Maize ZmFNSI Homologs Interact with an NLR Protein to Modulate Hypersensitive Response


Abstract
:1. Introduction
2. Results
2.1. The Genes Encoding ZmFNSI and ZmS3H Are Induced in Rp1-D21 Compared to the Corresponding Wild Type
2.2. ZmFNSI-1 and ZmFNSI-2 Suppress Rp1-D21-Mediated HR in N. benthamiana
2.3. ZmFNSI-1 and ZmFNSI-2 Have No Obvious Suppression Roles on Other Elicitor-Induced HR
2.4. The Enzymatic Activity of ZmFNSI-1 and ZmFNSI-2 Might Not Be Required for Suppressing Rp1-D21-Mediated HR
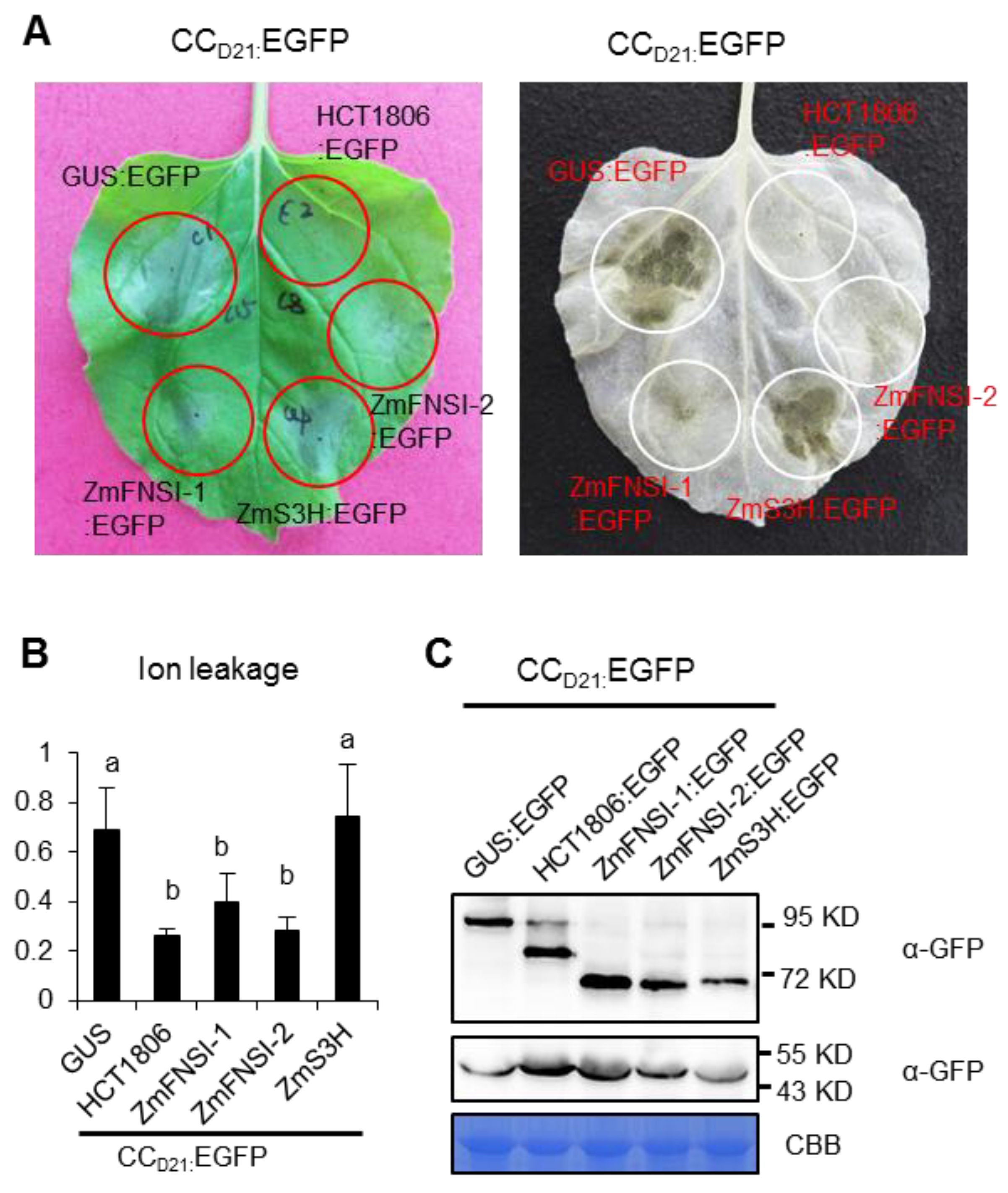
2.5. ZmFNSI-1 and ZmFNSI-2 Suppress CCD21-Mediated HR and Interact with CCD21
2.6. ZmFNSI-1 and ZmFNSI-2 Interact with HCT
2.7. ZmFNSI-1 and ZmFNSI-2 Form Stronger Homomers than ZmS3H
2.8. ZmFNSI-1 and ZmFNSI-2 do not Change the Subcellular Localization of CCD21
3. Discussion
3.1. ZmFNSI Functions in Plant Defense Response
3.2. FNSI Enzyme Activity Is not Required for HR and ZmFNSIs Are not General Cell Death Suppressors
3.3. FNSI/S5H and S3H have Overlapping and Different Roles in Plant Defense Response
3.4. ZmFNSI Regulates Rp1-D21-mediated HR possibly through Forming a Protein Complex
4. Materials and Methods
4.1. Plant Materials and Growth Condition
4.2. Sequence Alignment and Phylogenetic Analysis
4.3. Plasmid Construction
4.4. Agrobacterium tumefaciens-Mediated Transient Expression
4.5. Protein Analysis
4.6. Y2H
4.7. Confocal Microscopy
4.8. Accession Number
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Jones, J.D.; Dangl, J.L. The plant immune system. Nature 2006, 444, 323–329. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, W.; Feng, B.; Zhou, J.M.; Tang, D. Plant immune signaling: Advancing on two frontiers. J. Integr. Plant Biol. 2020, 62, 2–24. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bent, A.F.; Mackey, D. Elicitors, effectors, and R genes: The new paradigm and a lifetime supply of questions. Annu. Rev. Phytopathol. 2007, 45, 399–436. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Balint-Kurti, P. The plant hypersensitive response: Concepts, control and consequences. Mol. Plant Pathol. 2019, 20, 1163–1178. [Google Scholar] [CrossRef] [Green Version]
- Cui, H.; Tsuda, K.; Parker, J.E. Effector-triggered immunity: From pathogen perception to robust defense. Annu. Rev. Plant Biol. 2015, 66, 487–511. [Google Scholar] [CrossRef]
- Mur, L.A.; Kenton, P.; Lloyd, A.J.; Ougham, H.; Prats, E. The hypersensitive response; the centenary is upon us but how much do we know? J. Exp. Bot. 2008, 59, 501–520. [Google Scholar] [CrossRef] [Green Version]
- Kourelis, J.; van der Hoorn, R.A.L. Defended to the nines: 25 Years of resistance gene cloning identifies nine mechanisms for R protein function. Plant Cell 2018, 30, 285–299. [Google Scholar] [CrossRef] [Green Version]
