GPER1 and microRNA: Two Players in Breast Cancer Progression


Department of Pharmacy, Health and Nutrition Sciences, University of Calabria, 87036 Rende, Italy
Int. J. Mol. Sci. 2021, 22(1), 98; https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22010098
Submission received: 2 December 2020
/
Revised: 21 December 2020
/
Accepted: 22 December 2020
/
Published: 24 December 2020
(This article belongs to the Special Issue New Regulators and Modulators of MicroRNA 2022)
Abstract
:Breast cancer is the main cause of morbidity and mortality in women worldwide. However, the molecular pathogenesis of breast cancer remains poorly defined due to its heterogeneity. Several studies have reported that G Protein-Coupled Estrogen Receptor 1 (GPER1) plays a crucial role in breast cancer progression, by binding to estrogens or synthetic agonists, like G-1, thus modulating genes involved in diverse biological events, such as cell proliferation, migration, apoptosis, and metastasis. In addition, it has been established that the dysregulation of short sequences of non-coding RNA, named microRNAs (miRNAs), is involved in various pathophysiological conditions, including breast cancer. Recent evidence has indicated that estrogens may regulate miRNA expression and therefore modulate the levels of their target genes, not only through the classical estrogen receptors (ERs), but also activating GPER1 signalling, hence suggesting an alternative molecular pathway involved in breast tumor progression. Here, the current knowledge about GPER1 and miRNA action in breast cancer is recapitulated, reporting recent evidence on the liaison of these two players in triggering breast tumorogenic effects. Elucidating the role of GPER1 and miRNAs in breast cancer might provide new tools for innovative approaches in anti-cancer therapy.
1. Introduction
Breast cancer is the most diagnosed cancer among women, with about two million cases each year and a mortality rate of 6.6% of all cancer deaths [1]. Breast cancer mortality in western countries has progressively decreased over the past 25 years and more than 90% of patients heal when the treatment starts at the early stages of the disease [1,2]. Therapy management of breast cancer depend on its histological and molecular characteristics, based primarily on the expression of estrogen receptors (ERs), progesterone receptor (PR) and ERBB2 receptor (HER2), which allow to distinct the breast carcinoma in diverse molecular subtypes (i.e., Luminal A and B, Her2-enriched, Basal- and Normal-like, ER-negative and Triple-Negative Breast Cancers, TNBC) [3]. Of note, four distinct subtypes of ER-negative breast cancer and six TNBC subtypes have been identified [3]. Based on this evidence, correct therapeutic approaches in breast cancer patients have become increasingly important to meliorate the response to conventional treatments. Despite advances in breast cancer therapeutic and diagnostic approaches, the management of metastasis derived from disseminated primary tumor cells remains an issue to address. [2,4]. It has been estimates that about 30% of women affected by breast cancer will develop metastasis in their lifetime [1]. Moreover, over 90% of the deaths of breast cancer patients are caused by metastasis [1]. Although the incidence of breast cancer rises with age, the rate of increase is slower after menopause, when estrogen production is reduced, thus suggesting the hormone-dependence of this neoplasia [4]. In this regard, it has been well established that the cumulative exposure of the breast tissue to estrogens may affect the cell division rate and cause increased cell proliferation, hence acting as an important risk factor in breast cancer development [4]. Estrogens, in particular 17β-estradiol (E2), exert important biological effects in a wide variety of normal and malignant tissues, through their nuclear receptors, ERα and ERβ [5]. Nevertheless, E2 also induce non-genomic effects binding to G protein coupled estrogen receptor 1 (GPER1) and activating rapid transduction pathways involved in neoplastic transformation and development [6]. In addition, it has been showed that E2 may regulate the expression of short sequences of non-coding RNA, named microRNAs (miRNAs) [7,8,9], whose dysregulation in diverse patho-physiological conditions, including breast cancer, it has been recently demonstrated [10,11].
Here, GPER1 and miRNAs involvement in breast cancer progression is reviewed, underlining the more recent action of GPER1 in modulating miRNA expression.
2. GPER1 History, Expression and Localization
Briefly, the history of GPER1 starts in the 1997 when a 7-transmembrane receptor belonging to the large family of membrane G-protein coupled receptor (GPCR), named GPR30, was identified and cloned [12]. Although GPR30 showed a similar structure to the interleukin 8 receptor and the angiotensin II receptor type 1 [13,14], the treatment with chemokines or peptides did not lead to the activation of this receptor [14], suggesting thus that GPR30 could be an orphan. Some years later, studies performed in breast cancer cells lacking ERs, but expressing GPR30, evidenced the ability of E2 to induce rapidly cell cascades and trigger growth effects through GPR30 [15,16]. Interestingly, these responses were blocked, silencing GPR30 expression [16]. Next, experimental data indicated a direct bind to GPR30 by E2, suggesting that it may work as a membrane-bound ER [17,18]. These findings, together with in vivo data showing that hepatic injury decreased by E2/GPER1/protein kinase A (PKA) pathway activation [19], allowed to name in 2007 GPR30 officially as the novel estrogen receptor GPER1 [20]. To date, in November 2020 “GPR30 or GPER1 or GPER and breast cancer” keywords in PubMed database yielded 1361 papers, whose about 89% published during the past decade [https://pubmed.ncbi.nlm.nih.gov/].
GPER1 gene, located on the chromosome 7p22.3, encodes a protein of a putative molecular mass of 41 kDa and high homology with its murine counterpart [21]. Similar to other GPCR, the N-terminal region of GPER1 is outside the cell, whereas the C-terminal region is located intracellularly [21].
Regarding GPER1 expression, its protein is not only expressed in normal and tumoral estrogen-responsive tissues [12,16,22,23], but also in several other tissues, such as brain, lung, liver, adipose tissue, vasculature, as well as in immune cells [24,25,26]. Interestingly, the expression of GPER1 is variable and may depend on the specie and tissue, as well as on the age and gender [27,28]. For instance, in the mammary gland, GPER1 expression is lower during puberty and then increases in sexual maturity stages [29]. In a number of cancer types, like breast, endometrial and ovarian tumors, high levels of GPER1 have been associated with larger tumor size, Her-2 expression and metastasis, as well as with poor survival [30,31,32,33]. However, it has also been reported that in breast cancer low levels of GPER are associated with metastatic lymph nodes, absence of ER expression, poor patient disease-free survival, and overall survival [34].
The significance of GPER1 expression related to ER-positive breast cancer and in TNBC has been investigated. GPER1 has been detected in about 50 to 60% of breast cancer tissues [35]. Among these, GPER1 expression has been evidenced in the majority of TNBC, whereas co-expression of GPER1 and ER was about 40% of all cases examined [35]. Notably, in patients treated with tamoxifen the expression of GPER1 was found increased and associated with a poor prognosis and relapse-free survival, suggesting that GPER1 expression in ER-positive breast cancer is correlated to tamoxifen resistance [36].
In TNBC, GPER1 expression is related with augmented recurrence and associated with a younger age and a more aggressive disease. After a 36-month follow-up, it has been reported that the rate survival in patients affected by TNBC and expressing low levels of GPER1 was 90%, whereas in the cohort with high GPER1 levels only 78% of the patients survived after this time period [34].
Moreover, GPER-1 in TNBC has been associated with a significant shorter overall survival and relapse-free survival of premenopausal patients [37].
GPER1 localization has been controversially discussed for many years. Similar to most trans-membrane GPCRs, GPER1 is integrated into the membrane and its ligand-activation lead to cAMP synthesis, a typical plasma membrane event [17,38,39,40]. Several studies confirm that GPER1 is mainly placed to the plasma membrane of diverse cells, like mammary and uterine epithelial cells [41,42], myometrium cells [43], renal epithelia cells [44,45,46], and hippocampal neurons [39,47]. However, the localization of GPER1 also seems to depend on species, tissue, and cell types. In this context, it has been reported an intracellular localization of GPER1, in particular bound to endoplasmic reticulum and co-localized in the Golgi apparatus and nuclear membrane in endothelial [48], vascular smooth muscle cells [48] and pancreatic islet cells [49,50] as well as in diverse cancer cell types [18]. Of note, an intranuclear localization of GPER1 it has also been shown in the more representative components of tumor stroma, especially of breast cancer, named cancer associated fibroblasts (CAFs) [51,52]. These data suggest that the cellular localization of GPER1 may be varied by specific environmental signalling.
3. GPER1 Ligands and Signalling
Although GPER is a very ubiquitous receptor in the cell, the endogenous and canonical ligand of GPER has not yet been found. Therefore, diverse computational and experimental studies have been performed in order to identify natural or synthetic ligands of GPER1 ([53,54] and reviewed therein). Apart from E2 that binds to GPER1 with an estimated binding affinity of 3–6 nM [17,18], a great number of other compounds have been identified, like genistein, quercetin, bisphenol A and many pesticides [53,55,56,57]. Interestingly, data have indicated that the modulators or antagonists of the classical ERs, such as tamoxifen, raloxifene, and fulvestrant, may act as GPER1 agonists [17,58]. In this context, discovery that tamoxifen is a GPER1-agonist led to the conclusion that GPER1 may contribute to the tamoxifen resistance in breast malignancy. Indeed, studies have indicated an increased expression of GPER1 in breast tumors with an acquired tamoxifen resistance [36].
Identification and synthesis in 2006 of the highly selective GPER1 agonist, G-1 [59] has allowed to analyse its specific signalling pathways and to obtained selective information on the GPER1 action in diverse cell contexts. Indeed, by experiments of binding, it was evidenced that G-1 has high affinity for GPER1 (Kd = 10 nM), but it binds to ERs at concentrations of 10µM [60]. The next year, two novel GPER1 specific agonists, named GPER1-L1 and GPER1-L2, with binding affinities of about 100 nM were synthesized [61]. Moreover, it was reported that propyl pyrazole triol (PPT), usually used as ERα specific agonist, may act as GPER1 agonist at concentrations as low as 10–100 nM, while the ERβ specific agonist diarylpropionitrile (DPN) had no effects on GPER1 activity [62].
