Long Non-Coding RNAs, the Dark Matter: An Emerging Regulatory Component in Plants



1
State Key Laboratory for Conservation and Utilization of Subtropical Agro-Bioresources, South China Agricultural University, Guangzhou 510640, China
2
Guangdong Laboratory for Lingnan Modern Agriculture, South China Agricultural University, Guangzhou 510640, China
3
Key Laboratory of Biology and Germplasm Enhancement of Horticultural Crops in South China, Ministry of Agriculture and Rural Affairs, South China Agricultural University, Guangzhou 510640, China
*
Author to whom correspondence should be addressed.
Int. J. Mol. Sci. 2021, 22(1), 86; https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22010086
Submission received: 1 December 2020
/
Revised: 18 December 2020
/
Accepted: 19 December 2020
/
Published: 23 December 2020
(This article belongs to the Section Molecular Plant Sciences)
Abstract
:Long non-coding RNAs (lncRNAs) are pervasive transcripts of longer than 200 nucleotides and indiscernible coding potential. lncRNAs are implicated as key regulatory molecules in various fundamental biological processes at transcriptional, post-transcriptional, and epigenetic levels. Advances in computational and experimental approaches have identified numerous lncRNAs in plants. lncRNAs have been found to act as prime mediators in plant growth, development, and tolerance to stresses. This review summarizes the current research status of lncRNAs in planta, their classification based on genomic context, their mechanism of action, and specific bioinformatics tools and resources for their identification and characterization. Our overarching goal is to summarize recent progress on understanding the regulatory role of lncRNAs in plant developmental processes such as flowering time, reproductive growth, and abiotic stresses. We also review the role of lncRNA in nutrient stress and the ability to improve biotic stress tolerance in plants. Given the pivotal role of lncRNAs in various biological processes, their functional characterization in agriculturally essential crop plants is crucial for bridging the gap between phenotype and genotype.
1. Introduction
Approximately 90% of the genome is pervasively transcribed into RNA. However, only 1–2% of the transcribed genome encodes for putative protein-coding RNAs, whereas the remaining 98–99% RNAs remain uncharacterized or do not code for any functional protein and are regarded as “dark matter,” transcriptional “noise,” “junk DNA,” or an “experimental artifact” [1,2,3]. Such RNAs are considered to be non-coding RNAs (ncRNAs) [1]. These ncRNAs are of three types: regulatory ncRNAs, circular ncRNAs, and structural ncRNAs. The regulatory ncRNAs are grouped into two sub-groups based on their length: (1) small RNAs (sRNAs), which are 20–31 bp in length, including small interfering RNAs (siRNAs: 19–25 bp), piwi-interacting RNAs (piRNAs: 26–31), and microRNAs (miRNAs: 20–22 bp); and (2) long regulatory RNAs (long non-coding RNAs (lncRNAs) > 200 bp) [4]. Structural or housekeeping ncRNAs consist of transfer RNAs (tRNAs), ribosomal RNAs (rRNAs), small nucleolar RNAs (snoRNAs), and small nuclear RNAs (snRNAs) [5]. CircRNAs (circRNAs) have been recently discovered as a new group of ncRNAs, generally generated by alternative back splicing of pre-mRNA [6].
lncRNAs constitute approximately 80% of total ncRNAs and are thought to play a significant role or act as a critical component in various biological processes [7]. lncRNAs are > 200 bp in length and do not have discernible coding potential. However, lncRNAs share similarities with mRNAs, such as transcription by RNA polymerase II (Pol II), a cap structure at the 5′-end and 3′-poly (A) tail, and splicing [8]. Pol II transcribes most lncRNAs, whereas plant-specific RNA polymerases Pol IV/V can produce lncRNAs [9]. The lncRNAs can be regarded as gene regulatory factors and contribute to three modes of gene expression regulation—transcriptional, post-transcriptional, and epigenetic [4,10]. A significant proportion of lncRNAs (42%) have been found to be involved in transcriptional regulation through transcriptional interference. However, post-transcriptional regulations occur in the cytoplasm, whereas transcriptional and epigenetic modifications occur in the nucleus [11].
Plant lncRNAs are involved in diverse biological processes, including photomorphogenesis, flowering, phosphate homeostasis, stress tolerance, and fertility [9,12]. They also play a vital role as a molecular sink for the target mimics of miRNAs to process into shorter ncRNAs [13], in protein subcellular re-localization [14], as repressors of histone acetylation, in direct epigenetic silencing mediated by specific chromatin domains [15], and in regulating protein post-transcriptional regulation [13,16]. This review briefly delineates the general characteristics of lncRNAs, including classification, progress in annotation, and key underlying mechanisms in plant development and stress tolerance. We aim to elucidate the lncRNA-mediated regulation of important plant developmental processes such as vernalization, fertility, lateral root development, photomorphogenesis, nodulation, and fiber development, and provide guidance for developing optimal plant breeding strategies in future crop breeding programs.
1.1. Discovery of lncRNAs
During the pre-genomics era, the lncRNA H19—a highly conserved transcript with a length of 2.5 kb—was first discovered in mice in 1984, and was noted to be highly abundant in embryonic cell lineage (Figure 1) [17]. H19 is located upstream of insulin-like growth factor 2 (Igf2) gene on chromosome 11 in humans, and on chromosome 7 in mice; they are coregulated and share a common imprinting mechanism [18]. Since this discovery, with the advent of high throughput sequencing, several regulatory lncRNAs such as Airn, HOTAIR, MALAT1, and X-inactive-specific transcript (Xist) have been discovered and further characterized. The first plant lncRNA, EARLY NODULIN 40 (ENOD40), was isolated from nodule primordia in Medicago plants and found to be involved in symbiotic nodule organogenesis [19]. ENOD40 can induce cytoplasmic localization of nuclear RNA binding protein MtRBP1 in Medicago truncatula during nodule formation [20]. Subsequently, several researchers identified and characterized lncRNAs as a regulator of flowering time [21], anther development and male fertility [22,23], innate immunity [24], nutrient deficiency, and toxicity [25] (Figure 1).
1.2. Biogenesis of lncRNAs
lncRNAs belong to a heterogeneous subclass of > 200-nt ncRNA transcripts lacking coding potential and appear as an emerging field of research that has been accelerated and expanded in the last decade. lncRNAs act as riboregulators and are localized in both the cytoplasm and nucleus and are transcribed by RNA polymerase II or III or IV/V [26,27,28]. RNA Pol II transcribes lncRNAs with a 5′-cap and poly-adenylation at the 3′ tail from various genome regions, including enhancer or intron splicing regions [29]. RNA pol IV and V generate lncRNAs through RNA-dependent DNA methylation (RdDM), which serve as precursors for small interfering RNAs (siRNAs) [30]. lncRNAs, with their possible biogenesis, are illustrated in Figure 2.
1.3. Status of lncRNAs in Plants
To date, a plethora of putative lncRNAs have been predicted in various plant species, such as Arabidopsis thaliana, Solanum lycopersicum, Zea mays, Medicago, Oryza sativa, Solanum tuberosum, Triticum aestivum, Populus trichocarpa, Setaria italica, and Brassica rapa (Table 1). The number of identified lncRNAs varies considerably, ranging from 6480 in A. thaliana to 58,218 in S. tuberosum [11]. All of these putative lncRNAs play various roles in response to different biotic stresses such as tomato yellow leaf curl virus (TYLCV), infection by Sclerotinia sclerotiorum, Fusarium oxysporum, and Puccinia striiformis, as well as to abiotic or environmental stresses like drought, low and high temperatures, osmotic or hypoxic stress, and nitrogen or phosphate deficiency [11]. In Camelina (Camelina sativa L.), through drought-stress specific transcriptome analysis, 5390 putative lncRNAs were identified, including 1347 intergenic, 69 antisense, 670 sense, and 2681 intronic lncRNAs [31]. Mining more than 200 transcriptome datasets for Arabidopsis [32] revealed that 70% of lncRNAs (~40,000 candidate lncRNAs; >30,000 natural antisense transcripts (NATs); >6000 long intergenic ncRNAs (lincRNAs)) were transcribed from coding loci to be associated with antisense transcripts [33,34]. Similarly, large-scale transcriptome and genome resequencing between rice and its closely related wild relatives (858 accessions) revealed 3363 lncRNAs in developing panicles. Among these, 329 lncRNAs were downregulated [35]. Genome-scale lncRNAs annotation from 749 RNA sequencing experiments on Z. mays encompassing 17 different developmental stages uncovered 18,165 lncRNAs in Z. mays, of which 37.83% lncRNAs were conserved between maize and teosinte [36] These significant differences in expression strongly suggest that lncRNAs diversified during the domestication of crop plants. A total of 1299, 1885, and 1910 lncRNA candidates in B. rapa, B. napus, and B. oleracea were identified, respectively [37]. Mining the rice whole transcriptome RNA sequencing libraries, 2588 lncRNAs under nitrogen starvation [38] and 2224 putative lncRNAs encoding loci associated with the reproductive process were verified [39]. Moreover, a total of 682 lncRNAs in cassava [40], 7245 lncRNAs in Z. mays L. [41], and 2542 lncRNAs in P. trichocarpa [42] were identified. These studies of lncRNAs in plants laid the foundation for the functional characterization of lncRNAs and their potential role in growth, development, and tolerance to various environmental stresses [11,43].
In the past two decades, significant progress in plant lncRNA prediction and functional characterization has been achieved [44]. In planta, some important plant lncRNAs such as phosphate-limitation inducible gene 1 (OsPI1), trehalose phosphate synthase 11 (TPS11), ENOD40, induced by phosphate starvation 1 (IPS1), cooled assisted intronic non-coding RNA (COLDAIR), cold induced long antisense intragenic RNA (COOLAIR), and long day (LD)-specific male-fertility-associated RNA (LDMAR) have provided insights into their distinct biological roles in various fundamental processes including vernalization, male sterility, nodule formation, photomorphogenesis, and phosphate (Pi) uptake [19,21,45,46,47]. For instance, ENOD40 was the first plant lncRNA predicted to act as a riboregulator in M. truncatula, Oryza sativa, Glycine max, and during root nodule organogenesis [20]. COOLAIR and COLDAIR, antisense intronic lncRNA transcripts of flowering locus C (FLC), function as epigenetic repressors of FLC transcription during vernalization in Arabidopsis [46]. Phosphate 2 (PHO2) plays a critical role in phosphate signaling and is one of the targets of miR399 [48]. Phosphate-starvation-inducible lncRNAs At4 and AtIPS1 act as target mimics of miR399 and sequester miR399 to reduce miR399-mediated cleavage of PHO2 [45,48]. These findings suggest that lncRNAs may act as essential regulatory molecules in moderating plant responses to environmental changes [21].
There have been many efforts in the development of computational tools and high-throughput sequencing technologies, such as RNA capture sequencing (RNA CaptureSeq), whole-genome tiling arrays, the lncRNA microarray technique, serial expression of gene expression (SAGE), the expressed sequence tag (EST) method, and RNA-sequencing (RNA-seq). More than half a million lncRNA transcripts have been predicted in silico to understand their biological mechanisms [49]. Accumulating evidence suggests that lncRNAs are emerging regulators of diverse biological processes.
2. Annotation of lncRNAs
Next-generation sequencing (NGS) technology and computational approaches enable us to generate transcript fragments and align them with a reference genome to identify novel RNAs. Likewise, mining RNA-seq data can help readily identify lncRNAs. However, it is challenging to differentiate the coding RNA from non-coding RNAs in thousands of assembled transcripts [50].
2.1. Computational Tools and Resources for lncRNA Identification
In plants, some studies have used both experimental and computational techniques to identify and characterize lncRNAs. Some important computational techniques to distinguish ncRNAs from protein-coding transcripts (Table 2), including coding or non-coding (CONC) for analyzing large ncRNA datasets; and coding potential calculator (CPC: score <0) for known protein-coding transcripts [51]. PhyloCSF distinguishes coding transcripts from non-coding ones using codon substitution frequency (CSF) scores. The Coding-Potential Assessment Tool (CPAT) is based on a logistic regression model, whereas Coding-Non-Coding Index (CNCI) and CPC are based on the support vector machine (SVM) [52,53,54]. To identify lncRNAs from transcriptome sequencing data, two standalone tools based on deep neural networks (DeepLNC) and the SVM algorithm (iSeeRNA) have also been developed [55,56].
Moreover, bioinformatics researchers have developed several tools to predict the mechanism of lncRNA regulation (Table 2). lncRNA–mRNA/–DNA interactions based on nucleic acid thermodynamics can be predicted using the lncRNATargets platform [57]. lncRNA–miRNA interactions and regulatory RNA motifs/functional sites, as well as common transcription factors for lncRNAs can be identified using spongeScan, RegRNA, and TF2LncRNA, respectively [58,59,60].
2.2. Databases for lncRNAs
Over the past decade, the number of discovered lncRNAs has significantly expanded. Thus, there is a need to organize the identified lncRNAs with their biological functions in a user-friendly environment. Several such databases have been archived for identified lncRNAs (Table 1). PLncDB integrates large-scale multi-omic datasets of lncRNAs such as RNA-seq, tilling array, siRNAs, and epigenetics. It contains 16,227 lncRNAs identified from the A. thaliana genome only [32]. PNRD is a collection of nearly all kinds of ncRNAs, including lncRNAs in rice, Arabidopsis, maize, and poplar [65]. lncRNAdb is a database for a few plant species, such as A. thaliana, O. sativa, Vitis vinifera, M. truncatula, B. rapa G. max, S. lycopersicum, and Populus tremula. lncRNAdb provides comprehensive annotations for eukaryotic lncRNAs, including evolutionary conservation, gene expression data, genomic context, structural information, functional evidence, subcellular localization, and the transcript sequence and their experimentally verified biological functions [64]. NONCODE (V6) is an interactive database that presents a complete collection and annotation database of ncRNAs in 39 species, including 23 plants, such as Arabidopsis with 3796 lncRNAs, O. sativa with 1123, and Z. mays 4279 [62]. It provides basic information about lncRNA expression profiles and predicted functions. GREENC provides a comprehensive annotation of more than 120,000 plant-specific lncRNAs from 37 plant species. This database provides an inbuilt pipeline for lncRNA prediction and evaluates coding potential and folding energies of lncRNAs [63]. PLNlncRbase is a repository of 1187 experimentally verified lncRNAs in 43 plant species collected from approximately 200 publications [67]. CANTATAdb includes several computationally identified lncRNAs from 10 model plant species [66]. PlncRNADB contains 5000 lncRNAs from Arabidopsis, Z. mays, and Populus, serving as a reference database for lncRNA prediction and evaluating the coding potential of protein transcripts. It also contains several computational pipelines for lncRNAs. It is worth mentioning that PlncRNADB displays the relationships between lncRNAs and various RNA-binding proteins (RBPs), which can be visualized as lncRNA–protein networks [68]. The EVLncRNAs database contains annotatedand experimentally validated lncRNAs—1543 lncRNAs from 77 species, including 428 plant lncRNAs from 44 plant species developed by Zhou et al. [69].
3. Classification of lncRNAs
lncRNA classification is a challenging task that is under active development and is so far based on several distinct features—genomic location and context, mechanism of function, targeting mechanism, and effect exerted on DNA sequences. However, the classification of lncRNAs is immature because their state of annotations is at its beginning, and classifications based on transcript properties and their length, the orientation of their regulatory elements in relation to known genomic annotations and functions have been proposed [70]. We summarize lncRNA classification based on genomic context, including the orientation relative to nearby protein-coding genes and function modes.
3.1. Based on Genome Context
Based on the genomic context, lncRNAs are transcribed from various genomic loci, including sense and antisense orientations of coding genes, intergenic regions, promoter regions, and enhancers [12]. These lncRNAs are categorized as natural intronic RNAs (incRNAs), antisense transcripts (lncNATs), bidirectional lncRNAs (BI-lncRNAs), overlapping lncRNAs (OT-lncRNAs), and long intergenic ncRNAs (lincRNAs) (Figure 2) [71]. LncRNAs transcribed from protein-coding regions of a gene and of which the promoter region may be retained from a few or all of the protein-coding exons by alternative splicing are designated as sense lncRNAs (Figure 2A). Those originating exclusively from the intronic region, sometimes including the exonic region, of protein-coding genes are known as intronic lncRNAs (Figure 2B) [72]. Sense lncRNAs lack protein-coding potential; their expression levels are lower than their corresponding mRNAs [73]. COLDAIR is a well-studied example of sense and intronic lncRNAs in plants. Intergenic lncRNA is transcribed from intergenic regions of protein-coding genes either in a sense or an antisense orientation (Figure 2C). Such lncRNAs act as a precursor for other ncRNAs. A total of ~2000 intergenic lncRNAs have been identified in various plant species, which are unique, functionally diverse, and are a major focus of lncRNA research. ENOD40 [74] and LDMAR [47] are the two most famous examples of such lncRNAs. Those lncRNAs that originate in the relatively opposite direction to the protein-coding gene are categorized as natural antisense transcripts (NATs) or antisense lncRNAs (Figure 2D) [33].
