Galectin-2 Has Bactericidal Effects against Helicobacter pylori in a β-galactoside-Dependent Manner

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
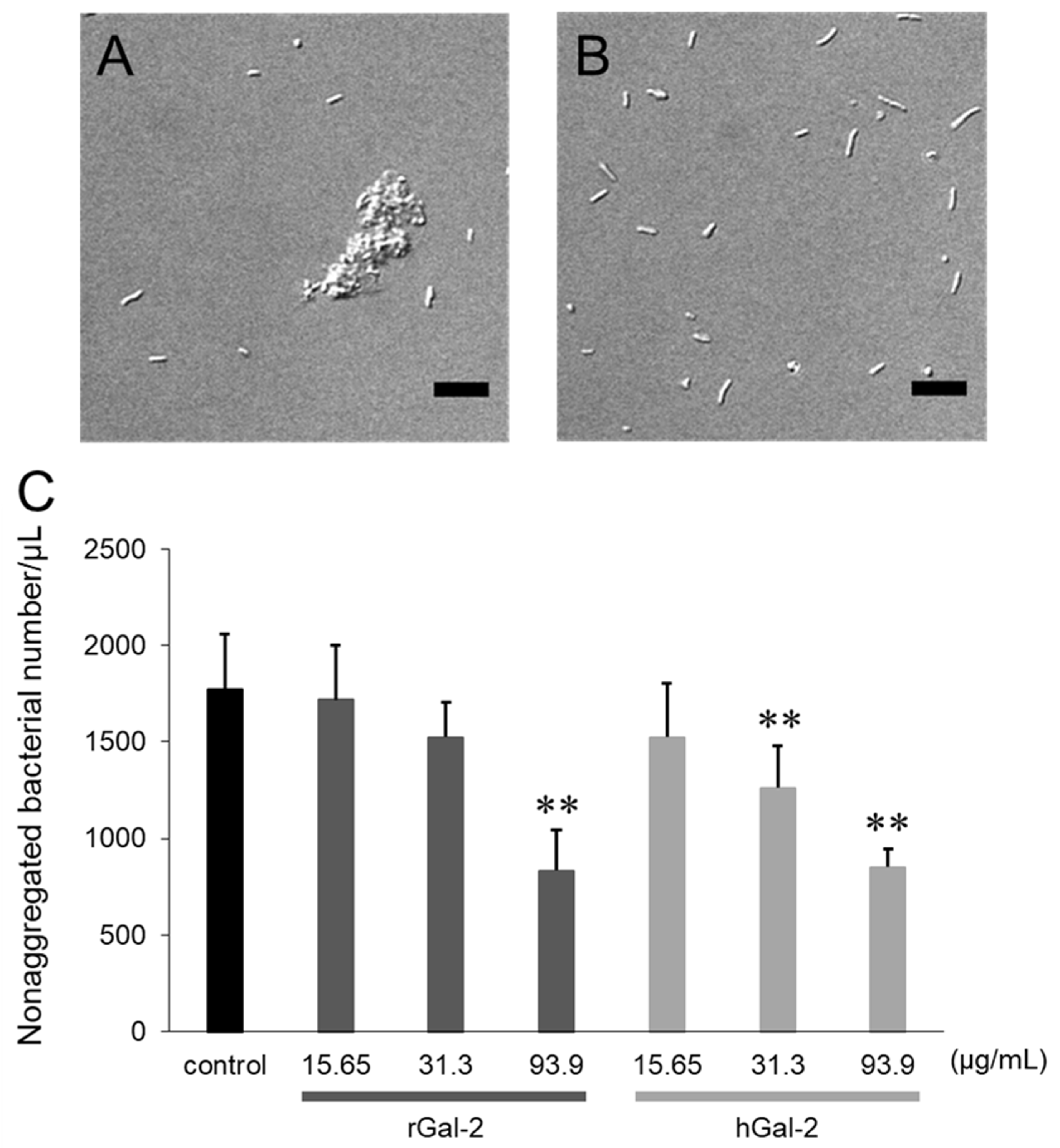
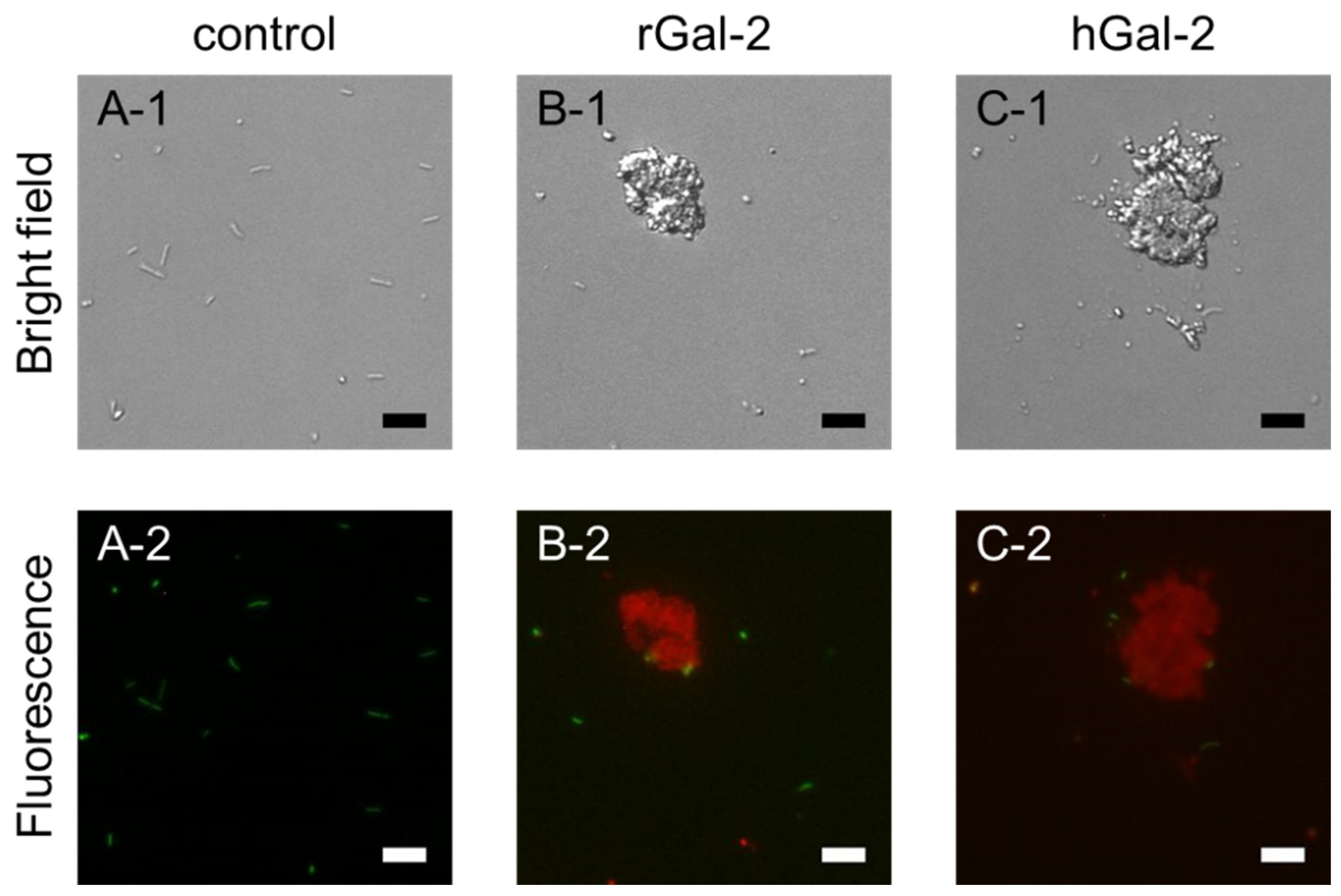
2.1. Gal-2 Induces Aggregation of H. pylori
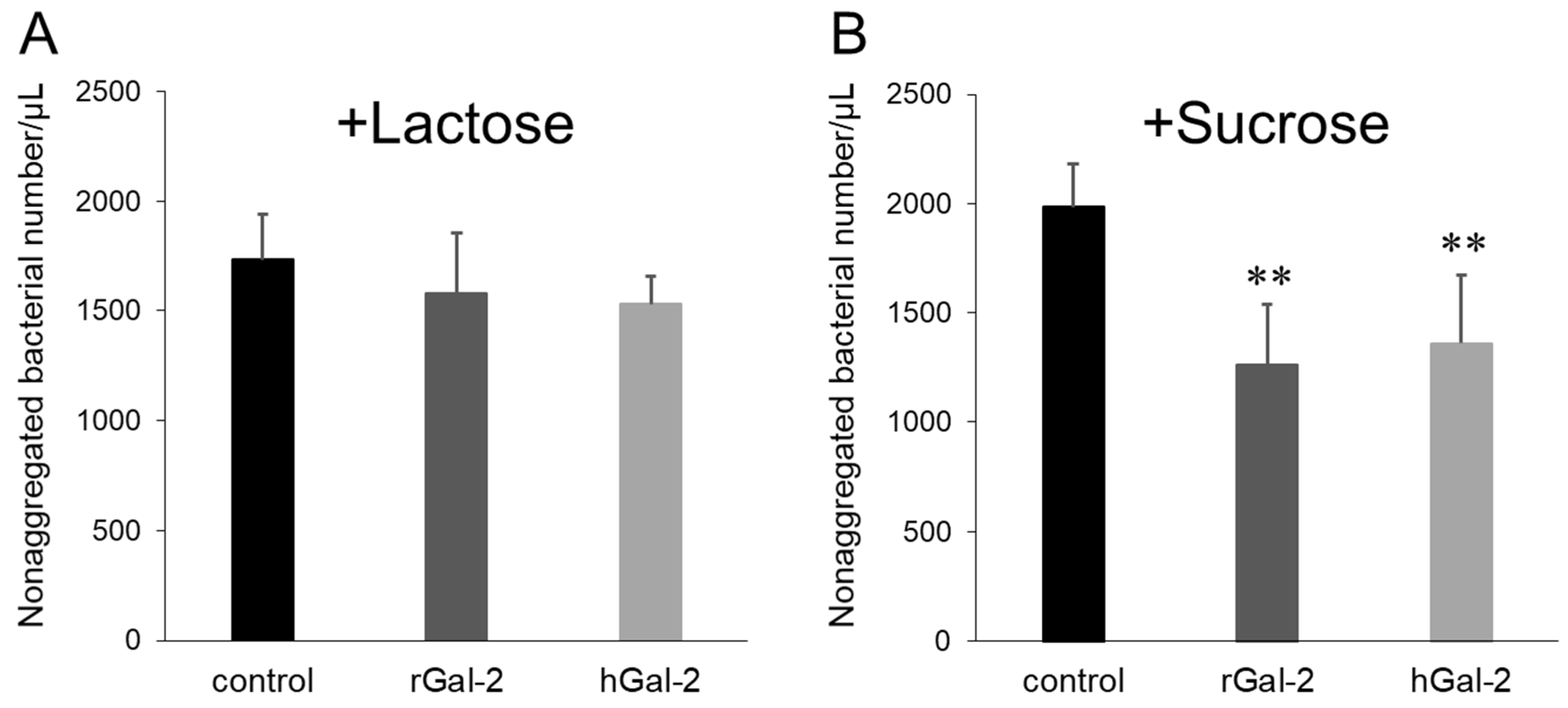
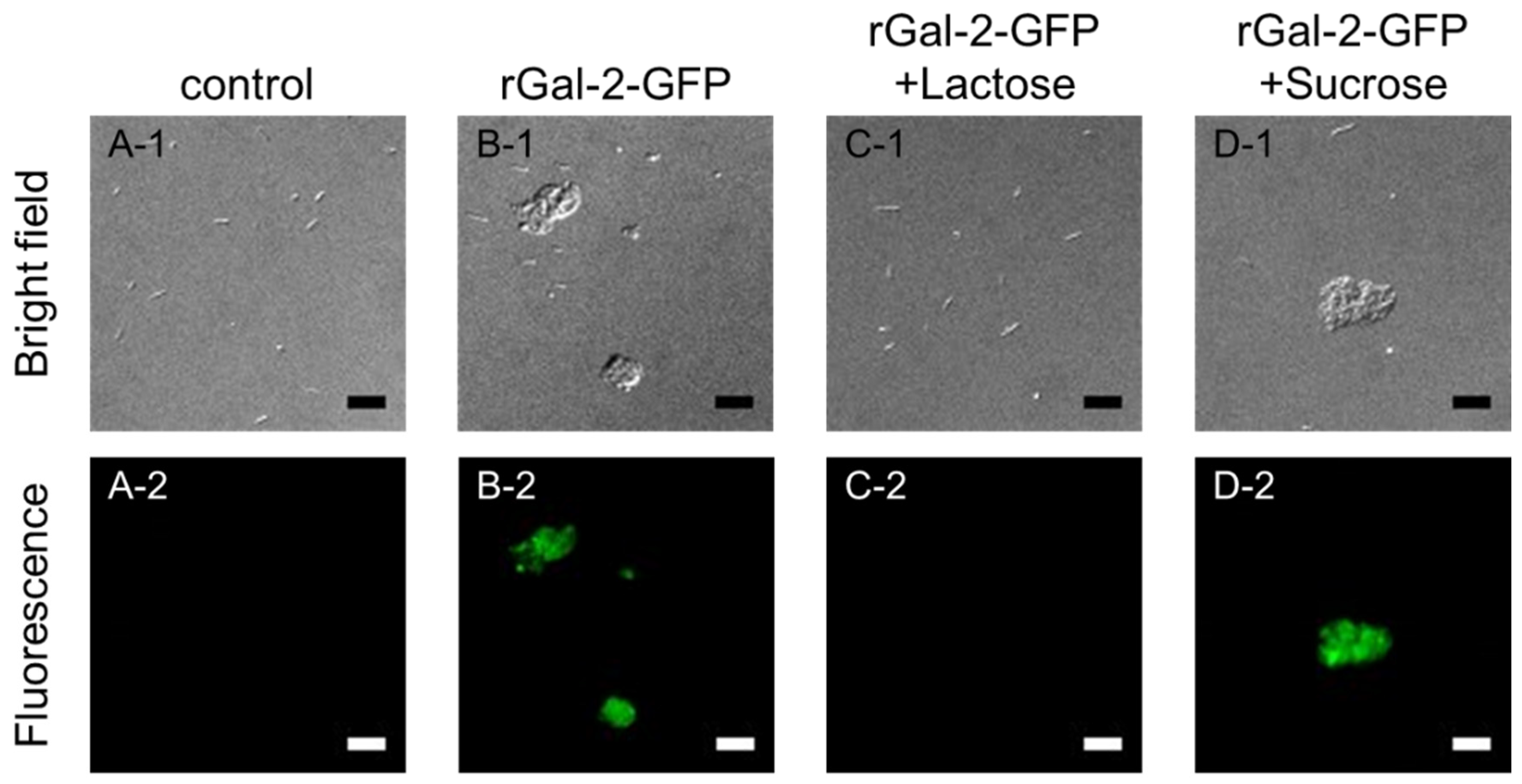
2.2. Gal-2-H. pylori Interaction Depends on β-galactosides
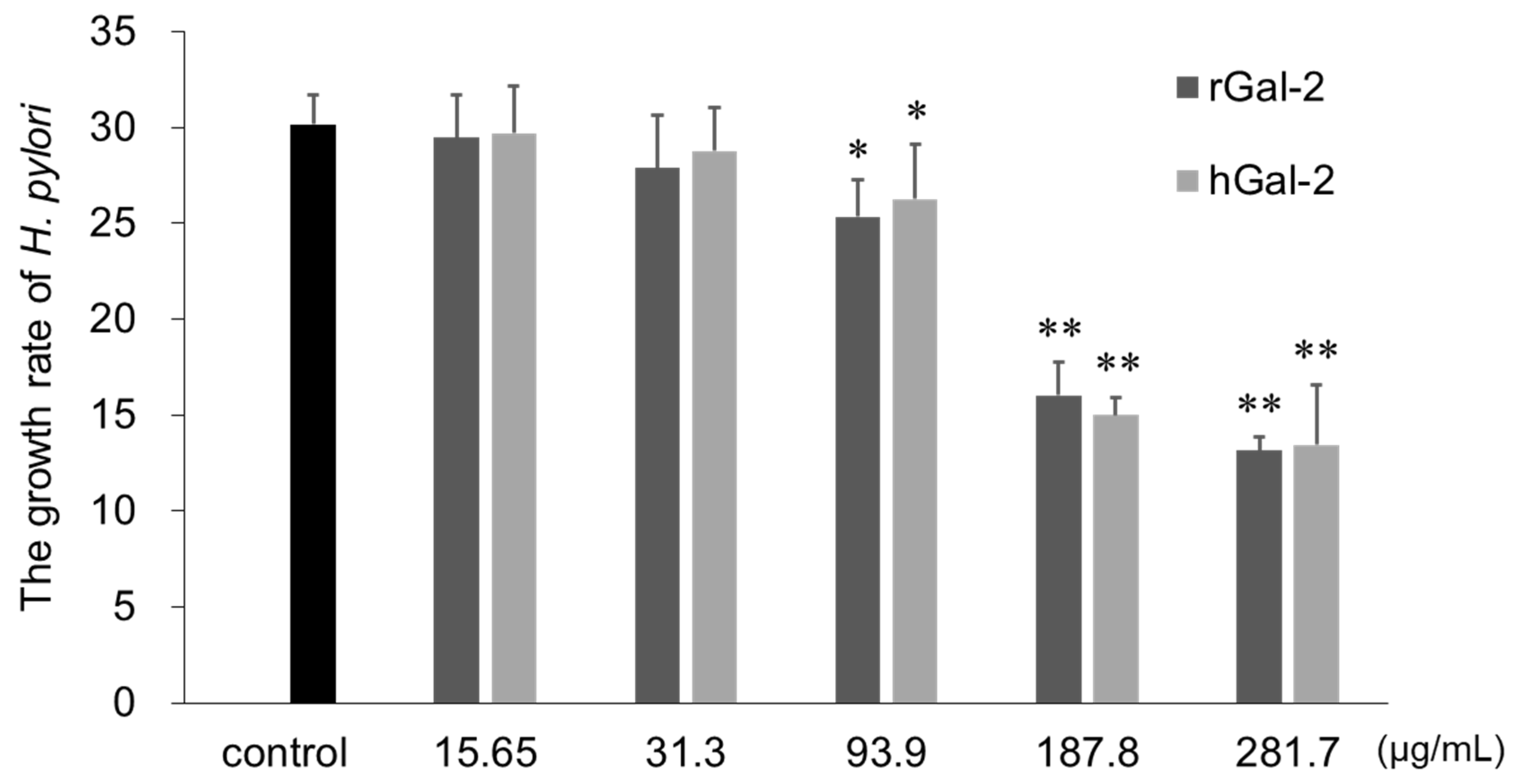
2.3. Gal-2 Inhibits H. pylori Growth
2.4. Gal-2 has Bactericidal Effects against H. pylori
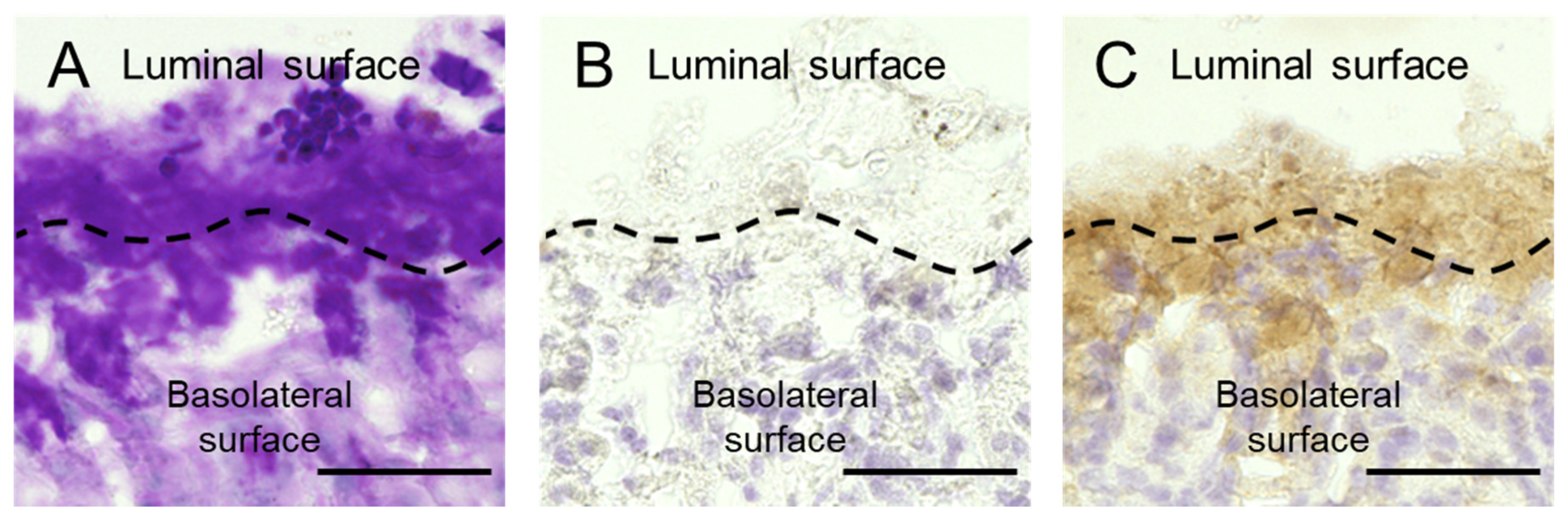
2.5. Gal-2 Distributes in Gastric Mucus
3. Discussion
4. Materials and Methods
4.1. H. pylori and Growth Condition
4.2. Preparation of H. pylori Suspension
4.3. Purification of Recombinant Galectin Proteins
4.4. Aggregation Assay
4.5. Inhibitory Activity of Gal-2 against H. pylori Growth
4.6. Bactericidal Effect of Gal-2
4.7. Preparation of Mouse Gastric Tissue Section
4.8. PAS Staining and IHC
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Robin Warren, J.; Marshall, B. Unidentified curved bacilli on gastric epithelium in active chronic gastritis. Lancet 1983, 321, 1273–1275. [Google Scholar] [CrossRef]
- Price, A.B.; Levi, J.; Dolby, J.M.; Dunscombe, P.L.; Smith, A.; Clark, J.; Stephenson, M.L. Campylobacter pyloridis in peptic ulcer disease: Microbiology, pathology, and scanning electron microscopy. Gut 1985, 26, 1183–1188. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pereira, M.I.; Medeiros, J.A. Role of helicobacter pylori in gastric mucosa-associated lymphoid tissue lymphomas. World J. Gastroenterol. 2014, 20, 684–698. [Google Scholar] [CrossRef] [PubMed]