As concern the antagonist compounds of GPER1, by screening a small molecules library (NIH-MLSMR), it was found G-15 as a selective GPER1 antagonist. G-15 is a synthetic substituted dihydroquinoline with similar structure as G-1, but lacking of the ethanone moiety [60]. G-15 shows a GPER binding affinity of 20 nM and only a minimal binding to ERs (Kd > 10 µM) [60]. Restoring the steric bulk of G-1, another GPER1 specific antagonist, named G-36, with an improved affinity to GPER1, was next developed [60]. Lappano et coll. reported that estriol, which is one of three endogenous estrogens (the other two are estradiol and estrone [63]), is an inhibitor of GPER1 activity [64]. As few years later, further experimental data showed that in breast cancer cells the synthetic molecule MIBE may bind to and block both ERα and GPER1 activity [61]. More recently, it has been identified the first peptide GPER1 ligand, termed ERalpha17p, corresponds to a portion of the hinge region/AF2 domain of the human ERα [65,66]. ERalpha17p shows an anti-tumoral activity in breast cancer cells [65,66].
A number of studies has indicated that, in estrogen-sensitive tumors, the activation of GPER1 induces several cascade responses leading to important biological events, like cell growth, migration and angiogenesis [62,67,68,69,70]. The network of signal transduction pathways mediated by GPER1 includes transactivation of the epidermal growth factor receptor (EGFR), activation of the mitogen activated protein kinase/extracellular regulated kinase (MAPK/ERK) and phosphatidylinositol 3-kinase (PI3K)/Akt cascades, calcium mobilization, and intracellular cyclic AMP production. In particular, in human breast cancer cells, the activation of MAPK/ERK signalling by E2-GPER1 binding involves the heterotrimeric G-proteins βγ-subunits and the cytosolic kinase src activation, suggesting the contribution of the heparin-bound EGF cleavage [15]. In particular, estrogenic GPER1 stimulation leads to the release of intracellular calcium from endoplasmic reticulum stores and the consequent activation of integrin α5β1, which in turn induces matrix metalloproteinase-dependent activation of the EGFR by release of membrane heparin-bound EGF [23]. In this manner, estrogenic action via GPER1 coordinates the release of local EGF-related polypeptides and EGFR phosphorylation, which, activating STAT5 and MAPK/ERK pathways, induces cellular activities associated with cell survival [23]. Besides ERK activation, the transactivation of the EGFR by E2 also activates PI3K/Akt pathway [70]. It has also been reported that, in breast cancer cells, the estrogenic stimulation of GPER1 causes activation of the Gαs protein which, in turn, leads to adenylyl cyclase activation and cAMP accumulation [17,21].
Growing evidence has shown that activation of GPER1 in several cancer cells, as well as in CAFs, may induce the expression of diverse genes, like c-fos, cyclin D1 and CTGF, involved in important biological effects, including cell proliferation and migration [71]. Moreover, GPER1 signalling triggers HIF-1α-dependent VEGF expression supporting its involvement in angiogenesis and progression of breast cancer [72]. Interestingly, estrogenic GPER1 activation is able to regulate the expression of the pro-inflammatory cytokine IL1β and its receptor IL1R1 in CAFs and breast cancer cells, respectively, suggesting a fine feedforward, which links tumor microenvironment with tumor cells toward the progression of breast cancer [73]. In this context, increasing data have indicated that the depletion of CAFs in tumor stroma led to decreased cancer growth and improved response to therapies [74]. These data suggest that tumor microenvironment is a fertile ground for innovative therapeutic approaches with the potential to ameliorate existing treatment and prevention options.
4. miRNA History and Biosynthesis
miRNAs were initially discovered in Caenorhabditis elegans as small temporal RNA (stRNA) [75] and characterized in subsequent studies [76,77,78]. miRNAs are a large family of small non-coding RNAs (about 22 nucleotides) presents in all species ranging from plants to humans [79]. miRNAs have regulatory and catalytic functions at the post-transcriptional level [80], by binding to the 3′- and, rarely, 5′-untranslated regions of one or more functionally related target mRNAs, thus repressing protein translation or initiating mRNA destabilization/degradation [81]. Considering that more than 60% of all mRNAs could be possible target of miRNAs [82], dysregulation of their expression may lead to important alterations in cellular processes such as cell proliferation, apoptosis, differentiation, and embryonic development [83]. Therefore, dysregulation of miRNAs expression may be associated with several pathologies including cardiovascular and blood diseases [84,85], diabetes [86] and cancer [83]. In this regard, it is important to mention that, depending on the cell context, the same miRNA may work as a tumor suppressor or show oncogenic activities [87].
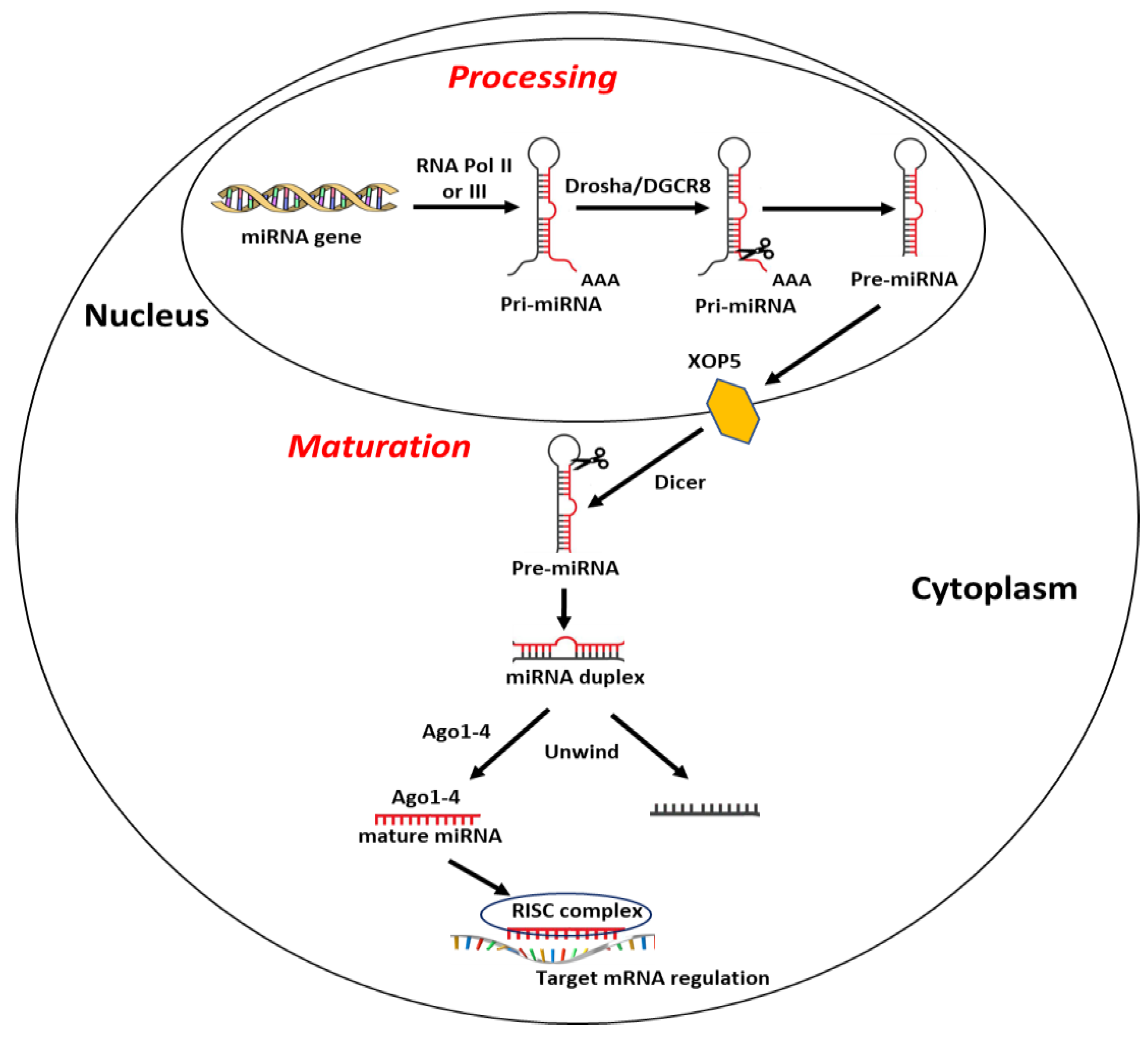
Most miRNA sequences are located within specific gene loci and about 30% of these are inside intronic gene sequences [88]. Briefly, biogenesis of miRNAs begins with their transcription in capped and polyadenylated long primary transcripts (pri-miRNAs) by RNA polymerase II or III [88]. Pri-miRNAs are then processed in the precursor miRNA (pre-miRNA), long 60–120 nucleotides, by the protein complex formed by RNase III endonuclease Drosha and the double-stranded RNA-binding protein DGCR8 (DiGeorge syndrome critical region gene 8) [89]. To prevent the nuclease degradation and easing its translocation from nucleus to cytoplasm, pre-miRNA is then assembled into the complex nucleocytoplasmic transporter factor Exportin-5 (XPO5) and Ran/GTP [90]. In the cytoplasm, the RNase II enzyme Dicer complex cuts the hairpin loop and produces a miRNA duplex of ~22 bp, containing a passenger strand, normally degraded, and a mature miRNA strand, which is loaded onto argonaute protein (Ago1–4) and united with the RNA-induced silencing complex (RISC) [91,92,93]. Finally, miRNA–RISC complex binds and regulates the expression of target mRNA [82] (Figure 1). Of note, a single miRNA may target and affect the expression of several mRNAs, and a single transcript may be targeted by numerous miRNAs [94].
