Dextran Sodium Sulfate-Induced Impairment of Protein Trafficking and Alterations in Membrane Composition in Intestinal Caco-2 Cell Line

 , , and
, , and
Abstract
:1. Introduction
2. Results
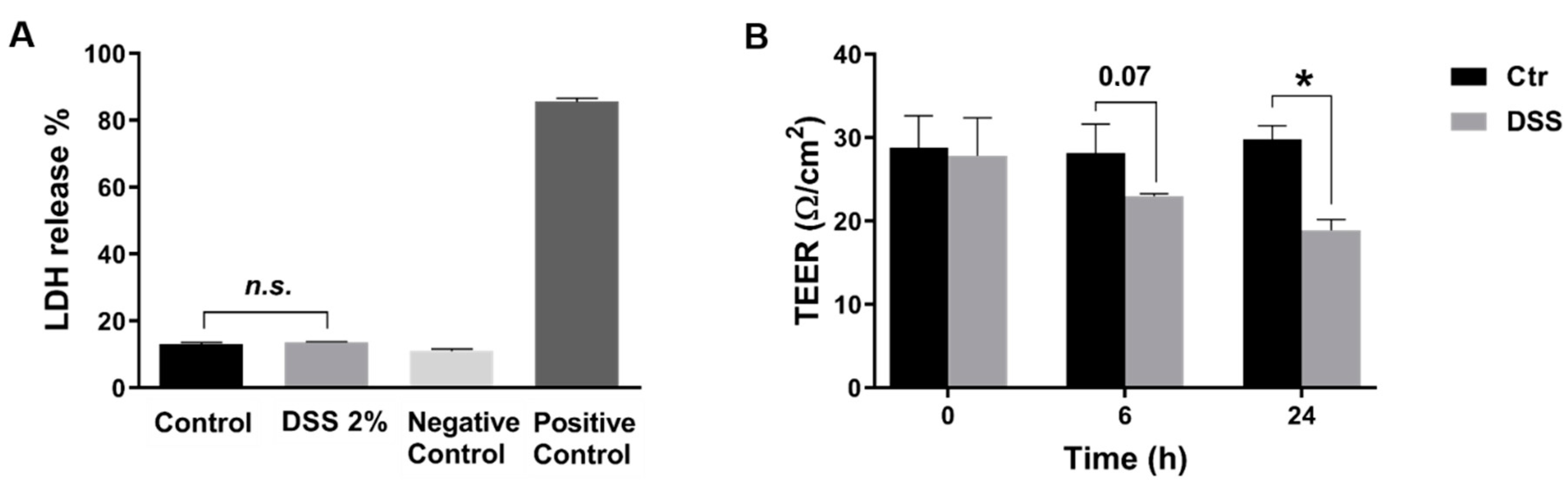
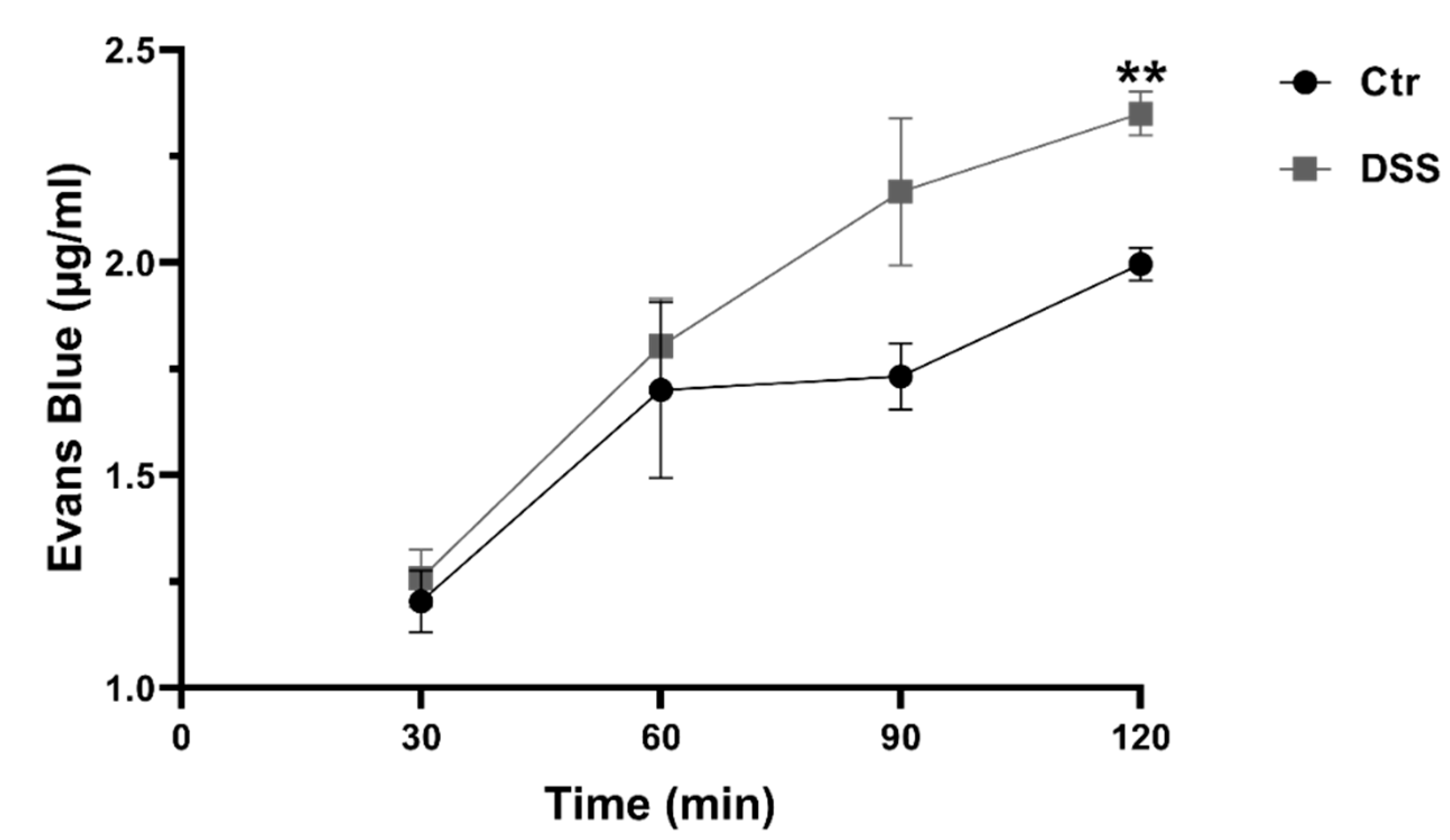
2.1. DSS Affects Membrane Integrity
2.2. DSS Induces ER Stress
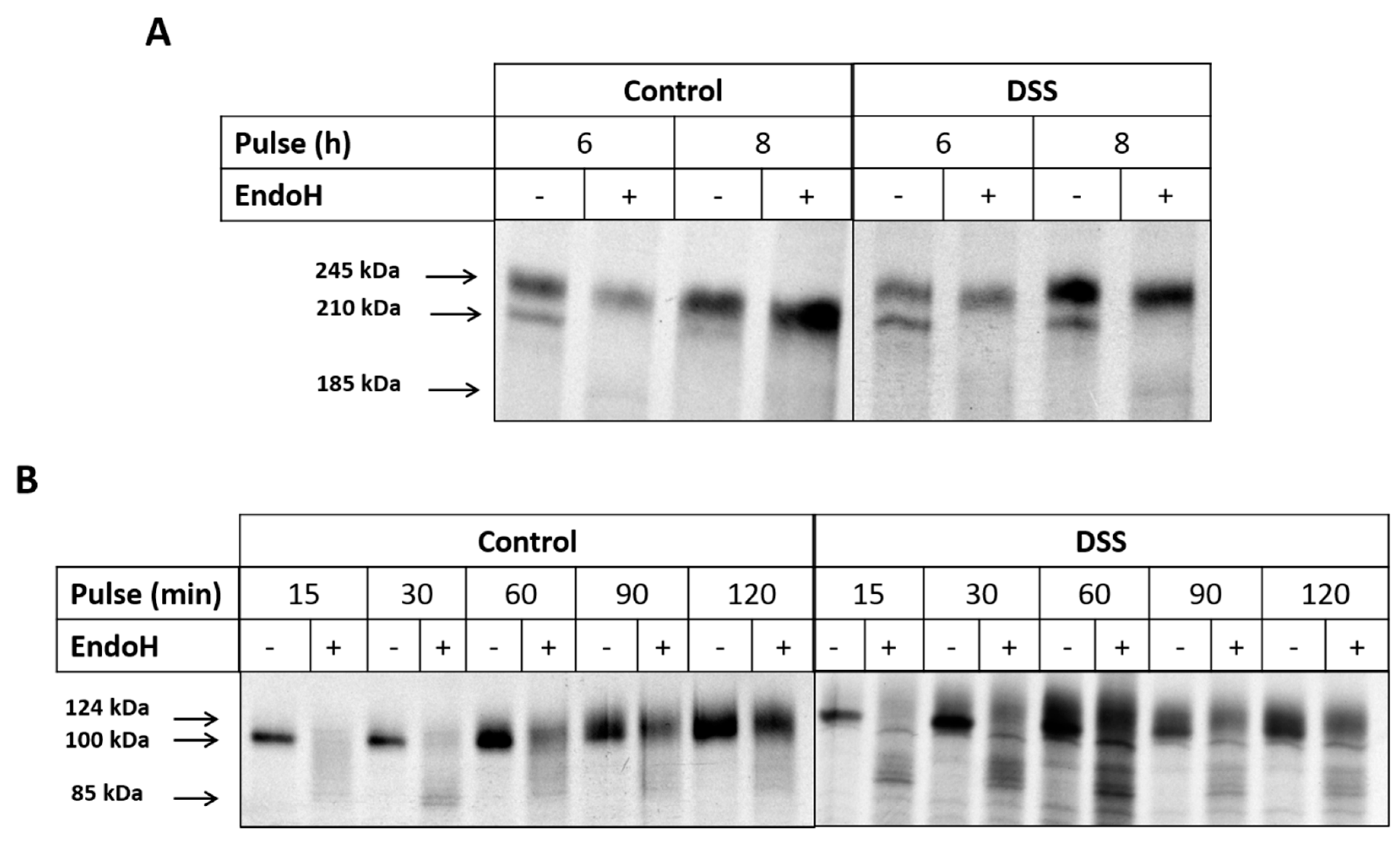
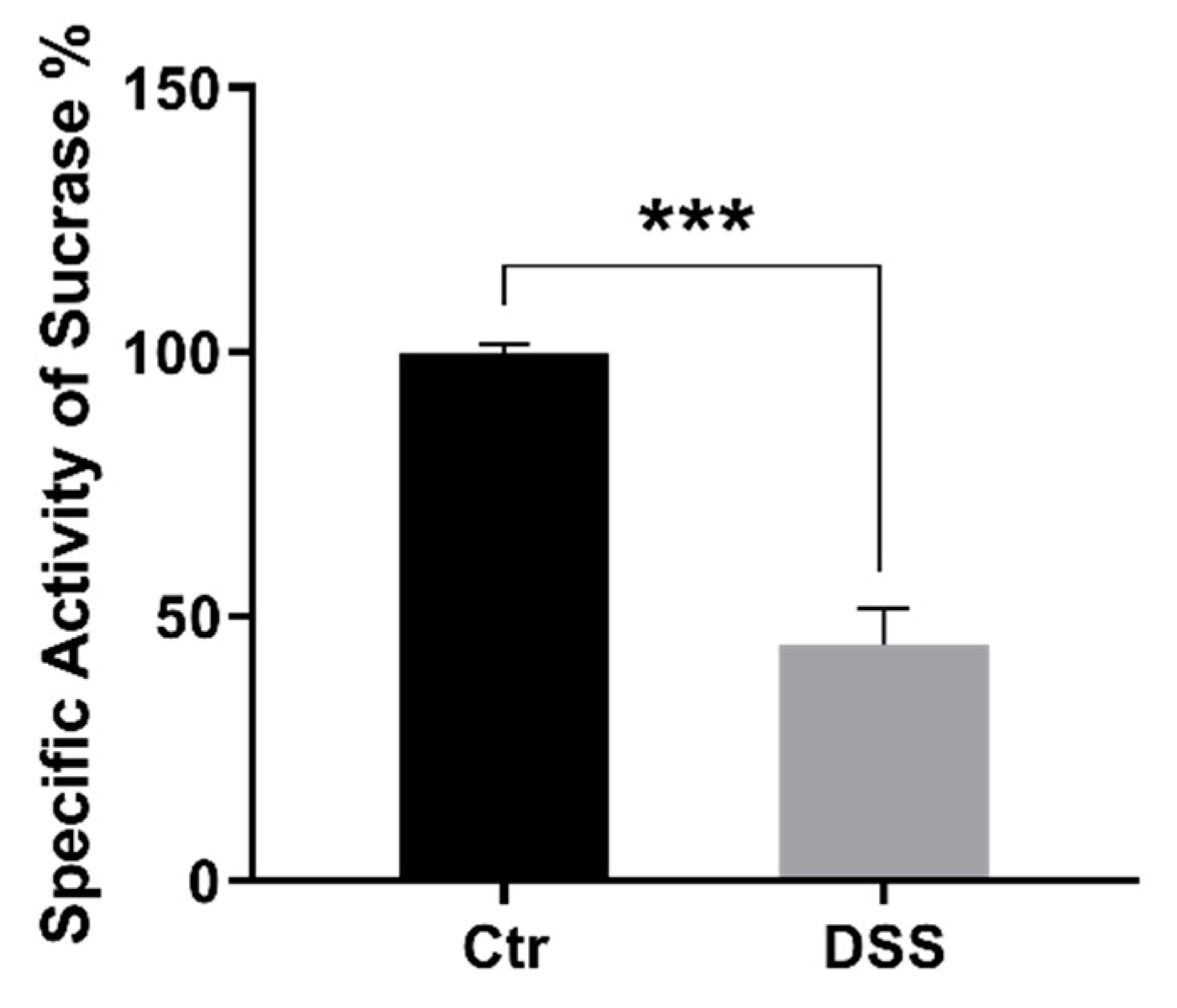
2.3. The Effect of DSS on Protein Trafficking
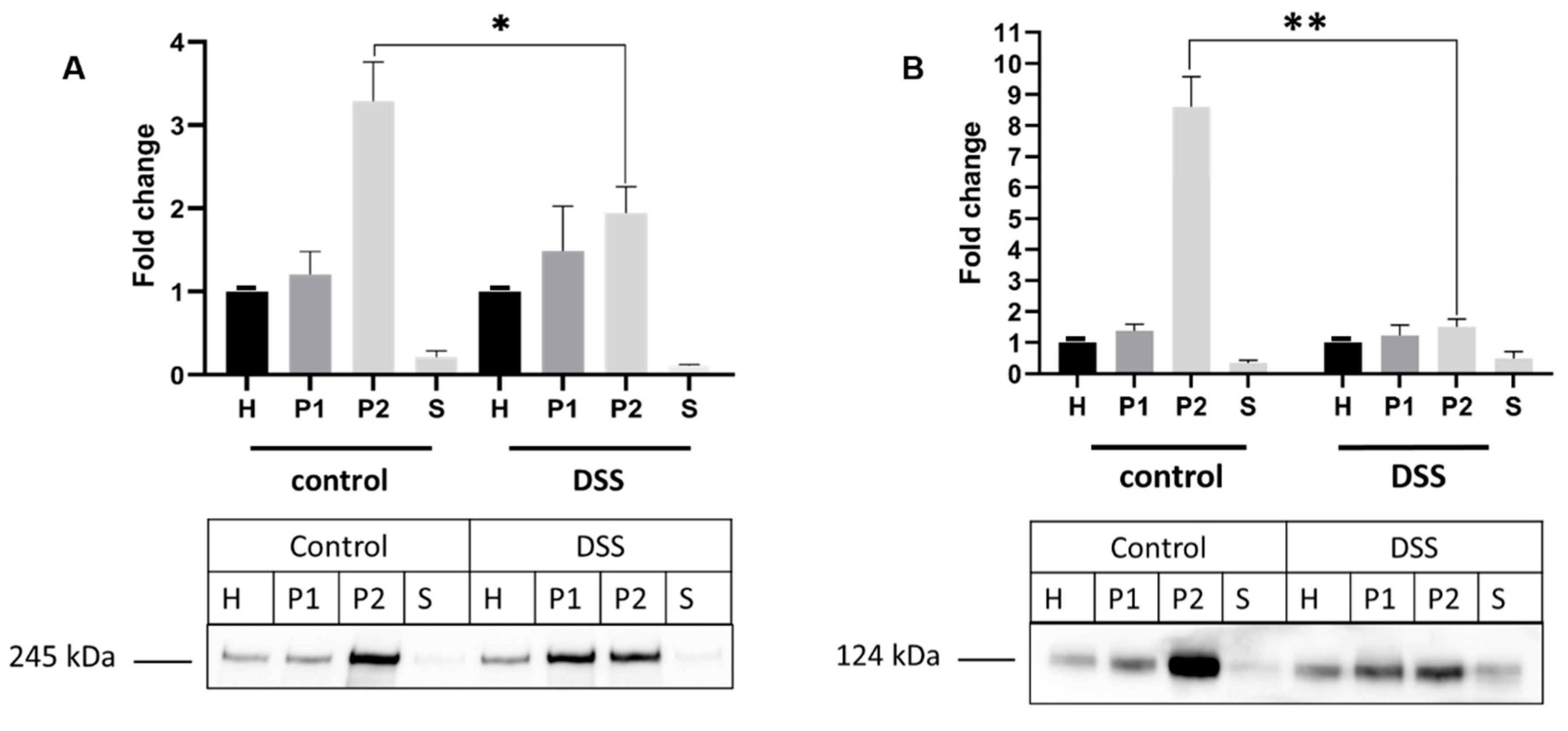
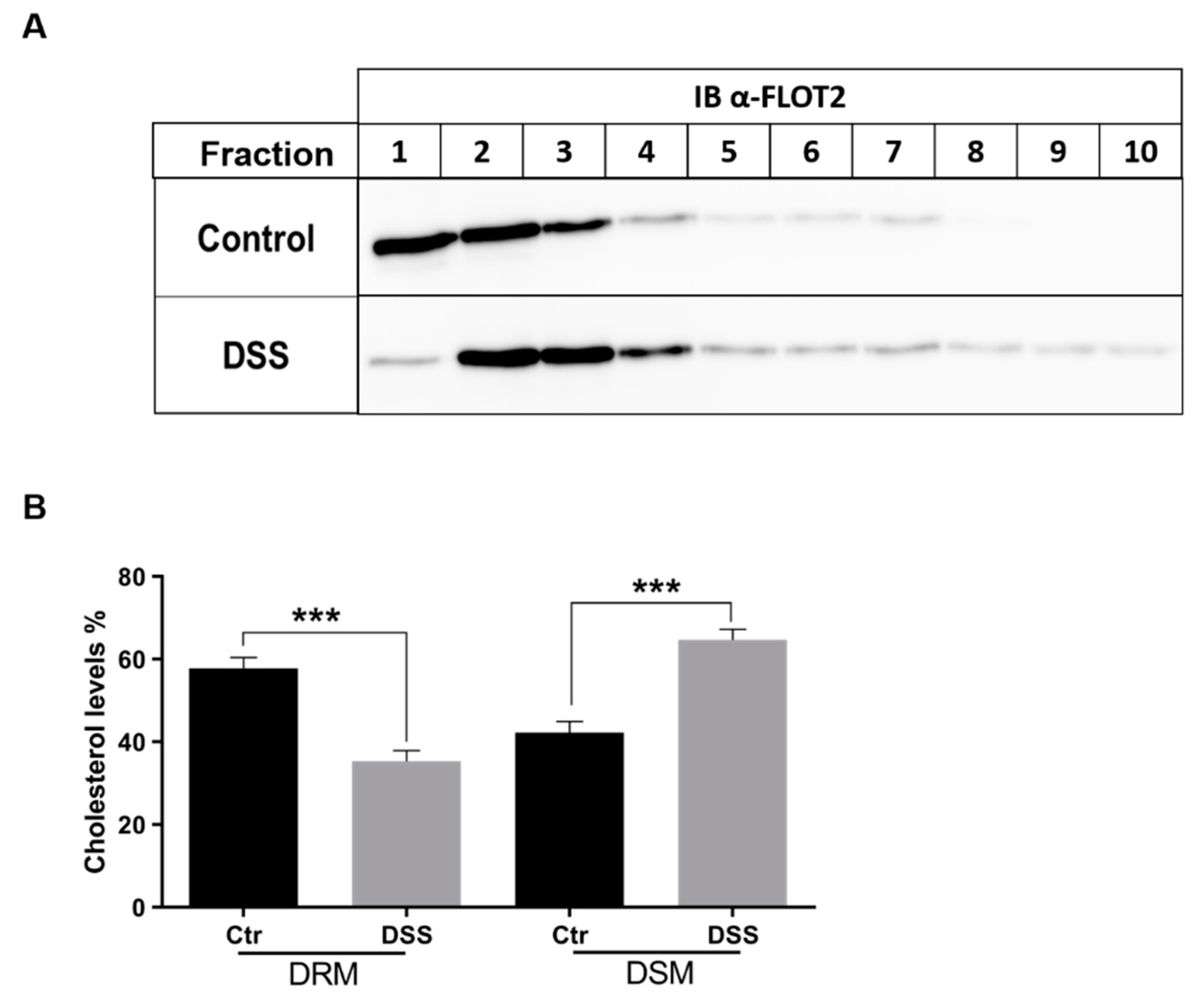
2.4. The Alteration of Membrane Composition Upon DSS Treatment
3. Discussion
4. Materials and Methods
4.1. Cell Culture
4.2. Measurement of Cell Death
4.3. Measurement of Trans-Epithelial Electrical Resistance
4.4. Evans Blue Permeability Assay (EBPA)
4.5. Total RNA Isolation and Reverse Transcription
4.6. Biosynthetic Labeling of Sucrase-Isomaltase and Dipeptidyl Peptidase-4
4.7. Brush Border Membranes Preparation
4.8. Detergent Resistant-Membrane Preparation
4.9. Lipid Extraction and Lipid Composition Analysis
4.10. Western Blotting
4.11. Statistical Analysis
5. Conclusion
Author Contributions
Funding
Conflicts of Interest
References
- Hollander, D. Crohn’s disease—A permeability disorder of the tight junction? Gut 1988, 29, 1621. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pearson, A.; Eastham, E.; Laker, M.; Craft, A.; Nelson, R. Intestinal permeability in children with Crohn’s disease and coeliac disease. Br. Med. J. (Clin. Res. Ed.) 1982, 285, 20–21. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Buhner, S.; Buning, C.; Genschel, J.; Kling, K.; Herrmann, D.; Dignass, A.; Kuechler, I.; Krueger, S.; Schmidt, H.H.; Lochs, H. Genetic basis for increased intestinal permeability in families with Crohn’s disease: Role of CARD15 3020insC mutation? Gut 2006, 55, 342–347. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ogura, Y.; Bonen, D.K.; Inohara, N.; Nicolae, D.L.; Chen, F.F.; Ramos, R.; Britton, H.; Moran, T.; Karaliuskas, R.; Duerr, R.H. A frameshift mutation in NOD2 associated with susceptibility to Crohn’s disease. Nature 2001, 411, 603–606. [Google Scholar] [CrossRef]
- Sengupta, P.; Baird, B.; Holowka, D. Lipid rafts, fluid/fluid phase separation, and their relevance to plasma membrane structure and function. Semin. Cell Dev. Biol. 2007, 18, 583–590. [Google Scholar] [CrossRef] [Green Version]
- Friedrichson, T.; Kurzchalia, T.V. Microdomains of GPI-anchored proteins in living cells revealed by crosslinking. Nature 1998, 394, 802–805. [Google Scholar] [CrossRef] [Green Version]
- Bowie, R.V.; Donatello, S.; Lyes, C.; Owens, M.B.; Babina, I.S.; Hudson, L.; Walsh, S.V.; O’Donoghue, D.P.; Amu, S.; Barry, S.P. Lipid rafts are disrupted in mildly inflamed intestinal microenvironments without overt disruption of the epithelial barrier. Am. J. Physiol. Gastrointest. Liver Physiol. 2012, 302, G781–G793. [Google Scholar] [CrossRef]
- Okamoto, R.; Watanabe, M. Role of epithelial cells in the pathogenesis and treatment of inflammatory bowel disease. J. Gastroenterol. 2016, 51, 11–21. [Google Scholar] [CrossRef] [Green Version]
- Kaser, A.; Lee, A.-H.; Franke, A.; Glickman, J.N.; Zeissig, S.; Tilg, H.; Nieuwenhuis, E.E.; Higgins, D.E.; Schreiber, S.; Glimcher, L.H. XBP1 links ER stress to intestinal inflammation and confers genetic risk for human inflammatory bowel disease. Cell 2008, 134, 743–756. [Google Scholar] [CrossRef] [Green Version]
