Acetylated Polysaccharides From Pleurotus geesteranus Alleviate Lung Injury Via Regulating NF-κB Signal Pathway


Abstract
:1. Introduction
2. Results
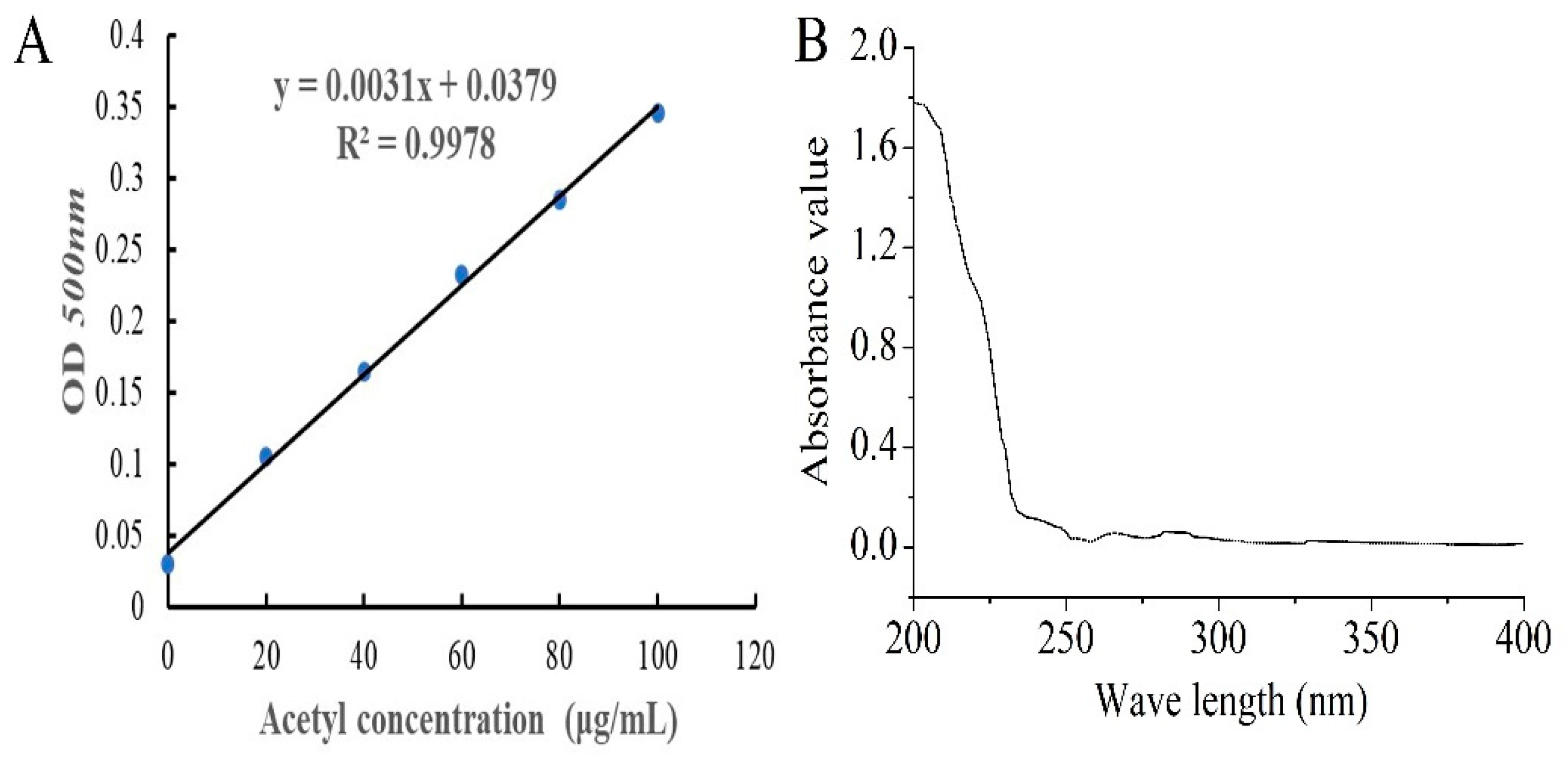
2.1. Preparation and Yield of AcPPS
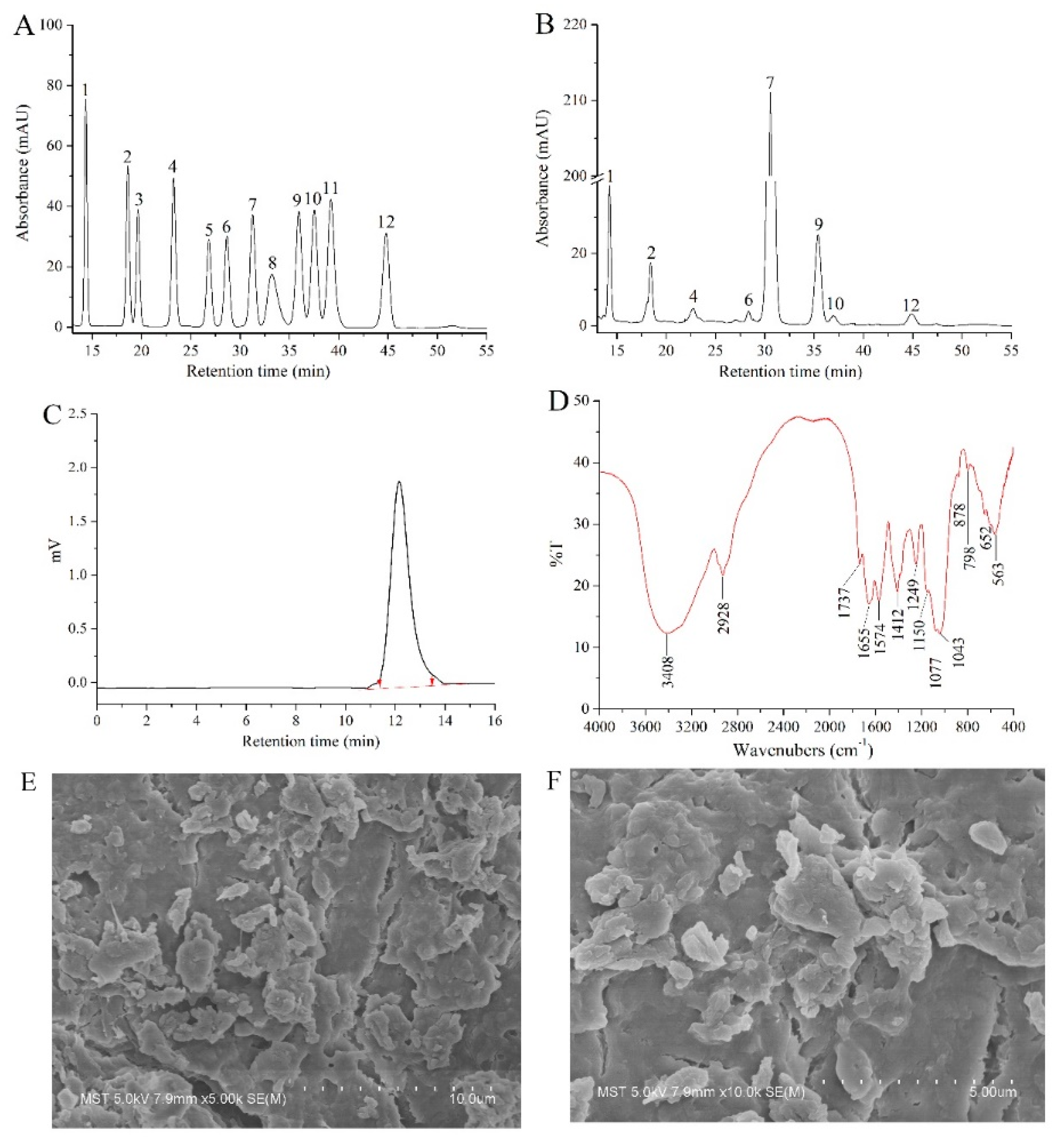
2.2. Structural Characterization
2.3. Acute Toxicity Analysis
2.4. Effects of AcPPS on Body Weight and Lung Index
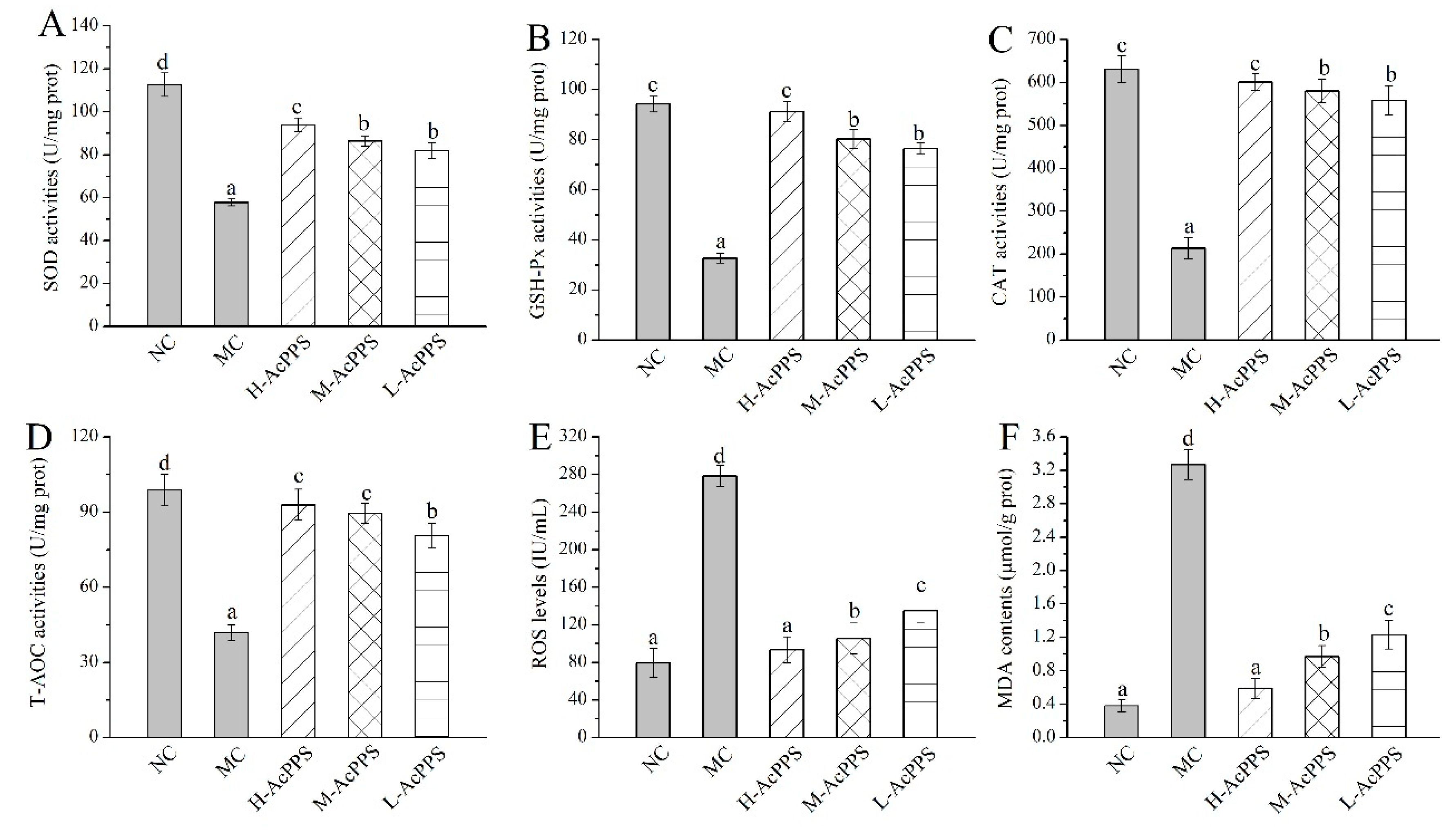
2.5. Effects of AcPPS on Oxidative Stress
2.6. Effects of AcPPS on Lipid Peroxidation
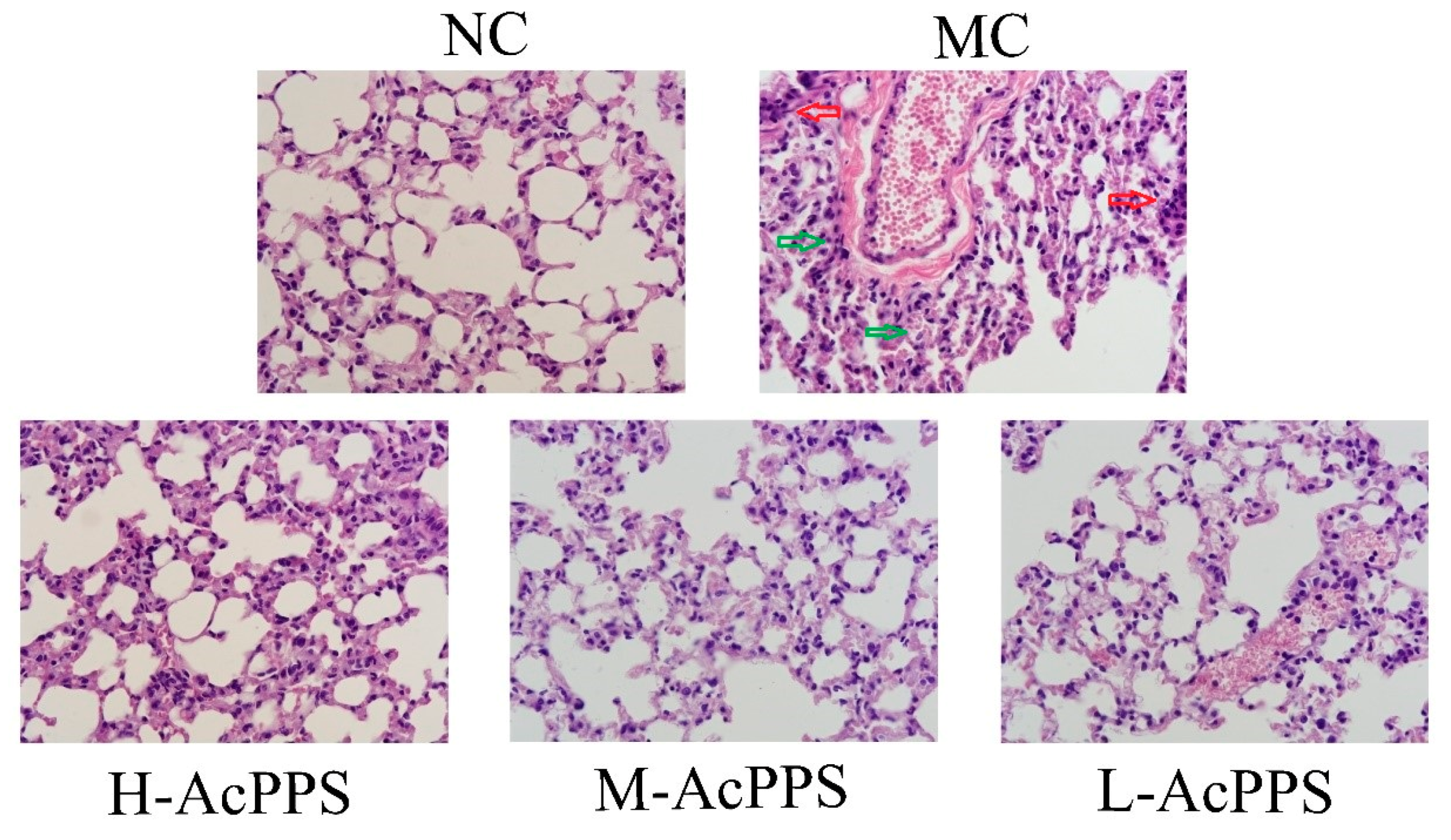
2.7. Histopathological Observations
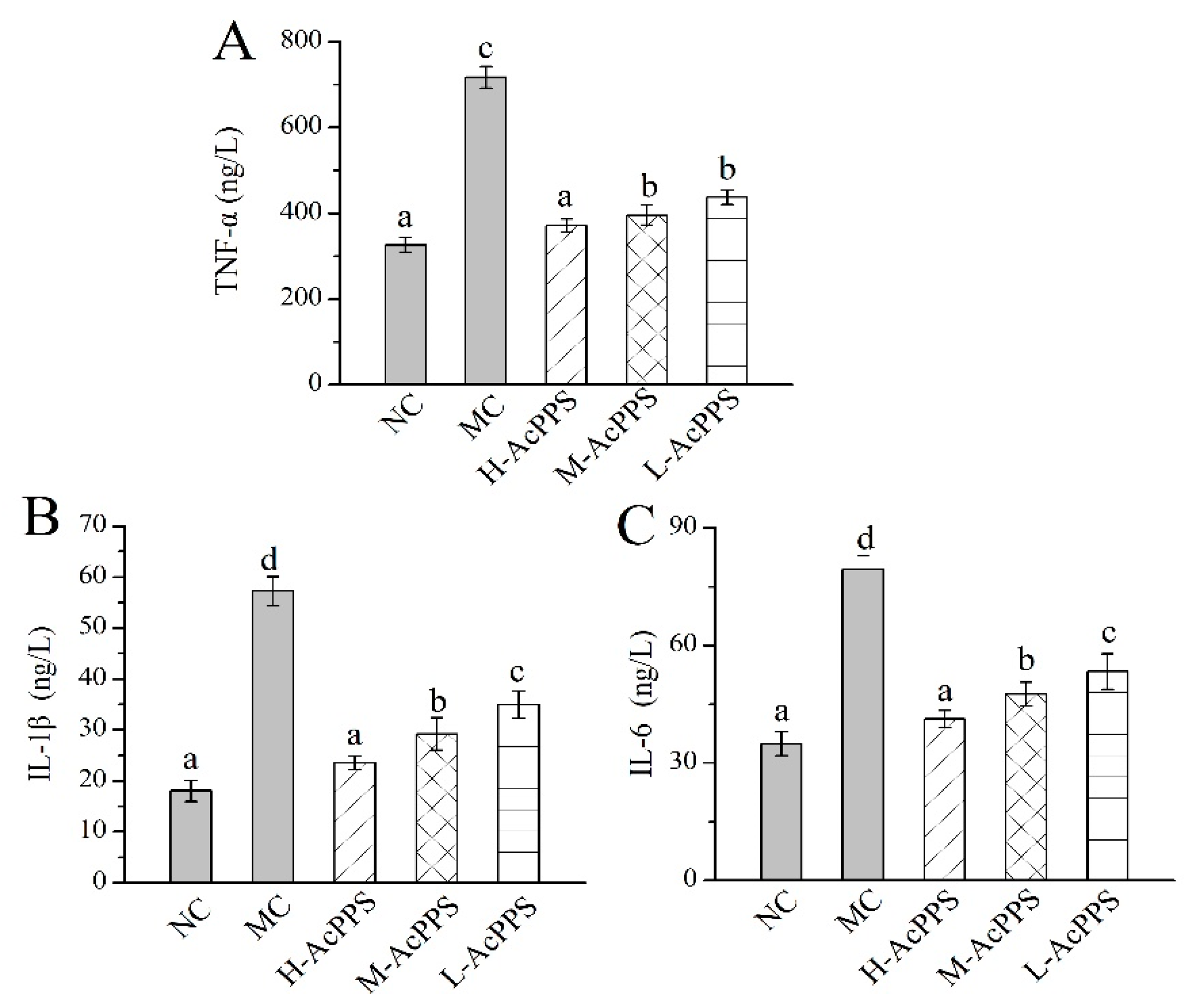
2.8. Effects of AcPPS on Inflammatory Index of BALF
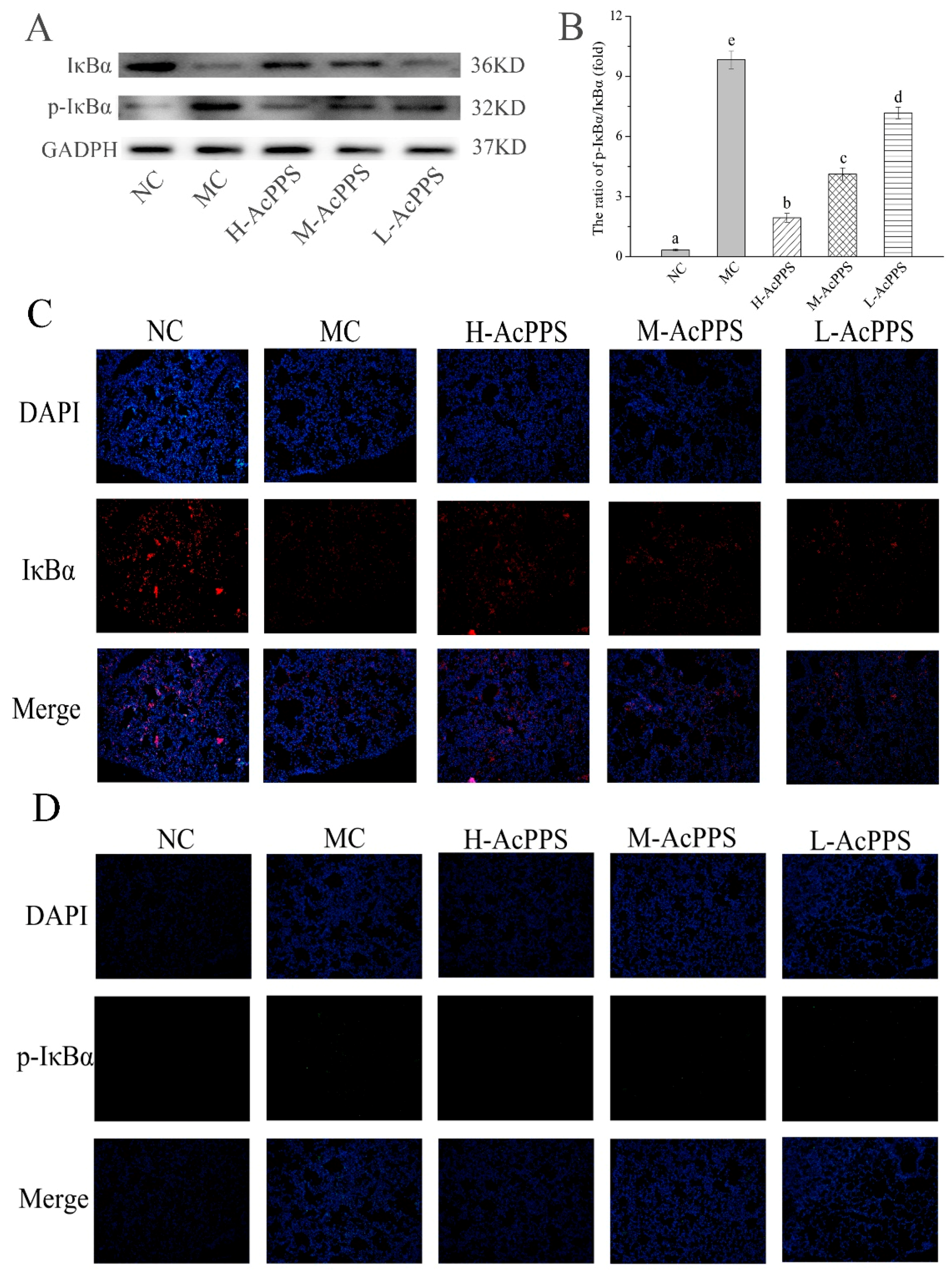
2.9. NF-κB Pathway Detection
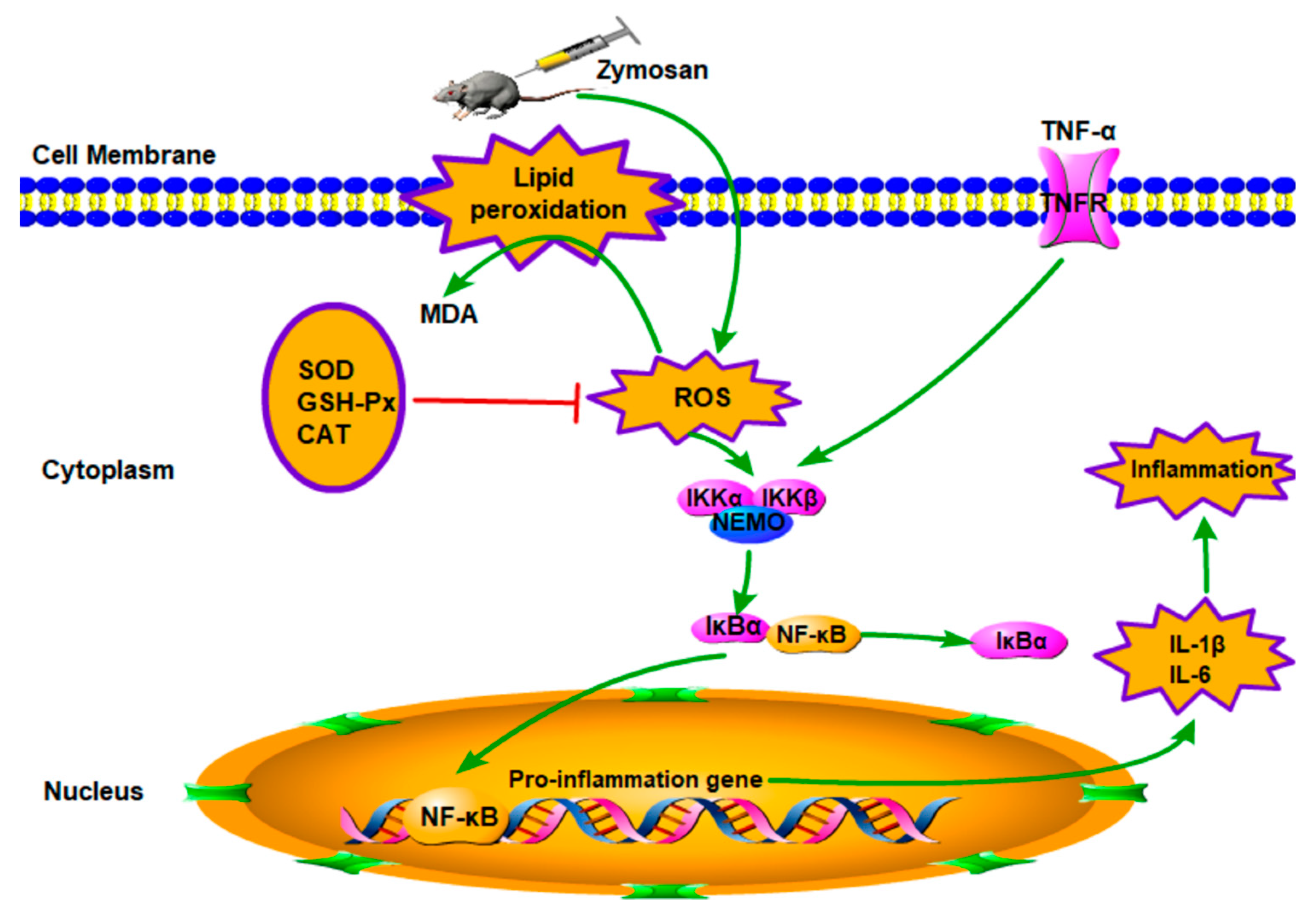
3. Discussion
4. Materials and Methods
4.1. Chemicals and Reagents
4.2. Materials and Methods
4.3. Preparation and Purification of AcPPS
4.3.1. Preparation of AcPPS
4.3.2. Standard Curve Plotting of Acetyl Content
4.3.3. The DS Assay of AcPPS
4.3.4. UV Analysis
4.4. HPGPC, GPC, FT-IR Spectroscopy, and SEM Analysis
4.5. Acute Toxicity Experiment
4.6. Animal Experiments
4.7. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Data Availability
Compliance with Ethical Standards
Abbreviations
| AcPPS | Acetylated Pleurotus geesteranus polysaccharides |
| ALI | Acute lung injury |
| BALF | Bronchoalveolar lavage fluid |
| GalN | Aminogalactose |
| Ara | Arabinose |
| CAT | Catalase |
| DS | Degree of substitution |
| ELISA | Enzyme-linked immunosorbent assay |
| FT-IR | Fourier transform infrared spectroscopy |
| Fuc | Fucose |
| Gal | Galactose |
| GalA | Galacturonic acid |
| GSH-Px | Glutathione peroxidase |
| GPC | Gel permeation chromatography |
| GlcN | Glucosamine |
| Glc | Glucose |
| GlcA | Glucuronic acid |
| HDL-C | High-density lipoprotein cholesterol |
| H&E | Hematoxylin-eosin |
| HPGPC | High-performance gel permeation chromatography |
| IF | Immunofluorescence |
| IL | Interleukins |
| LDL-C | Low density lipoprotein cholesterol |
| MDA | Malondialdehyde |
| Man | Mannose |
| MC | Model control |
| MODS | Multiple organ dysfunction syndrome |
| NC | Normal control |
| Mn | Number-average molar mass |