- International Agency for Research on Cancer. Infection with helicobacter pylori. in: Iarc monographs on the evaluation of carcinogenic risks to humans. Schisto Somiasis Liver Flukes Helicobacter Pylori 1994, 61, 177–241. [Google Scholar]
- Asaka, M. A new approach for elimination of gastric cancer deaths in japan. Int. J. Cancer 2013, 132, 1272–1276. [Google Scholar] [CrossRef] [Green Version]
- Asaka, M.; Kato, M.; Sakamoto, N. Roadmap to eliminate gastric cancer with helicobacter pylori eradication and consecutive surveillance in japan. J. Gastroenterol. 2014, 49, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Nishida, T.; Tsujii, M.; Tanimura, H.; Tsutsui, S.; Tsuji, S.; Takeda, A.; Inoue, A.; Fukui, H.; Yoshio, T.; Kishida, O.; et al. Comparative study of esomeprazole and lansoprazole in triple therapy for eradication of helicobacter pylori in japan. World J. Gastroenterol. 2014, 20, 4362–4369. [Google Scholar] [CrossRef]
- Ohtaka, M.; Miura, M.; Hanawa, M.; Hirose, Y.; Kitahashi, A.; Imamura, N.; Watanabe, I.; Takaso, K.; Shimura, N.; Yoda, Y.; et al. Efficacy and tolerability of second-line metronidazole triple therapy using vonoprazan for helicobacter pylori eradication in japan—comparative study: Vonoprazan vs. proton pump inhibitors. Open J. Gastroenterol. 2018, 8, 27–38. [Google Scholar] [CrossRef] [Green Version]
- Thung, I.; Aramin, H.; Vavinskaya, V.; Gupta, S.; Park, J.Y.; Crowe, S.E.; Valasek, M.A. Review article: The global emergence of helicobacter pylori antibiotic resistance. Aliment. Pharmacol. 2016, 43, 514–533. [Google Scholar] [CrossRef] [Green Version]