- Bogaert, S.; De Vos, M.; Olievier, K.; Peeters, H.; Elewaut, D.; Lambrecht, B.; Pouliot, P.; Laukens, D. Involvement of endoplasmic reticulum stress in inflammatory bowel disease: A different implication for colonic and ileal disease? PLoS ONE 2011, 6, e25589. [Google Scholar] [CrossRef] [Green Version]
- Navid, F.; Colbert, R.A. Causes and consequences of endoplasmic reticulum stress in rheumatic disease. Nat. Rev. Rheumatol. 2017, 13, 25. [Google Scholar] [CrossRef] [PubMed]
- Dieleman, L.A.; Ridwan, B.U.; Tennyson, G.S.; Beagley, K.W.; Bucy, R.P.; Elson, C.O. Dextran sulfate sodium-induced colitis occurs in severe combined immunodeficient mice. Gastroenterology 1994, 107, 1643–1652. [Google Scholar] [CrossRef]
- KITAJIMA, S.; TAKUMA, S.; MORIMOTO, M. Tissue distribution of dextran sulfate sodium (DSS) in the acute phase of murine DSS-induced colitis. J. Vet. Med. Sci. 1999, 61, 67–70. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sambuy, Y.; De Angelis, I.; Ranaldi, G.; Scarino, M.; Stammati, A.; Zucco, F. The Caco-2 cell line as a model of the intestinal barrier: Influence of cell and culture-related factors on Caco-2 cell functional characteristics. Cell Biol. Toxicol. 2005, 21, 1–26. [Google Scholar] [CrossRef] [PubMed]
- Araki, Y.; Sugihara, H.; Hattori, T. In vitro effects of dextran sulfate sodium on a Caco-2 cell line and plausible mechanisms for dextran sulfate sodium-induced colitis. Oncol. Rep. 2006, 16, 1357–1362. [Google Scholar] [CrossRef] [PubMed]
- Naim, H.; Sterchi, E.; Lentze, M. Structure, biosynthesis, and glycosylation of human small intestinal maltase-glucoamylase. J. Biol. Chem. 1988, 263, 19709–19717. [Google Scholar]