NATs play a major role in the transcriptional and post-transcriptional regulation of their corresponding protein-coding gene. NATs may act in cis or in trans to regulate gene expression or the array of functions in plants. COOLAIR is a cis-acting antisense lncRNA required for regulating flower time through the silencing of FLC [75]. Hidden treasure 1 (HID1) is a trans-acting gene involved in photomorphogenesis in Arabidopsis seedlings by suppressing phytochrome-interacting factor 3 (PIF3), a key repressor of photo-morphogenesis [76]. However, other types of lncRNAs also exist based on their origin in the genome. These include bidirectional lncRNA, enhancer lncRNA, and promoter-associated lncRNA (Figure 2E–G). Bidirectional lncRNAs are transcribed from transcripts in close proximity (<1000 bp) to the transcription start site of protein-coding genes but proceed in the opposite direction (Figure 2E) [77]. Enhancer lncRNAs are synthesized from the enhancer region of protein coding genes with or without a poly-(A) tail and mediate transcriptional regulation of protein coding genes (Figure 2F) [78]. Lastly, lncRNAs originate from promoter region protein coding genes and control the regulation of corresponding protein coding genes (Figure 2G) [79]. COLDWRAP is a 316-nt lncRNA, transcribed 225 nt upstream from the FLC gene [80].
3.2. Based on the Criteria of Functional Mode
Increasing evidence suggests that lncRNAs play various pivotal roles during plant life, including roles in chromatin remodeling, molecular imprinting, as transcriptional regulators, in RNA processing, and as ribonucleoprotein regulators. Here, we briefly summarize the classification of lncRNAs based on their function reported to date as signals, decoys, guides, scaffolds, and precursors for small interfering RNAs (siRNAs) (Figure 3).
lncRNAs may act as molecular signals in distinct biological processes and integrate developmental cues due to their specificity and diverse spatiotemporal expression in response to different biological stimuli. For instance, In Arabidopsis, COLDAIR is associated with the repression of FLC by recruiting polycomb repressive complex 2 (PRC2). This complex includes the curly leaf (CLF) subunit, possessing histone methyltransferase activity and catalyzing H3K27me3 deposition at FLC [21]. A drought-responsive lncRNA, drought induced lncRNA (DRIR), acts to regulate the activity of the FUT4 gene encoding fucosyltransferase, resulting in enhanced tolerance to drought (Figure 3A) [81].
lncRNAs may act as decoys, in the form of negative transcription regulators. They function as miRNA target mimics and titrating target proteins or regulators. In maize, degradome analysis revealed that 58 lncRNAs act as decoys. In Arabidopsis, lncRNAs—including IPS1, involved in inorganic P (Pi) homeostasis with PHO2 [45], and the alternative splicing competitor (ASCO) lncRNA—have been demonstrated to interact with miRNAs [82]. Similarly, lncRNAs are predicted to be target mimics for tomato miRNAs in response to infection with TYLCV [83] (Figure 3B).
Some lncRNAs serve as guides to bind or direct ribonucleoprotein complexes to specific target regions to regulate gene expression. COLDAIR directs the PRC2-mediated histone H3Lys27 methylation at FLC during vernalization in Arabidopsis (Figure 3C) [84]. Likewise, the ELF18-induced long-noncoding RNA 1 (ELENA1) lncRNA induced by infection of Pseudomonas syringae acts in cis/trans, imparting plant immunity through pathogenesis-related gene 1 (PR1) [24]. Several lncRNAs may serve in the scaffolding mechanism as a central podium for the assembly of various molecular components. APOLO, a polymerase II/V transcribed lncRNA, regulates chromatin loop formation (Figure 3D) [26].
lncRNAs may act as precursors or substrates for siRNAs, negatively regulating the target gene (Figure 3E) [85]. Among these, trans-acting small interfering RNAs (tasiRNAs) or phased secondary small interfering RNAs (phasiRNAs) are two endogenous classes of siRNAs [85,86]. Recently, various studies on lncRNA-derived siRNA havce been reported; among these, the miR390-TAS3-ARF pathway [87], the miR3954-lncRNA-phasiRNA-NAC pathway [88], the mul-miR3954-MuLnc1-si161579-MuCML27 [89], and miR2118-PMS1T-phasiRNAs [90] are well known in various plant species. For instance, a novel lncRNA, MuLnc1, was targeted by miR3954 to produce 21 nt phasiRNAs in mulberries. Among these phasiRNAs, si16157 can cleave calmodulin-like protein gene MulCML27, which plays an important role in response to stress. This suggests that from the complex regulatory network of ul-miR3954-MuLnc1-si161579-MuCML27, MuLnc1 may be considered as a candidate target lncRNA for genetic improvement in mulberries [89].
4. Role of lncRNAs in Plant Development
Currently, lncRNAs have become a hot topic in the molecular biology community. In plants, lncRNAs belong to a novel class of ncRNAs, and the regulatory mechanism of lncRNAs is scarce. Only 1% of plant-annotated lncRNAs have been characterized, and the emerging shreds of evidence support the fact that lncRNAs play a crucial role as regulators in a wide range of biological processes, including in regulation of mRNA processing and transcription mechanisms, nutrient metabolism, flowering, and male sterility through transcriptional modulation of the target transcript.
4.1. lncRNAs Regulate Flowering
Reproductive success of plants tightly linked to the proper timing of floral transition and morphogenesis. Flowering in the plants is controlled by various internal and external cues such as hormones, day length, and temperature. In Arabidopsis, vernalization downregulates the expression of the FLC gene to promote flowering in spring. Several lncRNA—COLDAIR, COOLAIR, and COLDWRAP—are involved in the fine-tuning of FLC expression. COLDAIR and COOLAIR are cold-inducible natural intronic lncRNAs of the MADS-box floral repressor transcription factor, FLC, which may participate in epigenetic reprogramming and serve as a transcription regulator, respectively [46,84]. COLDAIR is a sense lncRNA originating from a cryptic promoter, harboring the first intron of FLC. This promoter becomes active upon FLC repression. In cold conditions, COLDAIR recruits the PRC2 protein for epigenetic silencing of FLC through histone deacetylation of H3K9 and trimethylation of H3K27 modification at the floral repressor gene FLC [91].
In contrast, COOLAIR is an overlapping antisense-lncRNA which has polyadenylation properties for the FLC locus. The proximal 3′ polyadenylation downregulates FLC expression through H3K4 demethylation of FLC in the cold response, whereas distal polyadenylation activates FLC transcript accumulation [92]. AtNXD (neoxanthin-deficient 1) is a homeodomain protein with a divergent domain, localized to a heterochromatic region in the COOLAIR promoter. AtNXD directs and stabilizes R-loop formation in the COOLAIR promoter, leading to its downregulation and FLC accumulation and facilitating flowering [93]. However, during vernalization, COLDAIR directs the suppression of the FLC locus, whereas the repression depends on COOLAIR [94]. Therefore, during warm days AtNXD facilitates the suppression of COOLAIR while both COOLAIR and COLDAIR activate the PRC2 protein as an epigenetic modifier, thus promoting FLC expression (Figure 4A). COLDWRAP is transcribed from the promoter of FLC, recruiting PRC2 to repress FLC [80]. MAS is a cis-acting cold-inducible NAT lncRNA of MADS affecting flowering 4 (MAF4), localized in nucleus. MAS has been found to be present in the MAF4 locus, which extends from the 5′ transcriptional termination site (TTS) to the first intron at the 3′ end. MAS contains a polyadenylation site to bind and recruit WD40-containing repeat 5a (WDR5a) to MAF4 sites, thus enhancing H3K4me3 and activating MAF4 expression, fine-tuning the flowering time [95]. CDF5 long non-coding RNA (FLORE) is a natural antisense transcript lncRNA of cycling DOF factor 5 (CDF5). CDF5 is involved in circadian clock delay flowering by repressing flowering locus T (FT) transcription. FLORE/CDF5 antiphasic expression reflects mutual inhibitory regulation, which directly affects flowering time regulation. However, FLORE directs the activation of FT through repressing CDF that promotes flowering [96]. The molecular mechanism underlying this antiphasic regulation needs to be identified in detail.
4.2. lncRNAs as a Mediator of Reproductive Development
The floral meristem develops into different floral organs. With advances in functional genomics, especially in RNA-sequencing, researchers have discovered several lncRNAs that control floral and reproductive development. These lncRNAs include early flowering-completely dominant (Ef-cd), LDMAR, photoperiod-sensitive genic male sterility T (PMS1T), and LINC-AP2 [47,90,97,98]. LINC-AP2 is an intergenic lincRNA of APETALA2 (AP2) [98]. Plants with Turnip crinkle virus (TCV) infections showed reduced expression of AP2 and showed elevated expression of LINC-AP2, causing floral structure distortion. Brassica campestris male fertility 11 (BcMF11) is critical for pollen development and male fertility [23]. LncRNA AG-incRNA4 is transcribed from the second intron of the agamous (AG) gene in Arabidopsis and is involved in PRC2 complex component CLF-mediated epigenetic modification of AG, thereby contributing to repression of AG [99].
In dioecious liverwort, Marchantia polymorpha, an MYB-type transcription factor, female gametophyte MYB (MpFGMYB), plays a critical role in female sexual differentiation. Loss-of-function MpFGMYB results in female-to-male sex conversion. Suppressor of feminization (SUF), a cis-acting antisense lncRNA of MpFGMYB, is recognized as an important regulator of male sexual differentiation and can suppress its expression only in cis. During sex differentiation, the constitutive expression of SUF suppresses MpFGMYB expression, encouraging the accumulation of an unknown factor (M), thus subsequently activating Y-chromosome and autosomal genes to develop male gametophyte. On the other hand, a dominant “FEMINIZER” on the X chromosome acts through an unknown mechanism to suppress SUF expression. Subsequently, it activates the expression of MpFGMYB and autosomal and X-chromosomal genes to promote female sexual differentiation. The MpFGMYB/SUF locus acts as a bidirectional transcriptional toggle for sex differentiation in liverworts [100]. However, the mechanism by which the SUF lncRNA suppresses MpFGMYB expression remains uncertain.
A novel lncRNA named LDMAR has been suggested to regulate photo-sensitive male sterility in rice [47]. Moreover, a spontaneous point mutation leads to altered RNA secondary structure and subsequently increases DNA methylation in the LDMAR promoter region, which results in programmed cell death in developing anthers. Furthermore, a siRNA named Psi–LDMAR is derived from the promoter region of LDMAR involved in DNA methylation and represses its expression through RNA-dependent DNA methylation (RdDM) [101]. Genome-scale RNA-seq analysis predicted several lncRNAs, and DNA methylation-regulated lincRNAs from ripening tomatoes have suggested their regulatory role during fruit development and ripening [102,103]. Yu et al. [104] suggest that lncRNA2155 inactivation delays fruit ripening.
4.3. Modulating Other Developmental Processes
lncRNAs play a critical role in protein structure and function modification and cellular localization of target proteins. ENOD40, a 700-nucleotide-long lncRNA, participates in nodulation specifically in legumes, and is found in several non-legumes. It encodes two short peptides (ENDO40-I: 13 amino acid, ENDO-II: 27 amino acid) that bind to sucrose synthase. In this context, this suggested the role of ENOD40 in sucrose synthase for utilization of sucrose during nodulation [105]. M. truncatula ENOD40 binds to RNA binding protein 1 (RBP1), facilitating its translocation from the nucleus to the cytoplasm. However, Arabidopsis RBP1 plays a critical role in ABA-mediated stress tolerance and development (Figure 4B) [106].
Hidden treasure 1 (HID1) is the first known nuclear-localized lncRNA (236 nucleotides) that negatively regulates photomorphogenesis by binding directly to phytochromeinteracting factor 3 (PIF3), a repressor in photomorphogenesis [76]. auxin regulated promoter loop (APOLO) intergenic lncRNA, located about 5 kb upstream of PINOID (PID), a key polar auxin regulator, is transcribed by RNA Polymerase -II and -V. APOLO controls the PID gene expression and activates the formation of a chromatin loop surrounding the PID promoter. Auxins promote the unwinding of the APOLO-PID chromatin loop by triggering the demethylation of APOLO. RNA Pol II facilitates like heterochromatin protein 1 (LHP1) for loop formation. The APOLO transcripts produced by RNA Pol II/V recruit PRC1 and ARGONAUTE4 (AGO4) for loop closing and triggering DNA methylation. Finally, the APOLO lncRNA-mediated chromatin loop is reformed and represses PID (Figure 4C) [26]. MIKKI is a four-intron-containing retrotransposon element that controls root development by regulating the scarecrow like (SCL) gene expression. MIKKI specifically binds to miR171 to mimic its expression, thus interpreting its root development role [107]. Transcriptome analysis of flower and fruit tissues in Poncirus trifoliata, Fragaria vesca, and Cicer arietinum demonstrated tissue-specific expression of lncRNAs, indicating their potential role in regulating floral development and fruit ripening [108,109,110].
Conclusively, these findings suggesting the critical functions of lncRNAs in plant reproductive growth through different regulatory mechanisms. The lncRNAs, as mentioned earlier from various plant species, display high tissue-specific expression patterns, suggesting that lncRNAs might play a critical regulatory role in plant development. It is necessary to carry out further studies on the function and mechanism of lncRNAs (Table 3).
5. Role of lncRNAs in Biotic and Abiotic Stress
Globally, changes in the environment due to erratic patterns of seasons, rainfall, temperature, and pathogens affect and pose a serious challenge to global food security. Plants have adopted dynamic responses to such abiotic stresses through various complex molecular networks, including biochemical and physiological cascades of different signal transduction pathways [111]. Subsequently, discovering stress-responsive lncRNAs with their target genes in various plant species has enabled us to understand the molecular mechanism underlying such stress adaptations. Moreover, the large-scale identification of lncRNAs has revealed their participatory roles in stress responses in plants. Such studies have established the pivotal role of ncRNAs in mediating stress tolerance at transcriptional, post-transcriptional modification, and epigenetic levels. As mentioned, a total of 1187 experimental-verified stress-related lncRNAs from 43 plant species have been listed on PLNlncRbase under 17 different biotic and abiotic conditions [67]. In contrast, the regulation mechanism of lncRNAs in response to biotic and abiotic stresses, which allows the plant to survive in a harsh environment, needs to be explored further (Figure 5).
5.1. lncRNAs in Drought Stress
Emerging evidence suggests that lncRNAs could directly affect drought responses by regulating the transcription of various stress-responsive genes or by recruiting complex mechanisms such as endogenous target mimics (eTM), chromatin modulation, and antisense transcription-mediated modulation [30]. Recently, genome-wide transcriptome analysis has revealed a variety of stress-responsive lncRNAs in various plant species, including six in Arabidopsis [112], Populus [42], rice [113], maize [114], foxtail millet [115], cassava [40], and switchgrass [116]. Systematic RNA-seq analysis of P. trichocarpa provided the opportunity to explore 2500 lncRNAs of which ~504 lncRNAs were categorized as drought-responsive [42]. To survey the drought-responsive lncRNAs in foxtail millet, deep sequencing served to identify ~584 lncRNAs, of which 17 lincRNAs and 2 NAR lncRNAs were droughts responsive [115]. In cassava, strand-specific RNA-seq data revealed 153 NAT lncRNAs and 318 lncRNAs responding to drought and cold stress, respectively [40]. Considering the regulatory mechanism of lncRNAs, 98 lncRNAs were explored in a comprehensive landscape of the rice genome. Among them, two key drought-responsive NAT lncRNAs—Os02g0250700–01, targeting the late embryogenesis abundant protein gene, and Os02g0180800–01, targeting the cinnamoyl-CoA reductase gene—were also recovered [113].