- Nio-Kobayashi, J. Histological mapping and subtype-specific functions of galectins in health and disease. Trends Glycosci. Glycotechnol. 2018, 30, SE89–SE96. [Google Scholar] [CrossRef] [Green Version]
- Kasai, K.I.; Hirabayashi, J. Galectins: A family of animal lectins that decipher glycocodes. J. Biochem. 1996, 119, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Wang, G.; Ge, Z.; Rasko, D.A.; Taylor, D.E. Lewis antigens in helicobacter pylori: Biosynthesis and phase variation. Mol. Microbiol. 2000, 36, 1187–1196. [Google Scholar] [CrossRef] [PubMed]
- Appelmelk, B.J.; Monteiro, M.A.; Martin, S.L.; Moran, A.P.; Vandenbroucke-Grauls, C.M.J.E. Why helicobacter pylori has lewis antigens. Trends Microbiol. 2000, 8, 565–570. [Google Scholar] [CrossRef]
- Jin, C.; Kenny, D.T.; Skoog, E.C.; Padra, M.; Adamczyk, B.; Vitizeva, V.; Thorell, A.; Venkatakrishnan, V.; Lindén, S.K.; Karlsson, N.G. Structural diversity of human gastric mucin glycans. Mol. Cell. Proteom. 2017, 16, 743–758. [Google Scholar] [CrossRef]
- Robinson, B.S.; Arthur, C.M.; Kamili, N.A.; Stowell, S.R. Galectin regulation of host microbial interactions. Trends Glycosci. Glycotechnol. 2018, 30, SE185–SE198. [Google Scholar] [CrossRef]
- Fowler, M.; Thomas, R.J.; Atherton, J.; Roberts, I.S.; High, N.J. Galectin-3 binds to helicobacter pylori o-antigen: It is upregulated and rapidly secreted by gastric epithelial cells in response to h. pylori adhesion. Cell. Microbiol. 2006, 8, 44–54. [Google Scholar] [CrossRef]
- Park, A.M.; Hagiwara, S.; Hsu, D.K.; Liu, F.T.; Yoshie, O. Galectin-3 plays an important role in innate immunity to gastric infection by helicobacter pylori. Infect. Immun. 2016, 84, 1184–1193. [Google Scholar] [CrossRef] [Green Version]