- Hauri, H.P.; Sterchi, E.E.; Bienz, D.; Fransen, J.A.; Marxer, A. Expression and intracellular transport of microvillus membrane hydrolases in human intestinal epithelial cells. J. Cell Biol. 1985, 101, 838–851. [Google Scholar] [CrossRef] [Green Version]
- Browman, D.T.; Hoegg, M.B.; Robbins, S.M. The SPFH domain-containing proteins: More than lipid raft markers. Trends Cell Biol. 2007, 17, 394–402. [Google Scholar] [CrossRef]
- Langhorst, M.F.; Reuter, A.; Stuermer, C. Scaffolding microdomains and beyond: The function of reggie/flotillin proteins. Cell. Mol. Life Sci. CMLS 2005, 62, 2228–2240. [Google Scholar] [CrossRef] [Green Version]
- Danielsen, E.M.; De Haro Hernando, A.; Yassin, M.; Rasmussen, K.; Olsen, J. Short-term tissue permeability actions of dextran sulfate sodium studied in a colon organ culture system. Tissue Barriers 2020. [Google Scholar] [CrossRef] [Green Version]
- Hino, K.; Saito, A.; Asada, R.; Kanemoto, S.; Imaizumi, K. Increased susceptibility to dextran sulfate sodium-induced colitis in the endoplasmic reticulum stress transducer OASIS deficient mice. PLoS ONE 2014, 9, e88048. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pandurangan, A.K.; Ismail, S.; Saadatdoust, Z.; Esa, N.M. Allicin alleviates dextran sodium sulfate-(DSS-) induced ulcerative colitis in BALB/c mice. Oxidative Med. Cell. Longev. 2015, 2015, 605208. [Google Scholar] [CrossRef] [PubMed]
- Mankertz, J.; Schulzke, J.-D. Altered permeability in inflammatory bowel disease: Pathophysiology and clinical implications. Curr. Opin. Gastroenterol. 2007, 23, 379–383. [Google Scholar] [CrossRef] [PubMed]