| PMP | Polymethylpentene |
| PPS | Polysaccharides from P. geesteranus |
| ROS | Reactive oxygen species |
| Rha | Rhamnose |
| Rib | Ribose |
| SEM | Scanning electron microscope |
| SOD | Superoxide dismutase |
| SIRS | Systemic inflammatory response syndrome |
| T-AOC | Total antioxidant capacity |
| TC | Total cholesterol |
| TG | Triglyceride |
| TNF-α | Tumor necrosis factor-α |
| UV | Ultraviolet |
| VLDL-C | Very low-density lipoprotein cholesterol |
| Mw | Weight-average molecular weight |
| Xyl | Xylose |
| Mz | Z-average molecular weight |
| ZY | Zymosan |
| ZIGI | Zymosan-induced generalized inflammation |
References
- Bone, R.C. Immunologic dissonance: A continuing evolution in our understanding of the systemic inflammatory response syndrome (SIRS) and the multiple organ dysfunction syndrome (MODS). Ann. Intern. Med. 1996, 125, 680–687. [Google Scholar] [CrossRef] [PubMed]
- Zheng, G.Z.; Ren, H.J.; Li, H.Q.; Li, X.H.; Dong, T.C.; Xu, S.M.; Yan, Y.L.; Sun, B.K.; Bai, J.W.; Li, Y.S. Lycium barbarum polysaccharide reduces hyperoxic acute lung injury in mice through Nrf2 pathway. Biomed. Pharmacother. 2019, 111, 733–739. [Google Scholar] [CrossRef] [PubMed]
- Zhao, H.Y.; Zhao, M.; Wang, Y.; Li, F.C.; Zhang, Z.G. Glycyrrhizic Acid Prevents Sepsis-Induced Acute Lung Injury and Mortality in rats. J. Histochem. Cytochem. 2016, 64, 125–137. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, X.R.; Yin, L.H.; Fang, L.L.; Xu, L.N.; Sun, P.Y.; Xu, M.; Liu, K.X.; Peng, J.Y. Protective effects of dioscin against systemic inflammatory response syndromevia adjusting TLR2/MyD88/NF-κb signal pathway. Int. Immunopharmacol. 2018, 65, 458–469. [Google Scholar] [CrossRef] [PubMed]
- Goris, R.J.; Boekholtz, W.K.; van Bebber, I.P.; Nuytinck, J.K.; Schillings, P.H. Multiple-organ failure and sepsis without bacteria: An experimental model. Arch. Surg. 1986, 121, 897–901. [Google Scholar] [CrossRef]
- Hook, J.S.; Cao, M.; Potera, R.M.; Alsmadi, N.Z.; Schmidtke, D.W.; Moreland, J.G. Nox2 regulates platelet activation and NET formation in the lung. Front. Immunol. 2019, 10, 1472–1482. [Google Scholar] [CrossRef]
