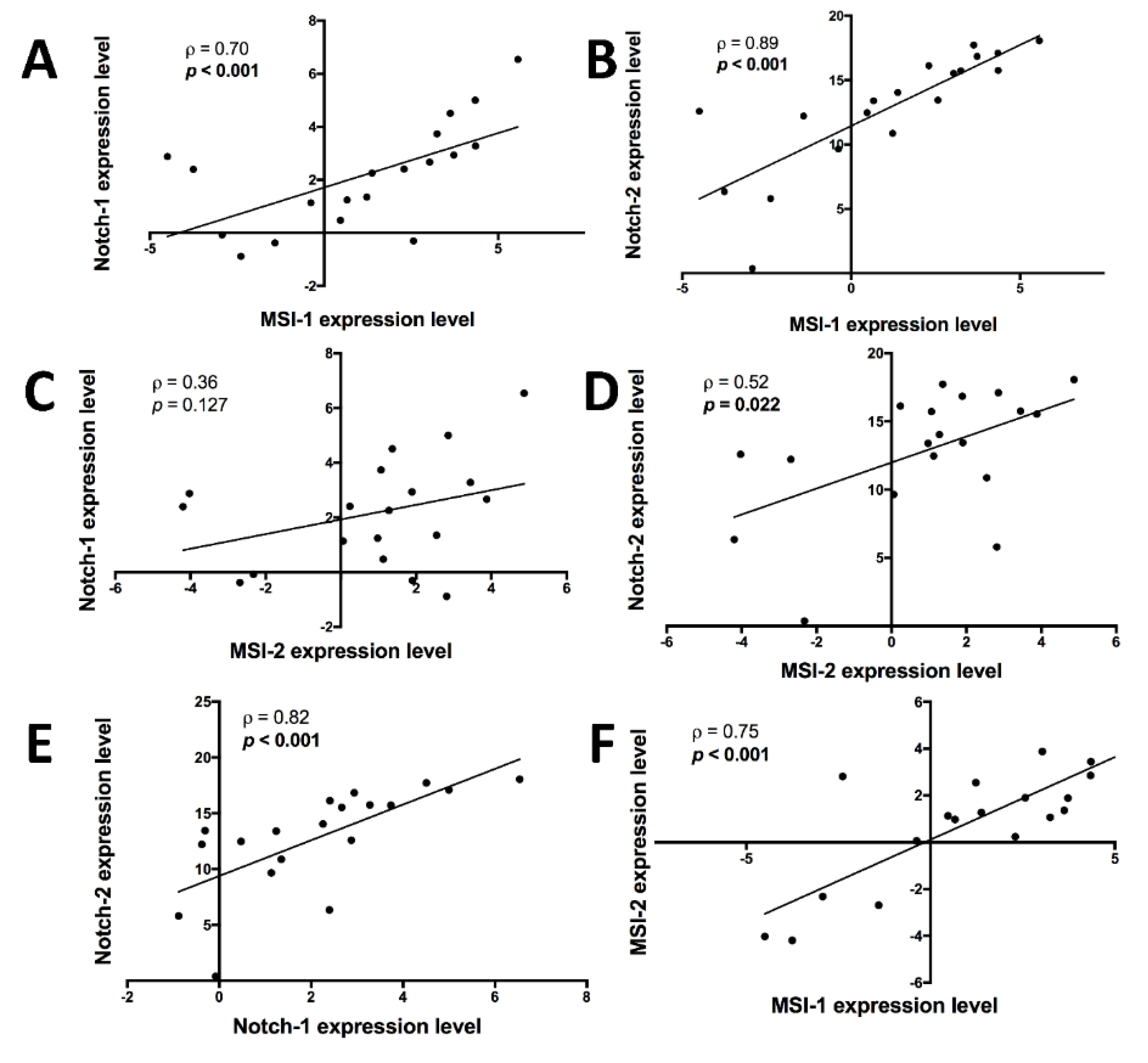
2.1. MSI Protein mRNAs Show Strong Correlations with Each Other and Notch Pathway Elements in Triple-Negative Breast Cancer Samples
2.2. MSI-1 and MSI-2 Small Interfering RNA (siRNA) Transfection Results in MSI-1 and MSI-2 Knockdown
2.3. Knockdown of MSI-1 and MSI-2 Suppresses the Notch Pathway
2.4. Breast Cancer Stem Cell Markers Are Downregulated after MSI Protein Knockdown
2.5. Decreased MSI Protein Expression Results in Increased Apoptosis and Reduced Proliferation
2.6. Low MSI Expression Leads to Decreased Radioresistance and Reduced Expression of DNA Repair-Related Proteins DNA-PKcs and EGFR
2.7. MSI-Downregulated Cells Exhibit Increased Cell Motility and Invasion
3. Discussion
3.1. MSI Protein Knockdown Critically Downregulates Stem Cell Characteristics and Cell Cycle Progression While Increasing Apoptosis Subsequent to Notch Pathway Inactivation
- We show that CD44 is downregulated via MSI knockdown in breast cancer. CD44 is a key stem cell marker in mammary malignancies and it has been shown that as few as 100 CD44(high) cells may promote tumorigenesis in breast cancer [25]. Our findings are in line with studies in colon cancer indicating a positive relationship between MSI proteins and CD44 [33,34].
- The mesenchymal protein vimentin has been described as a stem cell regulator in mouse models that leads to reduced regenerative capacity and is associated with tumor sphere formation [27].
- The cell cycle analysis findings were fairly similar to previous quantifications published by our group in endometrial carcinoma [14]. In sum, we see a strong increase in G1 phase cells at the expense of S and no substantial change in G2/M phase cell proportions after 72 hours, suggesting an anti-proliferative effect. This effect seems to slowly develop between 24 and 72 hours with its peak after 72 hours.
- The apoptosis assay demonstrated that the antiproliferative effect also seems to carry pro-apoptotic properties: Knockdown cells were significantly more likely to bind annexin V, indicating apoptotic features. Again, this is well in line with a previous study in endometrial cancer [14].