- Casellas, F.; Aguade, S.; Soriano, B.; Accarino, A.; Molero, J.; Guarner, L. Intestinal Permeability to 99m Tc--Diethylenetriaminopentaacetic Acid in Inflammatory Bowel Disease. Am. J. Gastroenterol. 1986, 81, 767–770. [Google Scholar] [PubMed]
- Fries, W.; Muja, C.; Crisafulli, C.; Cuzzocrea, S.; Mazzon, E. Dynamics of enterocyte tight junctions: Effect of experimental colitis and two different anti-TNF strategies. Am. J. Physiol. Gastrointest. Liver Physiol. 2008, 294, G938–G947. [Google Scholar] [CrossRef] [PubMed]
- Clayburgh, D.R.; Shen, L.; Turner, J.R. A porous defense: The leaky epithelial barrier in intestinal disease. Lab. Investig. 2004, 84, 282–291. [Google Scholar] [CrossRef] [Green Version]
- Peterson, L.W.; Artis, D. Intestinal epithelial cells: Regulators of barrier function and immune homeostasis. Nat. Rev. Immunol. 2014, 14, 141–153. [Google Scholar] [CrossRef]
- Vitale, S.; Strisciuglio, C.; Pisapia, L.; Miele, E.; Barba, P.; Vitale, A.; Cenni, S.; Bassi, V.; Maglio, M.; Del Pozzo, G. Cytokine production profile in intestinal mucosa of paediatric inflammatory bowel disease. PLoS ONE 2017, 12, e0182313. [Google Scholar] [CrossRef]
- Ron, D.; Walter, P. Signal integration in the endoplasmic reticulum unfolded protein response. Nat. Rev. Mol. Cell Biol. 2007, 8, 519–529. [Google Scholar] [CrossRef]
- Alfalah, M.; Wetzel, G.; Fischer, I.; Busche, R.; Sterchi, E.E.; Zimmer, K.-P.; Sallmann, H.-P.; Naim, H.Y. A novel type of detergent-resistant membranes may contribute to an early protein sorting event in epithelial cells. J. Biol. Chem. 2005, 280, 42636–42643. [Google Scholar] [CrossRef] [Green Version]
- Jacob, R.; Heine, M.; Alfalah, M.; Naim, H.Y. Distinct cytoskeletal tracks direct individual vesicle populations to the apical membrane of epithelial cells. Curr. Biol. 2003, 13, 607–612. [Google Scholar] [CrossRef] [Green Version]
