Transcriptional and Metabolic Changes Associated with Phytoglobin Expression during Germination of Barley Seeds


Abstract
:1. Introduction
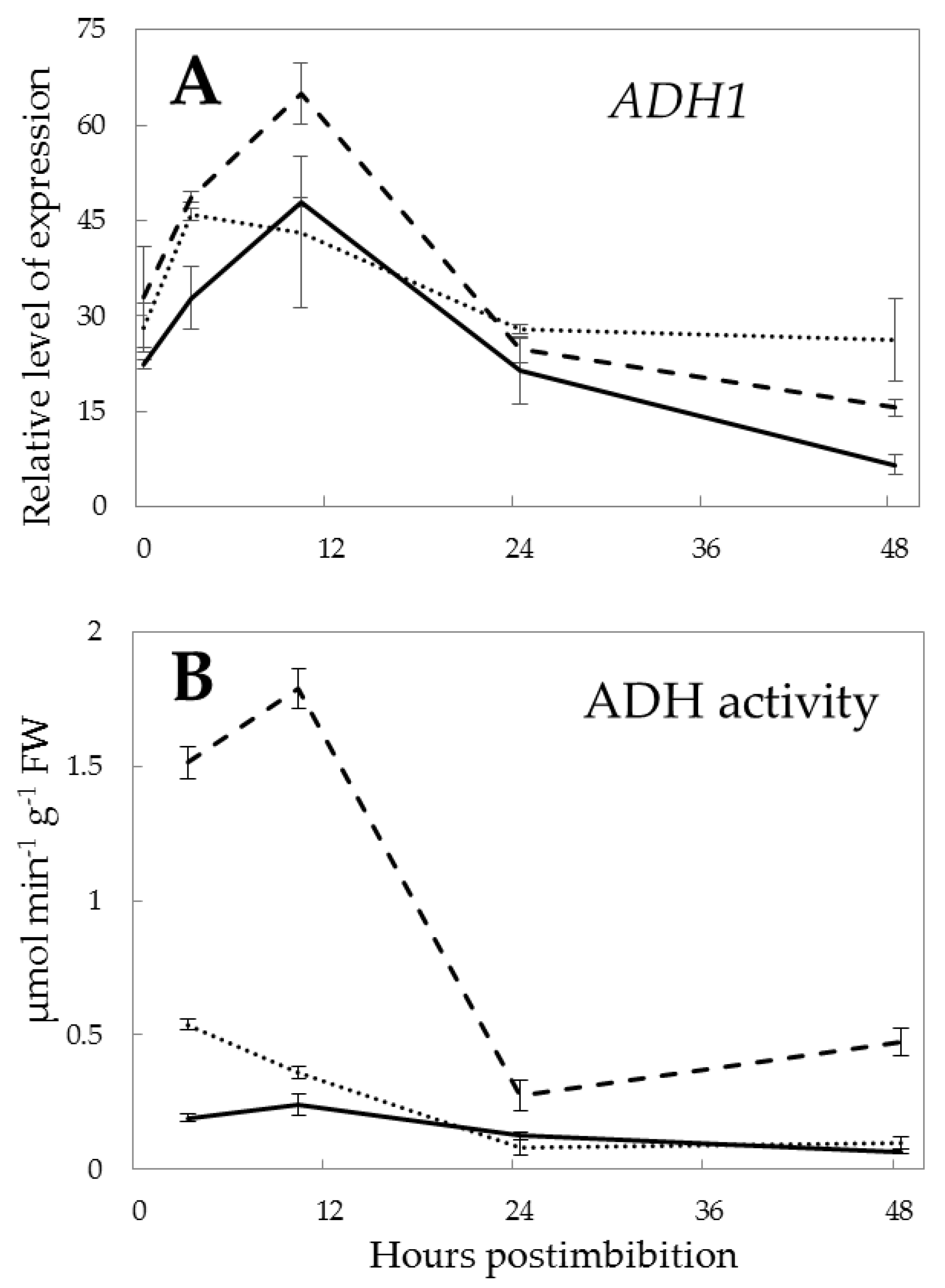
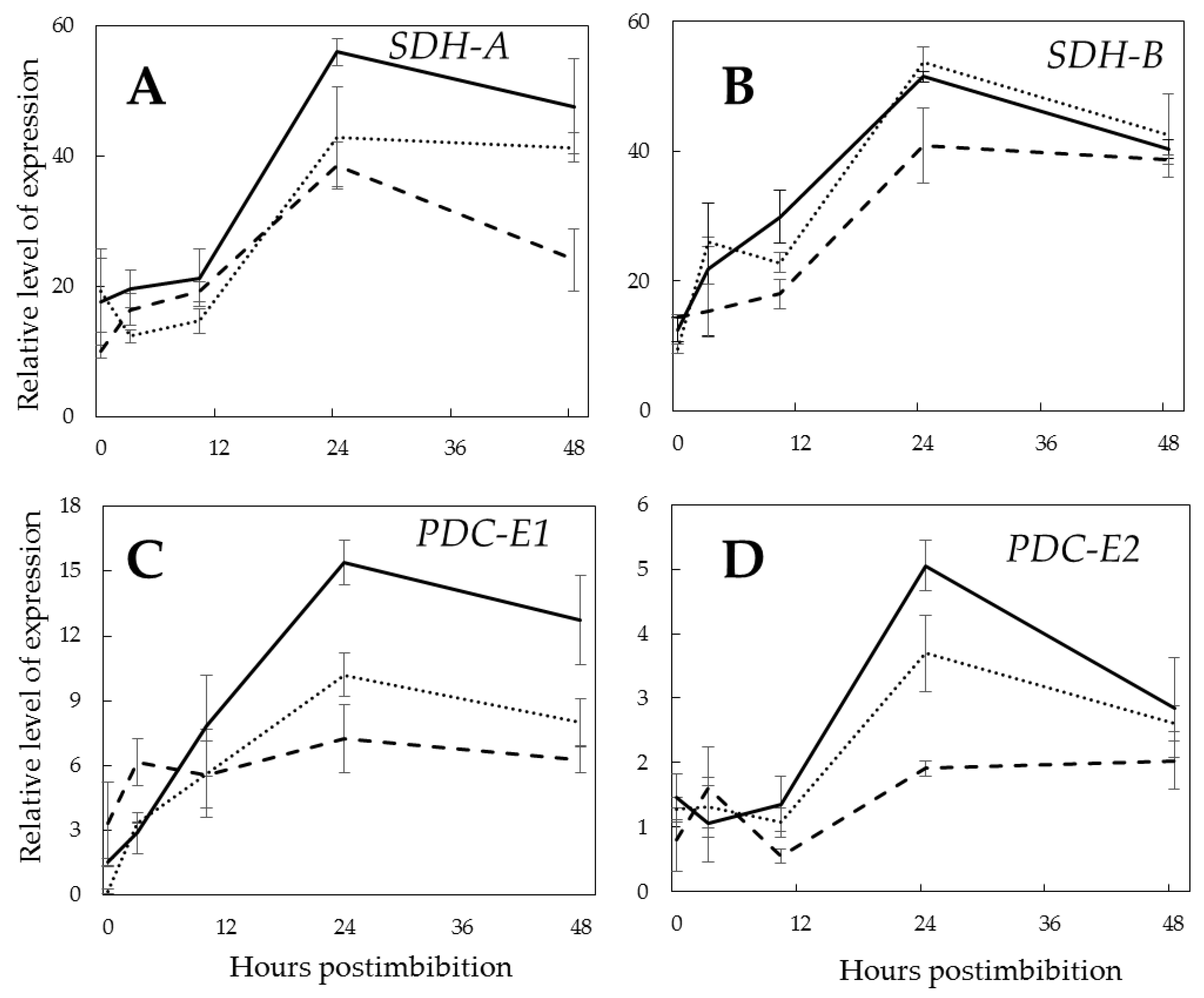
2. Results
3. Discussion
3.1. Anaerobic Conditions in Germinating Seeds and Expression Of Class 1 Phytoglobin
3.2. Energy Production During Seed Germination and the Role of NO
3.3. Fermentation and the Pgb-NO Cycle
3.4. S-Nitrosoglutathione Reductase
3.5. Pgb1 and Operation of the TCA Cycle
3.6. Pgb and the Non-Coupled Respiration
4. Materials and Methods
4.1. Plant Growth
4.2. ATP/ADP Ratio
4.3. NO Emission
4.4. Measurement of Protein Concentration
4.5. Total Protein S-Nitrosylation
4.6. Enzymatic Activity Assays
4.7. Gene Expression
4.8. Statistical Analysis
5. Conclusions
Author Contributions
Acknowledgments
Conflicts of Interest
Abbreviations
| ADH | Alcohol dehydrogenase |
| AOX | Alternative oxidase |
| DTNB | 5,5′-dithiol-bis (2-nitrobenzoic acid) |
| ETC | Electron transport chain |
| GAPDH | Glyceraldehyde 3-phosphate dehydrogenase |
| GSNO | S-nitrosoglutathione |
| GSNOR | S-nitrosoglutathione reductase |
| NDs | type II NAD(P)H dehydrogenases |
| NiR | Nitrite reductase |
| NO | Nitric oxide |
| NR | Nitrate reductase |
| PDC | Pyruvate dehydrogenase complex |
| Pgb | Phytoglobin |
| RNS | Reactive species of nitrogen |
| ROS | Reactive species of oxygen |
| SDH | Succinate dehydrogenase |
| TCA | Tricarboxylic acid |
References
- Qiu, J.; Bai, Y.; Fu, Y.B.; Wilmshurst, J.F. Spatial variation in temperature thresholds during seed germination of remnant Festuca hallii populations across the Canadian prairie. Environ. Exp. Bot. 2010, 67, 479–548. [Google Scholar] [CrossRef]
- Steinbrecher, T.; Leubner-Metzger, G. The biomechanics of seed germination. J. Exp. Bot. 2017, 68, 765–783. [Google Scholar] [CrossRef]
- Bykova, N.V.; Hu, J.; Ma, Z.; Igamberdiev, A.U. The role of reactive oxygen and nitrogen species in bioenergetics, metabolism and signaling during seed germination. In Reactive Oxygen and Reactive Nitrogen Species Signaling and Communication in Plants; Gupta, K.J., Igamberdiev, A.U., Eds.; Springer: Berlin, Germany, 2015; pp. 177–195. [Google Scholar] [CrossRef]
- Bethke, P.C. Rebirth and death: Nitric oxide and reactive oxygen species in seeds. SEB Exp. Biol. Ser. 2009, 62, 17–30. [Google Scholar] [CrossRef]