3.2. MSI Protein Knockdown Reduces Breast Cancer Radioresistance via Downregulation of EGFR and DNA-PKcs
- First and most importantly, the Notch pathway has been described to confer radioresistance through enhancing cancer stem cell properties [36,37]. Conversely, Notch signaling is upregulated after irradiation [38]. We have demonstrated a correlation between MSI expression and Notch elements in primary breast tissues as well as a downregulation of Notch signaling after MSI knockdown. Decreased Notch activity may thus confer reduced radioresistant properties.
- In glioblastoma, de Araujo and colleagues demonstrated a decrease in DNA-PKcs expression [21]. In our study, we demonstrate the same effect for breast cancer. A decrease in this key DNA repair protein is known to sensitize breast cancer to radiation [43] and the protein has been suggested as a therapeutic target [44]. Interestingly, the decrease in DNA-PKcs may also explain why we were unable to see changes in double strand breaks via γH2AX assay. The γH2AX assay does not directly measure double strand breaks, but rather the (related) γ-phosphorylation of histones. However, this process is mediated by DNA-PKcs as aptly summarized by An et al.: “DNA-PKcs plays a dominant role in the regulation of H2AX phosphorylation in response to DNA damage and cell cycle progression” [45]. Thus, with potentially more DNA damage, but less DNA-PKcs to indicate damage via histone phosphorylation, we believe it is ultimately understandable why no overall change was seen in the γH2AX assay.
- Lastly, EGFR is also well known for its mediation of radioresistant properties with new clinical trials to target its effect underway [29,46]. MSI knockdown downregulates EGFR, thus providing another potential mechanism to enhance radiosensitivity. EGFR also plays a key role for BCSC activity [47], demonstrating yet again that many of the aforementioned mechanisms are closely related.