- Dangl, J.L.; Jones, J.D. Plant pathogens and integrated defence responses to infection. Nature 2001, 411, 826–833. [Google Scholar] [CrossRef]
- Ellis, J.; Dodds, P.; Pryor, T. Structure, function and evolution of plant disease resistance genes. Curr. Opin. Plant Biol. 2000, 3, 278–284. [Google Scholar] [CrossRef]
- Monteiro, F.; Nishimura, M.T. Structural, functional, and genomic diversity of plant NLR proteins: An evolved resource for rational engineering of plant immunity. Annu. Rev. Phytopathol. 2018, 56, 243–267. [Google Scholar] [CrossRef]
- Hulbert, S.H. Structure and evolution of the rp1 complex conferring rust resistance in maize. Annu. Rev. Phytopathol. 1997, 35, 293–310. [Google Scholar] [CrossRef] [PubMed]
- Sudupak, M.A.; Bennetzen, J.L.; Hulbert, S.H. Unequal exchange and meiotic instability of disease-resistance genes in the Rp1 region of maize. Genetics 1993, 133, 119–125. [Google Scholar] [PubMed]
- Sun, Q.; Collins, N.C.; Ayliffe, M.; Smith, S.M.; Drake, J.; Pryor, T.; Hulbert, S.H. Recombination between paralogues at the Rp1 rust resistance locus in maize. Genetics 2001, 158, 423–438. [Google Scholar] [PubMed]
- Smith, S.M.; Steinau, M.; Trick, H.N.; Hulbert, S.H. Recombinant Rp1 genes confer necrotic or nonspecific resistance phenotypes. Mol. Genet. Genom. 2010, 283, 591–602. [Google Scholar] [CrossRef]
- Negeri, A.; Wang, G.F.; Benavente, L.; Kibiti, C.M.; Chaikam, V.; Johal, G.; Balint-Kurti, P. Characterization of temperature and light effects on the defense response phenotypes associated with the maize Rp1-D21 autoactive resistance gene. BMC Plant Biol. 2013, 13, 106. [Google Scholar] [CrossRef] [Green Version]
- Chintamanani, S.; Hulbert, S.H.; Johal, G.S.; Balint-Kurti, P.J. Identification of a maize locus that modulates the hypersensitive defense response, using mutant-assisted gene identification and characterization. Genetics 2010, 184, 813–825. [Google Scholar] [CrossRef] [Green Version]
- Chaikam, V.; Negeri, A.; Dhawan, R.; Puchaka, B.; Ji, J.; Chintamanani, S.; Gachomo, E.W.; Zillmer, A.; Doran, T.; Weil, C.; et al. Use of Mutant-Assisted Gene Identification and Characterization (MAGIC) to identify novel genetic loci that modify the maize hypersensitive response. Theor. Appl. Genet. 2011, 123, 985–997. [Google Scholar] [CrossRef]
- Olukolu, B.A.; Negeri, A.; Dhawan, R.; Venkata, B.P.; Sharma, P.; Garg, A.; Gachomo, E.; Marla, S.; Chu, K.; Hasan, A.; et al. A connected set of genes associated with programmed cell death implicated in controlling the hypersensitive response in maize. Genetics 2013, 193, 609–620. [Google Scholar] [CrossRef] [Green Version]
- Olukolu, B.A.; Wang, G.F.; Vontimitta, V.; Venkata, B.P.; Marla, S.; Ji, J.; Gachomo, E.; Chu, K.; Negeri, A.; Benson, J.; et al. A genome-wide association study of the maize hypersensitive defense response identifies genes that cluster in related pathways. PLoS Genet. 2014, 10, e1004562. [Google Scholar] [CrossRef] [Green Version]