- Nio-Kobayashi, J.; Takahashi-Iwanaga, H.; Iwanaga, T. Immunohistochemical localization of six galectin subtypes in the mouse digestive tract. J. Histochem. Cytochem. 2009, 57, 41–50. [Google Scholar] [CrossRef]
- Oka, T.; Murakami, S.; Arata, Y.; Hirabayashi, J.; Kasai, K.I.; Wada, Y.; Futai, M. Identification and cloning of rat galectin-2: Expression is predominantly in epithelial cells of the stomach. Arch. Biochem. Biophys. 1999, 361, 195–201. [Google Scholar] [CrossRef]
- Saal, I.; Lensch, M.; Lohr, M.; Manning, J.C.; Decaestecker, C.; André, S.; Kiss, R.; Salmon, I.; Gabius, H.J. Human galectin-2: Expression profiling by rt-pcr/immunohistochemistry and its introduction as a histochemical tool for ligand localization. Histol. Histopathol. 2005, 20, 1191–1208. [Google Scholar]
- Tamura, M.; Sato, D.; Nakajima, M.; Saito, M.; Sasaki, T.; Tanaka, T.; Hatanaka, T.; Takeuchi, T.; Arata, Y. Identification of galectin-2–mucin interaction and possible formation of a high molecular weight lattice. Biol. Pharm. Bull. 2017, 40, 1789–1795. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paclik, D.; Berndt, U.; Guzy, C.; Dankof, A.; Danese, S.; Holzloehner, P.; Rosewicz, S.; Wiedenmann, B.; Wittig, B.M.; Dignass, A.U.; et al. Galectin-2 induces apoptosis of lamina propria t lymphocytes and ameliorates acute and chronic experimental colitis in mice. J. Mol. Med. 2008, 86, 1395–1406. [Google Scholar] [CrossRef] [PubMed]
- Jung, J.H.; Kim, H.J.; Yeom, J.; Yoo, C.; Shin, J.; Yoo, J.; Kang, C.S.; Lee, C. Lowered expression of galectin-2 is associated with lymph node metastasis in gastric cancer. J. Gastroenterol. 2012, 47, 37–48. [Google Scholar] [CrossRef] [PubMed]