Moreover, eTM lincRNA340 acts as a target mimic for miR169, which controls the expression of the nuclear factor Y (NF-Y) gene under drought conditions (Figure 5A) [40]. lncRNAs may regulate the transcriptional expression of various genes. One possible role of lncRNAs as a transcriptional regulator in response to drought has been explored in Arabidopsis. A 755-nt-long nuclear-localized lncRNA designed as drought-induced lncRNA (DRIR) regulates several drought-responsive genes such as those involved in ABA-signaling (P5CS1, RD29A/B, and ABI5); annexin (ANNAT7); and aquaporin genes (TIP4, NIP1). Moreover, DRIR may act to influence the activity of fucosyltransferase 4 (FUT4) or transcription factor NAM/ATAF/CUC 3 (NAC3), resulting in improved tolerance to salinity and drought stresses (Qin et al., 2017), suggesting that lncRNA DRIR may confer drought tolerance by acting as a positive regulator. Conclusively, the emerging drought stress-responsive lncRNAs could act as a target mimic for various stress-responsive miRNAs that regulate different drought-responsive genes and transcription factors. These lncRNAs may act as a critical regulatory hub for various drought-responsive signal transduction pathways at various transcriptional and post-transcriptional levels.
5.2. lncRNAs in Salinity Stress
Like other abiotic stresses, salinity stress is one of the major limiting factors for plant growth and productivity. With the aid of “omics” approaches, the role of lncRNAs has been investigated in various plant species. For instance, 3030 lincRNAs and 275 lncNATs were identified from salt-stressed roots of soybean plants [117]. Salt stress also altered the accumulation of lncRNAs in Arabidopsis [118] and Medicago [119]. Moreover, in grapevine and C. sinensis, 1661 lncRNAs and 172 lncRNAs were found to be differentially expressed in roots under salt stress, respectively [120,121]. Similarly, in cotton, 44 lncRNAs were differentially expressed, of which lncRNA973 was observed to regulate salt stress-responsive genes, including those involved in the ROS scavenging system (Figure 5B) [122]. Recently, a total of 26 lncRNAs in salt-tolerant M-81E and 133 differentially expressed lncRNAs in salt-sensitive Roma strains of sweet sorghum were identified under salt-stress [123]. Strand-specific RNA-sequencing of salt-stressed root tissues of two closely related poplars (Populus euphratica and Populus alba var. pyramidalis) revealed a total of 10,646 and 10,531 lncRNAs in each [124]. Taken together, the identification of salt-responsive lncRNAs provides an opportunity to explore and improve our understanding of lncRNA-mediated gene regulation.
5.3. lncRNAs Controlling Nutrient Stress
The availability of certain nutrients in the soil can have severe effects on plant growth and development. Nutrient acquisition from the soil is one of the most critical physiological processes and mechanisms, including various membrane transporters involved in nutrient homeostasis [125]. Phosphorus (P) is one of the essential macronutrients contributing to plant growth and development, acting as a growth-limiting factor, and as a source of inorganic P (Pi) for ATP production [126]. However, the complex regulatory machines underlying P homeostasis in plants is scarce. Furthermore, plants have evolved several molecular and physiological mechanisms to improve Pi availability and P usage efficiency [127]. The role of lncRNAs in controlling phosphate availability in plants has been studied in several model plants such as Arabidopsis and rice [128,129]. In Arabidopsis, strand-specific RNA sequencing analysis of Pi-sufficient (P+) and Pi-deficient (P−) plants facilitated the identification of ~ 1212 lncRNAs, of which ~ 309 lncRNAs were differentially expressed under the mentioned conditions [128]. Npc536 is another lncRNA in Arabidopsis induced in leaves and roots by phosphate starvation and drought, which also affects root growth under salt stress conditions [112].
PHOSPHATE2 (PHO2) is critical for Pi acquisition, translocation, and homeostasis in Arabidopsis, and miR399 mimics its expression [130]. PHO2 expression is indispensable for Pi homeostasis when the Pi supply is ample. Upon Pi starvation, PHO2 is suppressed by an activated level of miR399, which directs the cleavage of PHO2 and thereby facilitates Pi uptake and translocation from root to shoot (Figure 5C). In Arabidopsis, the lncRNA IPS, sharing 23 nt complementary binding sites with miR399, acts as a target mimic for miR399 and controls the accumulation of target gene PHO2 under Pi starvation, facilitating its uptake [45]. In this context, lncRNA IPS binds to miR399, preventing it from acting on its target gene PHO2 when Pi is abundant in the soil. Elevated levels of PHO2 abolish the functions of phosphate transporter 1 (PHR1) [25], thereby preventing Pi toxicity in roots by maintaining an optimum uptake of Pi. Under P− conditions, miR399 directs the cleavage of the PHO2 transcript, which presumably activates PHR’s function and eventually allows for Pi accumulation in the root. In M. truncatula, three lncRNAs—known as phosphate deficiency induced lncRNAs (PDILs)—have been identified to regulate Pi-deficiency signaling. Among them, PDIL1 suppresses the degradation of MtPHO2 by miR399 to regulate Pi deficiency signaling and Pi transportation, whereas PDIL2/3 is involved in transcriptional regulation of Pi homeostasis [131].
Nitrogen (N) is another key essential nutrient for plant growth, development, and reproductive success. N serves as a major component of amino acids, energy-carrying molecule ATP (adenosine triphosphate), and N metabolism [132]. It is well established that N deficiency severely affects plant productivity, but molecular and genetic approaches have identified several regulatory and transporter genes controlling N use efficiency to help to improve plant productivity [133]. However, the molecular mechanism of N assimilation is not entirely known. Advancements in RNA-sequencing have further expanded our understanding of N-responsive lncRNAs’ roles in N homeostasis in plants. Studies showed that changes in N supply altered the expression of lncRNAs in rice [38], Populus [133], barley [134], and maize [41]. A study on lncRNAs in the rice genome under nitrogen starvation revealed the role of two cis-NAT-lncRNAs, cis-NATAMT1.1 and cis-NATAMT1.2, derived from the sense strand of AMT1 [38]. The regulatory mechanism of the nitrogen (N)-responsive lincRNA trans-acting siRNA3 (TAS3), targeting the auxin response factor 4 (ARF4) gene, is well understood in Arabidopsis [87], whereas its possible new target—nitrate transporter (NRT)—needs further validation [135]. Similarly, a nitrate-induced lncRNA, T5120, is regulated by NLP7, a master regulator of nitrate signaling. NLP7 directly binds to nitrate-responsive cis-element (NRE)-like the motif in T5120 and activates its transcription. Moreover, the expression of nitrate responsive gene NRT1.1 was also upregulated. These findings suggest the NLP7-T5120-NRT regulatory module’s role in nitrate signaling and subsequently in improved nitrate assimilation and nitrogen use efficiency (Figure 5D) [136].
Among various micronutrients, Boron (B) is an essential micronutrient required for growth and development, in processes such as phenolic metabolism, nitrate assimilation, membrane integrity, and cell wall biosynthesis. The emerging evidence suggests that B’s molecular regulation is complicated, limited to coding genes, and attributed to lncRNAs, a novel regulatory player in the regulation of gene expression. A genome-scale exploration of lncRNAs in response to B toxicity detected ~8000 lncRNAs [137], whereas under B deficiency conditions in P. trifoliata 2101 lncRNAs were detected, of which 729 were upregulated and 721 were downregulated [138].
5.4. lncRNAs as Candidate Player in Cold Stress Tolerance
Temperature, either low or high temperature, is also essential to plant growth and productivity as a limiting factor. High-throughput RNA-sequencing has enabled researchers to elucidate several quantitative trait locus (QTLs) and various regulatory genes involved in cold tolerance in plants [139]. Likewise, several lncRNAs have emerged as potential genes involved in cold tolerance, and this has been well documented in various plant species. For instance, in Arabidopsis, weather-dependent reproductive-process-related lncRNAs COOLAIR, COLDWRAP, and COLDAIR facilitate the induction of flowering in the appropriate season [46,80,93].
Vernalization is a well-characterized phenomenon in flowering plants, causing them to flower in the favorable conditions of spring [140]. In Arabidopsis, FLC is a repressor of flowering. COOLAIR, a NAT-lncRNA transcribed from the sense transcript of FLC, controls FLC repression during vernalization [141]. COLDAIR, transcribed from the first intron of the FLC gene, represses FLC locus’s expression through PRC2-mediated chromatin modification [46]. The FLC-promoter-derived lncRNA COLDWRAP could further interact with stable repression of the FLC during vernalization [140]. The operating mechanisms of cold-induced SVALKA and cryptic antisense CBF1 (asCBF1) lncRNAs have further advanced our understanding of the cold acclimation mechanism in plants [142]. In response to cold, MAS activated transcription of the corresponding cold responsive MAF4, recruiting the WDR5a complex for epigenetic modification at the MAF4 locus for its activation and subsequently suppressed flowering [95].
5.5. lncRNAs as Emerging Regulators in Plant Responses to Biotic Stresses
Plants have evolved a multi-layered defense system to counteract pathogens through inducible or constitutive defenses. These defense responses include physical/chemical barriers, pattern recognition receptors (PRRs), and nucleotide-binding site–leucine-rich repeats (NBS-LRRs). In contrast, immunity-related lncRNAs are less documented in the plants. However, recent advanced sequencing technology has disclosed the role of lncRNAs in plant–pathogen interactions. For instance, in tomatoes, NBS-LRRs are a major regulatory component of the response to pathogen invasion. miR482b regulates NBS-LRR genes during infection. lncRNA23468 prevents the pairing of miR482b and encourages the accumulation of NBS-LRRs in tomato plants infected with P. infestans [143]. A very recent example is ELENA1 lncRNA, which enhances resistance against disease caused by P. syringae and is involved in imparting plant immunity [24]. Similarly, lncRNA S-slylnc0957 acts as a negative regulator of TYLCV infection [144]. In wheat, 125 lncRNAs were responsive to powdery mildew infection [145]. Cui et al. [146] demonstrated that tomato lncRNA16397 is an antisense lncRNA transcript of SlGRX that enhances resistance to P. infestans by reducing reactive oxygen species (ROS) accumulation. Many other biotic stress-related lncRNAs have been identified, but their biological roles are yet to be investigated.
miR482 acts as a negative regulator of NBS-LRR genes [147]. Cui et al. [148] reported that 88 lncRNAs predicted to decoy 46 miRNAs could target 30 coding genes, particularly NBS-LRR genes [147]. Among these lncRNA-miRNA-mRNA regulatory modules, Sl-lncRNA15492-miR482-NBS-LRR affects innate immunity by maintaining Sl-NBS-LRR homeostasis. [149]. The Sl-lncRNA15492 lncRNA is 511 bp long, and pre-Sl-miR482a was located on the antisense sequence. Sl-lncRNA15492 acts as a positive regulator and Sl-miR482a acts as a negative regulator of tomatoes’ resistance to P. infestans. In this context, Sl-lncRNA15492 suppresses the expression of pre-Sl-miR482a to relieve the suppression of NBS-LRR genes and enhance tomato resistance against infection. However, when optimum levels of Sl-NBS-LRR accumulate, Sl-miR482a cleaves Sl-lncRNA15492, leading to an abundance of mature Sl-miR482a and decreasing the accumulation of NBS-LRR (Figure 5E) [149].
Considering the mounting evidence of lncRNAs in various biotic and abiotic stress responses in several plant species, researchers have provided novel insights into stress-responsive lncRNAs (SR-lncRNAs). However, the mechanisms involved in controlling the regulation of SR-lncRNAs are largely unknown and need further research. Table 3 presents several identified lncRNAs, along with their possible targets and mechanism of action contributing to the biotic and abiotic stress response in plants (Table 3).
6. Conclusions and Future Prospective
lncRNAs are among the most important types of ncRNAs emerging as potentially important players in various fundamental biological processes in both animals and plants. The development of high-throughput screening and other new techniques has helped to tailor the identification of lncRNAs and their biology. By contrast, our understanding of the regulation of synthesis and biogenesis of lncRNAs in plants remains scant. To date, the roles of very few plant lncRNAs have been explored. It has been found that lncRNAs are involved in induced responses to various abiotic and biotic stresses and that they play a crucial role in regulating gene expression during stress responses. The fine regulation of lncRNAs is essential for functional stress responses, and various studies have reported that the regulation of lncRNAs is associated with an array of responses. Emerging evidence suggests their regulatory roles in morphogenesis, tolerance to stress, response to pathogenic infection, and so on. As research into lncRNAs moves forward, aiming to uncover the regulatory mechanisms by which these lncRNAs act, novel functions in addition to those described here will likely be uncovered.
RNA-binding protein interactions play a pivotal role throughout the life of plants and are crucial for maintaining cellular processes and pathogenesis. LncRNA interactions with proteins may prove useful, and identifing proteins associated with lncRNA-mediated biological processes is critical. Consequently, the extension of sophisticated approaches like RNA-binding would undoubtedly improve our understanding of lncRNA biology and their systematic characterization. Loss and gain of function, RNA interference (RNAi), virus-induced gene silencing (VIGS), and recently developed systems such as transcription activator-like effector nucleases (TALENs) and clustered regulatory interspaced short palindromic repeats (CRISPR)/Cas9 systems can be effectivity employed to determine the precise function of lncRNAs in plants. Therefore, we suggest that lncRNAs may be developed into valuable molecular biology tools in plants to effectively control their response to various stresses. Increasing our understanding of precisely how lncRNAs regulate growth and development and stress responses will help to identify novel targets that will pave a path for developing new strategies for many plant processes.
Author Contributions
M.W. conceived the idea and wrote the manuscript; R.X. and Y.L. revised the manuscript. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
No new data or existing data were created/used or analyzed in this study.
Acknowledgments
This work was funded by the National Key Research and Developmental Program of China (2018YFD1000104). This work was also supported by awards from the National Natural Science Foundation of China (#31872063), the Special Support Program of Guangdong Province (2019TX05N193).