- Wang, G.F.; He, Y.; Strauch, R.; Olukolu, B.A.; Nielsen, D.; Li, X.; Balint-Kurti, P.J. Maize homologs of hydroxycinnamoyltransferase, a key enzyme in lignin biosynthesis, bind the nucleotide binding leucine-rich repeat Rp1 proteins to modulate the defense response. Plant Physiol. 2015, 169, 2230–2243. [Google Scholar]
- Wang, G.F.; Balint-Kurti, P.J. Maize homologs of CCoAOMT and HCT, two key enzymes in lignin biosynthesis, form complexes with the NLR Rp1 protein to modulate the defense response. Plant Physiol. 2016, 171, 2166–2177. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Padmavati, M.; Reddy, A.R. Flavonoid biosynthetic pathway and cereal defence response: An emerging trend in crop biotechnology. J. Plant Biochem. Biot. 1999, 8, 15–20. [Google Scholar] [CrossRef]
- Fofana, B.; Benhamou, N.; McNally, D.J.; Labbe, C.; Seguin, A.; Belanger, R.R. Suppression of induced resistance in cucumber through disruption of the flavonoid pathway. Phytopathology 2005, 95, 114–123. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gill, U.S.; Uppalapati, S.R.; Gallego-Giraldo, L.; Ishiga, Y.; Dixon, R.A.; Mysore, K.S. Metabolic flux towards the (iso)flavonoid pathway in lignin modified alfalfa lines induces resistance against Fusarium oxysporum f. sp medicaginis. Plant Cell Environ. 2018, 41, 1997–2007. [Google Scholar]
- George, V.C.; Dellaire, G.; Rupasinghe, H.P.V. Plant flavonoids in cancer chemoprevention: Role in genome stability. J. Nutr. Biochem. 2017, 45, 1–14. [Google Scholar] [CrossRef]
- Peters, N.K.; Frost, J.W.; Long, S.R. A plant flavone, luteolin, induces expression of Rhizobium meliloti nodulation genes. Science 1986, 233, 977–980. [Google Scholar] [CrossRef]
- Lanubile, A.; Ferrarini, A.; Maschietto, V.; Delledonne, M.; Marocco, A.; Bellin, D. Functional genomic analysis of constitutive and inducible defense responses to Fusarium verticillioides infection in maize genotypes with contrasting ear rot resistance. BMC Genom. 2014, 15, 710. [Google Scholar] [CrossRef] [Green Version]
- Falcone Ferreyra, M.L.; Emiliani, J.; Rodriguez, E.J.; Campos-Bermudez, V.A.; Grotewold, E.; Casati, P. The identification of maize and Arabidopsis type I FLAVONE SYNTHASEs links flavones with hormones and biotic interactions. Plant Physiol. 2015, 169, 1090–1107. [Google Scholar] [CrossRef] [Green Version]
- van Damme, M.; Huibers, R.P.; Elberse, J.; Van den Ackerveken, G. Arabidopsis DMR6 encodes a putative 2OG-Fe(II) oxygenase that is defense-associated but required for susceptibility to downy mildew. Plant J. 2008, 54, 785–793. [Google Scholar] [CrossRef]
- Kawai, Y.; Ono, E.; Mizutani, M. Evolution and diversity of the 2-oxoglutarate-dependent dioxygenase superfamily in plants. Plant J. 2014, 78, 328–343. [Google Scholar] [CrossRef]