5. miRNA Expression and Regulation in Breast Cancer
Currently, miRBase database (release 22.1) reports information about 1917 human precursors and 2656 mature miRNAs (http://www.mirbase.org/) whose ~ 2300 have been validated in a recent study [95]. The involvement of miRNAs in breast cancer development and progression it has been well demonstrated, thus miRNAs may be considered a potential diagnostic and prognostic markers, as well as therapeutic targets. To date, in November 2020, “miRNAs and breast cancer” keywords in PubMed database yielded 6227 papers, of which about 96% were published during the past decade (https://pubmed.ncbi.nlm.nih.gov/). In Table 1 are listed and referred the more representative miRNAs modulated in breast cancer [96,97,98,99,100,101,102,103,104,105,106,107,108,109,110,111,112,113,114,115,116,117,118,119,120,121,122,123,124,125,126,127,128,129,130,131,132,133,134,135,136,137,138,139,140,141,142,143,144,145,146,147,148,149,150,151,152,153,154,155,156,157,158,159,160,161,162,163,164,165,166,167,168,169,170,171,172,173,174,175], one of the first solid tumors profiled for miRNAs expression [122].
Studies performed by Iorio and colleagues revelled a miRNA differential modulation in breast cancer; for instance, the expression of miR-125b and miR-145 was found down-regulated, while miR-122 and miR-155 showed an up-regulation [122], suggesting a putative tumor suppressor or oncogenic action, respectively. miRNA-21 is the more expressed in breast tumor tissue as compared to matched normal tissue. In addition, miR-21 has been associated with advanced clinical stage, lymph node metastasis and poor prognosis [176]. Moreover, it has been reported that the over-expression of miR-21 is able to mediate cell survival and proliferation by targeting several potential tumor suppressor genes, like phosphatase and tensin homolog (PTEN) [177], tropomyosin 1 (TPM1) [178] and programmed cell death 4 (PDCD4) [179]. On the contrary, the ectopic expression of miR-205 in breast cancer cells reduced cell growth effects and improved the responsiveness to tyrosine kinase inhibitor, gefitinib [180].
miRNAs were also related to certain biological features of breast cancer, like ER and progesterone receptor (PR) expression, as well as the tumor stage and invasion [122]. In this context, it was reported that E2-ERα binding modulates miRNA-191 and miRNA-425 expression in breast cancer cells [181]. Furthermore, it has been shown that certain miRNAs may regulate ERα levels, impairing estrogenic pro-tumorogenic action in breast cancer [122]. For instance, miRNA-206 targets directly ERα [122], whereas miRNA-221 and miRNA-222 may confer a tamoxifen resistance, regulating the expression of p27 [182] and ERα [183].
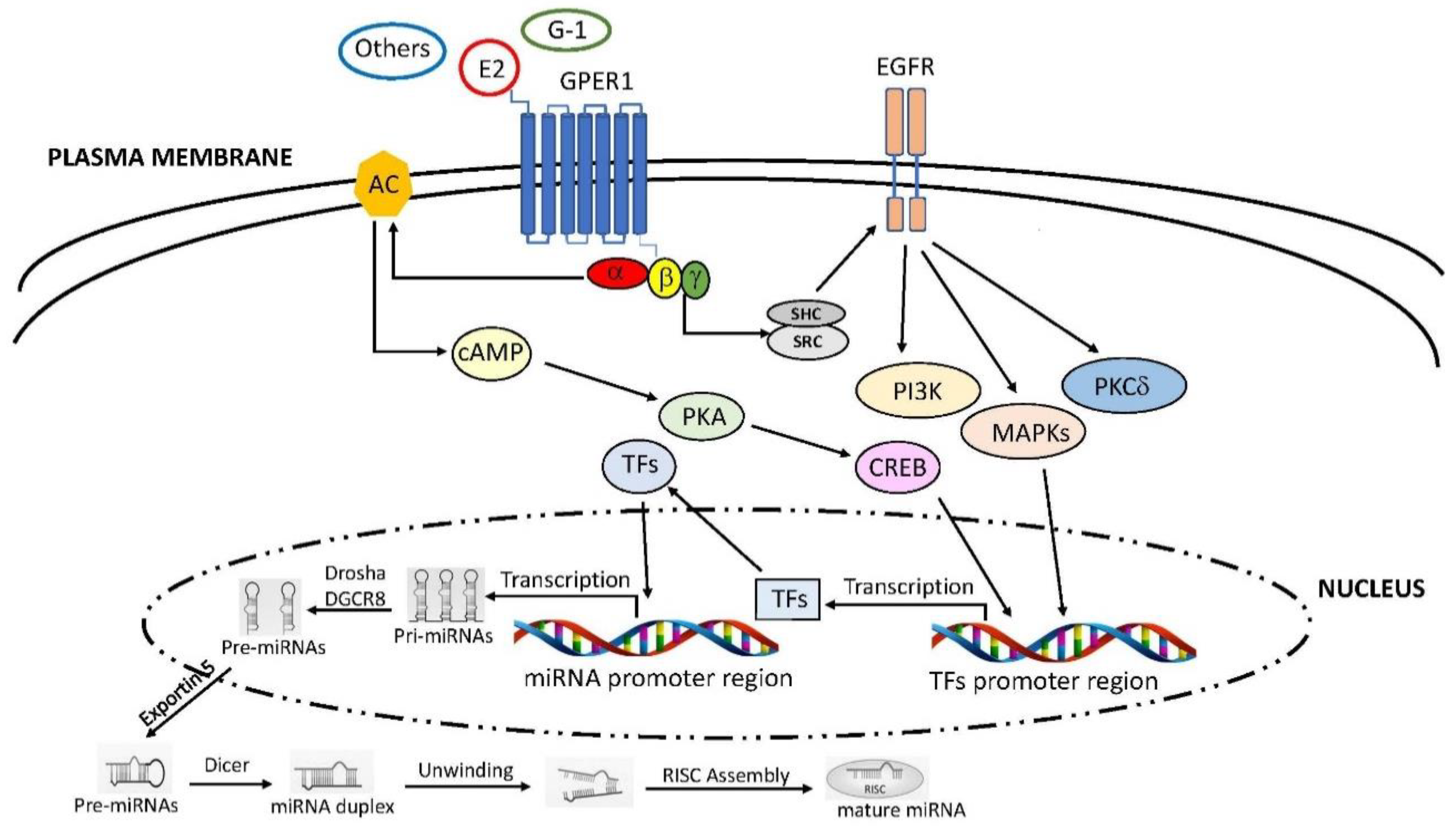
Emerging data have reported that estrogenic stimuli are able to regulate miRNAs expression not only through the classic ERs, but also by the GPER1 involvement [184,185] (Figure 2).
In particular, it has been shown that in breast cancer cells the activation of GPER by E2 or G-1 lead to down-regulation of the tumor-suppressor miR-148a, in a dose and time dependent manner [186]. In turn, the reduced levels of miR-148a increase the expression of its target gene, named human leukocyte antigen-G (HLA-G), known to be implicated in the tumor-driven immune escape in malignancies [186]. In subsequent experiments, Tao and collaborators also reported that miR-148a downregulation mediated by E2-GPER1 signalling induces migratory effects in triple-negative breast cancer cells by overexpressing the lncRNA HOTAIR [187]. Consistent with this evidence, data obtained in ER-negative and GPER1-positive cancer types have indicated that the over-expression of miR-144, induced by E2 and G-1 through the GPER1/PI3K/Akt/ERK/Elk1 axis, lead to reduced levels of the tumor-suppressor Runt-related transcription factor-1 (Runx-1) and the consequent induction of cell cycle progression [9]. Of note, these results were also confirmed in tumor xenografts upon G-1 treatment [9]. More recently, it has been shown that in ER-negative and GPER1-positive breast cancer cells, E2 is able to modulate the expression of diverse miRNAs [8]. Among these, the tumor suppressor miR-338-3p was similar down-regulated in both cell types. In particular, miR-338-3p levels were reduced by E2 or G1 through GPER1, causing an increased cancer cell progression mediated by the augmented expression of its target gene, the proto-oncogene c-Fos [8]. Interestingly, E2 was also able to trigger a differential landscape of miRNAs in GPER1 positive and ER-negative CAFs derived from primary and metastatic breast tumors [7], suggesting a fine regulation of miRNA expression by GPER in very different cancer microenvironments. In particular, E2 modulated miRNA expression three times more in CAFs derived from the metastatic breast cancer respect to CAFs obtained from primary breast tumors. Moreover, in CAFs derived from the metastatic breast cancer, the estrogen stimulation triggered a huge miRNA increase with respect to CAFs from primary breast tumors [7], providing further findings concerning the involvement of certain miRNAs in different phases of the metastatic process [188]. Interestingly, in CAFs derived from metastatic tumor the expression of one of the most induced miRNAs, named miR-623, returned to basal levels by silencing GPER1 expression, suggesting the involvement of this receptor in the regulation of miR-623 [189].
6. Conclusions
Various evidence suggests that GPER plays a crucial role in breast cancer in mediating estrogenic modulation of several target genes involved in proliferation, apoptosis, migration and metastasis. In addition, studies in the past decades have helped to elucidate the importance of miRNA regulation in a number of diseases, including breast cancer. Findings that GPER activation by E2 or G1 may regulate the expression of certain miRNAs and trigger tumorigenic effects in breast cancer, provide new knowledge of the molecular mechanism involved in this neoplasia that would be useful, not only toward innovative therapeutic approaches, but also to understand the failure of treatment in oncological patients.