- Ma, Z.; Marsolais, F.; Bykova, N.V.; Igamberdiev, A.U. Nitric oxide and reactive oxygen species mediate metabolic changes in barley seed embryo during germination. Front. Plant. Sci. 2016, 7, 138. [Google Scholar] [CrossRef] [Green Version]
- Igamberdiev, A.U.; Hill, R.D. Nitrate, NO and haemoglobin in plant adaptation to hypoxia: An alternative to classic fermentation pathways. J. Exp. Bot. 2004, 55, 2473–2482. [Google Scholar] [CrossRef] [Green Version]
- Gupta, K.J.; Hebelstrup, K.H.; Mur, L.A.; Igamberdiev, A.U. Plant hemoglobins: Important players at the crossroads between oxygen and nitric oxide. FEBS Lett. 2011, 585, 3843–3849. [Google Scholar] [CrossRef]
- Stoimenova, M.; Igamberdiev, A.U.; Gupta, K.J.; Hill, R.D. Nitrite-driven anaerobic ATP synthesis in barley and rice root mitochondria. Planta 2007, 226, 465–474. [Google Scholar] [CrossRef]
- Sakamoto, A.; Ueda, M.; Morikawa, H. Arabidopsis glutathione-dependent formaldehyde dehydrogenase is an S-nitrosoglutathione reductase. FEBS Lett. 2002, 515, 20–24. [Google Scholar] [CrossRef] [Green Version]
- Kwon, E.; Feechan, A.; Yun, B.-W.; Hwang, B.-H.; Pallas, J.A.; Kang, J.-G.; Loake, G.J. AtGSNOR1 function is required for multiple developmental programs in Arabidopsis. Planta 2012, 236, 887–900. [Google Scholar] [CrossRef] [PubMed]
- Luo, T.; Xian, M.; Zhang, C.; Zhang, C.; Hu, L.; Xu, Z. Associating transcriptional regulation for rapid germination of rapeseed (Brassica napus L.) under low temperature stress through weighted gene co-expression network analysis. Sci. Rep. 2019, 9, 55. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez, J.L.; De Diego, J.G.; Rodríguez, F.D.; Cervantes, E. Mitochondrial structures during seed germination and early seedling development in Arabidopsis thaliana. Biologia 2015, 70, 1019–1025. [Google Scholar] [CrossRef]
- Fernie, A.R.; Carrari, F.; Sweetlove, L.J. Respiratory metabolism: Glycolysis, the TCA cycle and mitochondrial electron transport. Curr. Opin. Plant Biol. 2004, 7, 254–261. [Google Scholar] [CrossRef] [PubMed]
- Eprintsev, A.T.; Fedorin, D.N.; Karabutova, L.A.; Igamberdiev, A.U. Expression of genes encoding subunits A and B of succinate dehydrogenase in germinating maize seeds is regulated by methylation of their promoters. J. Plant Physiol. 2016, 205, 33–40. [Google Scholar] [CrossRef] [PubMed]
- Millar, A.H.; Day, D.A. Nitric oxide inhibits the cytochrome oxidase but not the alternative oxidase of plant mitochondria. FEBS Lett. 1996, 39, 155–158. [Google Scholar] [CrossRef] [Green Version]