- Liu, Q.; Liu, H.; Gong, Y.; Tao, Y.; Jiang, L.; Zuo, W.; Yang, Q.; Ye, J.; Lai, J.; Wu, J.; et al. An atypical thioredoxin imparts early resistance to sugarcane mosaic virus in maize. Mol. Plant 2017, 10, 483–497. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Keskiaho, K.; Hieta, R.; Sormunen, R.; Myllyharju, J. Chlamydomonas reinhardtii has multiple prolyl 4-hydroxylases, one of which is essential for proper cell wall assembly. Plant Cell 2007, 19, 256–269. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, J.H.; Cheon, Y.M.; Kim, R.G.; Ahn, J.H. Analysis of flavonoids and characterization of the OsFNS gene involved in flavone biosynthesis in rice. J. Plant Biol. 2008, 51, 97–101. [Google Scholar] [CrossRef]
- Lee, Y.J.; Kim, J.H.; Kim, B.G.; Lim, Y.; Ahn, J.H. Characterization of flavone synthase I from rice. BMB Rep. 2008, 41, 68–71. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.; Zhao, L.; Zhao, J.; Li, Y.; Wang, J.; Guo, R.; Gan, S.; Liu, C.J.; Zhang, K. S5H/DMR6 encodes a salicylic acid 5-hydroxylase that fine-tunes salicylic acid homeostasis. Plant Physiol. 2017, 175, 1082–1093. [Google Scholar] [CrossRef] [Green Version]
- Zeilmaker, T.; Ludwig, N.R.; Elberse, J.; Seidl, M.F.; Berke, L.; Van Doorn, A.; Schuurink, R.C.; Snel, B.; Van den Ackerveken, G. DOWNY MILDEW RESISTANT 6 and DMR6-LIKE OXYGENASE 1 are partially redundant but distinct suppressors of immunity in Arabidopsis. Plant J. 2015, 81, 210–222. [Google Scholar] [CrossRef]
- Zhang, K.; Halitschke, R.; Yin, C.; Liu, C.J.; Gan, S.S. Salicylic acid 3-hydroxylase regulates Arabidopsis leaf longevity by mediating salicylic acid catabolism. Proc. Natl. Acad. Sci. USA 2013, 110, 14807–14812. [Google Scholar] [CrossRef] [Green Version]
- Wang, G.F.; Ji, J.; Ei-Kasmi, F.; Dangl, J.L.; Johal, G.; Balint-Kurti, P.J. Molecular and functional analyses of a maize autoactive NB-LRR protein identify precise structural requirements for activity. PLoS Pathog. 2015, 11, e1004674. [Google Scholar]
- Bai, S.; Liu, J.; Chang, C.; Zhang, L.; Maekawa, T.; Wang, Q.; Xiao, W.; Liu, Y.; Chai, J.; Takken, F.L.; et al. Structure-function analysis of barley NLR immune receptor MLA10 reveals its cell compartment specific activity in cell death and disease resistance. PLoS Pathog. 2012, 8, e1002752. [Google Scholar] [CrossRef] [Green Version]
- Gao, Z.; Chung, E.H.; Eitas, T.K.; Dangl, J.L. Plant intracellular innate immune receptor Resistance to Pseudomonas syringae pv. maculicola 1 (RPM1) is activated at, and functions on, the plasma membrane. Proc. Natl. Acad. Sci. USA 2011, 108, 7619–7624. [Google Scholar] [CrossRef] [Green Version]