Funding
This work was supported by Italian Ministry of Research and University (MIUR) MIUR ex 60%.
Conflicts of Interest
The author declares no conflict of interest.
References
- Globocan. Estimated Cancer Incidence, Mortality et Prevalence Worldwide in 2018. 2018. Available online: http://globocan.iarc.fr/Default.aspx (accessed on 23 December 2020).
- Harbeck, N.; Penault-Llorca, F.; Cortes, J.; Gnant, M.; Houssami, N.; Poortmans, P.; Ruddy, K.; Tsang, J.; Cardoso, F. Breast cancer. Nat. Rev. Dis. Primers 2019, 5, 66. [Google Scholar] [CrossRef] [PubMed]
- Yeo, S.K.; Guan, J.L. Breast Cancer: Multiple Subtypes within a Tumor? Trends Cancer 2017, 3, 753–760. [Google Scholar] [CrossRef] [PubMed]
- Fidler, I.J. The pathogenesis of cancer metastasis: The ‘seed and soil’ hypothesis revisited. Nat. Rev. Cancer 2003, 3, 453–458. [Google Scholar] [CrossRef] [PubMed]
- Heldring, N.; Pike, A.; Andersson, S.; Matthews, J.; Cheng, G.; Treuter, E.; Warner, M.; Hartman, J.; Tujague, M.; Stro, A. Estrogen Receptors: How Do They Signal and What Are Their Targets. Physiol. Rev. 2007, 87, 905–931. [Google Scholar] [CrossRef] [Green Version]
- Xu, S.; Yu, S.; Dong, D.; Lee, L.T.O. G Protein-Coupled Estrogen Receptor: A Potential Therapeutic Target in Cancer. Front. Endocrinol. 2019, 10, 725. [Google Scholar] [CrossRef] [Green Version]
- Vivacqua, A.; Muoio, M.G.; Miglietta, A.M.; Maggiolini, M. Differential MicroRNA Landscape Triggered by Estrogens in Cancer Associated Fibroblasts (CAFs) of Primary and Metastatic Breast Tumors. Cancers 2019, 11, 412. [Google Scholar] [CrossRef] [Green Version]
- Vivacqua, A.; Sebastiani, A.; Miglietta, A.M.; Rigiracciolo, D.C.; Cirillo, F.; Galli, G.R.; Talia, M.; Santolla, M.F.; Lappano, R.; Giordano, F.; et al. miR-338-3p Is Regulated by Estrogens through GPER in Breast Cancer Cells and Cancer-Associated Fibroblasts (CAFs). Cells 2018, 7, 203. [Google Scholar] [CrossRef] [Green Version]
- Vivacqua, A.; De Marco, P.; Santolla, M.F.; Cirillo, F.; Pellegrino, M.; Panno, M.L.; Abonante, S.; Maggiolini, M. Estrogenic gper signaling regulates mir144 expression in cancer cells and cancer-associated fibroblasts (cafs). Oncotarget 2015, 6, 16573–16587. [Google Scholar] [CrossRef]
- Rzeszutek, I.; Singh, A. Small RNAs, Big Diseases. Int. J. Mol. Sci. 2020, 21, 5699. [Google Scholar] [CrossRef]
- Vivacqua, A.; De Marco, P.; Belfiore, A.; Maggiolini, M. Recent Advances on the Role of microRNAs in both Insulin Resistance and Cancer. Curr. Pharm. Des. 2017, 23, 3658–3666. [Google Scholar] [CrossRef]
- Carmeci, C.; Thompson, D.A.; Ring, H.Z.; Francke, U.; Weigel, R.J. Identification of a gene (GPR30) with homology to the G-protein-coupled receptor superfamily associated with estrogen receptor expression in breast cancer. Genomics 1997, 45, 607–617. [Google Scholar] [CrossRef] [PubMed]
- Takada, Y.; Kato, C.; Kondo, S.; Korenaga, R.; Ando, J. Cloning of cDNAs encoding G protein-coupled receptor expressed in human endothelial cells exposed to fluid shear stress. Biochem. Biophys. Res. Commun. 1997, 240, 737–741. [Google Scholar] [CrossRef] [PubMed]
- Feng, Y.; Gregor, P. Cloning of a novel member of the G protein-coupled receptor family related to peptide receptors. Biochem. Biophys. Res. Commun. 1997, 231, 651–654. [Google Scholar] [CrossRef] [PubMed]
- Filardo, E.J.; Quinn, J.A.; Bland, K.I.; Frackelton, A.R., Jr. Estrogen-induced activation of Erk-1 and Erk-2 requires the G protein-coupled receptor homolog, GPR30, and occurs via trans-activation of the epidermal growth factor receptor through release of HB-EGF. Mol. Endocrinol. 2000, 14, 1649–1660. [Google Scholar] [CrossRef]
- Maggiolini, M.; Vivacqua, A.; Fasanella, G.; Recchia, A.G.; Sisci, D.; Pezzi, V.; Montanaro, D.; Musti, A.M.; Picard, D.; Andò, S. The G protein-coupled receptor GPR30 mediates c-fos up-regulation by 17beta-estradiol and phytoestrogens in breast cancer cells. J. Biol. Chem. 2004, 279, 27008–27016. [Google Scholar] [CrossRef] [Green Version]
- Thomas, P.; Pang, Y.; Filardo, E.J.; Dong, J. Identity of an estrogen membrane receptor coupled to a G protein in human breast cancer cells. Endocrinology 2005, 146, 624–632. [Google Scholar] [CrossRef] [Green Version]
- Revankar, C.M.; Cimino, D.F.; Sklar, L.A.; Arterburn, J.B.; Prossnitz, E.R. A transmembrane intracellular estrogen receptor mediates rapid cell signaling. Science 2005, 307, 1625–1630. [Google Scholar] [CrossRef] [Green Version]
- Hsieh, Y.C.; Yu, H.P.; Frink, M.; Suzuki, T.; Choudhry, M.A.; Schwacha, M.G.; Chaudry, I.H. G protein-coupled receptor 30-dependent protein kinase a pathway is critical in nongenomic effects of estrogen in attenuating liver injury after trauma-hemorrhage. Am. J. Pathol. 2007, 170, 1210–1218. [Google Scholar] [CrossRef] [Green Version]
- Alexander, S.P.; Mathie, A.; Peters, J.A. Guide to receptors and channels (GRAC), 5th edition. Br. J. Pharmacol. 2011, 164, S16–S19. [Google Scholar] [CrossRef] [Green Version]
- Filardo, E.J.; Quinn, J.A.; Frackelton, A.R., Jr.; Bland, K.I. Estrogen action via the G protein-coupled receptor, GPR30: Stimulation of adenylyl cyclase and cAMP-mediated attenuation of the epidermal growth factor receptor-to-MAPK signaling axis. Mol. Endocrinol. 2002, 16, 70–84. [Google Scholar] [CrossRef]
- Owman, C.; Blay, P.; Nilsson, C.; Lolait, S.J. Cloning of human cDNA encoding a novel heptahelix receptor expressed in Burkitt’s lymphoma and widely distributed in brain and peripheral tissues. Biochem. Biophys. Res. Commun. 1996, 228, 285–292. [Google Scholar] [CrossRef] [PubMed]
- Filardo, E.J.; Thomas, P. Minireview: G protein-coupled estrogen receptor-1, GPER-1: Its mechanism of action and role in female reproductive cancer, renal and vascular physiology. Endocrinology 2012, 153, 2953–2962. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kvingedal, A.M.; Smeland, E.B. A novel putative G-protein-coupled receptor expressed in lung, heart and lymphoid tissue. FEBS Lett. 1997, 407, 59–62. [Google Scholar] [CrossRef]
- Hugo, E.R.; Brandebourg, T.D.; Woo, J.G.; Loftus, J.; Alexander, J.W.; Ben-Jonathan, N. Bisphenol A at environmentally relevant doses inhibits adiponectin release from human adipose tissue explants and adipocytes. Environ. Health Perspect. 2008, 116, 1642–1647. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shi, H.F.; Kumar, S.P.; Liu, X. G protein-coupled estrogen receptor in energy homeostasis and obesity pathogenesis. Prog. Mol. Biol. Transl. Sci. 2013, 114, 193–250. [Google Scholar] [CrossRef] [Green Version]
- Baltgalvis, K.A.; Greising, S.M.; Warren, G.L.; Lowe, D.A. Estrogen regulates estrogen receptors and antioxidant gene expression in mouse skeletal muscle. PLoS ONE 2010, 5, e10164. [Google Scholar] [CrossRef] [Green Version]
- Pollanen, E.; Sipila, S.; Alen, M.; Ronkainen, P.H.; Ankarberg-Lindgren, C.; Puolakka, J.; Suominen, H.; Hämäläinen, E.; Turpeinen, U.; Konttinen, Y.T.; et al. Differential influence of peripheral and systemic sex steroids on skeletal muscle quality in pre-and postmenopausal women. Aging Cell 2011, 10, 650–660. [Google Scholar] [CrossRef] [Green Version]
- Prossnitz, E.R.; Arterburn, J.B.; Smith, H.O.; Oprea, T.I.; Sklar, L.A.; Hathaway, H.J. Estrogen signaling through the transmembrane G protein–coupled receptor GPR30. Ann. Rev. Physiol. 2008, 70, 165–190. [Google Scholar] [CrossRef]