- Clerc, P.; Rigoulet, M.; Leverve, X.; Fontaine, E. Nitric oxide increases oxidative phosphorylation efficiency. J. Bioenerg. Biomembr. 2007, 39, 158–166. [Google Scholar] [CrossRef]
- Millar, A.H.; Whelan, J.; Soole, K.L.; Day, D.A. Organization and regulation of mitochondrial respiration in plants. Annu. Rev. Plant Biol. 2011, 62, 79–104. [Google Scholar] [CrossRef]
- Wanniarachchi, V.R.; Dametto, L.; Sweetman, C.; Shavrukov, Y.; Day, D.A.; Jenkins, C.L.D.; Soole, K.L. Alternative respiratory pathway component genes (AOX and ND) in rice and barley and their response to stress. Int. J. Mol. Sci. 2018, 19, 915. [Google Scholar] [CrossRef] [Green Version]
- Vanlerberghe, G.C. Alternative oxidase: A mitochondrial respiratory pathway to maintain metabolic and signaling homeostasis during abiotic and biotic stress in plants. Int. J. Mol. Sci. 2013, 14, 6805–6847. [Google Scholar] [CrossRef]
- Daley, D.O.; Considine, M.J.; Howell, K.A.; Millar, A.H.; Day, D.A.; Whelan, J. Respiratory gene expression in soybean cotyledons during post-germinative development. Plant Mol. Biol. 2003, 51, 745–755. [Google Scholar] [CrossRef]
- Igamberdiev, A.U.; Bykova, N.V.; Shah, J.K.; Hill, R.D. Anoxic nitric oxide cycling in plants: Participating reactions and possible mechanisms. Physiol. Plant. 2010, 138, 393–404. [Google Scholar] [CrossRef]
- Rosental, L.; Nonogaki, H.; Fait, A. Activation and regulation of primary metabolism during seed germination. Seed Sci. Res. 2014, 24, 1–15. [Google Scholar] [CrossRef]
- Benamar, A.; Tallon, C.; Macherel, D. Membrane integrity and oxidative properties of mitochondria isolated from imbibing pea seeds after priming or accelerated ageing. Seed Sci. Res. 2003, 13, 35–45. [Google Scholar] [CrossRef]
- Ismail, A.M.; Ella, E.S.; Vergara, G.V.; Mackill, D.J. Mechanisms associated with tolerance to flooding during germination and early seedling growth in rice (Oryza sativa). Ann. Bot. 2009, 103, 197–209. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Igamberdiev, A.U.; Baron, K.; Manac’h-Little, N.; Stoimenova, M.; Hill, R.D. The haemoglobin/nitric oxide cycle: Involvement in flooding stress and effects on hormone signalling. Ann. Bot. 2005, 96, 557–564. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bewley, J.D.; Black, M. Seeds: Physiology of Development and Germination, 2nd ed.; Springer: Berlin, Germany, 1994. [Google Scholar]