Conflicts of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Abbreviations
| lncRNAs | Long non-coding RNAs |
| sRNAs | Small RNAs |
| miRNAs | MicroRNAs |
| ENOD4 | Early nodulin 40 |
| Igf2 | insulin-like growth factor 2 |
| TYLCV | Tomato yellow leaf curl virus |
| SAGE | Serial expression of gene expression |
| EST | Expressed sequence tag |
| NATs | Natural antisense transcripts |
| TTS | Transcriptional termination site |
| TCV | Turnip crinkle virus |
| SCL | Scarecrow like |
| NF-Y | Nuclear factor Y |
| N | Nitrogen |
| B | Boron |
| P | Phosphorous |
| Pi | inorganic Phosphorous |
References
- Ariel, F.; Romero-Barrios, N.; Jegu, T.; Benhamed, M.; Crespi, M. Battles and hijacks: Noncoding transcription in plants. Trends Plant Sci. 2015, 20, 362–371. [Google Scholar] [CrossRef]
- Doolittle, W.F. Is junk DNA bunk? A critique of ENCODE. Proc. Natl. Acad. Sci. USA 2013, 110, 5294–5300. [Google Scholar] [CrossRef] [Green Version]
- Liu, G.; Mattick, J.S.; Taft, R.J. A meta-analysis of the genomic and transcriptomic composition of complex life. Cell Cycle (Georget. Tex.) 2013, 12, 2061–2072. [Google Scholar] [CrossRef] [Green Version]
- Quan, M.; Chen, J.; Zhang, D. Exploring the secrets of long noncoding RNAs. Int. J. Mol. Sci. 2015, 16, 5467–5496. [Google Scholar] [CrossRef] [Green Version]
- Moseley, M.L.; Zu, T.; Ikeda, Y.; Gao, W.; Mosemiller, A.K.; Daughters, R.S.; Chen, G.; Weatherspoon, M.R.; Clark, H.B.; Ebner, T.J.; et al. Bidirectional expression of CUG and CAG expansion transcripts and intranuclear polyglutamine inclusions in spinocerebellar ataxia type 8. Nat. Genet. 2006, 38, 758–769. [Google Scholar] [CrossRef]
- Ye, C.Y.; Chen, L.; Liu, C.; Zhu, Q.H.; Fan, L. Widespread noncoding circular RNAs in plants. New Phytol. 2015, 208, 88–95. [Google Scholar] [CrossRef]
- Fok, E.T.; Scholefield, J.; Fanucchi, S.; Mhlanga, M.M. The emerging molecular biology toolbox for the study of long noncoding RNA biology. Epigenomics 2017, 9, 1317–1327. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.L.; Carmichael, G.G. Decoding the function of nuclear long non-coding RNAs. Curr. Opin. Cell Biol. 2010, 22, 357–364. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chekanova, J.A. Long non-coding RNAs and their functions in plants. Curr. Opin. Plant Biol. 2015, 27, 207–216. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, K.; Shi, Z.M.; Chang, Y.N.; Hu, Z.M.; Qi, H.X.; Hong, W. The ways of action of long non-coding RNAs in cytoplasm and nucleus. Gene 2014, 547, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Sanchita; Trivedi, P.K.; Asif, M.H. Updates on plant long non-coding RNAs (lncRNAs): The regulatory components. Plant Cell Tissue Organ Cult. (PCTOC) 2020, 140, 259–269. [Google Scholar] [CrossRef]
- Rai, M.I.; Alam, M.; Lightfoot, D.A.; Gurha, P.; Afzal, A.J. Classification and experimental identification of plant long non-coding RNAs. Genomics 2019, 111, 997–1005. [Google Scholar] [CrossRef] [PubMed]
- Karlik, E.; Ari, S.; Gozukirmizi, N. LncRNAs: Genetic and epigenetic effects in plants. Biotechnol. Biotechnol. Equip. 2019, 33, 429–439. [Google Scholar] [CrossRef] [Green Version]
- Dey, M.; Complainville, A.; Charon, C.; Torrizo, L.; Kondorosi, A.; Crespi, M.; Datta, S. Phytohormonal responses in enod40-overexpressing plants of Medicago truncatula and rice. Physiol. Plant. 2004, 120, 132–139. [Google Scholar] [CrossRef] [PubMed]
- Pant, B.D.; Buhtz, A.; Kehr, J.; Scheible, W.-R. MicroRNA399 is a long-distance signal for the regulation of plant phosphate homeostasis. Plant J. 2008, 53, 731–738. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, J.; Wang, H.; Chua, N.H. Long noncoding RNA transcriptome of plants. Plant Biotechnol. J. 2015, 13, 319–328. [Google Scholar] [CrossRef] [PubMed]
- Pachnis, V.; Belayew, A.; Tilghman, S.M. Locus unlinked to alpha-fetoprotein under the control of the murine raf and Rif genes. Proc. Natl. Acad. Sci. USA 1984, 81, 5523. [Google Scholar] [CrossRef] [Green Version]
- Keniry, A.; Oxley, D.; Monnier, P.; Kyba, M.; Dandolo, L.; Smits, G.; Reik, W. The H19 lincRNA is a developmental reservoir of miR-675 that suppresses growth and Igf1r. Nat. Cell Biol. 2012, 14, 659–665. [Google Scholar] [CrossRef]
- Crespi, M.D.; Jurkevitch, E.; Poiret, M.; d’Aubenton-Carafa, Y.; Petrovics, G.; Kondorosi, E.; Kondorosi, A. enod40, a gene expressed during nodule organogenesis, codes for a non-translatable RNA involved in plant growth. EMBO J. 1994, 13, 5099–5112. [Google Scholar] [CrossRef]
- Campalans, A.; Kondorosi, A.; Crespi, M. Enod40, a short open reading frame-containing mRNA, induces cytoplasmic localization of a nuclear RNA binding protein in Medicago truncatula. Plant Cell 2004, 16, 1047–1059. [Google Scholar] [CrossRef] [Green Version]
- Swiezewski, S.; Liu, F.; Magusin, A.; Dean, C. Cold-induced silencing by long antisense transcripts of an Arabidopsis Polycomb target. Nature 2009, 462, 799–802. [Google Scholar] [CrossRef] [PubMed]
- Ma, J.; Yan, B.; Qu, Y.; Qin, F.; Yang, Y.; Hao, X.; Yu, J.; Zhao, Q.; Zhu, D.; Ao, G. Zm401, a short-open reading-frame mRNA or noncoding RNA, is essential for tapetum and microspore development and can regulate the floret formation in maize. J. Cell. Biochem. 2008, 105, 136–146. [Google Scholar] [CrossRef] [PubMed]
- Song, J.-H.; Cao, J.-S.; Wang, C.-G. BcMF11, a novel non-coding RNA gene from Brassica campestris, is required for pollen development and male fertility. Plant Cell Rep. 2013, 32, 21–30. [Google Scholar] [CrossRef] [PubMed]
- Seo, J.S.; Sun, H.-X.; Park, B.S.; Huang, C.-H.; Yeh, S.-D.; Jung, C.; Chua, N.-H. ELF18-INDUCED LONG-NONCODING RNA Associates with Mediator to Enhance Expression of Innate Immune Response Genes in Arabidopsis. Plant Cell 2017, 29, 1024–1038. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bari, R.; Datt Pant, B.; Stitt, M.; Scheible, W.-R. PHO2, microRNA399, and PHR1 define a phosphate-signaling pathway in plants. Plant Physiol. 2006, 141, 988–999. [Google Scholar] [CrossRef] [Green Version]
- Ariel, F.; Jegu, T.; Latrasse, D.; Romero-Barrios, N.; Christ, A.; Benhamed, M.; Crespi, M. Noncoding transcription by alternative RNA polymerases dynamically regulates an auxin-driven chromatin loop. Mol. Cell 2014, 55, 383–396. [Google Scholar] [CrossRef] [Green Version]
- Datta, R.; Paul, S. Long non-coding RNAs: Fine-tuning the developmental responses in plants. J. Biosci. 2019, 44, 77. [Google Scholar] [CrossRef]
- Wierzbicki, A.T.; Haag, J.R.; Pikaard, C.S. Noncoding Transcription by RNA Polymerase Pol IVb/Pol V Mediates Transcriptional Silencing of Overlapping and Adjacent Genes. Cell 2008, 135, 635–648. [Google Scholar] [CrossRef] [Green Version]
- Quinn, J.J.; Chang, H.Y. Unique features of long non-coding RNA biogenesis and function. Nat. Rev. Genet. 2016, 17, 47–62. [Google Scholar] [CrossRef]
- Jha, U.C.; Nayyar, H.; Jha, R.; Khurshid, M.; Zhou, M.; Mantri, N.; Siddique, K.H.M. Long non-coding RNAs: Emerging players regulating plant abiotic stress response and adaptation. BMC Plant Biol. 2020, 20, 466. [Google Scholar] [CrossRef]
- Subburaj, S.; Jeon, Y.; Tu, L.; Jin, Y.-T.; Kumari, S.; Lee, G.-J. Genome-wide identification, functional prediction and expression profiling of long non-coding RNAs in Camelina sativa. Plant Growth Regul. 2018, 86, 49–63. [Google Scholar] [CrossRef]
- Jin, J.; Liu, J.; Wang, H.; Wong, L.; Chua, N.-H. PLncDB: Plant long non-coding RNA database. Bioinformatics 2013, 29, 1068–1071. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, H.; Chung, P.J.; Liu, J.; Jang, I.-C.; Kean, M.J.; Xu, J.; Chua, N.-H. Genome-wide identification of long noncoding natural antisense transcripts and their responses to light in Arabidopsis. Genome Res. 2014, 24, 444–453. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, J.; Jung, C.; Xu, J.; Wang, H.; Deng, S.; Bernad, L.; Arenas-Huertero, C.; Chua, N.-H. Genome-wide analysis uncovers regulation of long intergenic noncoding RNAs in Arabidopsis. Plant Cell 2012, 24, 4333–4345. [Google Scholar] [CrossRef] [Green Version]
- Zheng, X.M.; Chen, J.; Pang, H.B.; Liu, S.; Gao, Q.; Wang, J.R.; Qiao, W.H.; Wang, H.; Liu, J.; Olsen, K.M.; et al. Genome-wide analyses reveal the role of noncoding variation in complex traits during rice domestication. Sci. Adv. 2019, 5, eaax3619. [Google Scholar] [CrossRef] [Green Version]
- Han, L.; Mu, Z.; Luo, Z.; Pan, Q.; Li, L. New lncRNA annotation reveals extensive functional divergence of the transcriptome in maize. J. Integr. Plant Biol. 2019, 61, 394–405. [Google Scholar] [CrossRef]
- Zhang, J.; Wei, L.; Jiang, J.; Mason, A.S.; Li, H.; Cui, C.; Chai, L.; Zheng, B.; Zhu, Y.; Xia, Q.; et al. Genome-wide identification, putative functionality and interactions between lncRNAs and miRNAs in Brassica species. Sci. Rep. 2018, 8, 4960. [Google Scholar] [CrossRef] [Green Version]
- Shin, S.-Y.; Jeong, J.S.; Lim, J.Y.; Kim, T.; Park, J.H.; Kim, J.-K.; Shin, C. Transcriptomic analyses of rice (Oryza sativa) genes and non-coding RNAs under nitrogen starvation using multiple omics technologies. BMC Genom. 2018, 19, 532. [Google Scholar] [CrossRef]
- Zhang, Y.C.; Liao, J.Y.; Li, Z.Y.; Yu, Y.; Zhang, J.P.; Li, Q.F.; Qu, L.H.; Shu, W.S.; Chen, Y.Q. Genome-wide screening and functional analysis identify a large number of long noncoding RNAs involved in the sexual reproduction of rice. Genome Biol. 2014, 15, 512. [Google Scholar] [CrossRef] [Green Version]
- Li, S.; Yu, X.; Lei, N.; Cheng, Z.; Zhao, P.; He, Y.; Wang, W.; Peng, M. Genome-wide identification and functional prediction of cold and/or drought-responsive lncRNAs in cassava. Sci. Rep. 2017, 7, 45981. [Google Scholar] [CrossRef]
- Lv, Y.; Liang, Z.; Ge, M.; Qi, W.; Zhang, T.; Lin, F.; Peng, Z.; Zhao, H. Genome-wide identification and functional prediction of nitrogen-responsive intergenic and intronic long non-coding RNAs in maize (Zea mays L.). BMC Genom. 2016, 17, 350. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shuai, P.; Liang, D.; Tang, S.; Zhang, Z.; Ye, C.Y.; Su, Y.; Xia, X.; Yin, W. Genome-wide identification and functional prediction of novel and drought-responsive lincRNAs in Populus trichocarpa. J. Exp. Bot. 2014, 65, 4975–4983. [Google Scholar] [CrossRef] [PubMed]
- Bhatia, G.; Goyal, N.; Sharma, S.; Upadhyay, K.S.; Singh, K. Present Scenario of Long Non-Coding RNAs in Plants. Non-Coding RNA 2017, 3, 16. [Google Scholar] [CrossRef] [Green Version]
- Wu, R.; Su, Y.; Wu, H.; Dai, Y.; Zhao, M.; Lu, Q. Characters, functions and clinical perspectives of long non-coding RNAs. Mol. Genet. Genom. MGG 2016, 291, 1013–1033. [Google Scholar] [CrossRef] [PubMed]
- Franco-Zorrilla, J.M.; Valli, A.; Todesco, M.; Mateos, I.; Puga, M.I.; Rubio-Somoza, I.; Leyva, A.; Weigel, D.; Garcia, J.A.; Paz-Ares, J. Target mimicry provides a new mechanism for regulation of microRNA activity. Nat. Genet. 2007, 39, 1033–1037. [Google Scholar] [CrossRef] [PubMed]
- Heo, J.B.; Sung, S. Vernalization-mediated epigenetic silencing by a long intronic noncoding RNA. Science 2011, 331, 76–79. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ding, J.; Lu, Q.; Ouyang, Y.; Mao, H.; Zhang, P.; Yao, J.; Xu, C.; Li, X.; Xiao, J.; Zhang, Q. A long noncoding RNA regulates photoperiod-sensitive male sterility, an essential component of hybrid rice. Proc. Natl. Acad. Sci. USA 2012, 109, 2654–2659. [Google Scholar] [CrossRef] [Green Version]
- Shin, H.; Shin, H.S.; Chen, R.; Harrison, M.J. Loss of At4 function impacts phosphate distribution between the roots and the shoots during phosphate starvation. Plant J. 2006, 45, 712–726. [Google Scholar] [CrossRef]
- Sun, Y.-M.; Chen, Y.-Q. Principles and innovative technologies for decrypting noncoding RNAs: From discovery and functional prediction to clinical application. J. Hematol. Oncol. 2020, 13, 109. [Google Scholar] [CrossRef]
- Kang, C.; Liu, Z. An Easy-to-Follow Pipeline for Long Noncoding RNA Identification: A Case Study in Diploid Strawberry Fragaria vesca. Methods Mol. Biol. (Clifton N.J.) 2019, 1933, 223–243. [Google Scholar] [CrossRef]
- Wang, J.; Meng, X.; Dobrovolskaya, O.B.; Orlov, Y.L.; Chen, M. Non-coding RNAs and Their Roles in Stress Response in Plants. Genom. Proteom. Bioinform. 2017, 15, 301–312. [Google Scholar] [CrossRef] [PubMed]
- Kong, L.; Zhang, Y.; Ye, Z.-Q.; Liu, X.-Q.; Zhao, S.-Q.; Wei, L.; Gao, G. CPC: Assess the protein-coding potential of transcripts using sequence features and support vector machine. Nucleic Acids Res. 2007, 35, W345–W349. [Google Scholar] [CrossRef] [PubMed]
- Lin, M.F.; Jungreis, I.; Kellis, M. PhyloCSF: A comparative genomics method to distinguish protein coding and non-coding regions. Bioinformatics 2011, 27, i275–i282. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Park, H.J.; Dasari, S.; Wang, S.; Kocher, J.-P.; Li, W. CPAT: Coding-Potential Assessment Tool using an alignment-free logistic regression model. Nucleic Acids Res. 2013, 41, e74. [Google Scholar] [CrossRef] [PubMed]
- Sun, K.; Chen, X.; Jiang, P.; Song, X.; Wang, H.; Sun, H. iSeeRNA: Identification of long intergenic non-coding RNA transcripts from transcriptome sequencing data. BMC Genom. 2013, 14, S7. [Google Scholar] [CrossRef] [Green Version]
- Tripathi, R.; Patel, S.; Kumari, V.; Chakraborty, P.; Varadwaj, P.K. DeepLNC, a long non-coding RNA prediction tool using deep neural network. Netw. Modeling Anal. Health Inform. Bioinform. 2016, 5, 21. [Google Scholar] [CrossRef]
- Hu, R.; Sun, X. lncRNATargets: A platform for lncRNA target prediction based on nucleic acid thermodynamics. J. Bioinform. Comput. Biol. 2016, 14, 1650016. [Google Scholar] [CrossRef]
- Furió-Tarí, P.; Tarazona, S.; Gabaldón, T.; Enright, A.J.; Conesa, A. spongeScan: A web for detecting microRNA binding elements in lncRNA sequences. Nucleic Acids Res. 2016, 44, W176–W180. [Google Scholar] [CrossRef]
- Huang, H.-Y.; Chien, C.-H.; Jen, K.-H.; Huang, H.-D. RegRNA: An integrated web server for identifying regulatory RNA motifs and elements. Nucleic Acids Res. 2006, 34, W429–W434. [Google Scholar] [CrossRef] [Green Version]