- Ridsdale, A.; Denis, M.; Gougeon, P.-Y.; Ngsee, J.K.; Presley, J.F.; Zha, X. Cholesterol is required for efficient endoplasmic reticulum-to-Golgi transport of secretory membrane proteins. Mol. Biol. Cell 2006, 17, 1593–1605. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wetzel, G.; Heine, M.; Rohwedder, A.; Naim, H.Y. Impact of glycosylation and detergent-resistant membranes on the function of intestinal sucrase-isomaltase. Biol. Chem. 2009, 390, 545–549. [Google Scholar] [CrossRef]
- Chen, Y.; Gao, H.; Yin, Q.; Chen, L.; Dong, P.; Zhang, X.; Kang, J. ER stress activating ATF4/CHOP-TNF-α signaling pathway contributes to alcohol-induced disruption of osteogenic lineage of multipotential mesenchymal stem cell. Cell. Physiol. Biochem. 2013, 32, 743–754. [Google Scholar] [CrossRef] [PubMed]
- Denis, R.; Arruda, A.; Romanatto, T.; Milanski, M.; Coope, A.; Solon, C.; Razolli, D.; Velloso, L. TNF-α transiently induces endoplasmic reticulum stress and an incomplete unfolded protein response in the hypothalamus. Neuroscience 2010, 170, 1035–1044. [Google Scholar] [CrossRef]
- Marone, M.; Mozzetti, S.; De Ritis, D.; Pierelli, L.; Scambia, G. Semiquantitative RT-PCR analysis to assess the expression levels of multiple transcripts from the same sample. Biol. Proced. Online 2001, 3, 19–25. [Google Scholar] [CrossRef] [Green Version]
- Naim, H.; Sterchi, E.; Lentze, M. Biosynthesis and maturation of lactase-phlorizin hydrolase in the human small intestinal epithelial cells. Biochem. J. 1987, 241, 427–434. [Google Scholar] [CrossRef] [Green Version]
- Beaulieu, J.-F.; Nichols, B.; Quaroni, A. Posttranslational regulation of sucrase-isomaltase expression in intestinal crypt and villus cells. J. Biol. Chem. 1989, 264, 20000–20011. [Google Scholar]