- Kennedy, R.A.; Rumpho, M.E.; Fox, T.C. Anaerobic metabolism in plants. Plant Physiol. 1992, 100, 1–6. [Google Scholar] [CrossRef] [Green Version]
- Andrzejczak, O.A.; Havelund, J.F.; Wang, W.Q.; Kovalchuk, S.; Hagensen, C.E.; Hasler-Sheetal, H.; Jensen, O.N.; Rogowska-Wrzesinska, A.; Møller, I.M.; Hebelstrup, K.H. The hypoxic proteome and metabolome of barley (Hordeum vulgare L.) with and without phytoglobin priming. Int. J. Mol. Sci. 2020, 21, 1546. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Holman, T.J.; Jones, P.D.; Russell, L.; Medhurst, A.; Tomás, S.Ú.; Talloji, P.; Marquez, J.; Schmuths, H.; Tung, S.-A.; Taylor, I.; et al. The N-end rule pathway promotes seed germination and establishment through removal of ABA sensitivity in Arabidopsis. Proc. Natl. Acad. Sci. USA 2009, 106, 4549–4554. [Google Scholar] [CrossRef] [Green Version]
- Angelovici, R.; Fait, A.; Fernie, A.R.; Galili, G. A seed high-lysine trait is negatively associated with the TCA cycle and slows down Arabidopsis seed germination. New Phytol. 2011, 189, 148–159. [Google Scholar] [CrossRef]
- Hebelstrup, K.H.; Igamberdiev, A.U.; Hill, R.D. Metabolic effects of hemoglobin gene expression in plants. Gene 2007, 398, 86–93. [Google Scholar] [CrossRef]
- Sowa, A.W.; Duff, S.M.; Guy, P.A.; Hill, R.D. Altering hemoglobin levels changes energy status in maize cells under hypoxia. Proc. Natl. Acad. Sci. USA 1998, 95, 10317–10321. [Google Scholar] [CrossRef] [Green Version]
- Igamberdiev, A.U.; Serege’lyes, C.; Manac’h, N.; Hill, R.D. NADH dependent metabolism of nitric oxide in alfalfa root cultures expressing barley hemoglobin. Planta 2004, 219, 95–102. [Google Scholar] [CrossRef] [PubMed]
- Cochrane, D.W.; Shah, J.K.; Hebelstrup, K.H.; Igamberdiev, A.U. Expression of phytoglobin affects nitric oxide metabolism and energy state of barley plants exposed to anoxia. Plant Sci. 2017, 265, 124–130. [Google Scholar] [CrossRef] [PubMed]
- Sreenivasulu, N.; Usadel, B.; Winter, A.; Radchuk, V.; Scholz, U.; Stein, N.; Weschke, W.; Strickert, M.; Close, T.J.; Stitt, M.; et al. Barley grain maturation and germination: Metabolic pathway and regulatory network commonalities and differences highlighted by new MapMan/PageMan profiling tools. Plant Physiol. 2008, 146, 1738–1758. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weitbrecht, K.; Müller, K.; Leubner-Metzger, G. First off the mark: Early seed germination. J. Exp. Bot. 2011, 62, 3289–3309. [Google Scholar] [CrossRef] [Green Version]