- Takaishi, S.; Wang, T.C. Gene expression profiling in a mouse model of helicobacter-induced gastric cancer. Cancer Sci. 2007, 98, 284–293. [Google Scholar] [CrossRef]
- Lobsanov, Y.D.; Gitt, M.A.; Leffler, H.; Barondes, S.H.; Rini, J.M. X-ray crystal structure of the human dimeric s-lac lectin, l-14-ii, in complex with lactose at 2.9-å resolution. J. Biol. Chem. 1993, 268, 27034–27038. [Google Scholar]
- Hirabayashi, J.; Hashidate, T.; Arata, Y.; Nishi, N.; Nakamura, T.; Hirashima, M.; Urashima, T.; Oka, T.; Futai, M.; Muller, W.E.G.; et al. Oligosaccharide specificity of galectins: A search by frontal affinity chromatography. Biochim. Biophys. Acta Gen. Subj. 2002, 1572, 232–254. [Google Scholar] [CrossRef]
- Moran, A.P.; Knirel, Y.A.; Senchenkova, S.F.Y.N.; Widmalm, G.; Hynes, S.O.; Jansson, P.E. Phenotypic variation in molecular mimicry between helicobacter pylori lipopolysaccharides and human gastric epithelial cell surface glycoforms. acid-induced phase variation in lewisx and lewisy expression by h. pylori lipopolysaccharides. J. Biol. Chem. 2002, 277, 5785–5795. [Google Scholar] [CrossRef] [Green Version]
- Khamri, W.; Moran, A.P.; Worku, M.L.; Karim, Q.N.; Walker, M.M.; Annuk, H.; Ferris, J.A.; Appelmelk, B.J.; Eggleton, P.; Reid, K.B.M.; et al. Variations in helicobacter pylori lipopolysaccharide to evade the innate immune component surfactant protein d. Infect. Immun. 2005, 73, 7677–7686. [Google Scholar] [CrossRef] [Green Version]
- Appelmelk, B.J.; Shiberu, B.; Trinks, C.; Tapsi, N.; Zheng, P.Y.; Verboom, T.; Maaskant, J.; Hokke, C.H.; Schiphorst, W.E.; Blanchard, D.; et al. Phase variation in helicobacter pylori lipopolysaccharide. Infect. Immun. 1998, 66, 70–76. [Google Scholar] [CrossRef] [Green Version]