- Filardo, E.J.; Graeber, C.T.; Quinn, J.A.; Resnick, M.B.; Giri, D.; DeLellis, R.A.; Steinhoff, M.M.; Sabo, E. Distribution of GPR30, a seven membrane-spanning estrogen receptor, in primary breast cancer and its association with clinicopathologic determinants of tumor progression. Clin. Cancer Res. 2006, 12, 6359–6366. [Google Scholar] [CrossRef] [Green Version]
- Smith, H.O.; Leslie, K.K.; Singh, M.; Qualls, C.R.; Revankar, C.M.; Joste, N.; Prossnitz, E.R. GPR-30: A novel indicator of poor survival in endometrial carcinoma. Am. J. Obstet. Gynecol. 2007, 196, 386.e1–386.e11. [Google Scholar] [CrossRef]
- Filardo, E.J.; Quinn, J.A.; Sabo, E. Association of the membrane estrogen receptor, GPR30, with breast tumor metastasis and transactivation of the epidermal growth factor receptor. Steroids 2008, 73, 870–873. [Google Scholar] [CrossRef] [PubMed]
- Smith, H.O.; Arias-Pulido, H.; Kuo, D.; Howard, T.; Qualls, C.; Lee, S.J.; Verschraegen, C.; Hathaway, H.; Joste, N.; Prossnitz, E. GPR30 predicts poor survival for ovarian cancer. Gynecol. Oncol. 2009, 114, 465–471. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ignatov, T.; Weißenborn, C.; Poehlmann, A.; Lemke, A.; Semczuk, A.; Roessner, A.; Costa, S.D.; Kalinski, T.; Ignatov, A. GPER-1 expression decreases during breast cancer tumorigenesis. Cancer Investig. 2013, 31, 309–315. [Google Scholar] [CrossRef] [PubMed]
- Treeck, O.; Schüler-Toprak, S.; Ortmann, O. Estrogen Actions in Triple-Negative Breast Cancer. Cells 2020, 9, 2358. [Google Scholar] [CrossRef] [PubMed]
- Ignatov, A.; Ignatov, T.; Weissenborn, C.; Eggemann, H.; Bischoff, J.; Semczuk, A.; Roessner, A.; Costa, S.D.; Kalinski, T. G-protein-coupled estrogen receptor GPR30 and tamoxifen resistance in breast cancer. Breast Cancer Res. Treat. 2011, 128, 457–466. [Google Scholar] [CrossRef]
- Steiman, J.; Peralta, E.A.; Louis, S.; Kamel, O. Biology of the estrogen receptor, GPR30, in triple negative breast cancer. Am. J. Surg. 2013, 206, 698–703. [Google Scholar] [CrossRef]
- Kelly, M.J.; Levin, E.R. Rapid actions of plasma membrane estrogen receptors. Trends Endocrinol. Metab. 2001, 12, 152–156. [Google Scholar] [CrossRef]
- Funakoshi, T.; Yanai, A.; Shinoda, K.; Kawano, M.M.; Mizukami, Y. G protein-coupled receptor 30 is an estrogen receptor in the plasma membrane. Biochem. Biophys. Res. Commun. 2006, 346, 904–910. [Google Scholar] [CrossRef]
- Sandén, C.; Broselid, S.; Cornmark, L.; Andersson, K.; Daszkiewicz-Nilsson, J.; Mårtensson, U.E.; Olde, B.; Leeb-Lundberg, L.M. G protein-coupled estrogen receptor 1/G protein-coupled receptor 30 localizes in the plasma membrane and traffics intracellularly on cytokeratin intermediate filaments. Mol. Pharmacol. 2011, 79, 400–410. [Google Scholar] [CrossRef]
- Filardo, E.; Quinn, J.; Pang, Y.; Graeber, C.; Shaw, S.; Dong, J.; Thomas, P. Activation of the novel estrogen receptor G protein-coupled receptor 30 (GPR30) at the plasma membrane. Endocrinology 2007, 148, 3236–3245. [Google Scholar] [CrossRef] [Green Version]
- Gao, F.; Ma, X.; Ostmann, A.B.; Das, S.K. GPR30 activation opposes estrogen-dependent uterine growth via inhibition of stromal ERK1/2 and estrogen receptor alpha (ERα) phosphorylation signals. Endocrinology 2011, 152, 1434–1447. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maiti, K.; Paul, J.; Read, M.; Chan, E.; Riley, S.; Nahar, P.; Smith, R. G-1-activated membrane estrogen receptors mediate increased contractility of the human myometrium. Endocrinology 2011, 152, 2448–2455. [Google Scholar] [CrossRef] [Green Version]
- Cheng, S.B.; Graeber, C.T.; Quinn, J.A.; Filardo, E.J. Retrograde transport of the transmembrane estrogen receptor, G-protein-coupled-receptor-30 (GPR30/GPER) from the plasma membrane towards the nucleus. Steroids 2011, 76, 892–896. [Google Scholar] [CrossRef]
- Lindsey, S.H.; Yamaleyeva, L.M.; Brosnihan, K.B.; Gallagher, P.E.; Chappell, M.C. Estrogen receptor GPR30 reduces oxidative stress and proteinuria in the salt-sensitive female mRen2. Lewis rat. Hypertension 2011, 58, 665–671. [Google Scholar] [CrossRef] [Green Version]
- Cheng, S.B.; Dong, J.; Pang, Y.; LaRocca, J.; Hixon, M.; Thomas, P.; Filardo, E.J. Anatomical location and redistribution of G protein-coupled estrogen receptor-1 during the estrus cycle in mouse kidney and specific binding to estrogens but not aldosterone. Mol. Cell. Endocrinol. 2014, 382, 950–959. [Google Scholar] [CrossRef] [PubMed]
- Akama, K.T.; Thompson, L.I.; Milner, T.A.; McEwen, B.S. Post-synaptic density-95 (PSD-95) binding capacity of G-protein-coupled receptor 30 (GPR30), an estrogen receptor that can be identified in hippocampal dendritic spines. J. Biol. Chem. 2013, 288, 6438–6450. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meyer, M.R.; Fredette, N.C.; Howard, T.A.; Hu, C.; Ramesh, C.; Daniel, C.; Amann, K.; Arterburn, J.B.; Barton, M.; Prossnitz, E.R. G protein-coupled estrogen receptor protects from atherosclerosis. Sci. Rep. 2014, 4, 7564. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Balhuizen, A.; Kumar, R.; Amisten, S.; Lundquist, I.; Salehi, A. Activation of G protein-coupled receptor 30 modulates hormone secretion and counteracts cytokine-induced apoptosis in pancreatic islets of female mice. Mol. Cell. Endocrinol. 2010, 320, 16–24. [Google Scholar] [CrossRef] [Green Version]
- Meyer, M.R.; Clegg, D.J.; Prossnitz, E.R.; Barton, M. Obesity, insulin resistance and diabetes: Sex differences and role of oestrogen receptors. Acta Physiol. 2011, 203, 259–269. [Google Scholar] [CrossRef]
- Madeo, A.; Maggiolini, M. Nuclear alternate estrogen receptor GPR30 mediates 17beta-estradiol-induced gene expression and migration in breast cancer-associated fibroblasts. Cancer Res. 2010, 70, 6036–6046. [Google Scholar] [CrossRef] [Green Version]
- Lappano, R.; Rigiracciolo, D.C.; Belfiore, A.; Maggiolini, M.; De Francesco, E.M. Cancer associated fibroblasts: Role in breast cancer and potential as therapeutic targets. Expert Opin. Ther. Targets 2020, 24, 559–572. [Google Scholar] [CrossRef] [PubMed]
- Prossnitz, E.R.; Barton, M. The G-protein-coupled estrogen receptor GPER in health and disease. Nat. Rev. Endocrinol. 2011, 7, 715–726. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grande, F.; Occhiuzzi, M.A.; Lappano, R.; Cirillo, F.; Guzzi, R.; Garofalo, A.; Jacquot, Y.; Maggiolini, M.; Rizzuti, B. Computational Approaches for the Discovery of GPER Targeting Compounds. Front. Endocrinol. 2020, 11, 517. [Google Scholar] [CrossRef] [PubMed]
- Molina, L.; Bustamante, F.A.; Bhoola, K.D.; Figueroa, C.D.; Ehrenfeld, P. Possible role of phytoestrogens in breast cancer via GPER-1/GPR30 signaling. Clin. Sci. 2018, 132, 2583–2598. [Google Scholar] [CrossRef]
- Gao, F.; Huang, Y.; Zhang, L.; Liu, W. Involvement of estrogen receptor and GPER in bisphenol A induced proliferation of vascular smooth muscle cells. Toxicol. In Vitro 2019, 56, 156–162. [Google Scholar] [CrossRef]
- Castillo-Sanchez, R.; Ramirez-Ricardo, J.; Martinez-Baeza, E.; Cortes-Reynosa, P.; Candanedo-Gonzales, F.; Gomez, R.; Salazar, E.P. Bisphenol A induces focal adhesions assembly and activation of FAK, Src and ERK2 via GPER in MDA-MB-231 breast cancer cells. Toxicol. In Vitro 2020, 66, 104871. [Google Scholar] [CrossRef]
- Vivacqua, A.; Bonofiglio, D.; Recchia, A.G.; Musti, A.M.; Picard, D.; Andò, S.; Maggiolini, M. The G protein-coupled receptor GPR30 mediates the proliferative effects induced by 17b-estradiol and hydroxytamoxifen in endometrial cancer cells. Mol. Endocrinol. 2006, 20, 631–646. [Google Scholar] [CrossRef] [Green Version]