- Kamoun, S.; van West, P.; Vleeshouwers, V.G.; de Groot, K.E.; Govers, F. Resistance of Nicotiana benthamiana to Phytophthora infestans is mediated by the recognition of the elicitor protein INF1. Plant Cell 1998, 10, 1413–1426. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lacomme, C.; Santa Cruz, S. Bax-induced cell death in tobacco is similar to the hypersensitive response. Proc. Natl. Acad. Sci. USA 1999, 96, 7956–7961. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martin, K.; Kopperud, K.; Chakrabarty, R.; Banerjee, R.; Brooks, R.; Goodin, M.M. Transient expression in Nicotiana benthamiana fluorescent marker lines provides enhanced definition of protein localization, movement and interactions in planta. Plant J. 2009, 59, 150–162. [Google Scholar] [CrossRef] [PubMed]
- Wang, G.F.; Balint-Kurti, P.J. Cytoplasmic and nuclear localizations are important for the hypersensitive response conferred by maize autoactive Rp1-D21 protein. Mol. Plant Microbe Interact. 2015, 28, 1023–1031. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thomazella, D.P.D.T.; Brail, Q.; Dahlbeck, D.; Staskawicz, B. CRISPR-Cas9 mediated mutagenesis of a DMR6 ortholog in tomato confers broad-spectrum disease resistance. bioRxiv 2016. [Google Scholar] [CrossRef] [Green Version]
- Yang, Q.; He, Y.; Kabahuma, M.; Chaya, T.; Kelly, A.; Borrego, E.; Bian, Y.; El Kasmi, F.; Yang, L.; Teixeira, P.; et al. A gene encoding maize caffeoyl-CoA O-methyltransferase confers quantitative resistance to multiple pathogens. Nat. Genet. 2017, 49, 1364–1372. [Google Scholar] [CrossRef]
- Liu, Y.; Guo, Y.; Ma, C.; Zhang, D.; Wang, C.; Yang, Q. Transcriptome analysis of maize resistance to Fusarium graminearum. BMC Genom. 2016, 17, 477. [Google Scholar]
- Kebede, A.Z.; Johnston, A.; Schneiderman, D.; Bosnich, W.; Harris, L.J. Transcriptome profiling of two maize inbreds with distinct responses to Gibberella ear rot disease to identify candidate resistance genes. BMC Genom. 2018, 19, 131. [Google Scholar] [CrossRef] [Green Version]
- Musungu, B.M.; Bhatnagar, D.; Brown, R.L.; Payne, G.A.; OBrian, G.; Fakhoury, A.M.; Geisler, M. A network approach of gene co-expression in the Zea mays/Aspergillus flavus pathosystem to map host/pathogen interaction pathways. Front. Genet. 2016, 7, 206. [Google Scholar] [CrossRef] [Green Version]
- Shu, X.; Livingston, D.P., 3rd; Woloshuk, C.P.; Payne, G.A. Comparative histological and transcriptional analysis of maize kernels infected with Aspergillus flavus and Fusarium verticillioides. Front. Plant Sci. 2017, 8, 2075. [Google Scholar] [CrossRef]
- Asselin, J.E.; Lin, J.; Perez-Quintero, A.L.; Gentzel, I.; Majerczak, D.; Opiyo, S.O.; Zhao, W.; Paek, S.M.; Kim, M.G.; Coplin, D.L.; et al. Perturbation of maize phenylpropanoid metabolism by an AvrE family type III effector from Pantoea stewartii. Plant Physiol. 2015, 167, 1117–1135. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ge, X.; Li, G.J.; Wang, S.B.; Zhu, H.; Zhu, T.; Wang, X.; Xia, Y. AtNUDT7, a negative regulator of basal immunity in Arabidopsis, modulates two distinct defense response pathways and is involved in maintaining redox homeostasis. Plant Physiol. 