A Non-Invasive Tool for Real-Time Measurement of Sulfate in Living Cells


 ,
,
Abstract
:1. Introduction
2. Results
2.1. Designing and Engineering of the FRET-Based Sulfate Sensor
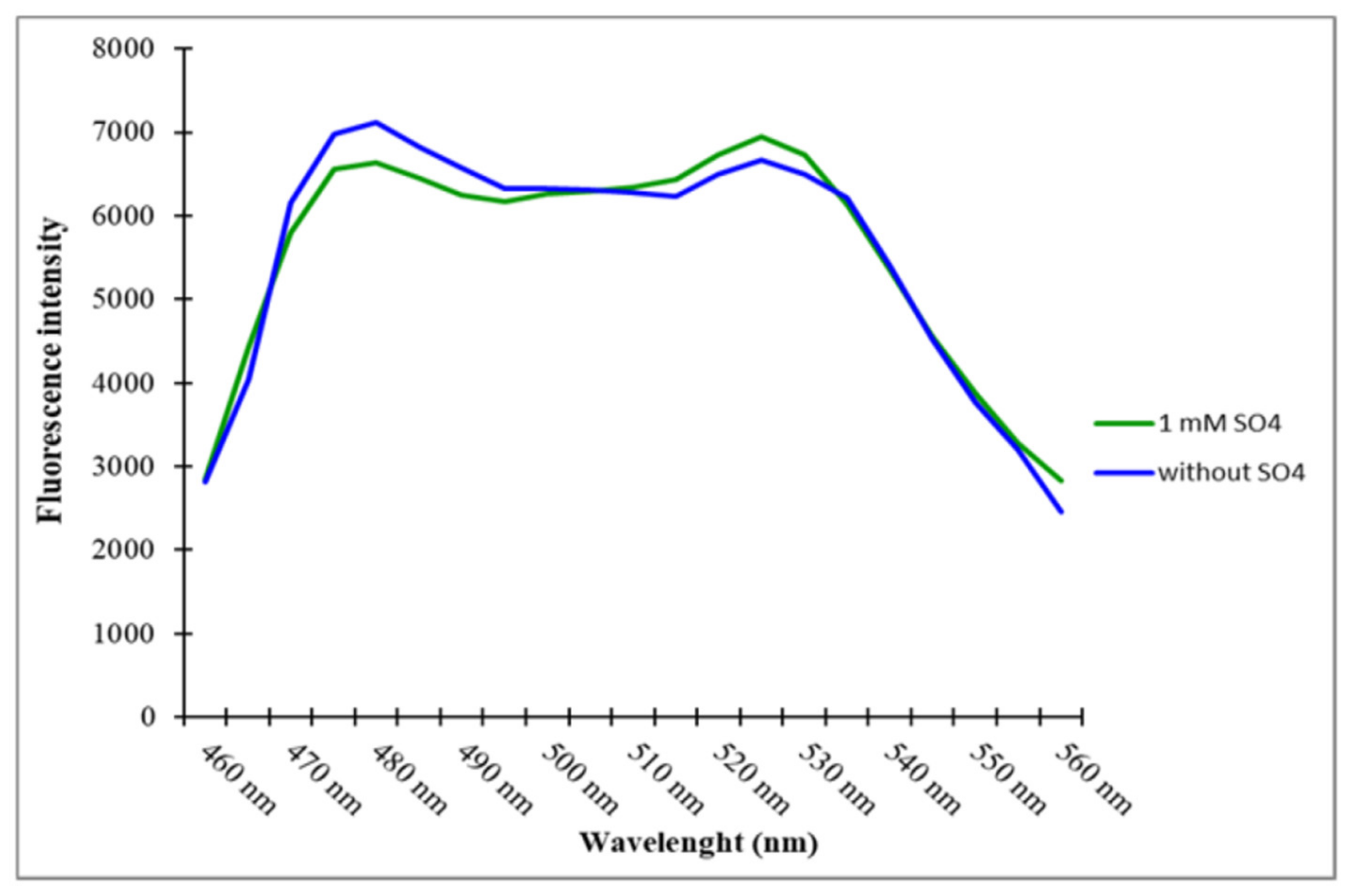
2.2. Purification and In Vitro Assay of the Sulfate Sensor
2.3. Affinity and Mutants of Sulfate Sensor
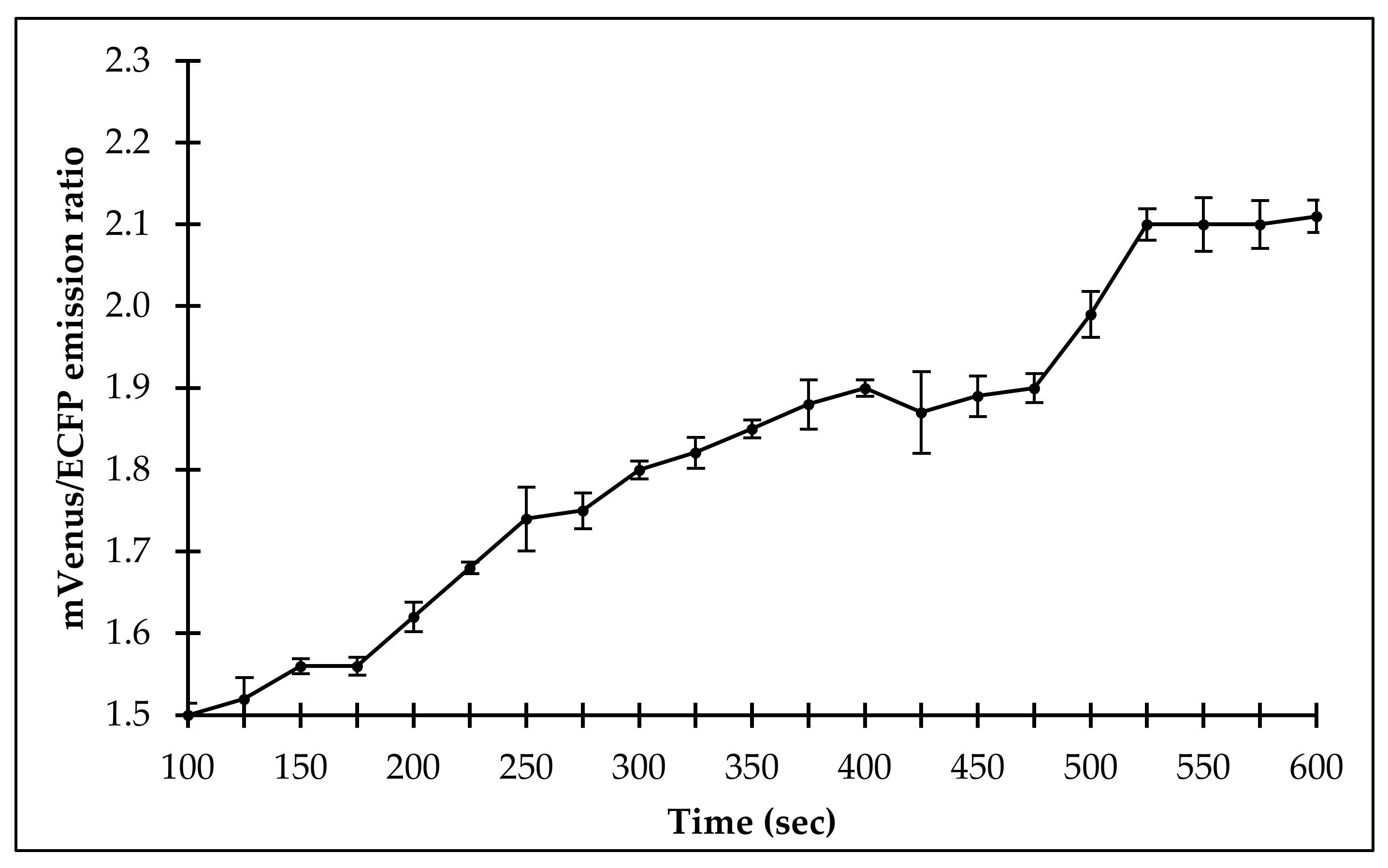
2.4. Measurement of Flux of Sulfate Uptake in Living Cells
2.4.1. In Bacteria
2.4.2. In Yeast
2.4.3. In Plant
3. Discussion
4. Materials and Methods
4.1. Molecular Docking
4.2. Designing and Construction of Sulfate Nanosensor
4.3. Expression and Purification of Sulfate Nanosensor Protein
4.4. Spectral Analysis, pH Stability, and Specificity of the Sulfate Sensor
4.5. Affinity (Kd) Analysis and Affinity Mutants of Sulfate Sensor
4.6. Measurement of Sulfate in Living Cells Using FLIP-SP
4.6.1. In Bacteria
4.6.2. In Yeast
4.6.3. In Plant
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| FRET | Fluorescent Resonance Energy Transfer |
| ECFP | Enhanced Cyan Fluorescent Protein |
| sbp | Sulfate Binding Protein |
References
- Buchner, P.; Takahashi, H.; Hawkesford, M.J. Plant sulphate transporters: Co-ordination of uptake, intracellular and long-distance transport. J. Exp. Bot. 2004, 55, 1765–1773. [Google Scholar] [CrossRef] [Green Version]
- Fahey, J.W.; Haristoy, X.; Dolan, P.M.; Kensler, T.W.; Scholtus, I.; Stephenson, K.K.; Talalay, P.; Lozniewski, A. Sulforaphane inhibits extracellular, intracellular, and antibiotic-resistant strains of Helicobacter pylori and prevents benzo[a]pyrene-induced stomach tumors. Proc. Natl. Acad. Sci. USA 2002, 99, 7610–7615. [Google Scholar] [CrossRef] [Green Version]
- Brosnan, J.T.; Brosnan, M.E. The sulfur-containing amino acids: an overview. J. Nutr. 2006, 136, 1636S–1640S. [Google Scholar] [CrossRef] [Green Version]
- Hirai, M.Y.; Fujiwara, T.; Awazuhara, M.; Kimura, T.; Noji, M.; Saito, K. Global expression profiling of sulfur-starved Arabidopsis by DNA macroarray reveals the role of O-acetyl-L-serine as a general regulator of gene expression in response to sulfur nutrition. Plant J. 2003, 33, 651–663. [Google Scholar] [CrossRef] [Green Version]