3.3. MSI Protein Knockdown Results in a Higher Cell Invasiveness and More Migration In Vivo, Possibly due to Downregulation of the LIF Receptor
4. Materials and Methods
4.1. Cell Line and Transfection
4.2. qPCR
4.3. Western Blot Analysis
4.4. Flow Cytometry
4.5. Cell Colony Formation
4.6. Cell Cycle Progression
4.7. γH2AX
4.8. Apoptosis
4.9. Invasion
4.10. Digital Holographic Microscopy
4.11. Primary Tissue
4.12. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| ALDH | Aldehyde dehydrogenase |
| BCSC | Breast cancer stem cell |
| Ctrl | Control |
| DAPI | 4’,6-diamidino-2-phenylindole |
| DNA-PKcs | DNA protein kinase catalytic subunit |
| EGFR | Epithelial growth factor receptor |
| EMT | Epithelial mesenchymal transition |
| FACS | Fluorescence-activated cell sorting |
| FCS | Fetal Calf Serum |
| LIFR | Leukemia inhibitory factor receptor |
| MDR | Multidrug resistance |
| Mnumb | Mammalian numb |
| MSI | Musashi (RNA-binding proteins) |
| MSI-1 | Musashi RNA-binding protein 1 |
| MSI-2 | Musashi RNA-binding protein 2 |
| OS | Overall survival |
| PE | Phycoerythrin |
| PEf | Plating efficacy |
| PI | Propidium iodide |
| qPCR | Quantitative polymerase chain reaction |
| s.e.m. | Standard error of the mean |
| SF | Surviving fraction |
| siRNA | Small interfering RNA |
| STR | Short tandem repeat |
| TBST | TBS-Tween |
| YAP | YES-associated protein |
References
- Al-Hajj, M.; Wicha, M.S.; Benito-Hernandez, A.; Morrison, S.J.; Clarke, M.F. Prospective identification of tumorigenic breast cancer cells. Proc. Natl. Acad. Sci. USA 2003, 100, 3983–3988. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zucchi, I.; Sanzone, S.; Astigiano, S.; Pelucchi, P.; Scotti, M.; Valsecchi, V.; Barbieri, O.; Bertoli, G.; Albertini, A.; Reinbold, R.A.; et al. The properties of a mammary gland cancer stem cell. Proc. Natl. Acad. Sci. USA 2007, 104, 10476–10481. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smalley, M.; Piggott, L.; Clarkson, R. Breast cancer stem cells: Obstacles to therapy. Cancer Lett. 2013, 338, 57–62. [Google Scholar] [CrossRef] [PubMed]
- Shao, J.; Fan, W.; Ma, B.; Wu, Y. Breast cancer stem cells expressing different stem cell markers exhibit distinct biological characteristics. Mol. Med. Rep. 2016, 14, 4991–4998. [Google Scholar] [CrossRef] [Green Version]
- Greve, B.; Kelsch, R.; Spaniol, K.; Eich, H.T.; Götte, M. Flow cytometry in cancer stem cell analysis and separation. Cytom. Part A 2012, 81, 284–293. [Google Scholar] [CrossRef]
- Ibrahim, S.A.; Gadalla, R.; El-Ghonaimy, E.A.; Samir, O.; Mohamed, H.T.; Hassan, H.; Greve, B.; El-Shinawi, M.; Mohamed, M.M.; Götte, M. Syndecan-1 is a novel molecular marker for triple negative inflammatory breast cancer and modulates the cancer stem cell phenotype via the IL-6/STAT3, Notch and EGFR signaling pathways. Mol. Cancer 2017, 16, 1–19. [Google Scholar] [CrossRef] [Green Version]