- Bologa, C.G.; Revankar, C.M.; Young, S.M.; Edwards, B.S.; Arterburn, J.B.; Kiselyov, A.S.; Parker, M.A.; Tkachenko, S.E.; Savchuck, N.P.; Sklar, L.A.; et al. Virtual and biomolecular screening converge on a selective agonist for GPR30. Nat. Chem. Biol. 2006, 2, 207–212. [Google Scholar] [CrossRef]
- Dennis, M.K.; Field, A.S.; Burai, R.; Ramesh, C.; Petrie, W.K.; Bologa, C.G.; Oprea, T.I.; Yamaguchi, Y.; Hayashi, S.-I.; Sklar, L.A.; et al. Identification of a GPER/GPR30 antagonist with improved estrogen receptor counterselectivity. J. Steroid Biochem. Mol. Biol. 2011, 127, 358–366. [Google Scholar] [CrossRef] [Green Version]
- Lappano, R.; Rosano, C.; Santolla, M.F.; Pupo, M.; De Francesco, E.M.; De Marco, P.; Ponassi, M.; Spallarossa, A.; Ranise, A.; Maggiolini, M. Two novel GPER agonists induce gene expression changes and growth effects in cancer cells. Cur. Cancer Drug Targets 2012, 12, 531–542. [Google Scholar] [CrossRef]
- Petrie, W.K.; Dennis, M.K.; Hu, C.; Dai, D.; Arterburn, J.B.; Smith, H.O.; Hathaway, H.J.; Prossnitz, E.R. G protein-coupled estrogen receptor-selective ligands modulate endometrial tumor growth. Obstet. Gynecol. Int. 2013, 2013. [Google Scholar] [CrossRef] [Green Version]
- Coelingh Bennink, H.J. Are all estrogens the same? Maturitas 2004, 47, 269–275. [Google Scholar] [CrossRef]
- Lappano, R.; Santolla, M.F.; Pupo, M.; Sinicropi, M.S.; Caruso, A.; Rosano, C.; Maggiolini, M. MIBE acts as antagonist ligand of both estrogen receptor a and GPER in breast cancer cells. Breast Cancer Res. 2012, 14, R12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lappano, R.; Mallet, C.; Rizzuti, B.; Grande, F.; Galli, G.R.; Byrne, C.; Broutin, I.; Boudieu, L.; Eschalier, A.; Jacquot, Y.; et al. The Peptide ERα17p Is a GPER Inverse Agonist that Exerts Antiproliferative Effects in Breast Cancer Cells. Cells 2019, 8, 590. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Trichet, M.; Lappano, R.; Belnou, M.; Salazar Vazquez, L.S.; Alves, I.; Ravault, D.; Sagan, S.; Khemtemourian, L.; Maggiolini, M.; Jacquot, Y. Interaction of the Anti-Proliferative GPER Inverse Agonist ERα17p with the Breast Cancer Cell Plasma Membrane: From Biophysics to Biology. Cells 2020, 9, 447. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pandey, D.P.; Lappano, R.; Albanito, L.; Madeo, A.; Maggiolini, M.; Picard, D. Estrogenic GPR30 signalling induces proliferation and migration of breast cancer cells through CTGF. EMBO J. 2009, 28, 523–532. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vivacqua, A.; Romeo, E.; De Marco, P.; De Francesco, E.M.; Abonante, S.; Maggiolini, M. GPER mediates the Egr-1 expression induced by 17beta-estradiol and 4-hydroxitamoxifen in breast and endometrial cancer cells. Breast Cancer Res. Treat. 2012, 133, 1025–1035. [Google Scholar] [CrossRef]
- De Francesco, E.M.; Sotgia, F.; Clarke, R.B.; Lisanti, M.P.; Maggiolini, M. G Protein-Coupled Receptors at the Crossroad between Physiologic and Pathologic Angiogenesis: Old Paradigms and Emerging Concepts. Int. J. Mol. Sci. 2017, 18, 2713. [Google Scholar] [CrossRef] [Green Version]
- Barton, M.; Filardo, E.J.; Lolait, S.J.; Thomas, P.; Maggiolini, M.; Prossnitz, E.R. Twenty years of the G protein-coupled estrogen receptor GPER: Historical and personal perspectives. J. Steroid Biochem. Mol. Biol. 2018, 4–15. [Google Scholar] [CrossRef] [Green Version]
- Molina, L.; Figueroa, C.D.; Bhoola, K.D.; Ehrenfeld, P. GPER-1/GPR30 a novel estrogen receptor sited in the cell membrane: Therapeutic coupling to breast cancer. Expert Opin. Ther. Targets 2017, 21, 755–766. [Google Scholar] [CrossRef]
- Lappano, R.; Rigiracciolo, D.; De Marco, P.; Avino, S.; Cappello, A.R.; Rosano, C.; Maggiolini, M.; De Francesco, E.M. Recent advances on the role of G protein-coupled receptors in hypoxia-mediated signaling. AAPS J. 2016, 18, 305–310. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Marco, P.; Lappano, R.; De Francesco, E.M.; Cirillo, F.; Pupo, M.; Avino, S.; Vivacqua, A.; Abonante, S.; Picard, D.; Maggiolini, M. GPER signalling in both cancer-associated fibroblasts and breast cancer cells mediates a feedforward IL1β/IL1R1 response. Sci. Rep. 2016, 6, 24354. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jung, Y.Y.; Kim, H.M.; Koo, J.S. The role of cancer-associated fibroblasts in breast cancer pathobiology. Histol. Histopathol. 2016, 31, 371–378. [Google Scholar] [CrossRef] [PubMed]
- Lee, R.; Feinbaum, R.; Ambros, V. The C. elegans heterochronic gene lin-4 encodes small RNAs with antisense complementarity to lin-14. Cell 1993, 75, 843–854. [Google Scholar] [CrossRef]
- Reinhart, B.J.; Slack, F.J.; Basson, M.; Pasquinelli, A.E.; Bettinger, J.C.; Rougvie, A.E.; Horvitz, H.R.; Ruvkun, G. The 21-nucleotide let-7 RNA regulates developmental timing in Caenorhabditis elegans. Nature 2000, 403, 901–906. [Google Scholar] [CrossRef]
- Lagos-Quintana, M.; Rauhut, R.; Lendeckel, W.; Tuschl, T. Identification of novel genes coding for small expressed RNAs. Science 2001, 294, 853–858. [Google Scholar] [CrossRef] [Green Version]
- Grimson, A.; Farh, K.K.; Johnston, W.K.; Garrett-Engele, P.; Lim, L.P.; Bartel, D.P. MicroRNA targeting specificity in mammals: Determinants beyond seed pairing. Mol. Cell 2007, 27, 91–105. [Google Scholar] [CrossRef] [Green Version]
- Bartel, D.P. MicroRNAs: Genomics, biogenesis, mechanism, and function. Cell 2004, 116, 281–297. [Google Scholar] [CrossRef] [Green Version]
- Pillai, R.S. MicroRNA function: Multiple mechanisms for a tiny RNA? RNA 2005, 11, 1753–1761. [Google Scholar] [CrossRef] [Green Version]
- Li, G.; Wu, X.; Qian, W.; Cai, H.; Sun, X.; Zhang, W.; Tan, S.; Wu, Z.; Qian, P.; Ding, K.; et al. CCAR1 5′ UTR as a natural miRancer of miR1254 overrides tamoxifen resistance. Cell Res. 2016, 26, 655–673. [Google Scholar] [CrossRef] [Green Version]
- Bartel, D.P. MicroRNAs: Target recognition and regulatory functions. Cell 2009, 136, 215–233. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Esquela-Kerscher, A.; Slack, F.J. Oncomirs—MicroRNAs with a role in cancer. Nat. Rev. Cancer 2006, 6, 259–269. [Google Scholar] [CrossRef]
- Chen, C.Z.; Li, L.; Lodish, H.F.; Bartel, D.P. MicroRNAs modulate hematopoietic lineage differentiation. Science 2004, 303, 83–86. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Latronico, M.V.; Catalucci, D.; Condorelli, G. MicroRNA and cardiac pathologies. Physiol. Genom. 2008, 34, 239–342. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miao, C.; Chang, J.; Zhang, G.; Fang, Y. MicroRNAs in type 1 diabetes: New research progress and potential directions. Biochem. Cell Biol. 2018, 96, 498–506. [Google Scholar] [CrossRef] [PubMed]
- Svoronos, A.A.; Engelman, D.M.; Slack, F.J. OncomiR or Tumor Suppressor? The Duplicity of MicroRNAs in Cancer. Cancer Res. 2016, 76, 3666–3670. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, S.; Gregory, R.I. MicroRNA biogenesis pathways in cancer. Nat. Rev. Cancer 2015, 15, 321–333. [Google Scholar] [CrossRef]
- Denli, A.M.; Tops, B.B.J.; Plasterk, R.H.A.; Ketting, R.F.; Hannon, G.J. Processing of primary microRNAs by the microprocessor complex. Nature 2004, 432, 231–235. [Google Scholar] [CrossRef]
- Okada, C.; Yamashita, E.; Lee, S.J.; Shibata, S.; Katahira, J.; Nakagawa, A.; Yoneda, Y.; Tsukihara, T. A high-resolution structure of the pre-microrna nuclear export machinery. Science 2009, 326, 1275–1279. [Google Scholar] [CrossRef]
- Hutvágner, G.; McLachlan, J.; Pasquinelli, A.E.; Bálint, E.; Tuschl, T.; Zamore, P.D. A cellular function for the RNA-interference enzyme Dicer in the maturation of the let-7 small temporal RNA. Science 2001, 293, 834–838. [Google Scholar] [CrossRef] [Green Version]