- Nikiforova, V.; Freitag, J.; Kempa, S.; Adamik, M.; Hesse, H.; Hoefgen, R. Transcriptome analysis of sulfur depletion in Arabidopsis thaliana: interlacing of biosynthetic pathways provides response specificity. Plant J. 2003, 33, 633–650. [Google Scholar] [CrossRef]
- LaLonde, S.; Ehrhardt, D.W.; Frommer, W.B. Shining light on signaling and metabolic networks by genetically encoded biosensors. Curr. Opin. Plant Boil. 2005, 8, 574–581. [Google Scholar] [CrossRef] [Green Version]
- Okumoto, S.; Jones, A.; Frommer, W.B. Quantitative Imaging with Fluorescent Biosensors. Annu. Rev. Plant Boil. 2012, 63, 663–706. [Google Scholar] [CrossRef]
- Liemburg-Apers, D.; Imamura, H.; Forkink, M.; Nooteboom, M.; Swarts, H.G.; Brock, R.; Smeitink, J.A.M.; Willems, P.H.; Koopman, W.J.H. Quantitative glucose and ATP sensing in mammalian cells. Pharm. Res. 2011, 28, 2745–2757. [Google Scholar] [CrossRef]
- Soleja, N.; Mohsin, M. Real time quantification of intracellular nickel using genetically encoded FRET-based nanosensor. Int. J. Boil. Macromol. 2019, 138, 648–657. [Google Scholar] [CrossRef]
- Fehr, M.; LaLonde, S.; Lager, I.; Wolff, M.W.; Frommer, W.B. In vivo imaging of the dynamics of glucose uptake in the cytosol of COS-7 cells by fluorescent nanosensors. J. Boil. Chem. 2003, 278, 19127–19133. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hol, W.G.J.; Van Duijnen, P.T.; Berendsen, H.J.C. The α-helix dipole and the properties of proteins. Nature 1978, 273, 443–446. [Google Scholar] [CrossRef] [PubMed]
- Trott, O.; Olson, A.J. AutoDock Vina: Improving the speed and accuracy of docking with a new scoring function, efficient optimization, and multithreading. J. Comput. Chem. 2010, 31, 455–461. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- He, J.J.; Quiocho, F.A. Dominant role of local dipoles in stabilizing uncompensated charges on a sulfate sequestered in a periplasmic active transport protein. Protein Sci. 1993, 2, 1643–1647. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khan, P.; Rahman, S.; Queen, A.; Manzoor, S.; Naz, F.; Hasan, G.M.; Luqman, S.; Kim, J.; Islam, A.; Ahmad, F.; et al. Elucidation of dietary polyphenolics as potential inhibitor of microtubule affinity regulating kinase 4: in silico and in vitro studies. Sci. Rep. 2017, 7, 9470. [Google Scholar] [CrossRef]