- Troschel, F.M.; Böhly, N.; Borrmann, K.; Braun, T.; Schwickert, A.; Kiesel, L.; Eich, H.T.; Götte, M.; Greve, B. miR-142-3p attenuates breast cancer stem cell characteristics and decreases radioresistance in vitro. Tumor Biol. 2018, 40, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Okano, H.; Imai, T.; Okabe, M. Musashi: A translational regulator of cell fate. J. Cell Sci. 2002, 115, 1355–1359. [Google Scholar]
- Okano, H.; Kawahara, H.; Toriya, M.; Nakao, K.; Shibata, S.; Imai, T. Function of RNA-binding protein Musashi-1 in stem cells. Exp. Cell Res. 2005, 306, 349–356. [Google Scholar] [CrossRef]
- Sakakibara, S.I.; Nakamura, Y.; Satoh, H.; Okano, H. RNA-binding protein Musashi2: Developmentally regulated expression in neural precursor cells and subpopulations of neurons in mammalian CNS. J. Neurosci. 2001, 21, 8091–8107. [Google Scholar] [CrossRef] [Green Version]
- Sakakibara, S.-I.; Nakamura, Y.; Yoshida, T.; Shibata, S.; Koike, M.; Takano, H.; Ueda, S.; Uchiyama, Y.; Noda, T.; Okano, H. RNA-binding protein Musashi family: Roles for CNS stem cells and a subpopulation of ependymal cells revealed by targeted disruption and antisense ablation. Proc. Natl. Acad. Sci. USA 2002, 99, 15194–15199. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fox, R.G.; Park, F.D.; Koechlein, C.S.; Kritzik, M.; Reya, T. Musashi signaling in stem cells and cancer. Ann. Rev. Cell Dev. Biol. 2015, 31, 249–267. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, P.X.; Li, Q.Y.; Yang, Z. Musashi-1 Expression is a Prognostic Factor in Ovarian Adenocarcinoma and Correlates with ALDH-1 Expression. Pathol. Oncol. Res. 2015, 21, 1133–1140. [Google Scholar] [CrossRef] [PubMed]
- Götte, M.; Greve, B.; Kelsch, R.; Müller-Uthoff, H.; Weiss, K.; Kharabi Masouleh, B.; Sibrowski, W.; Kiesel, L.; Buchweitz, O. The adult stem cell marker Musashi-1 modulates endometrial carcinoma cell cycle progression and apoptosis via Notch-1 and p21WAF1/CIP1. Int. J. Cancer 2011, 129, 2042–2049. [Google Scholar] [CrossRef]
- Götte, M.; Wolf, M.; Staebler, A.; Buchweitz, O.; Kelsch, R.; Schüring, A.N.; Kiesel, L. Increased expression of the adult stem cell marker Musashi-1 in endometriosis and endometrial carcinoma. J. Pathol. 2008, 215, 317–329. [Google Scholar] [CrossRef]
- Lagadec, C.; Vlashi, E.; Frohnen, P.; Alhiyari, Y.; Chan, M.; Pajonk, F. The RNA-binding protein musashi-1 regulates proteasome subunit expression in breast cancerand glioma-initiating cells. Stem Cells 2014, 32, 135–144. [Google Scholar] [CrossRef] [Green Version]
- Choi, Y.M.; Kim, K.B.; Lee, J.H.; Chun, Y.K.; An, I.S.; An, S.; Bae, S. DBC2/RhoBTB2 functions as a tumor suppressor protein via Musashi-2 ubiquitination in breast cancer. Oncogene 2017, 36, 2802–2812. [Google Scholar] [CrossRef]
- Katz, Y.; Li, F.; Lambert, N.J.; Sokol, E.S.; Tam, W.L.; Cheng, A.W.; Airoldi, E.M.; Lengner, C.J.; Gupta, P.B.; Yu, Z.; et al. Musashi proteins are post-transcriptional regulators of the epithelial-luminal cell state. Elife 2014, 3, 1–27. [Google Scholar] [CrossRef] [Green Version]
- Kang, M.H.; Jeong, K.J.; Kim, W.Y.; Lee, H.J.; Gong, G.; Suh, N.; Gyrffy, B.; Kim, S.; Jeong, S.Y.; Mills, G.B.; et al. Musashi RNA-binding protein 2 regulates estrogen receptor 1 function in breast cancer. Oncogene 2017, 36, 1745–1752. [Google Scholar] [CrossRef]