2007, 145, 204–215. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lan, W.; Lu, F.; Regner, M.; Zhu, Y.; Rencoret, J.; Ralph, S.A.; Zakai, U.I.; Morreel, K.; Boerjan, W.; Ralph, J. Tricin, a flavonoid monomer in monocot lignification. Plant Physiol. 2015, 167, 1284–1295. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jorgensen, K.; Rasmussen, A.V.; Morant, M.; Nielsen, A.H.; Bjarnholt, N.; Zagrobelny, M.; Bak, S.; Moller, B.L. Metabolon formation and metabolic channeling in the biosynthesis of plant natural products. Curr. Opin. Plant Biol. 2005, 8, 280–291. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.P.; Liu, B.; Sun, Y.; Chiang, V.L.; Sederoff, R.R. Enzyme-enzyme interactions in monolignol biosynthesis. Front. Plant Sci. 2019, 9, 1942. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Balint-Kurti, P.J.; Zwonitzer, J.C.; Wisser, R.J.; Carson, M.L.; Oropeza-Rosas, M.A.; Holland, J.B.; Szalma, S.J. Precise mapping of quantitative trait loci for resistance to southern leaf blight, caused by Cochliobolus heterostrophus race O, and flowering time using advanced intercross maize lines. Genetics 2007, 176, 645–657. [Google Scholar] [CrossRef] [Green Version]
- Lopez-Zuniga, L.O.; Wolters, P.; Davis, S.; Weldekidan, T.; Kolkman, J.M.; Nelson, R.; Hooda, K.S.; Rucker, E.; Thomason, W.; Wisser, R.; et al. Using maize chromosome segment substitution line populations for the identification of loci associated with multiple disease resistance. G3 Genes Genomes Genet. 2019, 9, 189–201. [Google Scholar] [CrossRef] [Green Version]
- Vlot, A.C.; Dempsey, D.A.; Klessig, D.F. Salicylic Acid, a multifaceted hormone to combat disease. Annu. Rev. Phytopathol. 2009, 47, 177–206. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Yang, Y.; Fang, B.; Gannon, P.; Ding, P.; Li, X.; Zhang, Y. Arabidopsis snc2-1D activates receptor-like protein-mediated immunity transduced through WRKY70. Plant Cell 2010, 22, 3153–3163. [Google Scholar] [CrossRef] [Green Version]
- Sun, Y.; Zhu, Y.-X.; Balint-Kurti, P.J.; Guan-Feng, W. Fine-tuning immunity: Players and regulators for plant NLRs. Trends Plant Sci. 2020. [Google Scholar] [CrossRef]
- Wang, G.; Roux, B.; Feng, F.; Guy, E.; Li, L.; Li, N.; Zhang, X.; Lautier, M.; Jardinaud, M.F.; Chabannes, M.; et al. The decoy substrate of a pathogen effector and a pseudokinase specify pathogen-induced modified-self recognition and immunity in plants. Cell Host Microbe 2015, 18, 285–295. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seto, D.; Koulena, N.; Lo, T.; Menna, A.; Guttman, D.S.; Desveaux, D. Expanded type III effector recognition by the ZAR1 NLR protein using ZED1-related kinases. Nat. Plants 2017, 3, 17027. [Google Scholar] [CrossRef] [PubMed]