- Sakakura, M.; Tamura, M.; Fujii, N.; Takeuchi, T.; Hatanaka, T.; Kishimoto, S.; Arata, Y.; Takahashi, H. Structural mechanisms for the s-nitrosylation-derived protection of mouse galectin-2 from oxidation-induced inactivation revealed by nmr. FEBS J. 2018, 285, 1129–1145. [Google Scholar] [CrossRef]
- Stowell, S.R.; Arthur, C.M.; Dias-Baruffi, M.; Rodrigues, L.C.; Gourdine, J.P.; Heimburg-Molinaro, J.; Ju, T.; Molinaro, R.J.; Rivera-Marrero, C.; Xia, B.; et al. Innate immune lectins kill bacteria expressing blood group antigen. Nat. Med. 2010, 16, 295–301. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barrow, H.; Guo, X.; Wandall, H.H.; Pedersen, J.W.; Fu, B.; Zhao, Q.; Chen, C.; Rhodes, J.M.; Yu, L.G. Serum galectin-2, -4, and -8 are greatly increased in colon and breast cancer patients and promote cancer cell adhesion to blood vascular endothelium. Clin. Cancer Res. 2011, 17, 7035–7046. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ahmed, H.; Fink, N.E.; Pohl, J.; Vasta, G.R. Galectin-1 from bovine spleen: Biochemical characterization, carbohydrate specificity and tissue-specific isoform profiles. J. Biochem. 1996, 120, 1007–1019. [Google Scholar] [CrossRef] [PubMed]
- Niki, T.; Fujita, K.; Rosen, H.; Hirashima, M.; Masaki, T.; Hattori, T.; Hoshino, K. Plasma galectin-9 concentrations in normal and diseased condition. Cell. Physiol. Biochem. 2018, 50, 1856–1868. [Google Scholar] [CrossRef] [PubMed]