- Shimada, Y.; Inomata, M.; Suzuki, H.; Hayashi, M.; Abdul Waheed, A.; Ohno-Iwashita, Y. Separation of a cholesterol-enriched microdomain involved in T-cell signal transduction. FEBS J. 2005, 272, 5454–5463. [Google Scholar] [CrossRef]
- Bligh, E.G.; Dyer, W.J. A rapid method of total lipid extraction and purification. Can. J. Biochem. Physiol. 1959, 37, 911–917. [Google Scholar] [CrossRef] [Green Version]
- Brogden, G.; Propsting, M.; Adamek, M.; Naim, H.Y.; Steinhagen, D. Isolation and analysis of membrane lipids and lipid rafts in common carp (Cyprinus carpio L.). Comp. Biochem. Physiol. Part B Biochem. Mol. Biol. 2014, 169, 9–15. [Google Scholar] [CrossRef]
- Branitzki-Heinemann, K.; Möllerherm, H.; Völlger, L.; Husein, D.M.; de Buhr, N.; Blodkamp, S.; Reuner, F.; Brogden, G.; Naim, H.Y.; von Köckritz-Blickwede, M. Formation of neutrophil extracellular traps under low oxygen level. Front. Immunol. 2016, 7, 518. [Google Scholar] [CrossRef] [Green Version]
- Brogden, G.; Shammas, H.; Maalouf, K.; Naim, S.L.; Wetzel, G.; Amiri, M.; von Köckritz-Blickwede, M.; Das, A.M.; Naim, H.Y. Case study on the pathophysiology of Fabry disease: Abnormalities of cellular membranes can be reversed by substrate reduction in vitro. Biosci. Rep. 2017, 37, BSR20160402. [Google Scholar] [CrossRef]
- Hodroj, M.H.; Jardaly, A.; Abi Raad, S.; Zouein, A.; Rizk, S. Andrographolide potentiates the antitumor effect of topotecan in acute myeloid leukemia cells through an intrinsic apoptotic pathway. Cancer Manag. Res. 2018, 10, 1079–1088. [Google Scholar] [CrossRef] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer | Sequence |