- Baudin, M.; Hassan, J.A.; Schreiber, K.J.; Lewis, J.D. Analysis of the ZAR1 immune complex reveals determinants for immunity and molecular interactions. Plant Physiol. 2017, 174, 2038–2053. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, J.; Wang, J.; Hu, M.; Wu, S.; Qi, J.; Wang, G.; Han, Z.; Qi, Y.; Gao, N.; Wang, H.W.; et al. Ligand-triggered allosteric ADP release primes a plant NLR complex. Science 2019, 364, eaav5868. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Hu, M.; Wang, J.; Qi, J.; Han, Z.; Wang, G.; Qi, Y.; Wang, H.W.; Zhou, J.M.; Chai, J. Reconstitution and structure of a plant NLR resistosome conferring immunity. Science 2019, 364, eaav5870. [Google Scholar] [CrossRef]
- Hu, G.; Richter, T.E.; Hulbert, S.H.; Pryor, T. Disease lesion mimicry caused by mutations in the rust resistance gene rp1. Plant Cell 1996, 8, 1367–1376. [Google Scholar] [CrossRef]
- Bolus, S.; Akhunov, E.; Coaker, G.; Dubcovsky, J. Dissection of cell death induction by wheat stem rust resistance protein Sr35 and its matching effector AvrSr35. Mol. Plant Microbe Interact. MPMI 2020, 33, 308–319. [Google Scholar] [CrossRef] [Green Version]
- Slootweg, E.J.; Spiridon, L.N.; Martin, E.C.; Tameling, W.I.L.; Townsend, P.D.; Pomp, R.; Roosien, J.; Drawska, O.; Sukarta, O.C.A.; Schots, A.; et al. Distinct roles of non-overlapping surface regions of the coiled-coil domain in the potato immune receptor Rx1. Plant Physiol. 2018, 178, 1310–1331. [Google Scholar] [CrossRef] [Green Version]
- Tameling, W.I.; Nooijen, C.; Ludwig, N.; Boter, M.; Slootweg, E.; Goverse, A.; Shirasu, K.; Joosten, M.H. RanGAP2 mediates nucleocytoplasmic partitioning of the NB-LRR immune receptor Rx in the Solanaceae, thereby dictating Rx function. Plant Cell 2010, 22, 4176–4194. [Google Scholar] [CrossRef] [Green Version]
- Li, J.; Huang, H.N.; Zhu, M.; Huang, S.; Zhang, W.H.; Dinesh-Kumar, S.P.; Tao, X.R. A plant immune receptor adopts a two-step recognition mechanism to enhance viral effector perception. Mol. Plant 2019, 12, 248–262. [Google Scholar] [CrossRef] [Green Version]
- Larkin, M.A.; Blackshields, G.; Brown, N.P.; Chenna, R.; McGettigan, P.A.; McWilliam, H.; Valentin, F.; Wallace, I.M.; Wilm, A.; Lopez, R.; et al. Clustal W and Clustal X version 2.0. Bioinformatics 2007, 23, 2947–2948. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tamura, K.; Stecher, G.; Peterson, D.; Filipski, A.; Kumar, S. MEGA6: Molecular Evolutionary Genetics Analysis version 6.0. Mol. Biol. Evol. 2013, 30, 2725–2729. [Google Scholar] [CrossRef] [PubMed] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name | Accession Number | Chromosomal Position | FC in B73 × H95 Isogenic Lines | FC in Mo17 × H95 Isogenic Lines | Rp1-D21-Mediated HR Phenotype |
|---|---|---|---|---|---|
| ZmFNSI-1 | Zm00001d029744 | Chr1: 85,068,069..85,072,051 | 17.23 | 55.62 | Suppressed |