- Chendrimada, T.P.; Gregory, R.I.; Kumaraswamy, E.; Norman, J.; Cooch, N.; Nishikura, K.; Shiekhattar, R. TRBP recruits the Dicer complex to Ago2 for microRNA processing and gene silencing. Nature 2005, 436, 740–744. [Google Scholar] [CrossRef]
- Kawamata, T.; Yoda, M.; Tomari, Y. Multilayer checkpoints for microrna authenticity during risc assembly. EMBO Rep. 2011, 12, 944–949. [Google Scholar] [CrossRef] [Green Version]
- Kim, V.N.; Nam, J.W. Genomics of microRNA. Trends Genet. 2006, 22, 165–173. [Google Scholar] [CrossRef] [PubMed]
- Alles, J.; Fehlmann, T.; Fischer, U.; Backes, C.; Galata, V.; Minet, M.; Hart, M.; Abu-Halima, M.; Grässer, F.A.; Lenhof, H.P.; et al. An estimate of the total number of true human miRNAs. Nucleic Acids Res. 2019, 47, 3353–3364. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fu, S.W.; Chen, L.; Man, Y.G. miRNA biomarkers in breast cancer detection and management. J. Cancer 2011, 2, 2116–2122. [Google Scholar] [CrossRef] [PubMed]
- Johnson, S.M.; Grosshans, H.; Shingara, J.; Byrom, M.; Jarvis, R.; Cheng, A.; Labourier, E.; Reinert, K.L.; Brown, D.; Slack, F.J. RAS is regulated by the let-7 microRNA family. Cell 2005, 120, 635–647. [Google Scholar] [CrossRef] [Green Version]
- Wu, L.; Fan, J.; Belasco, J.G. MicroRNAs direct rapid deadenylation of mRNA. Proc. Natl. Acad. Sci. USA 2006, 103, 4034–4039. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mayr, C.; Hemann, M.T.; Bartel, D.P. Disrupting the pairing between let-7 and Hmga2 enhances oncogenic transformation. Science 2007, 315, 1576–1579. [Google Scholar] [CrossRef] [Green Version]
- Ma, L.; Teruya-Feldstein, J.; Weinberg, R.A. Tumour invasion and metastasis initiated by microRNA-10b in breast cancer. Nature 2007, 449, 682–688. [Google Scholar] [CrossRef]
- Loh, H.Y.; Norman, B.P.; Lai, K.S.; Rahman, N.M.A.N.A.; Alitheen, N.B.M.; Osman, M.A. The regulatory role of microRNAs in breast cancer. Int. J. Mol. Sci. 2019, 20, 4940. [Google Scholar] [CrossRef] [Green Version]
- Guo, X.; Connick, M.C.; Vanderhoof, J.; Ishak, M.A.; Hartley, R.S. MicroRNA-16 modulates HuR regulation of cyclin E1 in breast cancer cells. Int. J. Mol. Sci. 2015, 16, 7112–7132. [Google Scholar] [CrossRef] [PubMed]
- Chu, J.; Zhu, Y.; Liu, Y.; Sun, L.; Lv, X.; Wu, Y.; Hu, P.; Su, F.; Gong, C.; Song, E.; et al. E2F7 overexpression leads to tamoxifen resistance in breast cancer cells by competing with E2F1 at miR-15a/16 promoter. Oncotarget 2015, 6, 31944–31957. [Google Scholar] [CrossRef] [PubMed]
- Hossain, A.; Kuo, M.T.; Saunders, G.F. Mir-17-5p regulates breast cancer cell proliferation by inhibiting translation of AIB1 mRNA. Mol. Cell. Biol. 2006, 26, 8191–8201. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, Z.; Wang, C.; Wang, M.; Li, Z.; Casimiro, M.C.; Liu, M.; Wu, K.; Whittle, J.; Ju, X.; Hyslop, T.; et al. A cyclin D1/microRNA 17/20 regulatory feedback loop in control of breast cancer cell proliferation. J. Cell Biol. 2008, 182, 509–517. [Google Scholar] [CrossRef]
- Li, P.; Guo, Y.; Bledsoe, G.; Yang, Z.; Chao, L.; Chao, J. Kallistatin induces breast cancer cell apoptosis and autophagy by modulating Wnt signaling and microRNA synthesis. Exp. Cell Res. 2016, 340, 305–314. [Google Scholar] [CrossRef] [Green Version]
- Song, Y.K.; Wang, Y.; Wen, Y.Y.; Zhao, P.; Bian, Z.J. MicroRNA-22 suppresses breast cancer cell growth and increases paclitaxel sensitivity by targeting NRAS. Technol. Cancer Res. Treat. 2018, 17. [Google Scholar] [CrossRef]
- Roscigno, G.; Puoti, I.; Giordano, I.; Donnarumma, E.; Russo, V.; Affinito, A.; Adamo, A.; Quintavalle, C.; Todaro, M.; Condorelli, G.; et al. MiR-24 induces chemotherapy resistance and hypoxic advantage in breast cancer. Oncotarget 2017, 8, 19507–19521. [Google Scholar] [CrossRef] [Green Version]
- Huang, Z.M.; Ge, H.F.; Yang, C.C.; Cai, Y.; Chen, Z.; Tian, W.Z.; Tao, J.L. MicroRNA-26a-5p inhibits breast cancer cell growth by suppressing RNF6 expression. Kaohsiung J. Med. Sci. 2019, 35, 467–473. [Google Scholar] [CrossRef] [Green Version]
- Liu, J.; Li, X.; Wang, M.; Xiao, G.; Yang, G.; Wang, H.; Li, Y.; Sun, X.; Qin, S.; Du, N.; et al. A miR-26a/E2F7 feedback loop contributes to tamoxifen resistance in ER-positive breast cancer. Int. J. Oncol. 2018, 53, 1601–1612. [Google Scholar] [CrossRef] [Green Version]
- Tormo, E.; Adam-Artigues, A.; Ballester, S.; Pineda, B.; Zazo, S.; González-Alonso, P.; Albanell, J.; Rovira, A.; Rojo, F.; Lluch, A.; et al. The role of miR-26a and miR-30b in HER2+ breast cancer trastuzumab resistance and regulation of the CCNE2 gene. Sci. Rep. 2017, 7, 1–9. [Google Scholar] [CrossRef]
- Mertens-Talcott, S.U.; Chintharlapalli, S.; Li, X.; Safe, S. The oncogenic microRNA-27a targets genes that regulate specificity protein transcription factors and the G2-M checkpoint in MDA-MB-231 breast cancer cells. Cancer Res. 2007, 67, 11001–11011. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Y.; Cai, B.; Shen, L.; Dong, Y.; Lu, Q.; Sun, S.; Liu, S.; Ma, S.; Ma, P.X.; Chen, J. MiRNA-29b suppresses tumor growth through simultaneously inhibiting angiogenesis and tumorigenesis by targeting Akt3. Cancer Lett. 2017, 397, 111–119. [Google Scholar] [CrossRef]
- Shukla, K.; Sharma, A.K.; Ward, A.; Will, R.; Hielscher, T.; Balwierz, A.; Breunig, C.; Münstermann, E.; König, R.; Keklikoglou, I.; et al. MicroRNA-30c-2-3p negatively regulates NF-κB signaling and cell cycle progression through downregulation of TRADD and CCNE1 in breast cancer. Mol. Oncol. 2015, 9, 1106–1119. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Augoff, K.; McCue, B.; Plow, E.F.; Sossey-Alaoui, K. MiR-31 and its host gene lncRNA LOC554202 are regulated by promoter hypermethylation in triple-negative breast cancer. Mol. Cancer 2012, 11. [Google Scholar] [CrossRef] [Green Version]
- Van Schooneveld, E.; Wildiers, H.; Vergote, I.; Vermeulen, P.B.; Dirix, L.Y.; Van Laere, S.J. Dysregulation of microRNAs in breast cancer and their potential role as prognostic and predictive biomarkers in patient management. Breast Cancer Res. 2015, 17, 21. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, Y.; Liu, A.Y.; Fan, C.; Zheng, H.; Li, Y.; Zhang, C.; Wu, S.; Yu, D.; Huang, Z.; Liu, F. MicroRNA-33b inhibits breast cancer metastasis by targeting HMGA2, SALL4 and Twist1. Sci. Rep. 2015, 5, 9995. [Google Scholar] [CrossRef]
- Li, G.; Yao, L.; Zhang, J.; Li, X.; Dang, S.; Zeng, K.; Zhou, Y.; Gao, F. Tumor-suppressive microRNA-34a inhibits breast cancer cell migration and invasion via targeting oncogenic TPD52. Tumor Biol. 2016, 37, 7481–7491. [Google Scholar] [CrossRef] [PubMed]
- Pakravan, K.; Babashah, S.; Sadeghizadeh, M.; Mowla, S.J.; Mossahebi-Mohammadi, M.; Ataei, F.; Dana, N.; Javan, M. MicroRNA-100 shuttled by mesenchymal stem cell-derived exosomes suppresses in vitro angiogenesis through modulating the mTOR/HIF-1α/VEGF signaling axis in breast cancer cells. Cell. Oncol. 2017, 40, 457–470. [Google Scholar] [CrossRef]
- Guan, H.; Dai, Z.; Ma, Y.; Wang, Z.; Liu, X.; Wang, X. MicroRNA-101 inhibits cell proliferation and induces apoptosis by targeting EYA1 in breast cancer. Int. J. Mol. Med. 2016, 37, 1643–1651. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; Zeng, H.; Li, H.; Chen, T.; Wang, L.; Zhang, K.; Chen, J.; Wang, R.; Li, Q.; Wang, S. MicroRNA-101 inhibits growth, proliferation and migration and induces apoptosis of breast cancer cells by targeting sex-determining region Y-Box 2. Cell. Physiol. Biochem. 2017, 43, 717–732. [Google Scholar] [CrossRef]