|---|---|
| XBP1 fw XBP1rev | TGGCCGGGTCTGCTGAGTCCG ATCCATGGGGAGATGTTCTGG |
| ATF4 fw ATF4 rev | GTTCTCCAGCGACAAGGCTA ATCCTGCTTGCTGTTGTTGG |
| CHOP fw CHOP rev | AGAACCAGGAAACGGAAACAGA TCTCCTTCATGCGCTGCTTT |
| BiP fw BiP rev | TGTTCAACCAATTATCAGCAAACTC TTCTGCTGTATCCTCTTCACCAGT |
| TNF-α fw TNF-α rev | CTTCTGCCTGCTGCACTTTG AGCTGCCCCTCAGCTTGAG |
| IL-1 α fw IL-1 α rev | CTGAAGAAGAGACGGTTGAGTT AGGGCGTCATTCAGGATGAA |
| IL-6 fw IL-6 rev | GTAGCCGCCCCACACAGA AGCCATCTTTGGAAGGTTCAGG |
| IL-10 fw IL-10 rev | GCCTTGTCTGAGATGATCCAGT CCACGGCCTTGCTCTTGTT |
| GAPDH Fw GAPDH rev | CATGGCCTTCCGTGTTCCTA CCTGCTTCACCACCTTCTTGAT |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Toutounji, M.; Wanes, D.; El-Harakeh, M.; El-Sabban, M.; Rizk, S.; Naim, H.Y. Dextran Sodium Sulfate-Induced Impairment of Protein Trafficking and Alterations in Membrane Composition in Intestinal Caco-2 Cell Line. Int. J. Mol. Sci. 2020, 21, 2726. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21082726
Toutounji M, Wanes D, El-Harakeh M, El-Sabban M, Rizk S, Naim HY. Dextran Sodium Sulfate-Induced Impairment of Protein Trafficking and Alterations in Membrane Composition in Intestinal Caco-2 Cell Line. International Journal of Molecular Sciences. 2020; 21(8):2726. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21082726
Chicago/Turabian StyleToutounji, Mohamad, Dalanda Wanes, Mohammad El-Harakeh, Marwan El-Sabban, Sandra Rizk, and Hassan Y. Naim. 2020. "Dextran Sodium Sulfate-Induced Impairment of Protein Trafficking and Alterations in Membrane Composition in Intestinal Caco-2 Cell Line" International Journal of Molecular Sciences 21, no. 8: 2726. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21082726