- Estevam, R.B.; Da Silva, N.M.J.W.; Da Silva, W.; Fonseca, F.M.; De Oliveira, A.G.; De Lima Pereira, S.A.; Pereira, T.L.; Adad, S.J.; Rodrigues, V.J.; Rodrigues, D.B.R. Modulation of galectin-3 and galectin 9 in gastric mucosa of patients with chronic gastritis and positive helicobacter pylori infection. Pathol. Res. Pract. 2017, 213, 1276–1281. [Google Scholar] [CrossRef]
- Van der Laan, A.M.; Schirmer, S.H.; De Vries, M.R.; Koning, J.J.; Volger, O.L.; Fledderus, J.O.; Bastiaansen, A.J.N.M.; Hollander, M.R.; Baggen, J.M.; Koch, K.T.; et al. Galectin-2 expression is dependent on the rs7291467 polymorphism and acts as an inhibitor of arteriogenesis. Eur. Heart J. 2012, 33, 1076–1084. [Google Scholar] [CrossRef] [Green Version]
- Yildirim, C.; Vogel, D.Y.S.; Hollander, M.R.; Baggen, J.M.; Fontijn, R.D.; Nieuwenhuis, S.; Haverkamp, A.; De Vries, M.R.; Quax, P.H.A.; Garcia-Vallejo, J.J.; et al. Galectin-2 induces a proinflammatory, anti-arteriogenic phenotype in monocytes, macrophages. PLoS ONE 2015, 10, e0124347. [Google Scholar] [CrossRef]
- Arata, Y.; Hirabayashi, J.; Kasai, K.I. Sugar binding properties of the two lectin domains of the tandem repeat-type galectin lec-1 (n32) of caenorhabditis elegans. detailed analysis by an improved frontal affinity chromatography method. J. Biol. Chem. 2001, 276, 3068–3077. [Google Scholar] [CrossRef] [Green Version]
- Tamura, M.; Sasai, A.; Ozawa, R.; Saito, M.; Yamamoto, K.; Takeuchi, T.; Ohtake, K.; Tateno, H.; Hirabayashi, J.; Kobayashi, J.; et al. Identification of the cysteine residue responsible for oxidative inactivation of mouse galectin-2. J. Biochem. 2016, 160, 233–241. [Google Scholar] [CrossRef] [Green Version]