| ZmFNSI-2 | Zm00001d027423 | Chr1: 4,909,652..4,914,685 | 235.02 | 28.01 | Suppressed |
| ZmS3H | Zm00001d002564 | Chr2: 15,538,202..15,540,469 | 8.56 | 4.18 | No effect |
| HCT1806 | Zm00001d027946 | Chr1: 18,086,164..18,087,981 | 292.03 | 183.11 | Suppressed |
| HCT4918 | Zm00001d027948 | Chr1: 18,147,157..18,148,937 | 1115 | 568.5 | Suppressed |
| CCoAOMT2 | Zm00001d045206 | Chr9: 16,074,658..16,083,126 | 2.07 | 1.70 | Suppressed |
| Peptidoglycan related genes | Zm00001d043988 | Chr3: 215,679,092..215,679,979 | 250.59 | 69.97 | No effect |
| Zm00001d027325 | Chr1: 2,974,258..2,976,935 | 2.36 | 3.06 | No effect | |
| Zm00001d053695 | Chr4: 239,190,056..239,192,283 | 22.92 | 12.59 | No effect | |
| Wall associated kinase | Zm00001d003019 | Chr2: 29,627,526..29,635,338 | 5.82 | 4.84 | Autoactive HR |
| Zm00001d003021 | Chr2: 29,666,263..29,670,199 | 6.08 | 4.61 | Autoactive HR | |
| Auxin-responsive genes | Zm00001d033460 | Chr1: 263,312,679..263,313,005 | 24.76 | 177.41 | No effect |
| Zm00001d031666 | Chr1: 195785845-195789120 | 18.10 | 21.15 | No effect | |
| Zm00001d028167 | Chr1: 25,047,706..25,050,015 | 41.12 | 84.14 | No effect | |
| MTHFR2 | Zm00001d034602 | Chr1: 297,605,177..297,611,407 | 4.33 | 4.76 | No effect |
| Calmodulin binding proteins | Zm00001d023843 | Chr10: 24,286,776..24,290,769 | 157.85 | 85.47 | Autoactive HR |
| Zm00001d052525 | Chr4: 192,080,883..192,084,967 | 4.55 | 4.41 | No effect | |
| Zm00001d004916 | Chr2: 148,560,206..148,564,300 | 7.69 | 2.17 | No effect | |
| EF hand family | Zm00001d043258 | Chr3: 193,663,756..193,664,037 | 10.66 | 45.62 | Partially suppressed |
| UDP-glycosyltransferase | Zm00001d014126 | Chr5: 33,393,733..33,395,460 | 5.45 | 1.91 | No effect |
| Cytochrome B5 isoforms | Zm00001d017425 | Chr5: 195,722,803..195,723,646 | 123.41 | 163.02 | Enhanced |
| Zm00001d011081 | Chr8: 138,418,132..138,420,302 | 2.27 | 2.74 | Enhanced |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhu, Y.-X.; Ge, C.; Ma, S.; Liu, X.-Y.; Liu, M.; Sun, Y.; Wang, G.-F. Maize ZmFNSI Homologs Interact with an NLR Protein to Modulate Hypersensitive Response. Int. J. Mol. Sci. 2020, 21, 2529. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21072529
Zhu Y-X, Ge C, Ma S, Liu X-Y, Liu M, Sun Y, Wang G-F. Maize ZmFNSI Homologs Interact with an NLR Protein to Modulate Hypersensitive Response. International Journal of Molecular Sciences. 2020; 21(7):2529. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21072529
Chicago/Turabian StyleZhu, Yu-Xiu, Chunxia Ge, Shijun Ma, Xiao-Ying Liu, Mengjie Liu, Yang Sun, and Guan-Feng Wang. 2020. "Maize ZmFNSI Homologs Interact with an NLR Protein to Modulate Hypersensitive Response" International Journal of Molecular Sciences 21, no. 7: 2529. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21072529