- Lin, J.C.; Tsai, J.T.; Chao, T.Y.; Ma, H.I.; Chien, C.S.; Liu, W.H. MSI1 associates glioblastoma radioresistance via homologous recombination repair, tumor invasion and cancer stem-like cell properties. Radiother. Oncol. 2018, 129, 352–363. [Google Scholar] [CrossRef]
- de Araujo, P.R.; Gorthi, A.; da Silva, A.E.; Tonapi, S.S.; Vo, D.T.; Burns, S.C.; Qiao, M.; Uren, P.J.; Yuan, Z.M.; Bishop, A.J.R.; et al. Musashi1 Impacts Radio-Resistance in Glioblastoma by Controlling DNA-Protein Kinase Catalytic Subunit. Am. J. Pathol. 2016, 186, 2271–2278. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sureban, S.M.; May, R.; George, R.J.; Dieckgraefe, B.K.; McLeod, H.L.; Ramalingam, S.; Bishnupuri, K.S.; Natarajan, G.; Anant, S.; Houchen, C.W. Knockdown of RNA Binding Protein Musashi-1 Leads to Tumor Regression In Vivo. Gastroenterology 2008, 134, 1448–1458. [Google Scholar] [CrossRef] [PubMed]
- Kudinov, A.E.; Karanicolas, J.; Golemis, E.A.; Boumber, Y. Musashi RNA-Binding Proteins as Cancer Drivers and Novel Therapeutic Targets. Clin. Cancer Res. 2017, 23, 2143–2153. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bouras, T.; Pal, B.; Vaillant, F.; Harburg, G.; Asselin-Labat, M.L.; Oakes, S.R.; Lindeman, G.J.; Visvader, J.E. Notch Signaling Regulates Mammary Stem Cell Function and Luminal Cell-Fate Commitment. Cell Stem Cell 2008, 3, 429–441. [Google Scholar] [CrossRef] [Green Version]
- Jaggupilli, A.; Elkord, E. Significance of CD44 and CD24 as cancer stem cell markers: An enduring ambiguity. Clin. Dev. Immunol. 2012, 2012. [Google Scholar] [CrossRef] [Green Version]
- Fang, Y.; Yuan, Y.; Zhang, L.L.; Lu, J.W.; Feng, J.F.; Hu, S.N. Downregulated GBX2 gene suppresses proliferation, invasion and angiogenesis of breast cancer cells through inhibiting the Wnt/β-catenin signaling pathway. Cancer Biomark. 2018, 23, 405–418. [Google Scholar] [CrossRef]
- Peuhu, E.; Virtakoivu, R.; Mai, A.; Wärri, A.; Ivaska, J. Epithelial vimentin plays a functional role in mammary gland development. Development 2017, 144, 4103–4113. [Google Scholar] [CrossRef] [Green Version]
- Feng, Y.; Dai, X.; Li, X.; Wang, H.; Liu, J.; Zhang, J.; Du, Y.; Xia, L. EGF signalling pathway regulates colon cancer stem cell proliferation and apoptosis. Cell Prolif. 2012, 45, 413–419. [Google Scholar] [CrossRef]
- Cuneo, K.C.; Nyati, M.K.; Ray, D.; Lawrence, T.S. EGFR targeted therapies and radiation: Optimizing efficacy by appropriate drug scheduling and patient selection. Pharmacol. Ther. 2015, 154, 67–77. [Google Scholar] [CrossRef] [Green Version]
- Ramirez Williams, L.; Brüggemann, K.; Hubert, M.; Achmad, N.; Kiesel, L.; Schäfer, S.D.; Greve, B.; Götte, M. γ-Secretase inhibition affects viability, apoptosis, and the stem cell phenotype of endometriotic cells. Acta Obstet. Gynecol. Scand. 2019, 98, 1565–1574. [Google Scholar] [CrossRef] [Green Version]