- Jiang, Q.; Wang, J.; Wang, Y.; Ma, R.; Wu, X.; Li, Y. TF2LncRNA: Identifying common transcription factors for a list of lncRNA genes from ChIP-Seq data. Biomed. Res. Int. 2014, 2014, 317642. [Google Scholar] [CrossRef] [Green Version]
- Sun, L.; Luo, H.; Bu, D.; Zhao, G.; Yu, K.; Zhang, C.; Liu, Y.; Chen, R.; Zhao, Y. Utilizing sequence intrinsic composition to classify protein-coding and long non-coding transcripts. Nucleic Acids Res. 2013, 41, e166. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Li, H.; Fang, S.; Kang, Y.; Wu, W.; Hao, Y.; Li, Z.; Bu, D.; Sun, N.; Zhang, M.Q.; et al. NONCODE 2016: An informative and valuable data source of long non-coding RNAs. Nucleic Acids Res. 2015, 44, D203–D208. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gallart, A.P.; Pulido, T.H.; De Lagrán, I.A.M.; Sanseverino, W.; Cigliano, R.A. GREENC: A Wiki-based database of plant lncRNAs. Nucleic Acids Res. 2015, 44, D1161–D1166. [Google Scholar] [CrossRef]
- Quek, X.C.; Thomson, D.W.; Maag, J.L.V.; Bartonicek, N.; Signal, B.; Clark, M.B.; Gloss, B.S.; Dinger, M.E. lncRNAdb v2.0: Expanding the reference database for functional long noncoding RNAs. Nucleic Acids Res. 2014, 43, D168–D173. [Google Scholar] [CrossRef]
- Yi, X.; Zhang, Z.; Ling, Y.; Xu, W.; Su, Z. PNRD: A plant non-coding RNA database. Nucleic Acids Res. 2014, 43, D982–D989. [Google Scholar] [CrossRef] [Green Version]
- Szcześniak, M.W.; Rosikiewicz, W.; Makałowska, I. CANTATAdb: A Collection of Plant Long Non-Coding RNAs. Plant Cell Physiol. 2015, 57, e8. [Google Scholar] [CrossRef] [Green Version]
- Xuan, H.; Zhang, L.; Liu, X.; Han, G.; Li, J.; Li, X.; Liu, A.; Liao, M.; Zhang, S. PLNlncRbase: A resource for experimentally identified lncRNAs in plants. Gene 2015, 573, 328–332. [Google Scholar] [CrossRef]
- Bai, Y.; Dai, X.; Ye, T.; Zhang, P.; Yan, X.; Gong, X.; Liang, S.; Chen, M. PlncRNADB: A repository of plant lncRNAs and lncRNA-RBP protein interactions. Curr. Bioinform. 2019, 14, 621–627. [Google Scholar] [CrossRef]
- Zhou, B.; Zhao, H.; Yu, J.; Guo, C.; Dou, X.; Song, F.; Hu, G.; Cao, Z.; Qu, Y.; Yang, Y.; et al. Experimentally Validated Plant lncRNAs in EVLncRNAs Database. In Plant Long Non-Coding RNAs: Methods and Protocols; Chekanova, J.A., Wang, H.-L.V., Eds.; Springer: New York, NY, USA, 2019; pp. 431–437. [Google Scholar] [CrossRef]
- Jarroux, J.; Morillon, A.; Pinskaya, M. History, Discovery, and Classification of lncRNAs. In Long Non Coding RNA Biology; Rao, M.R.S., Ed.; Springer: Singapore, 2017; pp. 1–46. [Google Scholar] [CrossRef]
- Nejat, N.; Mantri, N. Emerging roles of long non-coding RNAs in plant response to biotic and abiotic stresses. Crit. Rev. Biotechnol. 2018, 38, 93–105. [Google Scholar] [CrossRef]
- Ma, L.; Bajic, V.B.; Zhang, Z. On the classification of long non-coding RNAs. RNA Biol. 2013, 10, 924–933. [Google Scholar] [CrossRef]
- Cabili, M.N.; Trapnell, C.; Goff, L.; Koziol, M.; Tazon-Vega, B.; Regev, A.; Rinn, J.L. Integrative annotation of human large intergenic noncoding RNAs reveals global properties and specific subclasses. Genes Dev. 2011, 25, 1915–1927. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kouchi, H.; Takane, K.i.; So, R.B.; Ladha, J.K.; Reddy, P.M. Rice ENOD40: Isolation and expression analysis in rice and transgenic soybean root nodules. Plant J. 1999, 18, 121–129. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ietswaart, R.; Rosa, S.; Wu, Z.; Dean, C.; Howard, M. Cell-size-dependent transcription of FLC and its antisense long non-coding RNA COOLAIR explain cell-to-cell expression variation. Cell Syst. 2017, 4, 622–635.e629. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.; Fan, X.; Lin, F.; He, G.; Terzaghi, W.; Zhu, D.; Deng, X.W. Arabidopsis noncoding RNA mediates control of photomorphogenesis by red light. Proc. Natl. Acad. Sci. USA 2014, 111, 10359. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Derrien, T.; Johnson, R.; Bussotti, G.; Tanzer, A.; Djebali, S.; Tilgner, H.; Guernec, G.; Martin, D.; Merkel, A.; Knowles, D.G.; et al. The GENCODE v7 catalog of human long noncoding RNAs: Analysis of their gene structure, evolution, and expression. Genome Res. 2012, 22, 1775–1789. [Google Scholar] [CrossRef] [Green Version]
- Devaux, Y.; Zangrando, J.; Schroen, B.; Creemers, E.E.; Pedrazzini, T.; Chang, C.P.; Dorn, G.W., 2nd; Thum, T.; Heymans, S. Long noncoding RNAs in cardiac development and ageing. Nat. Rev. Cardiol. 2015, 12, 415–425. [Google Scholar] [CrossRef]
- Lubas, M.; Andersen, P.R.; Schein, A.; Dziembowski, A.; Kudla, G.; Jensen, T.H. The Human Nuclear Exosome Targeting Complex Is Loaded onto Newly Synthesized RNA to Direct Early Ribonucleolysis. Cell Rep. 2015, 10, 178–192. [Google Scholar] [CrossRef] [Green Version]
- Kim, D.-H.; Sung, S. Vernalization-Triggered Intragenic Chromatin Loop Formation by Long Noncoding RNAs. Dev. Cell 2017, 40, 302–312.e304. [Google Scholar] [CrossRef]
- Borah, P.; Das, A.; Milner, M.J.; Ali, A.; Bentley, A.R.; Pandey, R. Long Non-Coding RNAs as Endogenous Target Mimics and Exploration of Their Role in Low Nutrient Stress Tolerance in Plants. Genes 2018, 9, 459. [Google Scholar] [CrossRef] [Green Version]
- Bardou, F.; Ariel, F.; Simpson, C.G.; Romero-Barrios, N.; Laporte, P.; Balzergue, S.; Brown, J.W.; Crespi, M. Long noncoding RNA modulates alternative splicing regulators in Arabidopsis. Dev. Cell 2014, 30, 166–176. [Google Scholar] [CrossRef] [Green Version]
- Yang, Y.; Liu, T.; Shen, D.; Wang, J.; Ling, X.; Hu, Z.; Chen, T.; Hu, J.; Huang, J.; Yu, W.; et al. Tomato yellow leaf curl virus intergenic siRNAs target a host long noncoding RNA to modulate disease symptoms. PLOS Pathogens 2019, 15, e1007534. [Google Scholar] [CrossRef] [PubMed]
- Wood, C.C.; Robertson, M.; Tanner, G.; Peacock, W.J.; Dennis, E.S.; Helliwell, C.A. The Arabidopsis thaliana vernalization response requires a polycomb-like protein complex that also includes Vernalization Insensitive 3. Proc. Natl. Acad. Sci. USA 2006, 103, 14631–14636. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, H.M.; Chen, L.T.; Patel, K.; Li, Y.H.; Baulcombe, D.C.; Wu, S.H. 22-Nucleotide RNAs trigger secondary siRNA biogenesis in plants. Proc. Natl. Acad. Sci. USA 2010, 107, 15269–15274. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fei, Q.; Xia, R.; Meyers, B.C. Phased, Secondary, Small Interfering RNAs in Posttranscriptional Regulatory Networks. Plant Cell 2013, 25, 2400. [Google Scholar] [CrossRef] [Green Version]
- Marin, E.; Jouannet, V.; Herz, A.; Lokerse, A.S.; Weijers, D.; Vaucheret, H.; Nussaume, L.; Crespi, M.D.; Maizel, A. miR390, Arabidopsis TAS3 tasiRNAs, and Their AUXIN RESPONSE FACTOR Targets Define an Autoregulatory Network Quantitatively Regulating Lateral Root Growth. Plant Cell 2010, 22, 1104. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Ke, L.; Wu, G.; Xu, Y.; Wu, X.; Xia, R.; Deng, X.; Xu, Q. miR3954 is a trigger of phasiRNAs that affects flowering time in citrus. Plant J. 2017, 92, 263–275. [Google Scholar] [CrossRef]
- Gai, Y.P.; Yuan, S.S.; Zhao, Y.N.; Zhao, H.N.; Zhang, H.L.; Ji, X.L. A Novel LncRNA, MuLnc1, Associated with Environmental Stress in Mulberry (Morus multicaulis). Front. Plant Sci. 2018, 9, 669. [Google Scholar] [CrossRef] [Green Version]
- Fan, Y.; Yang, J.; Mathioni, S.M.; Yu, J.; Shen, J.; Yang, X.; Wang, L.; Zhang, Q.; Cai, Z.; Xu, C.; et al. PMS1T, producing phased small-interfering RNAs, regulates photoperiod-sensitive male sterility in rice. Proc. Natl. Acad. Sci. USA 2016, 113, 15144. [Google Scholar] [CrossRef] [Green Version]
- Crevillen, P.; Yang, H.; Cui, X.; Greeff, C.; Trick, M.; Qiu, Q.; Cao, X.; Dean, C. Epigenetic reprogramming that prevents transgenerational inheritance of the vernalized state. Nature 2014, 515, 587–590. [Google Scholar] [CrossRef] [Green Version]
- Liu, F.; Marquardt, S.; Lister, C.; Swiezewski, S.; Dean, C. Targeted 3′ processing of antisense transcripts triggers Arabidopsis FLC chromatin silencing. Science 2010, 327, 94–97. [Google Scholar] [CrossRef]
- Song, J.; Angel, A.; Howard, M.; Dean, C. Vernalization-a cold-induced epigenetic switch. J. Cell Sci. 2012, 125, 3723–3731. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Helliwell, C.A.; Robertson, M.; Finnegan, E.J.; Buzas, D.M.; Dennis, E.S. Vernalization-Repression of Arabidopsis FLC Requires Promoter Sequences but Not Antisense Transcripts. PLoS ONE 2011, 6, e21513. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.; Li, J.; Lian, B.; Gu, H.; Li, Y.; Qi, Y. Global identification of Arabidopsis lncRNAs reveals the regulation of MAF4 by a natural antisense RNA. Nat. Commun. 2018, 9, 5056. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Henriques, R.; Wang, H.; Liu, J.; Boix, M.; Huang, L.F.; Chua, N.H. The antiphasic regulatory module comprising CDF5 and its antisense RNA FLORE links the circadian clock to photoperiodic flowering. New Phytol. 2017, 216, 854–867. [Google Scholar] [CrossRef] [Green Version]
- Fang, J.; Zhang, F.; Wang, H.; Wang, W.; Zhao, F.; Li, Z.; Sun, C.; Chen, F.; Xu, F.; Chang, S.; et al. Ef-cdlocus shortens rice maturity duration without yield penalty. Proc. Natl. Acad. Sci. USA 2019, 116, 18717. [Google Scholar] [CrossRef] [Green Version]
- Gao, R.; Liu, P.; Irwanto, N.; Loh, D.R.; Wong, S.-M. Upregulation of LINC-AP2 is negatively correlated with AP2 gene expression with Turnip crinkle virus infection in Arabidopsis thaliana. Plant Cell Rep. 2016, 35, 2257–2267. [Google Scholar] [CrossRef]
- Wu, H.-W.; Deng, S.; Xu, H.; Mao, H.-Z.; Liu, J.; Niu, Q.-W.; Wang, H.; Chua, N.-H. A noncoding RNA transcribed from the AGAMOUS (AG) second intron binds to CURLY LEAF and represses AG expression in leaves. New Phytol. 2018, 219, 1480–1491. [Google Scholar] [CrossRef] [Green Version]
- Hisanaga, T.; Okahashi, K.; Yamaoka, S.; Kajiwara, T.; Nishihama, R.; Shimamura, M.; Yamato, K.T.; Bowman, J.L.; Kohchi, T.; Nakajima, K. A cis-acting bidirectional transcription switch controls sexual dimorphism in the liverwort. EMBO J. 2019, 38, e100240. [Google Scholar] [CrossRef]
- Ding, J.; Shen, J.; Mao, H.; Xie, W.; Li, X.; Zhang, Q. RNA-directed DNA methylation is involved in regulating photoperiod-sensitive male sterility in rice. Mol. Plant 2012, 5, 1210–1216. [Google Scholar] [CrossRef] [Green Version]
- Wang, M.; Zhao, W.; Gao, L.; Zhao, L. Genome-wide profiling of long non-coding RNAs from tomato and a comparison with mRNAs associated with the regulation of fruit ripening. BMC Plant Biol. 2018, 18, 75. [Google Scholar] [CrossRef] [Green Version]
- Zhu, B.; Yang, Y.; Li, R.; Fu, D.; Wen, L.; Luo, Y.; Zhu, H. RNA sequencing and functional analysis implicate the regulatory role of long non-coding RNAs in tomato fruit ripening. J. Exp. Bot. 2015, 66, 4483–4495. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, T.; Tzeng, D.T.W.; Li, R.; Chen, J.; Zhong, S.; Fu, D.; Zhu, B.; Luo, Y.; Zhu, H. Genome-wide identification of long non-coding RNA targets of the tomato MADS box transcription factor RIN and function analysis. Ann. Bot. 2019, 123, 469–482. [Google Scholar] [CrossRef] [PubMed]
- Röhrig, H.; John, M.; Schmidt, J. Modification of soybean sucrose synthase by S-thiolation with ENOD40 peptide A. Biochem. Biophys. Res. Commun. 2004, 325, 864–870. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sousa, C.; Johansson, C.; Charon, C.; Manyani, H.; Sautter, C.; Kondorosi, A.; Crespi, M. Translational and structural requirements of the early nodulin gene enod40, a short-open reading frame-containing RNA, for elicitation of a cell-specific growth response in the alfalfa root cortex. Mol. Cell. Biol. 2001, 21, 354–366. [Google Scholar] [CrossRef] [Green Version]
- Cho, J.; Paszkowski, J. Regulation of rice root development by a retrotransposon acting as a microRNA sponge. eLife 2017, 6, e30038. [Google Scholar] [CrossRef] [Green Version]
- Khemka, N.; Singh, V.K.; Garg, R.; Jain, M. Genome-wide analysis of long intergenic non-coding RNAs in chickpea and their potential role in flower development. Sci. Rep. 2016, 6, 33297. [Google Scholar] [CrossRef] [Green Version]
- Wang, C.Y.; Liu, S.R.; Zhang, X.Y.; Ma, Y.J.; Hu, C.G.; Zhang, J.Z. Genome-wide screening and characterization of long non-coding RNAs involved in flowering development of trifoliate orange (Poncirus trifoliata L. Raf.). Sci. Rep. 2017, 7, 43226. [Google Scholar] [CrossRef] [Green Version]
- Kang, C.; Liu, Z. Global identification and analysis of long non-coding RNAs in diploid strawberry Fragaria vesca during flower and fruit development. BMC Genom. 2015, 16, 815. [Google Scholar] [CrossRef] [Green Version]
- Lamaoui, M.; Jemo, M.; Datla, R.; Bekkaoui, F. Heat and Drought Stresses in Crops and Approaches for Their Mitigation. Front. Chem. 2018, 6, 26. [Google Scholar] [CrossRef]
- Amor, B.B.; Wirth, S.; Merchan, F.; Laporte, P.; d’Aubenton-Carafa, Y.; Hirsch, J.; Maizel, A.; Mallory, A.; Lucas, A.; Deragon, J.M.; et al. Novel long non-protein coding RNAs involved in Arabidopsis differentiation and stress responses. Genome Res. 2009, 19, 57–69. [Google Scholar] [CrossRef] [Green Version]
- Chung, P.J.; Jung, H.; Jeong, D.-H.; Ha, S.-H.; Choi, Y.D.; Kim, J.-K. Transcriptome profiling of drought responsive noncoding RNAs and their target genes in rice. BMC Genom. 2016, 17, 563. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, W.; Han, Z.; Guo, Q.; Liu, Y.; Zheng, Y.; Wu, F.; Jin, W. Identification of maize long non-coding RNAs responsive to drought stress. PLoS ONE 2014, 9, e98958. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qi, X.; Xie, S.; Liu, Y.; Yi, F.; Yu, J. Genome-wide annotation of genes and noncoding RNAs of foxtail millet in response to simulated drought stress by deep sequencing. Plant Mol. Biol. 2013, 83, 459–473. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Tang, G.; Peng, X.; Sun, F.; Liu, S.; Xi, Y. Long non-coding RNAs of switchgrass (Panicum virgatum L.) in multiple dehydration stresses. BMC Plant Biol. 2018, 18, 79. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, R.; Li, M.; Zhang, H.; Duan, L.; Sun, X.; Jiang, Q.; Zhang, H.; Hu, Z. Continuous salt stress-induced long non-coding RNAs and DNA methylation patterns in soybean roots. BMC Genom. 2019, 20, 730. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Di, C.; Yuan, J.; Wu, Y.; Li, J.; Lin, H.; Hu, L.; Zhang, T.; Qi, Y.; Gerstein, M.B.; Guo, Y.; et al. Characterization of stress-responsive lncRNAs in Arabidopsis thaliana by integrating expression, epigenetic and structural features. Plant J. 2014, 80, 848–861. [Google Scholar] [CrossRef]