- Liu, Z.J.; Jiao, Y.C.; Lu, H.Y.; Shu, X.L.; Chen, Q.H. Chemical characterization, antioxidant properties and anticancer activity of exopolysaccharides from Floccularia luteovirens. Carbohydr. Polym. 2020, 229, 115432. [Google Scholar] [CrossRef]
- Chabot, F.; Mitchell, J.A.; Gutteridge, J.M.; Evans, T.W. Reactive oxygen species in acute lung injury. Eur Respir J. 1998, 11, 745–757. [Google Scholar] [CrossRef]
- Gutteridge, J.M.C.; Quinlan, G.J. Reactive oxygen species, antioxidant protection and lung injury. In ARDS Acute Respiratory Distress in Adults; Springer: New York, NY, USA, 1996; pp. 167–195. [Google Scholar] [CrossRef]
- Kellner, M.; Noonepalle, S.; Lu, Q.; Srivastava, A.; Zemskov, E.; Black, S.M. ROS signaling in the pathogenesis of acute lung injury (ALI) and acute respiratory distress syndrome (ARDS). Adv. Exp. Med. Biol. 2017, 967, 105–137. [Google Scholar] [CrossRef]
- Li, W.A.; Fu, K.Q.; Lv, X.P.; Wang, Y.; Wang, J.F.; Li, H.T.; Tian, W.R.; Gao, R.F. Lactoferrin suppresses lipopolysaccharide-induced endometritis in mice via down-regulation of the NF-κB pathway. Int. Immunopharmacol. 2015, 28, 695–699. [Google Scholar] [CrossRef]
- Tsedensodnom, O.; Vacaru, A.M.; Howarth, D.L.; Yin, C.; Sadler, K.C. Ethanol metabolism and oxidative stress are required for unfolded protein response activation and steatosis in zebrafish with alcoholic liver disease. Dis. Model. Mech. 2013, 6, 1213–1226. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fu, Z.L.; Xia, L.; De, J.; Zhu, M.X.; Li, H.; Lu, Y.; Chen, D.F. Beneficial effects on H1N1-induced acute lung injury and structure characterization of anti-complementary acidic polysaccharides from Juniperus pingii var. wilsonii. Int. J. Biol. Sci. 2019, 129, 246–253. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Zhang, H.; Wang, Y.X.; Nie, S.P.; Li, C.; Xie, M.Y. Acetylation and carboxymethylation of the polysaccharide from Ganoderma atrum and their antioxidant and immunomodulating activities. Food Chem. 2014, 156, 279–288. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.X.; Chen, J.L.; Lei, L.; Li, F.H.; Tang, Y.; Yuan, Y.; Zhang, Y.Q.; Wu, S.R.; Yin, R.; Ming, J. Acetylation of polysaccharide from Morchella angusticeps peck enhances its immune activation and anti-inflammatory activities in macrophage RAW264.7 cells. Food Chem. Toxicol. 2019, 125, 38–45. [Google Scholar] [CrossRef]