- Venkatesh, V.; Nataraj, R.; Thangaraj, G.S.; Karthikeyan, M.; Gnanasekaran, A.; Kaginelli, S.B.; Kuppanna, G.; Kallappa, C.G.; Basalingappa, K.M. Targeting Notch signalling pathway of cancer stem cells. Stem Cell Investig. 2018, 5, 5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Angeloni, V.; Tiberio, P.; Appierto, V.; Daidone, M.G. Implications of stemness-related signaling pathways in breast cancer response to therapy. Semin. Cancer Biol. 2015, 31, 43–51. [Google Scholar] [CrossRef] [PubMed]
- Chiou, G.Y.; Yang, T.W.; Huang, C.C.; Tang, C.Y.; Yen, J.Y.; Tsai, M.C.; Chen, H.Y.; Fadhilah, N.; Lin, C.C.; Jong, Y.J. Musashi-1 promotes a cancer stem cell lineage and chemoresistance in colorectal cancer cells. Sci. Rep. 2017, 7, 1–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schulenburg, A.; Cech, P.; Herbacek, I.; Marian, B.; Wrba, F.; Valent, P.; Ulrich-Pur, H. CD44-positive colorectal adenoma cells express the potential stem cell markers musashi antigen (msi1) and ephrin B2 receptor (EphB2). J. Pathol. 2007, 213, 152–160. [Google Scholar] [CrossRef]
- Safa, A.R. Resistance to cell death and its modulation in cancer stem cells. Crit. Rev. Oncog. 2016, 21, 203–219. [Google Scholar] [CrossRef]
- Yadav, P.; Shankar, B.S. Radio resistance in breast cancer cells is mediated through TGF-β signalling, hybrid epithelial-mesenchymal phenotype and cancer stem cells. Biomed. Pharmacother. 2019, 111, 119–130. [Google Scholar] [CrossRef]
- Shen, Y.; Chen, H.; Zhang, J.; Chen, Y.; Wang, M.; Ma, J.; Hong, L.; Liu, N.; Fan, Q.; Lu, X.; et al. Increased Notch signaling enhances radioresistance of malignant stromal cells induced by glioma stem/progenitor cells. PLoS ONE 2015, 10, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Lagadec, C.; Vlashi, E.; Alhiyari, Y.; Phillips, T.M.; Bochkur Dratver, M.; Pajonk, F. Radiation-induced notch signaling in breast cancer stem cells. Int. J. Radiat. Oncol. Biol. Phys. 2013, 87, 609–618. [Google Scholar] [CrossRef] [Green Version]
- Wu, C.T.; Lin, W.Y.; Chang, Y.H.; Chen, W.C.; Chen, M.F. Impact of CD44 expression on radiation response for bladder cancer. J. Cancer 2017, 8, 1137–1144. [Google Scholar] [CrossRef] [Green Version]
- Tsubouchi, K.; Minami, K.; Hayashi, N.; Yokoyama, Y.; Mori, S.; Yamamoto, H.; Koizumi, M. The CD44 standard isoform contributes to radioresistance of pancreatic cancer cells. J. Radiat. Res. 2017, 58, 816–826. [Google Scholar] [CrossRef] [Green Version]
- Theys, J.; Jutten, B.; Habets, R.; Paesmans, K.; Groot, A.J.; Lambin, P.; Wouters, B.G.; Lammering, G.; Vooijs, M. E-Cadherin loss associated with EMT promotes radioresistance in human tumor cells. Radiother. Oncol. 2011, 99, 392–397. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, C.Y.; Lin, H.H.; Tang, M.J.; Wang, Y.K. Vimentin contributes to epithelial-mesenchymal transition ancer cell mechanics by mediating cytoskeletal organization and focal adhesion maturation. Oncotarget 2015, 6, 15966–15983. [Google Scholar] [PubMed] [Green Version]