- Queen, A.; Khan, P.; Idrees, D.; Azam, A.; Hassan, I. Biological evaluation of p-toluene sulphonylhydrazone as carbonic anhydrase IX inhibitors: an approach to fight hypoxia-induced tumors. Int. J. Boil. Macromol. 2018, 106, 840–850. [Google Scholar] [CrossRef]
- Kaper, T.; Looger, L.L.; Takanaga, H.; Platten, M.; Steinman, L.; Frommer, W.B. Nanosensor detection of an immunoregulatory tryptophan influx/kynurenine efflux cycle. PLoS Boil. 2007, 5, e257. [Google Scholar] [CrossRef]
- Cheng, G.; Karunakaran, R.; East, A.K.; Poole, P.S. Multiplicity of sulfate and molybdate transporters and their role in nitrogen fixation in Rhizobium leguminosarum bv.viciae Rlv3841. Mol. Plant-Microbe Interact. 2016, 29, 143–152. [Google Scholar] [CrossRef] [Green Version]
- Pereira, C.T.; Roesler, C.; Faria, J.N.; Fessel, M.; Balan, A. Sulfate-Binding Protein (Sbp) from Xanthomonas citri: structure and functional insights. Mol. Plant-Microbe Interact. 2017, 30, 578–588. [Google Scholar] [CrossRef]
- Scheffel, F.; Demmer, U.; Warkentin, E.; Hülsmann, A.; Schneider, E.; Ermler, U. Structure of the ATPase subunit CysA of the putative sulfate ATP-binding cassette (ABC) transporter from Alicyclobacillus acidocaldarius. FEBS Lett. 2005, 579, 2953–2958. [Google Scholar] [CrossRef] [Green Version]
- Baird, G.S.; Zacharias, D.A.; Tsien, R.Y. Circular permutation and receptor insertion within green fluorescent proteins. Proc. Natl. Acad. Sci. USA 1999, 96, 11241–11246. [Google Scholar] [CrossRef] [Green Version]
- Zimmer, M. Green fluorescent protein (GFP): Applications, structure, and related photophysical behavior. Chem. Rev. 2002, 102, 759–782. [Google Scholar] [CrossRef] [PubMed]