- Ni, Y.L.; Shen, H.T.; Su, C.H.; Chen, W.Y.; Huang-Liu, R.; Chen, C.J.; Chen, S.P.; Kuan, Y.H. Nerolidol suppresses the inflammatory response during lipopolysaccharide-induced acute lung injury via the modulation of antioxidant enzymes and the AMPK/Nrf-2/HO-1 pathway. Oxid. Med. Cell. Longev. 2019, 16, 1–11. [Google Scholar] [CrossRef]
- Greig, F.H.; Kennedy, S.; Spickett, C.M. Physiological effects of oxidized phospholipids and their cellular signaling mechanisms in inflammation. Free Radical Biol. Med. 2012, 52, 266–280. [Google Scholar] [CrossRef]
- Yeh, C.H.; Yang, J.J.; Yang, M.L.; Li, Y.C.; Kuan, Y.H. Rutin decreases lipopolysaccharide-induced acute lung injury via inhibition of oxidative stress and the MAPK-NF-κB pathway. Free Radical Biol. Med. 2014, 69, 249–257. [Google Scholar] [CrossRef]
- Kawahara, K.I. Beyond free radical scavenging: Beneficial effects of edaravone (Radicut) in various diseases (Review). Exp. Ther. Med. 2012, 3, 3–8. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Z.; Zhou, J.; Song, D.Q.; Sun, Y.H.; Liao, C.L.; Jiang, X. Gastrodin protects against LPS-induced acute lung injury by activating Nrf2 signaling pathway. Oncotarget 2017, 8, 32147–32156. [Google Scholar] [CrossRef] [Green Version]
- Welbourn, C.R.; Young, Y. Endotoxin, septic shock and acute lung injury: Neutrophils, macrophages and inflammatory mediators. British J. Surg. 1992, 79, 998–1003. [Google Scholar] [CrossRef]
- Qi, T.J.; Xu, F.; Yan, X.X.; Li, S.A.; Li, H.T. Sulforaphane exerts anti-inflammatory effects against lipopolysaccharide-induced acute lung injury in mice through the Nrf2/ARE pathway. Int. J. Mol. Med. 2015, 37, 182–188. [Google Scholar] [CrossRef] [Green Version]
- Yang, B.; Huang, W.; Han, J.; Liang, Z. Study of the role of epidermal growth factor on lung fluid transport in rabbits with acute lung injury caused by endotoxin. Exp. Ther. Med. 2012, 4, 611–614. [Google Scholar] [CrossRef] [PubMed]