- Wang, T.-Z.; Liu, M.; Zhao, M.-G.; Chen, R.; Zhang, W.-H. Identification and characterization of long non-coding RNAs involved in osmotic and salt stress in Medicago truncatula using genome-wide high-throughput sequencing. BMC Plant Biol. 2015, 15, 131. [Google Scholar] [CrossRef] [Green Version]
- Jin, Z.; Gao, S.; Ma, W.; Lyu, X.; Cao, X.; Yao, Y. Identification and functional prediction of salt stress-related long noncoding RNAs in grapevine roots. Environ. Exp. Bot. 2020, 179, 104215. [Google Scholar] [CrossRef]
- Wan, S.; Zhang, Y.; Duan, M.; Huang, L.; Wang, W.; Xu, Q.; Yang, Y.; Yu, Y. Integrated Analysis of Long Non-coding RNAs (lncRNAs) and mRNAs Reveals the Regulatory Role of lncRNAs Associated with Salt Resistance in Camellia sinensis. Front. Plant Sci. 2020, 11, 218. [Google Scholar] [CrossRef]
- Zhang, X.; Dong, J.; Deng, F.; Wang, W.; Cheng, Y.; Song, L.; Hu, M.; Shen, J.; Xu, Q.; Shen, F. The long non-coding RNA lncRNA973 is involved in cotton response to salt stress. BMC Plant Biol. 2019, 19, 459. [Google Scholar] [CrossRef]
- Sun, X.; Zheng, H.; Li, J.; Liu, L.; Zhang, X.; Sui, N. Comparative Transcriptome Analysis Reveals New lncRNAs Responding to Salt Stress in Sweet Sorghum. Front. Bioeng. Biotechnol. 2020, 8, 331. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, J.; Bai, X.; Luo, W.; Feng, Y.; Shao, X.; Bai, Q.; Sun, S.; Long, Q.; Wan, D. Genome-Wide Identification of Long Noncoding RNAs and Their Responses to Salt Stress in Two Closely Related Poplars. Front. Genet. 2019, 10, 777. [Google Scholar] [CrossRef] [PubMed]
- Tegeder, M.; Masclaux-Daubresse, C. Source and sink mechanisms of nitrogen transport and use. New Phytol. 2018, 217, 35–53. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Adnan, M.; Shah, Z.; Fahad, S.; Arif, M.; Alam, M.; Khan, I.A.; Mian, I.A.; Basir, A.; Ullah, H.; Arshad, M.; et al. Phosphate-Solubilizing Bacteria Nullify the Antagonistic Effect of Soil Calcification on Bioavailability of Phosphorus in Alkaline Soils. Sci. Rep. 2017, 7, 16131. [Google Scholar] [CrossRef] [PubMed]
- Wang, F.; Deng, M.; Xu, J.; Zhu, X.; Mao, C. Molecular mechanisms of phosphate transport and signaling in higher plants. Semin. Cell Dev. Biol. 2018, 74, 114–122. [Google Scholar] [CrossRef]
- Yuan, J.; Zhang, Y.; Dong, J.; Sun, Y.; Lim, B.L.; Liu, D.; Lu, Z.J. Systematic characterization of novel lncRNAs responding to phosphate starvation in Arabidopsis thaliana. BMC Genom. 2016, 17, 655. [Google Scholar] [CrossRef] [Green Version]
- Jabnoune, M.; Secco, D.; Lecampion, C.; Robaglia, C.; Shu, Q.; Poirier, Y. A rice cis-natural antisense RNA acts as a translational enhancer for its cognate mRNA and contributes to phosphate homeostasis and plant fitness. Plant Cell 2013, 25, 4166–4182. [Google Scholar] [CrossRef] [Green Version]
- Huang, T.-K.; Han, C.-L.; Lin, S.-I.; Chen, Y.-J.; Tsai, Y.-C.; Chen, Y.-R.; Chen, J.-W.; Lin, W.-Y.; Chen, P.-M.; Liu, T.-Y.; et al. Identification of downstream components of ubiquitin-conjugating enzyme PHOSPHATE2 by quantitative membrane proteomics in Arabidopsis roots. Plant Cell 2013, 25, 4044–4060. [Google Scholar] [CrossRef] [Green Version]
- Wang, T.; Zhao, M.; Zhang, X.; Liu, M.; Yang, C.; Chen, Y.; Chen, R.; Wen, J.; Mysore, K.S.; Zhang, W.H. Novel phosphate deficiency-responsive long non-coding RNAs in the legume model plant Medicago truncatula. J. Exp. Bot. 2017, 68, 5937–5948. [Google Scholar] [CrossRef] [Green Version]
- Hawkesford, M.J.; Griffiths, S. Exploiting genetic variation in nitrogen use efficiency for cereal crop improvement. Curr. Opin. Plant Biol. 2019, 49, 35–42. [Google Scholar] [CrossRef]
- Chen, M.; Wang, C.; Bao, H.; Chen, H.; Wang, Y. Genome-wide identification and characterization of novel lncRNAs in Populus under nitrogen deficiency. Mol. Genet. Genom. 2016, 291, 1663–1680. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.; Jiang, Q.; Jiang, P.; Zhang, W.; Huang, J.; Liu, C.; Halford, N.G.; Lu, R. Novel low-nitrogen stress-responsive long non-coding RNAs (lncRNA) in barley landrace B968 (Liuzhutouzidamai) at seedling stage. BMC Plant Biol. 2020, 20, 142. [Google Scholar] [CrossRef] [PubMed]
- Fukuda, M.; Nishida, S.; Kakei, Y.; Shimada, Y.; Fujiwara, T. Genome-Wide Analysis of Long Intergenic Noncoding RNAs Responding to Low-Nutrient Conditions in Arabidopsis thaliana: Possible Involvement of Trans-Acting siRNA3 in Response to Low Nitrogen. Plant Cell Physiol. 2019, 60, 1961–1973. [Google Scholar] [CrossRef] [PubMed]
- Liu, F.; Xu, Y.; Chang, K.; Li, S.; Liu, Z.; Qi, S.; Jia, J.; Zhang, M.; Crawford, N.M.; Wang, Y. The long noncoding RNA T5120 regulates nitrate response and assimilation in Arabidopsis. New Phytol. 2019, 224, 117–131. [Google Scholar] [CrossRef]
- Unver, T.; Tombuloglu, H. Barley long non-coding RNAs (lncRNA) responsive to excess boron. Genomics 2020, 112, 1947–1955. [Google Scholar] [CrossRef]
- Zhou, G.F.; Zhang, L.P.; Li, B.X.; Sheng, O.; Wei, Q.J.; Yao, F.X.; Guan, G.; Liu, G.D. Genome-Wide Identification of Long Non-coding RNA in Trifoliate Orange (Poncirus trifoliata (L.) Raf) Leaves in Response to Boron Deficiency. Int. J. Mol. Sci. 2019, 20, 5419. [Google Scholar] [CrossRef] [Green Version]
- Jha, U.C.; Bohra, A.; Jha, R. Breeding approaches and genomics technologies to increase crop yield under low-temperature stress. Plant Cell Rep. 2017, 36, 1–35. [Google Scholar] [CrossRef]
- Kim, D.H.; Doyle, M.R.; Sung, S.; Amasino, R.M. Vernalization: Winter and the timing of flowering in plants. Annu. Rev. Cell Dev. Biol. 2009, 25, 277–299. [Google Scholar] [CrossRef] [Green Version]
- Castaings, L.; Bergonzi, S.; Albani, M.C.; Kemi, U.; Savolainen, O.; Coupland, G. Evolutionary conservation of cold-induced antisense RNAs of FLOWERING LOCUS C in Arabidopsis thaliana perennial relatives. Nat. Commun. 2014, 5, 4457. [Google Scholar] [CrossRef] [Green Version]
- Kindgren, P.; Ard, R.; Ivanov, M.; Marquardt, S. Transcriptional read-through of the long non-coding RNA SVALKA governs plant cold acclimation. Nat. Commun. 2018, 9, 4561. [Google Scholar] [CrossRef] [Green Version]
- Jiang, N.; Cui, J.; Shi, Y.; Yang, G.; Zhou, X.; Hou, X.; Meng, J.; Luan, Y. Tomato lncRNA23468 functions as a competing endogenous RNA to modulate NBS-LRR genes by decoying miR482b in the tomato-Phytophthora infestans interaction. Hortic. Res. 2019, 6, 28. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, J.; Yang, Y.; Jin, L.; Ling, X.; Liu, T.; Chen, T.; Ji, Y.; Yu, W.; Zhang, B. Re-analysis of long non-coding RNAs and prediction of circRNAs reveal their novel roles in susceptible tomato following TYLCV infection. BMC Plant Biol. 2018, 18, 104. [Google Scholar] [CrossRef] [PubMed]
- Xin, M.; Wang, Y.; Yao, Y.; Song, N.; Hu, Z.; Qin, D.; Xie, C.; Peng, H.; Ni, Z.; Sun, Q. Identification and characterization of wheat long non-protein coding RNAs responsive to powdery mildew infection and heat stress by using microarray analysis and SBS sequencing. BMC Plant Biol. 2011, 11, 61. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cui, J.; Luan, Y.; Jiang, N.; Bao, H.; Meng, J. Comparative transcriptome analysis between resistant and susceptible tomato allows the identification of lncRNA16397 conferring resistance to Phytophthora infestans by co-expressing glutaredoxin. Plant J. 2017, 89, 577–589. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.; Xia, R.; Kuang, H.; Meyers, B.C. The Diversification of Plant NBS-LRR Defense Genes Directs the Evolution of MicroRNAs That Target Them. Mol. Biol. Evol. 2016, 33, 2692–2705. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cui, J.; Jiang, N.; Hou, X.; Wu, S.; Zhang, Q.; Meng, J.; Luan, Y. Genome-Wide Identification of lncRNAs and Analysis of ceRNA Networks During Tomato Resistance to Phytophthora infestans. Phytopathology 2020, 110, 456–464. [Google Scholar] [CrossRef]
- Jiang, N.; Cui, J.; Hou, X.; Yang, G.; Xiao, Y.; Han, L.; Meng, J.; Luan, Y. Sl-lncRNA15492 interacts with Sl-miR482a and affects Solanum lycopersicum immunity against Phytophthora infestans. Plant J. 2020, 103, 1561–1574. [Google Scholar] [CrossRef]
- Zhang, G.; Chen, D.; Zhang, T.; Duan, A.; Zhang, J.; He, C. Transcriptomic and functional analyses unveil the role of long non-coding RNAs in anthocyanin biosynthesis during sea buckthorn fruit ripening. DNA Res. 2018, 25, 465–476. [Google Scholar] [CrossRef]
- Wunderlich, M.; Groß-Hardt, R.; Schöffl, F. Heat shock factor HSFB2a involved in gametophyte development of Arabidopsis thaliana and its expression is controlled by a heat-inducible long non-coding antisense RNA. Plant Mol. Biol. 2014, 85, 541–550. [Google Scholar] [CrossRef] [Green Version]
- Cho, J.; Koo, D.-H.; Nam, Y.-W.; Han, C.-T.; Lim, H.-T.; Bang, J.-W.; Hur, Y. Isolation and characterization of cDNA clones expressed under male sex expression conditions in a monoecious cucumber plant (Cucumis sativus L. cv. Winter Long). Euphytica 2005, 146, 271–281. [Google Scholar] [CrossRef]
- Yang, W.-C.; Katinakis, P.; Hendriks, P.; Smolders, A.; de Vries, F.; Spee, J.; van Kammen, A.; Bisseling, T.; Franssen, H. Characterization of GmENOD40, a gene showing novel patterns of cell-specific expression during soybean nodule development. Plant J. 1993, 3, 573–585. [Google Scholar] [CrossRef] [PubMed]
- Liao, X.; Wang, J.; Zhu, S.; Xie, Q.; Wang, L.; Yu, H.; Ye, Z.; Yang, C. Transcriptomic and functional analyses uncover the regulatory role of lncRNA000170 in tomato multicellular trichome formation. Plant J. 2020, 104, 18–29. [Google Scholar] [CrossRef] [PubMed]
- Li, R.; Fu, D.; Zhu, B.; Luo, Y.; Zhu, H. CRISPR/Cas9-mediated mutagenesis of lncRNA1459 alters tomato fruit ripening. Plant J. 2018, 94, 513–524. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Held, M.A.; Penning, B.; Brandt, A.S.; Kessans, S.A.; Yong, W.; Scofield, S.R.; Carpita, N.C. Small-interfering RNAs from natural antisense transcripts derived from a cellulose synthase gene modulate cell wall biosynthesis in barley. Proc. Natl. Acad. Sci. USA 2008, 105, 20534. [Google Scholar] [CrossRef] [Green Version]
- Zubko, E.; Meyer, P. A natural antisense transcript of the Petunia hybrida Sho gene suggests a role for an antisense mechanism in cytokinin regulation. Plant J. 2007, 52, 1131–1139. [Google Scholar] [CrossRef] [Green Version]
- Wang, H.V.; Chekanova, J.A. Long Noncoding RNAs in Plants. Adv. Exp. Med. Biol. 2017, 1008, 133–154. [Google Scholar] [CrossRef]
- Qin, T.; Zhao, H.; Cui, P.; Albesher, N.; Xiong, L. A Nucleus-Localized Long Non-Coding RNA Enhances Drought and Salt Stress Tolerance. Plant Physiol. 2017, 175, 1321–1336. [Google Scholar] [CrossRef] [Green Version]
- Burleigh, S.H.; Harrison, M.J. The down-regulation of Mt4-like genes by phosphate fertilization occurs systemically and involves phosphate translocation to the shoots. Plant Physiol. 1999, 119, 241–248. [Google Scholar] [CrossRef] [Green Version]
- Wasaki, J.; Yonetani, R.; Shinano, T.; Kai, M.; Osaki, M. Expression of the OsPI1 gene, cloned from rice roots using cDNA microarray, rapidly responds to phosphorus status. New Phytol. 2003, 158, 239–248. [Google Scholar] [CrossRef]
- Li, R.; Jin, J.; Xu, J.; Wang, L.; Li, J.; Lou, Y.; Baldwin, I.T. Long non-coding RNAs associate with jasmonate-mediated plant defense against herbivores. Plant Cell Environ. 2020. [Google Scholar] [CrossRef]
- Yu, Y.; Zhou, Y.-F.; Feng, Y.-Z.; He, H.; Lian, J.-P.; Yang, Y.-W.; Lei, M.-Q.; Zhang, Y.-C.; Chen, Y.-Q. Transcriptional landscape of pathogen-responsive lncRNAs in rice unveils the role of ALEX1 in jasmonate pathway and disease resistance. Plant Biotechnol. J. 2020, 18, 679–690. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, C.; Muchhal, U.S.; Raghothama, K.G. Differential expression of TPS11, a phosphate starvation-induced gene in tomato. Plant Mol. Biol. 1997, 33, 867–874. [Google Scholar] [CrossRef] [PubMed]
- Du, Q.; Wang, K.; Zou, C.; Xu, C.; Li, W.X. The PILNCR1-miR399 Regulatory Module Is Important for Low Phosphate Tolerance in Maize. Plant Physiol. 2018, 177, 1743–1753. [Google Scholar] [CrossRef] [PubMed] [Green Version]
Figure 1.
Timeline of the discovery of some of the important regulatory long non-coding RNAs (lncRNAs) in RNA biology from 1984 to 2019. The first lncRNA was the imprinted H19 gene, followed by the discovery of the XIST lncRNA gene during the pre-genomic era. In plants, lncRNA endo40 was first discovered to be involved in nodulation. Later, COOLAIR, COLDAIR, LDMR, PHO2, APOLO, LINC-AP2, MAS, and other lncRNA discoveries provided insights into their distinct biological roles in different processes such as vernalization, male sterility, sex-differentiation photomorphogenesis, and phosphate (Pi) uptake.
Figure 1.
Timeline of the discovery of some of the important regulatory long non-coding RNAs (lncRNAs) in RNA biology from 1984 to 2019. The first lncRNA was the imprinted H19 gene, followed by the discovery of the XIST lncRNA gene during the pre-genomic era. In plants, lncRNA endo40 was first discovered to be involved in nodulation. Later, COOLAIR, COLDAIR, LDMR, PHO2, APOLO, LINC-AP2, MAS, and other lncRNA discoveries provided insights into their distinct biological roles in different processes such as vernalization, male sterility, sex-differentiation photomorphogenesis, and phosphate (Pi) uptake.

Figure 2.