- Bergonzelli, G.E.; Granato, D.; Pridmore, R.D.; Marvin-Guy, L.F.; Donnicola, D.; Corthésy-Theulaz, I.E. GroEL of lactobacillus johnsonii la1 (ncc 533) is cell surface associated: Potential role in interactions with the host and the gastric pathogen helicobacter pylori. Infect. Immun. 2006, 74, 425–434. [Google Scholar] [CrossRef] [Green Version]
- Ensgraber, M.; Loos, M. A 66-kilodalton heat shock protein of salmonella typhimurium is responsible for binding of the bacterium to intestinal mucus. Infect. Immun. 1992, 60, 3072–3078. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ota, H.; Katsuyama, T. Alternating laminated array of two types of mucin in the human gastric surface mucous layer. Histochem. J. 1992, 24, 86–92. [Google Scholar] [CrossRef] [PubMed]






© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sasaki, T.; Saito, R.; Oyama, M.; Takeuchi, T.; Tanaka, T.; Natsume, H.; Tamura, M.; Arata, Y.; Hatanaka, T. Galectin-2 Has Bactericidal Effects against Helicobacter pylori in a β-galactoside-Dependent Manner. Int. J. Mol. Sci. 2020, 21, 2697. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21082697
Sasaki T, Saito R, Oyama M, Takeuchi T, Tanaka T, Natsume H, Tamura M, Arata Y, Hatanaka T. Galectin-2 Has Bactericidal Effects against Helicobacter pylori in a β-galactoside-Dependent Manner. International Journal of Molecular Sciences. 2020; 21(8):2697. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21082697
Chicago/Turabian StyleSasaki, Takaharu, Rei Saito, Midori Oyama, Tomoharu Takeuchi, Toru Tanaka, Hideshi Natsume, Mayumi Tamura, Yoichiro Arata, and Tomomi Hatanaka. 2020. "Galectin-2 Has Bactericidal Effects against Helicobacter pylori in a β-galactoside-Dependent Manner" International Journal of Molecular Sciences 21, no. 8: 2697. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21082697