- Zhao, H.L.; Eguchi, S.; Alam, A.; Ma, D.Q. The role of nuclear factor-erythroid 2 related factor 2 (Nrf-2) in the protection against lung injury. Am. J. Physiol. Lung Cell Mol. Physiol. 2017, 312, L155–L162. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Enshasy, H.A.; Hatti-Kaul, R. Mushroom immunomodulators: Unique molecules with unlimited applications. Trends Biotechnol. 2013, 31, 668–677. [Google Scholar] [CrossRef] [PubMed]
- Zhao, L.Y.; Lai, S.S.; Huang, R.; Wu, M.Y.; Gao, N.; Xu, L.; Qin, H.B.; Peng, W.L.; Zhao, J.H. Structure and anticoagulant activity of fucosylated glycosaminoglycan degraded by deaminative cleavage. Carbohydr. Polym. 2013, 98, 1514–1523. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Wang, Z.Q.; Wu, H.H.; Jia, W.; Teng, L.S.; Song, J.; Yang, X.G.; Wang, D. Sarcodon imbricatus polysaccharides protect against cyclophosphamide-induced immunosuppression via regulating Nrf2-mediated oxidative stress. Int. J. Biol. Macromol. 2018, 8, 157–162. [Google Scholar] [CrossRef]
- Zhang, M.; Zhu, L.; Cui, S.W.; Wang, Q.; Zhou, T.; Shen, H.S. Fractionation, partial characterization and bioactivity of water-soluble polysaccharides and polysaccharide-protein complexes from Pleurotus geesteranus. Int. J. Biol. Macromol. 2011, 48, 5–12. [Google Scholar] [CrossRef]
- Mao, D.B.; Ma, Y.P.; Geng, L.J.; Zhao, A.J.; Zheng, J.Q.; Xu, C.P. Fermentation characteristics in stirred-tank reactor of exopolysaccharides with hypolipidemic activity produced by Pleurotus geesteranus 5#. An. Acad. Bras. Cienc. 2013, 85, 1473–1481. [Google Scholar] [CrossRef]
- Song, X.L.; Liu, Z.H.; Zhang, J.J.; Zhang, C.; Dong, Y.H.; Ren, Z.Z.; Gao, Z.; Liu, M.; Zhao, H.J.; Jia, L. Antioxidative and hepatoprotective effects of enzymatic and acidic-hydrolysis of Pleurotus geesteranus mycelium polysaccharides on alcoholic liver diseases. Carbohydr. Polym. 2018, 201, 75–86. [Google Scholar] [CrossRef]
- Chan, Y.Y.; Chan, E.; Chan, S.W.; Sze, S.Y.; Chan, M.F.; Tsui, S.H.; Leung, K.Y.; Chan, R.Y.; Chung, I.Y. Enhancement of in vitro and in vivo anticancer activities of polysaccharide peptide from Grifola frondosa by chemical modifications. Pharm. Biol. 2011, 49, 1114–1120. [Google Scholar] [CrossRef]
- Yuan, Q.X.; Zhao, L.Y.; Li, Z.H.; Harqin, C.; Peng, Y.F.; Liu, J.K. Physicochemical analysis, structural elucidation and bioactivities of a high-molecular-weight polysaccharide from Phellinus igniarius mycelia. Int. J. Biol. Macromol. 2018, 120, 1855–1864. [Google Scholar] [CrossRef] [PubMed]