Biogenesis of lncRNAs and their classification based on genomic context. (A) Sense lncRNAs transcribed from protein-coding transcripts in the sense and antisense directions by alternative splicing. (B) Intronic lncRNAs synthesized from the intronic region, sometimes including exonic sequences. These lncRNAs are also transcribed in both the sense and antisense directions of their corresponding transcripts. (C) Those lncRNAs transcribed from intergenic regions of protein-coding transcripts are either in the sense or antisense direction between two protein-coding genes and are designated as intergenic lncRNAs. (D) Natural antisense lncRNAs arise antisense to the corresponding protein-coding gene. (E) Bi-directional lncRNAs, synthesized from the promoter of the protein coding-gene but in the opposite direction. (F) Enhancer lncRNAs arise from the enhancer region of the protein-coding gene, with or without the 3′-poly(A) tail modification. (G) LncRNAs transcribed from the promoter region of protein coding-genes, controlling the transcriptional regulation of related genes.
Figure 2.
Biogenesis of lncRNAs and their classification based on genomic context. (A) Sense lncRNAs transcribed from protein-coding transcripts in the sense and antisense directions by alternative splicing. (B) Intronic lncRNAs synthesized from the intronic region, sometimes including exonic sequences. These lncRNAs are also transcribed in both the sense and antisense directions of their corresponding transcripts. (C) Those lncRNAs transcribed from intergenic regions of protein-coding transcripts are either in the sense or antisense direction between two protein-coding genes and are designated as intergenic lncRNAs. (D) Natural antisense lncRNAs arise antisense to the corresponding protein-coding gene. (E) Bi-directional lncRNAs, synthesized from the promoter of the protein coding-gene but in the opposite direction. (F) Enhancer lncRNAs arise from the enhancer region of the protein-coding gene, with or without the 3′-poly(A) tail modification. (G) LncRNAs transcribed from the promoter region of protein coding-genes, controlling the transcriptional regulation of related genes.

Figure 3.
lncRNA classification based on their mechanisms of action—(A) as signal molecules to activate the transcription of the protein-coding transcript under a specific stimulus; (B) as decoys binding to miRNA or regulatory factors (RFs) and subsequently altering the corresponding function; (C) as guides, recruiting the chromatin-modifying complex that acts in cis or in trans of the target protein-coding transcript; (D) as scaffolds, recruiting multiple protein complexes and participating in chromatin modification to repress the target gene; (E) as substrates for miRNA-mediated phased secondary small interfering RNA (phasiRNA) generation, which functions in targeting several downstream genes.
Figure 3.
lncRNA classification based on their mechanisms of action—(A) as signal molecules to activate the transcription of the protein-coding transcript under a specific stimulus; (B) as decoys binding to miRNA or regulatory factors (RFs) and subsequently altering the corresponding function; (C) as guides, recruiting the chromatin-modifying complex that acts in cis or in trans of the target protein-coding transcript; (D) as scaffolds, recruiting multiple protein complexes and participating in chromatin modification to repress the target gene; (E) as substrates for miRNA-mediated phased secondary small interfering RNA (phasiRNA) generation, which functions in targeting several downstream genes.

Figure 4.
Representative lncRNAs involved in various developmental processes. (A) Regulation of gene expression by COOLAIR and COLDAIR lncRNAs during vernalization. Under high-temperature conditions, the R-loop in the COOLAIR promoter is activated by NDX proteins, and the COOLAIR transcript binds to the single-stranded DNA, promoting FLC locus expression. Upon exposure to cold conditions, COLDAIR binds to PRC2, whereas COOLAIR induces methylation in the FLC gene promoter to suppress its transcription. (B) The function of ENOD40 lncRNA in post-transcription of nodulation. (C) Regulation of lncRNA APOLO produced from the PID locus and its subsequent response to auxin polar transportation. In response to auxin, DNA demethylation of APOLO lncRNA is activated through chromatin loop activation, and RNA Pol II-transcribed APOLO recruits LHP1, thus increasing APOLO and PID expression. In contrast, Pol IV/V activates the PID promoter’s DNA methylation by recruiting AGO4 and PRC1 on the APOLO gene to repress PID synthesis.
Figure 4.
Representative lncRNAs involved in various developmental processes. (A) Regulation of gene expression by COOLAIR and COLDAIR lncRNAs during vernalization. Under high-temperature conditions, the R-loop in the COOLAIR promoter is activated by NDX proteins, and the COOLAIR transcript binds to the single-stranded DNA, promoting FLC locus expression. Upon exposure to cold conditions, COLDAIR binds to PRC2, whereas COOLAIR induces methylation in the FLC gene promoter to suppress its transcription. (B) The function of ENOD40 lncRNA in post-transcription of nodulation. (C) Regulation of lncRNA APOLO produced from the PID locus and its subsequent response to auxin polar transportation. In response to auxin, DNA demethylation of APOLO lncRNA is activated through chromatin loop activation, and RNA Pol II-transcribed APOLO recruits LHP1, thus increasing APOLO and PID expression. In contrast, Pol IV/V activates the PID promoter’s DNA methylation by recruiting AGO4 and PRC1 on the APOLO gene to repress PID synthesis.

Figure 5.
LncRNAs controlling abiotic and biotic stress responses in plants. (A) In drought conditions, lncRNA lincRNA340 is induced to repress miR169, which relieves nuclear factor Y (NF-Y) gene expression to improve stress tolerance. (B) lncRNA973 acts as a positive regulator of salt-responsive genes in ROS (reactive oxygen species) to enhance salinity tolerance. (C) IPS1 decoys miR399 to regulate the expression of PHO2 and help maintain phosphate homeostasis in the root. (D) lncRNA T5120 functions in NLP7-mediated nitrate regulation in roots. (E) The aggressive interaction between Sl-miR482a and Sl-lncRNA15492 in tomatoes helps to maintain Sl-NBS-LRR homeostasis, thereby affecting the resistance of tomatoes to P. infestans.
Figure 5.
LncRNAs controlling abiotic and biotic stress responses in plants. (A) In drought conditions, lncRNA lincRNA340 is induced to repress miR169, which relieves nuclear factor Y (NF-Y) gene expression to improve stress tolerance. (B) lncRNA973 acts as a positive regulator of salt-responsive genes in ROS (reactive oxygen species) to enhance salinity tolerance. (C) IPS1 decoys miR399 to regulate the expression of PHO2 and help maintain phosphate homeostasis in the root. (D) lncRNA T5120 functions in NLP7-mediated nitrate regulation in roots. (E) The aggressive interaction between Sl-miR482a and Sl-lncRNA15492 in tomatoes helps to maintain Sl-NBS-LRR homeostasis, thereby affecting the resistance of tomatoes to P. infestans.


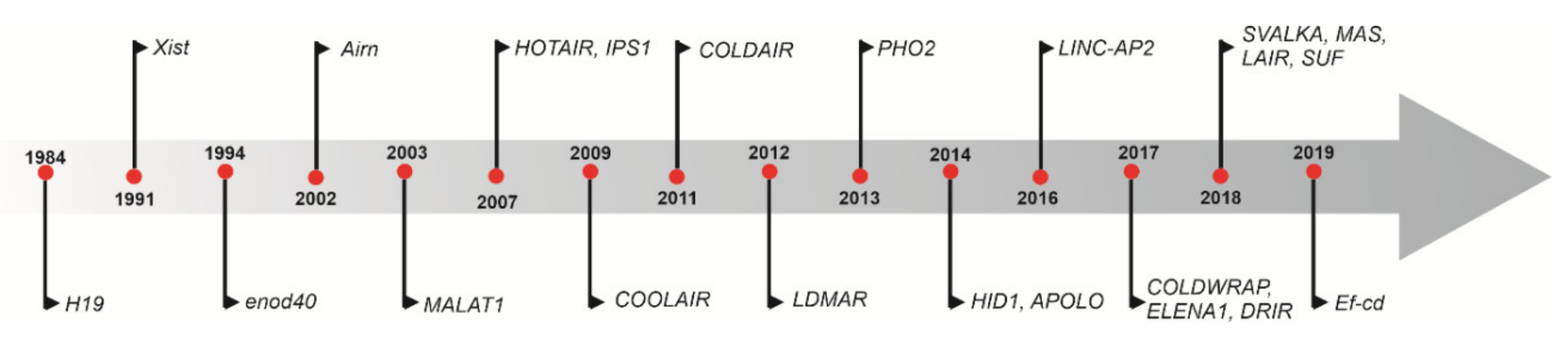{kind=link}
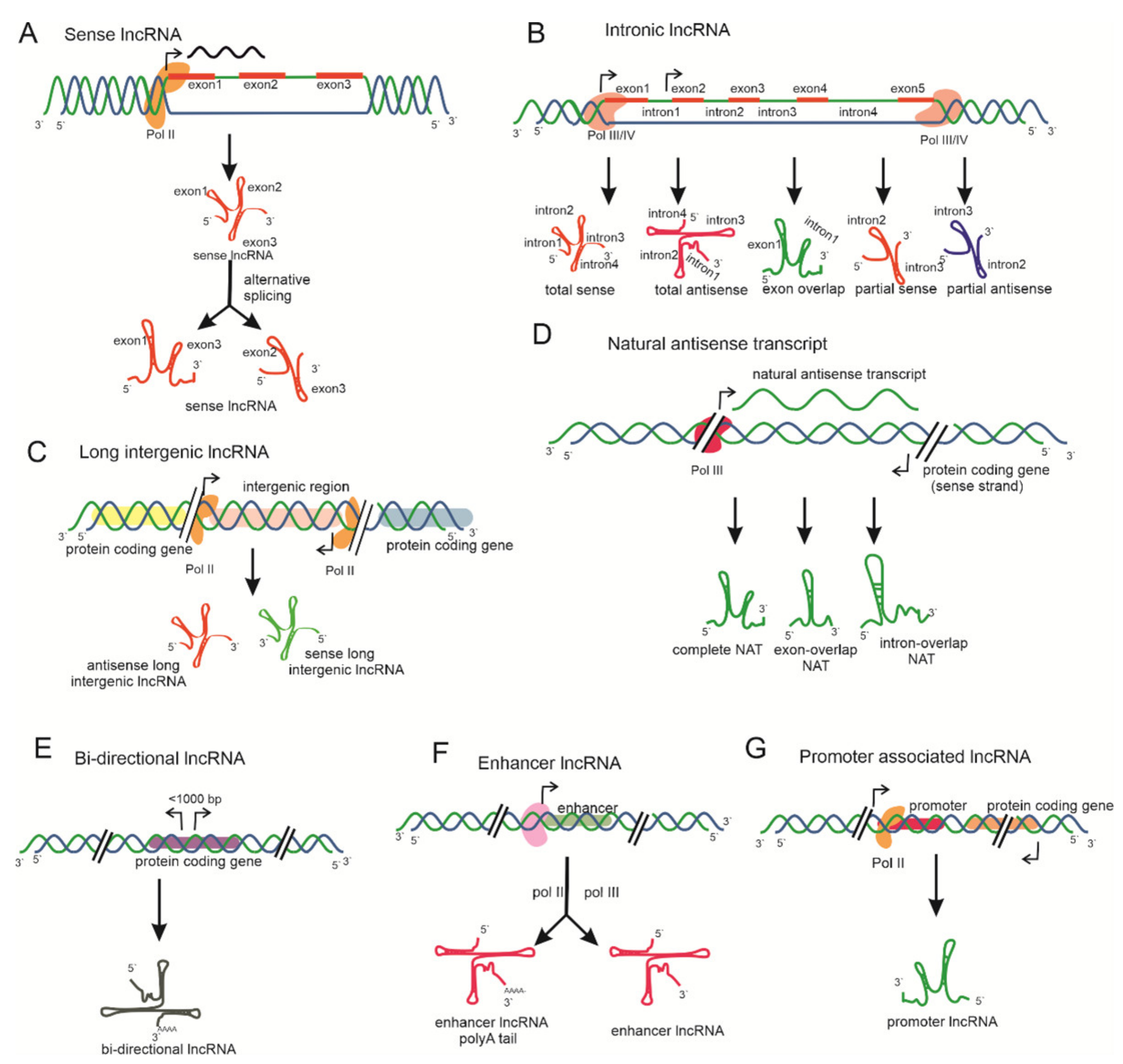{kind=link}
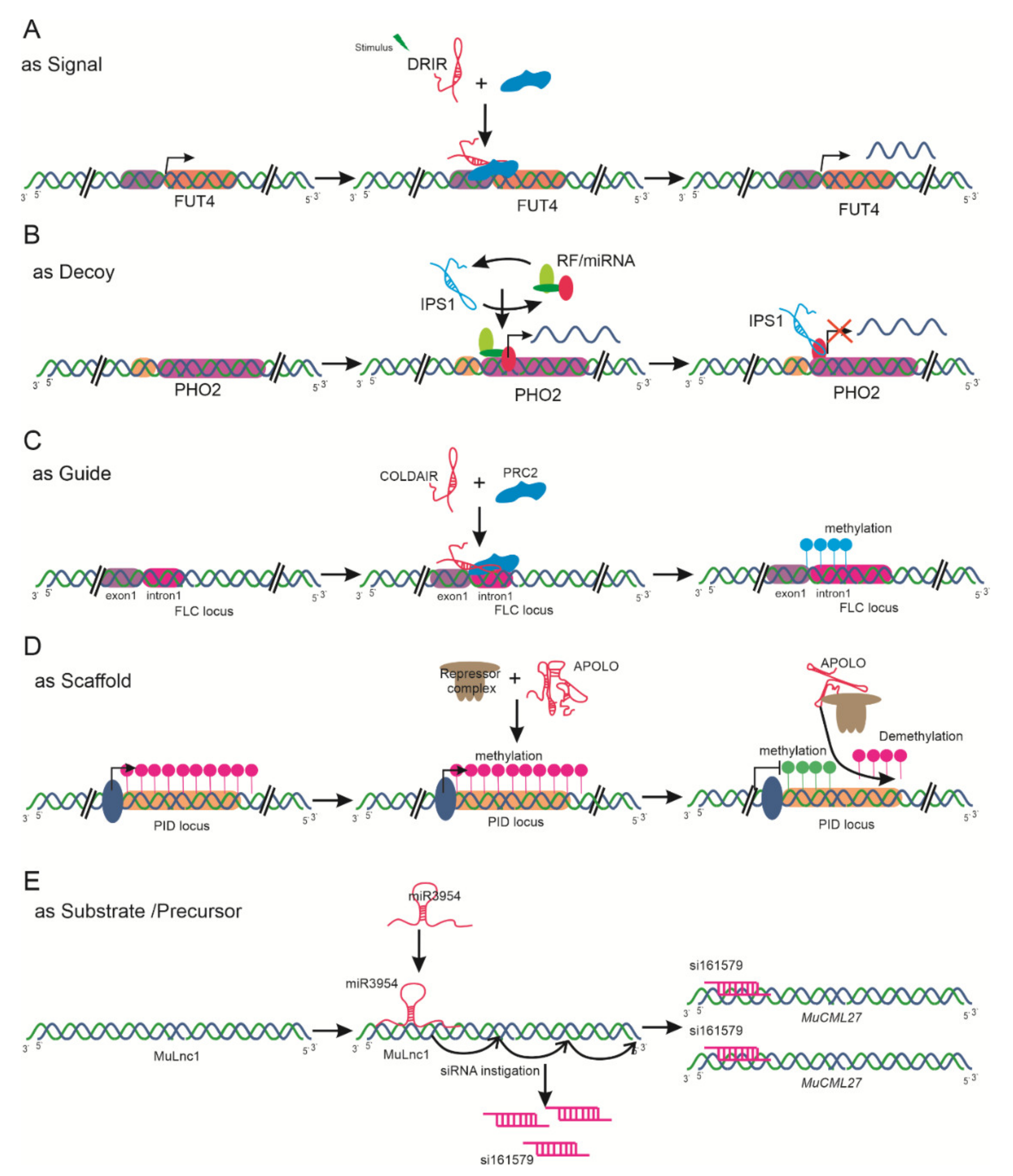{kind=link}
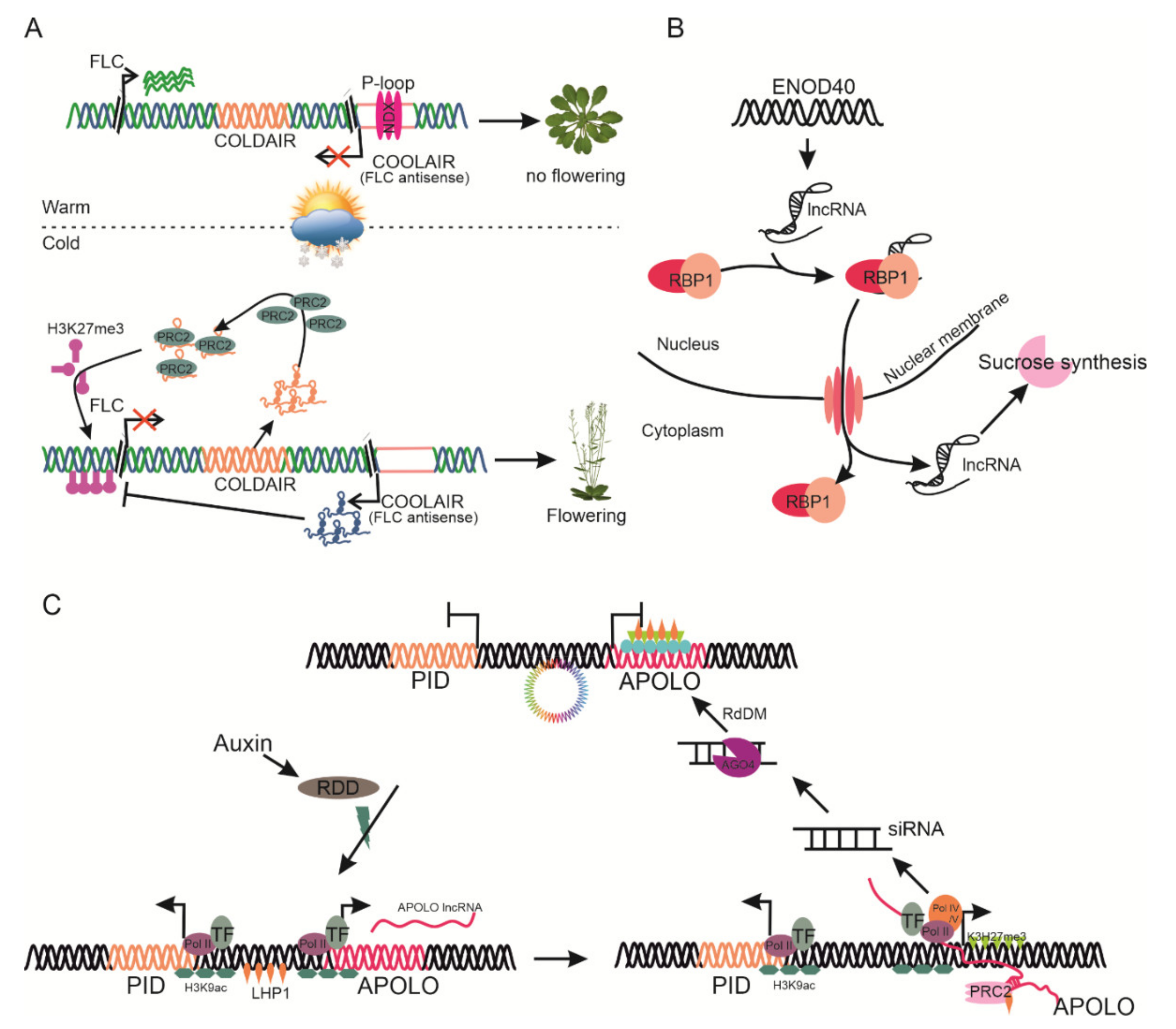{kind=link}
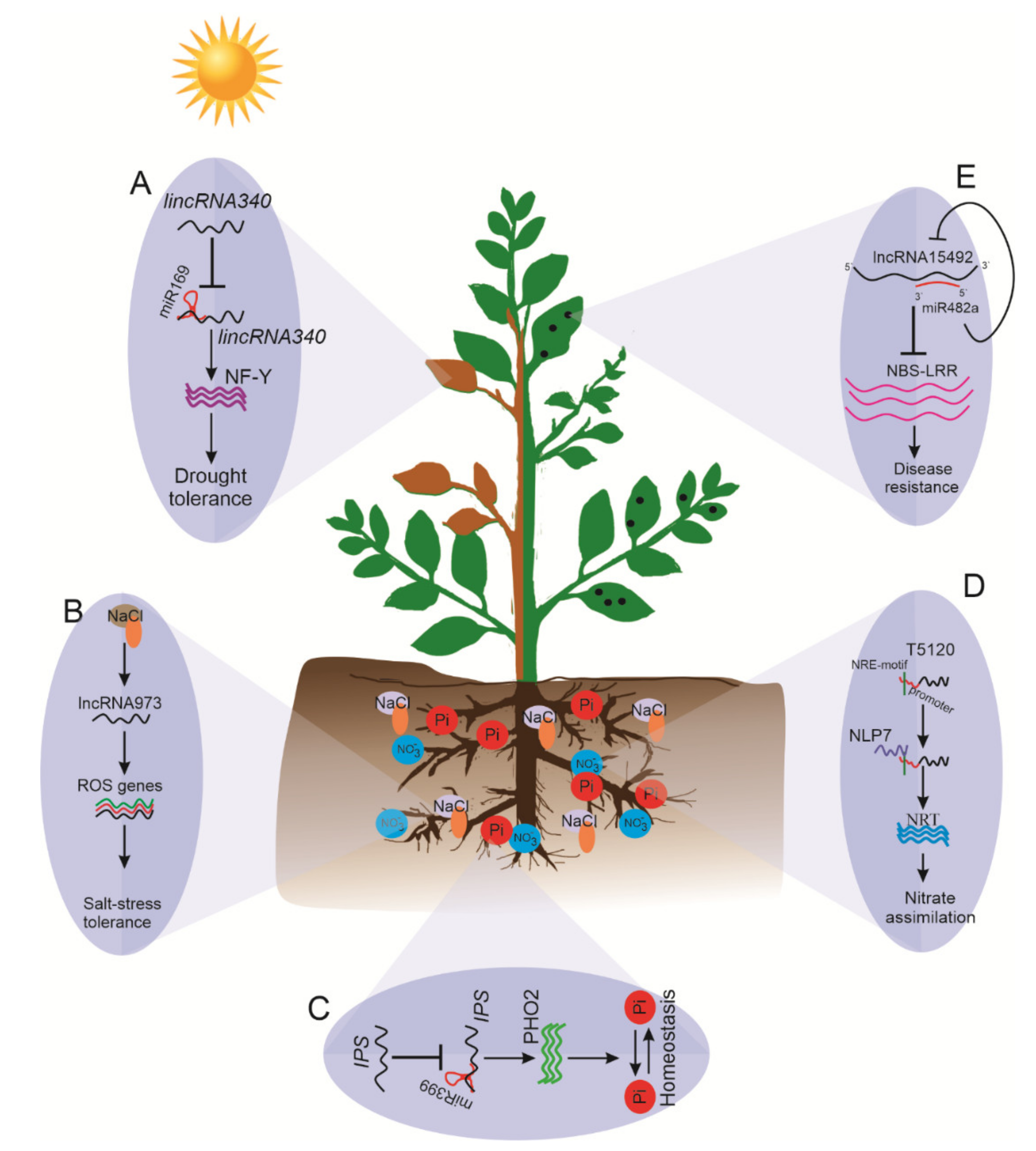{kind=link}
Table 1.
List of genome-scale identifications of plant-specific lncRNAs in various species and under different abiotic and biotic conditions. EST: expressed sequence tag.
Table 1.
List of genome-scale identifications of plant-specific lncRNAs in various species and under different abiotic and biotic conditions. EST: expressed sequence tag.
| Name of Plants | No. of lncRNAs | Identification Approach | Stress |
|---|---|---|---|
| Actinidia chinenesis | 7051 | Transcriptome data | |