- Ciszewski, W.M.; Tavecchio, M.; Dastych, J.; Curtin, N.J. DNA-PK inhibition by NU7441 sensitizes breast cancer cells to ionizing radiation and doxorubicin. Breast Cancer Res. Treat. 2014, 143, 47–55. [Google Scholar] [CrossRef] [PubMed]
- Mohiuddin, I.S.; Kang, M.H. DNA-PK as an Emerging Therapeutic Target in Cancer. Front. Oncol. 2019, 9, 1–8. [Google Scholar] [CrossRef]
- An, J.; Huang, Y.C.; Xu, Q.Z.; Zhou, L.J.; Shang, Z.F.; Huang, B.; Wang, Y.; Liu, X.D.; Wu, D.C.; Zhou, P.K. DNA-PKcs plays a dominant role in the regulation of H2AX phosphorylation in response to DNA damage and cell cycle progression. BMC Mol. Biol. 2010, 11, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Harari, P.M.; Huang, S.M. Combining EGFR inhibitors with radiation or chemotherapy: Will preclinical studies predict clinical results? Int. J. Radiat. Oncol. Biol. Phys. 2004, 58, 976–983. [Google Scholar] [CrossRef]
- S Steelman, L.; Fitzgerald, T.; Lertpiriyapong, K.; Cocco, L.; Y Follo, M.; M Martelli, A.; M Neri, L.; Marmiroli, S.; Libra, M.; Candido, S.; et al. Critical Roles of EGFR Family Members in Breast Cancer and Breast Cancer Stem Cells: Targets for Therapy. Curr. Pharm. Des. 2016, 22, 2358–2388. [Google Scholar] [CrossRef] [Green Version]
- Chen, D.; Sun, Y.; Wei, Y.; Zhang, P.; Rezaeian, A.H.; Teruya-Feldstein, J.; Gupta, S.; Liang, H.; Lin, H.-K.; Hung, M.-C.; et al. LIFR is a breast cancer metastasis suppressor upstream of the Hippo-YAP pathway and a prognostic marker. Nat. Med. 2012, 18, 1511–1517. [Google Scholar] [CrossRef] [Green Version]
- Iorns, E.; Ward, T.M.; Dean, S.; Jegg, A.; Thomas, D.; Murugaesu, N.; Sims, D.; Mitsopoulos, C.; Fenwick, K.; Kozarewa, I.; et al. Whole genome in vivo RNAi screening identifies the leukemia inhibitory factor receptor as a novel breast tumor suppressor. Breast Cancer Res. Treat. 2012, 135, 79–91. [Google Scholar] [CrossRef]
- Johnson, R.W.; Finger, E.C.; Olcina, M.M.; Vilalta, M.; Aguilera, T.; Miao, Y.; Merkel, A.R.; Johnson, J.R.; Sterling, J.A.; Wu, J.Y.; et al. Induction of LIFR confers a dormancy phenotype in breast cancer cells disseminated to the bone marrow. Nat. Cell Biol. 2016, 18, 1078–1089. [Google Scholar] [CrossRef] [Green Version]
- Viola, M.; Brüggemann, K.; Karousou, E.; Caon, I.; Caravà, E.; Vigetti, D.; Greve, B.; Stock, C.; De Luca, G.; Passi, A.; et al. MDA-MB-231 breast cancer cell viability, motility and matrix adhesion are regulated by a complex interplay of heparan sulfate, chondroitin−/dermatan sulfate and hyaluronan biosynthesis. Glycoconj. J. 2017, 34, 411–420. [Google Scholar] [CrossRef]
- Greve, B.; Sheikh-Mounessi, F.; Kemper, B.; Ernst, I.; Götte, M.; Eich, H.T. Survivin, a target to modulate the radiosensitivity of Ewing’s sarcoma. Strahlenther. Onkol. 2012, 188, 1038–1047. [Google Scholar] [CrossRef] [PubMed]
- Greve, B.; Dreffke, K.; Rickinger, A.; Könemann, S.; Fritz, E.; Eckardt-Schupp, F.; Amler, S.; Sauerland, C.; Braselmann, H.; Sauter, W.; et al. Multicentric investigation of ionising radiation-induced cell death as a predictive parameter of individual radiosensitivity. Apoptosis 2009, 14, 226–235. [Google Scholar] [CrossRef] [PubMed]