- Gupta, K.J.; Igamberdiev, A.U. The anoxic plant mitochondrion as a nitrite: NO reductase. Mitochondrion 2011, 11, 537–543. [Google Scholar] [CrossRef]
- Gupta, K.J.; Shah, J.K.; Brotman, Y.; Jahnke, K.; Willmitzer, L.; Kaiser, W.M.; Bauwe, H.; Igamberdiev, A.U. Inhibition of aconitase by nitric oxide leads to induction of the alternative oxidase and to a shift of metabolism towards biosynthesis of amino acids. J. Exp. Bot. 2012, 63, 1773–1784. [Google Scholar] [CrossRef] [Green Version]
- Corpas, F.J.; Alché, J.D.; Barroso, J.B. Current overview of S-nitrosoglutathione (GSNO) in higher plants. Front. Plant Sci. 2013, 4, 126. [Google Scholar] [CrossRef] [Green Version]
- Frungillo, L.; Skelly, M.J.; Loake, G.J.; Spoel, S.H.; Salgado, I. S-nitrosothiols regulate nitric oxide production and storage in plants through the nitrogen assimilation pathway. Nat. Commun. 2014, 5, 5401. [Google Scholar] [CrossRef] [Green Version]
- Bignon, E.; Allega, M.F.; Lucchetta, M.; Tiberti, M.; Papaleo, E. Computational structural biology of S-nitrosylation of cancer targets. Front. Oncol. 2018, 8, 272. [Google Scholar] [CrossRef] [Green Version]
- Botha, F.C.; Potgieter, G.P.; Botha, A-M. Respiratory metabolism and gene expression during seed germination. Plant Growth Regul. 1992, 11, 211–224. [Google Scholar] [CrossRef]
- Oyedotun, K.S.; Lemire, B.D. The quaternary structure of the Saccharomyces cerevisiae succinate dehydrogenase. Homology modeling, cofactor docking, and molecular dynamics simulation studies. J. Biol. Chem. 2004, 279, 9424–9431. [Google Scholar] [CrossRef] [Green Version]
- Eastmond, P.J.; Graham, I.A. Re-examining the role of the glyoxylate cycle in oilseeds. Trends Plant Sci. 2001, 6, 72–78. [Google Scholar] [CrossRef]
- Weraduwage, S.M.; Micallef, M.C.; Marillia, E.; Taylor, D.C.; Grodzinski, B.; Micallef, B.J. Increased mtpdh activity through antisense inhibition of mitochondrial pyruvate dehydrogenase kinase enhances inflorescence initiation, and inflorescence growth and harvest index at elevated CO2 in Arabidopsis thaliana. Front. Plant Sci. 2016, 7, 95. [Google Scholar] [CrossRef] [Green Version]
- Simonin, V.; Galina, A. Nitric oxide inhibits succinate dehydrogenase-driven oxygen consumption in potato tuber mitochondria in an oxygen tension-independent manner. Biochem. J. 2013, 449, 263–273. [Google Scholar] [CrossRef] [Green Version]