- Zhao, P.; Zhou, H.F.; Zhao, C.C.; Li, X.; Wang, Y.; Wang, Y.; Huang, L.Q.; Gao, W.Y. Purification, characterization and immunomodulatory activity of fructans from Polygonatum odoratum and P. cyrtonema. Carbohydr. Polym. 2019, 214, 44–52. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Zhou, C.; Huang, S.; Jiang, C. Selenium polysaccharide SPMP-2a from Pleurotus geesteranus alleviates H2O2-induced oxidative damage in HaCaT cells. BioMed Res. Int. 2017, 2017, 4940384. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meng, M.; Cheng, D.; Han, L.R.; Chen, Y.Y.; Wang, C.L. Isolation, purification, structural analysis and immunostimulatory activity of water-soluble polysaccharides from Grifola Frondosa fruiting body. Carbohydr. Polym. 2017, 157, 1134–1143. [Google Scholar] [CrossRef]
- Song, Y.; Yang, Y.; Zhang, Y.Y.; Duan, L.S.; Zhou, C.L.; Ni, Y.Y.; Liao, X.J.; Li, Q.H.; Hu, X.S. Effect of acetylation on antioxidant and cytoprotective activity of polysaccharides isolated from pumpkin (Cucurbita pepo, lady godiva). Carbohydr. Polym. 2013, 98, 689–691. [Google Scholar] [CrossRef]
- Liu, S.S.; Zhang, J.Y.; Pang, Q.; Song, S.D.; Miao, R.C.; Chen, W.; Zhou, Y.Y.; Liu, C. The protective role of curcumin in zymosan-induced multiple organ dysfunction syndrome in mice. Shock 2016, 45, 209–219. [Google Scholar] [CrossRef]
- Li, B.; Cramer, D.; Wagner, S.; Hansen, R.; King, C.; Kakar, S.; Ding, C.L.; Yan, J. Yeast glucan particles activate murine resident macrophages to secrete proinflammatory cytokines via MyD88 and Syk kinase-dependent pathways. Clin. Immunol. 2007, 124, 170–181. [Google Scholar] [CrossRef] [Green Version]
- Xie, K.L.; Yu, Y.H.; Zhang, Z.S.; Liu, W.B.; Pei, Y.P.; Xiong, L.Z.; Hou, L.C.; Wang, G.L. Hydrogen gas improves survival rate and organ damage in zymosan-induced generalized inflammation model. Shock 2010, 34, 495–501. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.Y.; Choi, G.E.; Yoo, H.J.; Kim, H.S. Interferon potentiates toll-like receptor-induced prostaglandin D2 production through positive feedback regulation between signal transducer and activators of transcription 1 and reactive oxygen species. Front. Immunol. 2017, 8, 1720–1731. [Google Scholar] [CrossRef] [Green Version]