| Arabidopsis | 1212 | Transcriptome data | Phosphate starvation, drought stress, salt stress Heat, cold, salt, |
| 76 | Genome data | ||
| B. napus | 3181 | Transcriptome data | Cold and heat |
| 10,001 | Transcriptome data | Sclerotinia stem rot by Sclerotinia sclerotiorum | |
| C. arietinum | 2248 | Transcriptome data | |
| C.s sativus | 3274 | Transcriptome data | |
| D. purpurea | 2660 | Transcriptome data | Cold, dehydration |
| F. vesca | 5884 | Transcriptome data | |
| G. arboretum | 5996 | Transcriptome data | |
| G. hirsutum | 1117 | Transcriptome data | Salt |
| H. annus | 25,327 | Transcriptome data | |
| M. truncatula | 503 | EST and Genome data | Osmotic and salt |
| 23,324 | Transcriptome data | ||
| M. notabilis | 1133 | Transcriptome data | |
| M. acuminata | 905 | Transcriptome data | Drought, Fusarium wilt disease |
| 5294 | Transcriptome data | ||
| O sativa | 2224 | Transcriptome data | |
| P. ginseng | 3688 | Transcriptome data | |
| P. trichocarpa | 2542 | Transcriptome data | Drought |
| Populus sp. | 7655 | Transcriptome data | |
| P. tomentosa | 1377 | Transcriptome data | |
| P. persica | 1417 | Deep Sequencing | |
| S. miltiorrhiza | 5446 | Transcriptome data | Methyl-jasmonate |
| S. italica | 584 | Transcriptome data | Drought |
| S. lycopersicum | 3679 | Transcriptome data | |
| 1565 | Transcriptome data | ||
| S. tuberosum | 1113 | Transcriptome data | |
| T. aestivum | 58,218 | Transcriptome data | Heat stress, powdery mildew |
| 125 | Transcriptome data | ||
| Z. mays | 1223 | Transcriptome data | Drought stress |
| 20,163 | Genome and transcriptome data | ||
| 664 | Genome data | ||
| 7245 | Transcriptome data |
Table 2.
A summarized list of bioinformatics tools, resources, and archived databases predicting lncRNAs in plants.
Table 2.
A summarized list of bioinformatics tools, resources, and archived databases predicting lncRNAs in plants.
| Name | Description | Weblink | |
|---|---|---|---|
| Prediction tools | PhlyoCSF [53] | lncRNA coding potential calculator using codon substitution frequency (CSF) scores | https://github.com/mlin/PhyloCSF |
| CPC [52] | lncRNA coding potential using sequence features and support vector machine (SVM) | http://cpc.cbi.pku.edu.cn/ | |
| CNCI [61] | Calculation of lncRNA coding potential by profiling adjoining nucleotide triplets | https://github.com/www-bioinfo-org/CNCI | |
| CPAT [54] | Calculation of lncRNA coding potential using a logistic regression model | http://rna-cpat.sourceforge.net/ | |
| DeepLNC [56] | lncRNA prediction using deep neural network | http://bioserver.iiita.ac.in/deeplnc/ | |
| iSeeRNA [55] | lncRNA prediction using SVM algorithm | http://137.189.133.71/iSeeRNA/ | |
| lncRNATargets [57] | lncRNA target prediction based on nucleic acid thermodynamics | http://www.herbbol.org:8001/lrt/index.php | |
| spongeScan [58] | detection of miRNA elements in sponge lncRNAs | http://spongescan.rc.ufl.edu/ | |
| TF2LncRNA [60] | Identification of transcription factors for lncRNAs | http://mlg.hit.edu.cn/tf2lncrna | |
| RegRNA [59] | For identifying regulatory RNA motifs | http://regrna2.mbc.nctu.edu.tw/index.html | |
| Database | NONCODE [62] | Complete collection and annotation of ncRNAs from 16 species, including Arabidopsis as the only plant species; 3853 lncRNA transcripts and 2477 lncRNA genes | http://www.noncode.org/ |
| GREENC [63] | A wiki-based plant lncRNA database from 37 plant species; 120000 lncRNAs | http://greenc.sciencedesigners.com/ | |
| lncRNAdb [64] | Literature describing functions of lncRNAs from Arabidopsis, rice, and others | http://lncrnadb.org | |
| PLncDB [32] | Arabidopsis lncRNAs; >13000 lncRNAs | http://chualab.rockefeller.edu/gbrowse2/homepage.html | |
| PNRD [65] | lncRNAs from Arabidopsis, poplar, maize, and rice; 5573 lncRNAs | http://structuralbiology.cau.edu.cn/PNRD/ | |
| CANTATAdb [66] | lncRNAs from 10 model plant species; 45117 lncRNAs | http://yeti.amu.edu.pl/CANTATA/ | |
| PLNlncRbase [67] | Experimentally verified plant lncRNAs in 43 species: 1187 lncRNAs | http://bioinformatics.ahau.edu.cn/PLNlncRbase/pcsb | |
| PLncRNAdb [68] | Plant-specific lncRNAs with a distinct annotation like information lncRNAs and various RNA-binding proteins (RBPs); 5000 lncRNAs | http://bis.zju.edu.cn/PlncRNADB/index.php | |
| EVLncRNAs [69] | Experimentally validated lncRNAs in 77 plants and animals; 1543 lncRNAs | http://biophy.dzu.edu.cn/EVLncRNAs |
Table 3.
Representative studies showing lncRNA-mediated growth, development, and stress tolerance in plants over the last decade.
Table 3.
Representative studies showing lncRNA-mediated growth, development, and stress tolerance in plants over the last decade.
| Role | Species | lncRNA | Function | Reference |
|---|---|---|---|---|
| Development | M. truncatula | ENOD40 | Nodule development | [14] |
| Arabidopsis | COOLAIR | Flowering | [21] | |
| B. campestris | MF11 | Male sterility | [22] | |
| B. campestris | ZM40 | Male sterility | [23] | |
| Arabidopsis | APOLO | Polar auxin transport | [26] | |
| Arabidopsis | NATs | Photomorphogenesis | [33] | |
| Arabidopsis | COLDAIR | Flowering | [46] | |
| O. sativa | LDMAR | Male sterility | [47] | |
| Arabidopsis | HIDI | Photomorphogenesis | [69] | |
| Arabidopsis | COLDWRAP | Flowering | [80] | |
| Arabidopsis | ASCO | Regulation of auxin | [82] | |
| Arabidopsis | MAS | Flowering | [95] | |
| Arabidopsis | CDF5 | Circadian | [96] | |
| Arabidopsis | AtR8 | Hypoxic stress | [96] | |
| Arabidopsis | FLORE | Flowering | [96] | |
| Arabidopsis | LINC-AP2 | Flowering | [98] | |
| M. polymorpha | SUF | Sexual differentiation | [100] | |
| O. sativa | MIKKI | Root Growth | [107] | |
| S. buckthorn | LNC1/2 | Anthocyanin biosynthesis | [150] | |
| Arabidopsis | asHSFB2a | Vegetative and gametophytic development | [151] | |
| C. sativus | CsM10 | Sex differentiation | [152] | |
| G. max | GmENOD40 | Nodule development | [153] | |
| S. lycopersicum | lncRNA000170 | Trichome formation | [154] | |
| S. lycopersicum | lncRNA1459 | Fruit ripening | [155] | |
| H. vulgare | HvCesA6 | Cell wall synthesis | [156] | |
| P. hybrid | SHO | Cytokinin biosynthesis | [157] | |
| Arabidopsis | ASL | Flowering | [158] | |
| Biotic and abiotic stress | Arabidopsis | ELENA1 | Pathogen infection | [24] |
| Arabidopsis | IPS1 | Phosphate starvation | [45] | |
| Arabidopsis | T5120 | Nitrate assimilation | [136] | |
| Arabidopsis | SVALKA | Cold-induced | [142] | |
| S. lycoperscium | lncRNA23468, lncRNA16397 | P. infestans infection | [143,146] | |
| S. lycoperscium | S-slylnc0957 | Pathogen infection | [144] | |
| Arabidopsis | DRIR | Drought tolerance | [159] | |
| M. truncatula | Mt4 | Phosphate starvation | [160] | |
| O. sativa | OsPI1 | Phosphate starvation | [161] | |
| O. sativa | ALEX1 | JA-mediated disease resistance | [162] | |
| S. lycoperscium | TPS11 | Phosphate starvation | [163] | |
| Z. mays | PILNCR1 | Phosphate starvation | [164] | |
| N. attenuata | JAL1/3 | JA-mediated herbivore resistance | [165] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Waseem, M.; Liu, Y.; Xia, R. Long Non-Coding RNAs, the Dark Matter: An Emerging Regulatory Component in Plants. Int. J. Mol. Sci. 2021, 22, 86. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22010086
AMA Style
Waseem M, Liu Y, Xia R. Long Non-Coding RNAs, the Dark Matter: An Emerging Regulatory Component in Plants. International Journal of Molecular Sciences. 2021; 22(1):86. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22010086
Chicago/Turabian StyleWaseem, Muhammad, Yuanlong Liu, and Rui Xia. 2021. "Long Non-Coding RNAs, the Dark Matter: An Emerging Regulatory Component in Plants" International Journal of Molecular Sciences 22, no. 1: 86. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22010086
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.