- Brown, G.C. Nitric oxide and mitochondrial respiration. Biochim. Biophys. Acta 1999, 1411, 351–369. [Google Scholar] [CrossRef] [Green Version]
- Pergo, É.M.; Ishii-Iwamoto, E.L. Changes in energy metabolism and antioxidant defense systems during seed germination of the weed species Ipomoea triloba L. and the responses to allelochemicals. J. Chem. Ecol. 2011, 37, 500–513. [Google Scholar] [CrossRef]
- Møller, I.M. Plant mitochondria and oxidative stress: Electron transport, NADPH turnover, and metabolism of reactive oxygen species. Annu. Rev. Plant Physiol. Plant Mol. Biol. 2001, 52, 561–591. [Google Scholar] [CrossRef] [Green Version]
- Velada, I.; Cardoso, H.G.; Ragonezi, C.; Nogales, A.; Ferreira, A.; Valadas, V.; Arnholdt-Schmitt, B. Alternative oxidase gene family in Hypericum perforatum L.: Characterization and expression at the post-germinative phase. Front. Plant Sci. 2016, 7, 1043. [Google Scholar] [CrossRef] [Green Version]
- De Oliveira, H.C.; Wulff, A.; Saviani, E.E.; Salgado, I. Nitric oxide degradation by potato tuber mitochondria: Evidence for the involvement of external NAD(P)H dehydrogenases. Biochim. Biophys. Acta 2008, 1777, 470–476. [Google Scholar] [CrossRef] [Green Version]
- Selinski, J.; Hartmann, A.; Deckers-Hebestreit, G.; Day, D.A.; Whelan, J.; Scheibea, R. Alternative oxidase isoforms are differentially activated by tricarboxylic acid cycle intermediates. Plant Physiol. 2018, 176, 1423–1432. [Google Scholar] [CrossRef] [Green Version]
- Hebelstrup, K.H.; Christiansen, M.W.; Carciofi, M.; Tauris, B.; Brinch-Pedersen, H.; Holm, P.B. UCE: A uracil excision (USER)-based toolbox for transformation of cereals. Plant Methods 2010, 6, 15. [Google Scholar] [CrossRef] [Green Version]
- Hebelstrup, K.H.; Shah, J.K.; Simpson, C.; Schjoerring, J.K.; Mandon, J.; Cristescu, S.M.; Harren, F.J.; Christiansen, M.W.; Mur, L.A.; Igamberdiev, A.U. An assessment of the biotechnological use of hemoglobin modulation in cereals. Physiol. Plant. 2014, 150, 593–603. [Google Scholar] [CrossRef]
- Dordas, C.; Hasinoff, B.B.; Igamberdiev, A.U.; Manac’h, N.; Rivoal, J.; Hill, R.D. Expression of a stress-induced hemoglobin affects NO levels produced by alfalfa root cultures under hypoxic stress. Plant J. 2003, 35, 763–770. [Google Scholar] [CrossRef]