- Giepmans, B.N.G. The fluorescent toolbox for assessing protein location and function. Science 2006, 312, 217–224. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Merzlyak, E.M.; Goedhart, J.; Shcherbo, D.; Bulina, E.M.; Shcheglov, A.S.; Fradkov, A.F.; Gaintzeva, A.; Lukyanov, K.A.; Lukyanov, S.; Gadella, T.W.J.; et al. Bright monomeric red fluorescent protein with an extended fluorescence lifetime. Nat. Methods 2007, 4, 555–557. [Google Scholar] [CrossRef] [PubMed]
- Fehr, M.; Frommer, W.B.; LaLonde, S. Visualization of maltose uptake in living yeast cells by fluorescent nanosensors. Proc. Natl. Acad. Sci. USA 2002, 99, 9846–9851. [Google Scholar] [CrossRef] [Green Version]
- Griesbeck, O.; Baird, G.S.; Campbell, R.E.; Zacharias, D.A.; Tsien, R.Y. Reducing the environmental sensitivity of yellow fluorescent protein: mechanism and applications. J. Boil. Chem. 2001, 276, 29188–29194. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.; Campbell, R.E.; Ting, A.Y.; Tsien, R.Y. Erratum: Creating new fluorescent probes for cell biology. Nat. Rev. Mol. Cell Boil. 2003, 4, 80. [Google Scholar] [CrossRef]
- Mukherjee, P.; Banerjee, S.; Wheeler, A.; Ratliff, L.A.; Irigoyen, S.; García, L.R.; Lockless, S.W.; Versaw, W.K. Live imaging of inorganic phosphate in plants with cellular and subcellular resolution. Plant Physiol. 2015, 167, 628–638. [Google Scholar] [CrossRef] [Green Version]
- Lindenburg, L.; Vinkenborg, J.L.; Oortwijn, J.; Aper, S.J.A.; Merkx, M. MagFRET: the first genetically encoded fluorescent Mg2+ sensor. PLoS ONE 2013, 8, e82009. [Google Scholar] [CrossRef] [Green Version]
- Nakanishi, Y.; Iida, S.; Ueoka-Nakanishi, H.; Niimi, T.; Tomioka, R.; Maeshima, M. Exploring dynamics of molybdate in living animal cells by a genetically encoded FRET nanosensor. PLoS ONE 2013, 8, e58175. [Google Scholar] [CrossRef]
- Mohsin, M.; Abdin, M.; Nischal, L.; Kardam, H.; Ahmad, A. Genetically encoded FRET-based nanosensor for in vivo measurement of leucine. Biosens. Bioelectron. 2013, 50, 72–77. [Google Scholar] [CrossRef]