- Schwickert, A.; Weghake, E.; Brüggemann, K.; Engbers, A.; Brinkmann, B.F.; Kemper, B.; Seggewiß, J.; Stock, C.; Ebnet, K.; Kiesel, L.; et al. MicroRNA MIR-142-3p Inhibits Breast Cancer Cell Invasiveness by Synchronous Targeting of WASL, Integrin Alpha V, and Additional Cytoskeletal Elements. PLoS ONE 2015, 10, 1–22. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kemper, B.; von Bally, G. Digital holographic microscopy for live cell applications and technical inspection. Appl. Opt. 2008, 47, A52–A61. [Google Scholar] [CrossRef] [PubMed]
- Eggers, J.C.; Martino, V.; Reinbold, R.; Schäfer, S.D.; Kiesel, L.; Starzinski-Powitz, A.; Schüring, A.N.; Kemper, B.; Greve, B.; Götte, M. MicroRNA miR-200b affects proliferation, invasiveness and stemness of endometriotic cells by targeting ZEB1, ZEB2 and KLF4. Reprod. Biomed. Online 2016, 32, 434–445. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kemper, B.; Bauwens, A.; Vollmer, A.; Ketelhut, S.; Langehanenberg, P.; Müthing, J.; Karch, H.; von Bally, G. Label-free quantitative cell division monitoring of endothelial cells by digital holographic microscopy. J. Biomed. Opt. 2010, 15, 036009. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Characteristic | N (%) |
|---|---|
| Age (years) | |
| Range | 34–63 |
| Mean ± SD | 52 ± 8 |
| Menopausal status | |
| Premenopausal | 8 (42.1%) |
| Postmenopausal | 11 (57.9%) |
| Tumor size (cm) | |
| T1 | 3 (15.8%) |
| T2 | 9 (47.4%) |
| T3 | 5 (26.3%) |
| T4 | 2 (10.5%) |
| Tumor grade | |
| G I | 1 (5.3%) |
| G II | 17 (89.4%) |
| G III | 1 (5.3%) |
| Positive axillary lymph nodes | |
| N0 | 2 (10.5%) |
| N1 | 9 (47.4%) |
| N2 | 5 (26.3%) |
| N3 | 3 (15.8%) |
| Lymphovascular invasion | |
| Negative | 11 (57.9%) |
| Positive | 8 (42.1%) |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Troschel, F.M.; Minte, A.; Ismail, Y.M.; Kamal, A.; Abdullah, M.S.; Ahmed, S.H.; Deffner, M.; Kemper, B.; Kiesel, L.; Eich, H.T.; et al. Knockdown of Musashi RNA Binding Proteins Decreases Radioresistance but Enhances Cell Motility and Invasion in Triple-Negative Breast Cancer. Int. J. Mol. Sci. 2020, 21, 2169. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21062169
Troschel FM, Minte A, Ismail YM, Kamal A, Abdullah MS, Ahmed SH, Deffner M, Kemper B, Kiesel L, Eich HT, et al. Knockdown of Musashi RNA Binding Proteins Decreases Radioresistance but Enhances Cell Motility and Invasion in Triple-Negative Breast Cancer. International Journal of Molecular Sciences. 2020; 21(6):2169. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21062169
Chicago/Turabian StyleTroschel, Fabian M., Annemarie Minte, Yahia Mahmoud Ismail, Amr Kamal, Mahmoud Salah Abdullah, Sarah Hamdy Ahmed, Marie Deffner, Björn Kemper, Ludwig Kiesel, Hans Theodor Eich, and et al. 2020. "Knockdown of Musashi RNA Binding Proteins Decreases Radioresistance but Enhances Cell Motility and Invasion in Triple-Negative Breast Cancer" International Journal of Molecular Sciences 21, no. 6: 2169. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21062169