- Planchet, E.; Gupta, K.J.; Sonoda, M.; Kaiser, W.M. Nitric oxide emission from tobacco leaves and cell suspensions: Rate limiting factors and evidence for the involvement of mitochondrial electron transport. Plant J. 2005, 41, 732–743. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Molina, I.; Salles, C.; Nicolas, M.; Crouzet, J. Grape alcohol dehydrogenase. II Kinetics studies: Mechanism, substrate and coenzyme specificity. Am. J. Enol. Vitic. 1987, 38, 60–64. [Google Scholar]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer | Forward Sequence | Reverse Sequence | Accession Number |
|---|---|---|---|
| Pgb 1 | 5′-ACCAACCCCAAGCTCAAGAC-3′ | 5′-CTGCCACGCCGTATTTCAAG-3′ | U94968.1 |
| NR | 5′-CAACACCAACTCCGTCAT-3′ | 5′-CTGAGTATGCGTATCCCTTG-3′ | X57844.1 |
| NiR | 5′-GACATCGGCTTCATGGGCT-3′ | 5′-GCACGGCCTTCTTGTACACC-3′ | S78730.1 |
| ADH1 | 5‘-GATCTGCTCAGGATCAACAC-3′ | 5′-GTGGAAGTCCCTACGAAATG-3′ | AF253472.1 |
| ADH3 | 5′-GTCTCTCAACTGGACTTGGTG-3′ | 5′-TAGCTTGTTCGTATTTTGCAGG-3′ | X12734.1 |
| SDH-A | 5′-CAGTGAAGGTGAGCGTTTCA-3′ | 5′-CACCAGCAAAAATAGCAGCA-3′ | AK376855.1 |
| SDH-B | 5′-TGTACGAGTGCATCCTCTGC-3′ | 5′-TCGTCGTTGATGGAGTCAAG-3′ | AK372209.1 |
| PDC-E1 | 5′-ATTGAATTCCGCCCTTGACG-3′ | 5′-GCCAGTAAAACCAGCCTCTG-3′ | AK353615.1 |
| PDC-E2 | 5′-TGCAGGGATGGAAAGAGCTT-3′ | 5′-GGTTGGAGCTGCTTCATACG-3′ | AK362954.1 |
| AOX1a | 5′-CGTCAACCACTTCGCATCGG-3′ | 5′-GCCCTCATTTCCTCGGAAGC-3′ | AK363239.1 |
| AOX1d1 | 5′-CACTACGCATCCGACATCCA-3′ | 5′-CAACAATCCATCCAAATTAACG-3′ | AK365405.1 |
| NDB2 | 5′-CGTCCACTGTCGCTCTGC-3′ | 5′-GGCATCCTCCACTTCCTTCAG-3′ | AK367948.1 |
| NDB3 | 5′-GCAAAATCCAGCTACTGGCG-3′ | 5′-TTCACGCACCCTTAGCCATT-3′ | AK354220.1 |
| GAPDH | 5′-GCTCAAGGGTATCATGGGTTACG-3′ | 5′GCAATTCCACCCTTAGCATCAAAG-3′ | AB120301.1 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zafari, S.; Hebelstrup, K.H.; Igamberdiev, A.U. Transcriptional and Metabolic Changes Associated with Phytoglobin Expression during Germination of Barley Seeds. Int. J. Mol. Sci. 2020, 21, 2796. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21082796
Zafari S, Hebelstrup KH, Igamberdiev AU. Transcriptional and Metabolic Changes Associated with Phytoglobin Expression during Germination of Barley Seeds. International Journal of Molecular Sciences. 2020; 21(8):2796. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21082796
Chicago/Turabian StyleZafari, Somaieh, Kim H. Hebelstrup, and Abir U. Igamberdiev. 2020. "Transcriptional and Metabolic Changes Associated with Phytoglobin Expression during Germination of Barley Seeds" International Journal of Molecular Sciences 21, no. 8: 2796. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21082796