- Chang, L.; Karin, M. Mammalian map kinase signalling cascades. Nature 2001, 410, 37–40. [Google Scholar] [CrossRef]
- Liu, J.Z.; Wang, X.; Peng, Z.W.; Zhang, T.; Wu, H.; Yu, W.H.; Kong, D.Q.; Liu, Y.; Bai, H.; Liu, R.; et al. The effects of insulin pre-administration in mice exposed to ethanol: Alleviating hepatic oxidative injury through anti-oxidative, anti-apoptotic activities and deteriorating hepatic steatosis through SRBEP-1c activation. Int. J. Biol. Sci. 2015, 11, 569–586. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, J.J.; Mao, D.; Yong, Y.Y.; Li, J.L.; Wei, H.; Lu, L. Hepatoprotective and hypolipidemic effects of water-soluble polysaccharidic extract of Pleurotus eryngii. Food Chem. 2012, 130, 687–694. [Google Scholar] [CrossRef]
- Sahanfirat, S.; Temizresitoglu, M.; Guden, D.S.; Kucukkavruk, S.P.; Tunctan, B.; Sari, A.N.; Kocak, Z.; Malik, K.U. Protection by mTOR inhibition on zymosan-induced systemic inflammatory response and oxidative/nitrosative stress: Contribution of mTOR/MEK1/ERK1/2/IKKβ/IκB-α/NF-κB signalling pathway. Inflammation 2018, 41, 276–298. [Google Scholar] [CrossRef] [PubMed]
- Von Asmuth, E.J.; Maessen, J.G.; van der Linden, C.J.; Buurman, W.A. Tumour necrosis factor alpha (TNF-alpha) and interleukin 6 in a zymosan-induced shock model. Scand. J. Immunol. 1990, 32, 313–319. [Google Scholar] [CrossRef]
- Miao, S.S.; Mao, X.H.; Pei, R.; Miao, S.P.; Xiang, C.; Lv, Y.J.; Yang, X.G.; Sun, J.; Jia, S.S.; Liu, Y.P. Antitumor activity of polysaccharides from Lepista sordida against laryngocarcinoma in vitro and in vivo. Int. J. Biol. Macromol. 2013, 60, 235–240. [Google Scholar] [CrossRef] [PubMed]
- Dueñaschasco, M.T.; Rodríguezcarvajal, M.A.; Tejero, M.; FranceRodriguez, G.; Espartero, J.L.; Irastorza-Iribas, A.; Gil-Serranob, A.M. Structural analysis of the exopolysaccharide produced by Pediococcus damnosus 2.6. Carbohydr. Res. 1997, 303, 453–458. [Google Scholar] [CrossRef]