- Ameen, S.; Ahmad, M.; Mohsin, M.; Qureshi, M.I.; Ibrahim, M.M.; Abdin, M.Z.; Ahmad, A. Designing, construction and characterization of genetically encoded FRET-based nanosensor for real time monitoring of lysine flux in living cells. J. Nanobiotechnol. 2016, 14, 49. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ahmad, M.; Ameen, S.; Siddiqi, T.O.; Khan, P.; Ahmad, A. Live cell monitoring of glycine betaine by FRET-based genetically encoded nanosensor. Biosens. Bioelectron. 2016, 86, 169–175. [Google Scholar] [CrossRef] [PubMed]
- Pflugrath, J.W.; Quiocho, F.A. Sulphate sequestered in the sulphate-binding protein of Salmonella typhimurium is bound solely by hydrogen bonds. Nature 1985, 314, 257–260. [Google Scholar] [CrossRef] [PubMed]
- Pflugrath, J.W.; Quiocho, F.A. The 2 Å resolution structure of the sulfate-binding protein involved in active transport in Salmonella typhimurium. J. Mol. Boil. 1988, 200, 163–180. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| S.no | Sensor Name | Sequences | Kd(µM) | Dynamic Range (µM) | ∆Rmax § |
|---|---|---|---|---|---|
| 1 | FLIP-WT (WT) | Wild type | 10 | 0.3 µM–90 µM | 0.7 |
| 2 | FLIP-130 (M-1) | S130I | 8 | 0.2 µM–50 µM | 0.6 |
| 3 | FLIP-132 (M-2) | G132Q | 45 | 40 µM–800 µM | 0.4 |
| 4 | FLIP-192 (M-3) | W192A | 6 | 0.5 µM–80 µM | 0.7 |
| 5 | FLIP-11 (M-4) | D11G | no binding | ||
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fatima, U.; Okla, M.K.; Mohsin, M.; Naz, R.; Soufan, W.; Al-Ghamdi, A.A.; Ahmad, A. A Non-Invasive Tool for Real-Time Measurement of Sulfate in Living Cells. Int. J. Mol. Sci. 2020, 21, 2572. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21072572
Fatima U, Okla MK, Mohsin M, Naz R, Soufan W, Al-Ghamdi AA, Ahmad A. A Non-Invasive Tool for Real-Time Measurement of Sulfate in Living Cells. International Journal of Molecular Sciences. 2020; 21(7):2572. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21072572
Chicago/Turabian StyleFatima, Urooj, Mohammad K. Okla, Mohd Mohsin, Ruphi Naz, Walid Soufan, Abdullah A. Al-Ghamdi, and Altaf Ahmad. 2020. "A Non-Invasive Tool for Real-Time Measurement of Sulfate in Living Cells" International Journal of Molecular Sciences 21, no. 7: 2572. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21072572