- Zhang, J.J.; Gao, X.; Pan, Y.G.; Xu, N.; Jia, L. Toxicology and immunology of Ganoderma Lucidum polysaccharides in Kunming mice and Wistar rats. Int. J. Biol. Macromol. 2016, 85, 302–310. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Groups | Body Weight (g) | Lung Weight (g) | Lung Index (%) | ||
|---|---|---|---|---|---|
| Treatment Time by Zymosan 1 d | Treatment Time by Zymosan 4 d | Treatment Time by Zymosan 7 d | |||
| NC | 42.78 ± 0.24 a | 43.40 ± 0.19 d | 44.50 ± 0.14 d | 0.33 ± 0.09 a | 0.74 ± 0.09 a |
| MC | 42.60 ± 0.26 a | 39.61 ± 0.17 a | 38.97 ± 0.23 a | 0.53 ± 0.07 c | 1.36 ± 0.12 c |
| H-AcPPS | 42.37 ± 0.29 a | 41.68 ± 0.14 c | 43.18 ± 0.16 c | 0.34 ± 0.02 a | 0.79 ± 0.07 a |
| M-AcPPS | 42.16 ± 0.12 a | 41.07 ± 0.18 b | 42.33 ± 0.25 b | 0.35 ± 0.04 a | 0.83 ± 0.14 a |
| L-AcPPS | 42.68 ± 0.15 a | 41.24 ± 0.23 c | 42.06 ± 0.14 b | 0.40 ± 0.03 b | 0.95 ± 0.11 b |
| NC | MC | H-AcPPS | M-AcPPS | L-AcPPS | ||
|---|---|---|---|---|---|---|
| Serum lipid parameters | TC (mmol/L) | 1.36 ± 0.11 a | 2.28 ± 0.15 c | 1.47 ± 0.08 a | 1.53 ± 0.12 a | 1.87 ± 0.09 b |
| TG (mmol/L) | 0.75 ± 0.02 a | 1.83 ± 0.09 d | 0.80 ± 0.05 a | 0.97 ± 0.08 b | 1.06 ± 0.07 c | |
| HDL-C (mmol/L) | 2.16 ± 0.15 c | 1.01 ± 0.08 a | 2.03 ± 0.11 c | 1.95 ± 0.09 b | 1.78 ± 0.07 b | |
| LDL-C (mmol/L) | 0.55 ± 0.06 a | 1.71 ± 0.12 d | 0.67 ± 0.07 a | 0.91 ± 0.05 b | 1.18 ± 0.15 c | |
| vLDL-C (mmol/L) | 0.34 ± 0.05 a | 1.26 ± 0.11 d | 0.45 ± 0.02 a | 0.61 ± 0.09 b | 0.78 ± 0.04 c |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Song, X.; Zhang, J.; Li, J.; Jia, L. Acetylated Polysaccharides From Pleurotus geesteranus Alleviate Lung Injury Via Regulating NF-κB Signal Pathway. Int. J. Mol. Sci. 2020, 21, 2810. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21082810
Song X, Zhang J, Li J, Jia L. Acetylated Polysaccharides From Pleurotus geesteranus Alleviate Lung Injury Via Regulating NF-κB Signal Pathway. International Journal of Molecular Sciences. 2020; 21(8):2810. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21082810
Chicago/Turabian StyleSong, Xinling, Jianjun Zhang, Jian Li, and Le Jia. 2020. "Acetylated Polysaccharides From Pleurotus geesteranus Alleviate Lung Injury Via Regulating NF-κB Signal Pathway" International Journal of Molecular Sciences 21, no. 8: 2810. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21082810