- Iorio, M.V.; Ferracin, M.; Liu, C.G.; Veronese, A.; Spizzo, R.; Sabbioni, S.; Magri, E.; Pedriali, M.; Fabbri, M.; Campiglio, M.; et al. MicroRNA gene expression deregulation in human breast cancer. Cancer Res. 2005, 65, 7065–7070. [Google Scholar] [CrossRef] [Green Version]
- Shi, P.; Chen, C.; Li, X.; Wei, Z.; Liu, Z.; Liu, Y. MicroRNA-124 suppresses cell proliferation and invasion of triple negative breast cancer cells by targeting STAT3. Mol. Med. Rep. 2019, 49, 3667–3675. [Google Scholar] [CrossRef] [Green Version]
- Jin, T.; Kim, H.S.; Choi, S.K.; Hwang, E.H.; Woo, J.; Ryu, H.S.; Kim, K.; Moon, A.; Moon, W.K. MicroRNA-200c/141 upregulates SerpinB2 to promote breast cancer cell metastasis and reduce patient survival. Oncotarget 2017, 8, 32769–32782. [Google Scholar] [CrossRef] [Green Version]
- Ferracin, M.; Bassi, C.; Pedriali, M.; Pagotto, S.; D’Abundo, L.; Zagatti, B.; Corrà, F.; Musa, G.; Callegari, E.; Lupini, L.; et al. MiR-125b targets erythropoietin and its receptor and their expression correlates with metastatic potential and ERBB2/HER2 expression. Mol. Cancer 2013, 12, 130. [Google Scholar] [CrossRef] [Green Version]
- Alhasan, L. MiR-126 modulates angiogenesis in breast cancer by targeting VEGF-A-mRNA. Asian Pac. J. Cancer Prev. 2019, 20, 193–197. [Google Scholar] [CrossRef] [Green Version]
- Sui, Y.; Zhang, X.; Yang, H.; Wei, W.; Wang, M. MicroRNA-133a acts as a tumour suppressor in breast cancer through targeting LASP1. Oncol. Rep. 2018, 39, 473–482. [Google Scholar] [CrossRef] [Green Version]
- Hua, K.; Jin, J.; Zhao, J.; Song, J.; Song, H.; Li, D.; Maskey, N.; Zhao, B.; Wu, C.; Xu, H.; et al. MiR-135b, upregulated in breast cancer, promotes cell growth and disrupts the cell cycle by regulating LATS2. Int. J. Oncol. 2016, 48, 1997–2006. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.; Liu, D.; Feng, Z.; Mao, J.; Zhang, C.; Lu, Y.; Li, J.; Zhang, Q.; Li, Q.; Li, L. MicroRNA-138 modulates metastasis and EMT in breast cancer cells by targeting vimentin. Biomed. Pharmacother. 2016, 77, 135–141. [Google Scholar] [CrossRef]
- Lu, Y.; Qin, T.; Li, J.; Wang, L.; Zhang, Q.; Jiang, Z.; Mao, J. MicroRNA-140-5p inhibits invasion and angiogenesis through targeting VEGF-A in breast cancer. Cancer Gene Ther. 2017, 24, 386–392. [Google Scholar] [CrossRef] [Green Version]
- Zhang, N.; Li, Z.; Bai, F.; Ji, N.; Zheng, Y.; Li, Y.; Chen, J.; Mao, X. MicroRNA expression profiles in benign prostatic hyperplasia. Mol. Med. Rep. 2018, 17, 3853–3858. [Google Scholar] [CrossRef] [Green Version]
- Sachdeva, M.; Zhu, S.; Wu, F.; Wu, H.; Walia, V.; Kumar, S.; Elble, R.; Watabe, K.; Mo, Y.Y. p53 represses c-Myc through induction of the tumor suppressor miR-145. Proc. Natl. Acad. Sci. USA 2009, 106, 3207–3212. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bhaumik, D.; Scott, G.K.; Schokrpur, S.; Patil, C.K.; Campisi, J.; Benz, C.C. Expression of microRNA-146 suppresses NF-κB activity with reduction of metastatic potential in breast cancer cells. Oncogene 2008, 27, 5643–5647. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shah, R.; Rosso, K.; Nathanson, D.S. Pathogenesis, prevention, diagnosis and treatment of breast cancer. World J. Clin. Oncol. 2014, 5, 283–298. [Google Scholar] [CrossRef] [PubMed]
- Mandujano-Tinoco, E.A.; García-Venzor, A.; Melendez-Zajgla, J.; Maldonado, V. New emerging roles of microRNAs in breast cancer. Breast Cancer Res. Treat. 2018, 171, 247–259. [Google Scholar] [CrossRef] [PubMed]
- Jiang, S.; Zhang, L.F.; Zhang, H.W.; Hu, S.; Lu, M.H.; Liang, S.; Li, B.; Li, Y.; Li, D.; Wang, E.D.; et al. A novel miR-155/miR-143 cascade controls glycolysis by regulating hexokinase 2 in breast cancer cells. EMBO J. 2012, 31, 1985–1998. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nagpal, N.; Ahmad, H.M.; Chameettachal, S.; Sundar, D.; Ghosh, S.; Kulshreshtha, R. HIF-inducible miR-191 promotes migration in breast cancer through complex regulation of TGFβ signaling in hypoxic microenvironment. Sci. Rep. 2015, 5, 1–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sharma, S.; Nagpal, N.; Ghosh, P.C.; Kulshreshtha, R. P53-miR-191-SOX4 regulatory loop affects apoptosis in breast cancer. RNA 2017, 23, 1237–1246. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xie, F.Y.; Hosany, S.; Zhong, S.; Jiang, Y.; Zhang, F.; Lin, L.L.; Wang, X.B.; Gao, S.M.; Hu, X.Q. MicroRNA-193a inhibits breast cancer proliferation and metastasis by downregulating WT1. PLoS ONE 2017, 12, e0185565. [Google Scholar] [CrossRef] [Green Version]
- Yang, L.; Cai, Y.; Zhang, D.; Sun, J.; Xu, C.; Zhao, W.; Jiang, W.; Pan, C. MiR-195/miR-497 regulate CD274 expression of immune regulatory ligands in triple-negative breast cancer. J. Breast Cancer 2018, 21, 371–381. [Google Scholar] [CrossRef]
- McAnena, P.; Tanriverdi, K.; Curran, C.; Gilligan, K.; Freedman, J.E.; Brown, J.A.L.; Kerin, M.J. Circulating microRNAs miR-331 and miR-195 differentiate local luminal a from metastatic breast cancer. BMC Cancer 2019, 19, 436. [Google Scholar] [CrossRef] [Green Version]
- Hong, H.; Yu, H.; Yuan, J.; Guo, C.; Cao, H.; Li, W.; Xiao, C. MicroRNA-200B impacts breast cancer cell migration and invasion by regulating Ezrin-radixin-moesin. Med. Sci. Monit. 2016, 22, 1946–1952. [Google Scholar] [CrossRef]
- Zhang, G.; Zhang, W.; Li, B.; Stringer-Reasor, E.; Chu, C.; Sun, L.; Bae, S.; Chen, D.; Wei, S.; Jiao, K.; et al. MicroRNA-200c and microRNA-141 are regulated by a FOXP3-KAT2B axis and associated with tumor metastasis in breast cancer. Breast Cancer Res. 2017, 19. [Google Scholar] [CrossRef]
- Wang, X.; Qiu, W.; Zhang, G.; Xu, S.; Gao, Q.; Yang, Z. MicroRNA-204 targets JAK2 in breast cancer and induces cell apoptosis through the STAT3/BCl-2/surviving pathway. Int. J. Clin. Exp. Pathol. 2015, 8, 5017–5025. [Google Scholar]
- Hong, B.S.; Ryu, H.S.; Kim, N.; Kim, J.; Lee, E.; Moon, H.; Kim, K.H.; Jin, M.S.; Kwon, N.H.; Kim, S.; et al. Tumor suppressor miRNA-204-5p regulates growth, metastasis, and immune microenvironment remodeling in breast cancer. Cancer Res. 2019, 79, 1520–1534. [Google Scholar] [CrossRef]
- Liu, J.; Mao, Q.; Liu, Y.; Hao, X.; Zhang, S.; Zhang, J. Analysis of miR-205 and miR-155 expression in the blood of breast cancer patients. Chin. J. Cancer Res. 2013, 2546–2554. [Google Scholar] [CrossRef]
- Elgamal, O.A.; Park, J.K.; Gusev, Y.; Azevedo-Pouly, A.C.P.; Jiang, J.; Roopra, A.; Schmittgen, T.D. Tumor suppressive function of mir-205 in breast cancer is linked to HMGB3 regulation. PLoS ONE 2013, 8. [Google Scholar] [CrossRef]
- Ren, Y.Q.; Wang, H.J.; Zhang, Y.Q.; Liu, Y.B. WBP2 modulates G1/S transition in ER+ breast cancer cells and is a direct target of miR-206. Cancer Chemother. Pharmacol. 2017, 79, 1003–1011. [Google Scholar] [CrossRef]
- Camps, C.; Saini, H.K.; Mole, D.R.; Choudhry, H.; Reczko, M.; Guerra-Assunção, J.A.; Tian, Y.M.; Buffa, F.M.; Harris, A.L.; Hatzigeorgiou, A.G.; et al. Integrated analysis of microRNA and mRNA expression and association with HIF binding reveals the complexity of microRNA expression regulation under hypoxia. Mol. Cancer 2014, 13. [Google Scholar] [CrossRef] [Green Version]
- Costales, M.G.; Haga, C.L.; Velagapudi, S.P.; Childs-Disney, J.L.; Phinney, D.G.; Disney, M.D. Small molecule inhibition of microRNA-210 reprograms an oncogenic hypoxic circuit. J. Am. Chem. Soc. 2017, 139, 3446–3455. [Google Scholar] [CrossRef] [Green Version]
- Chen, L.L.; Zhang, Z.J.; Yi, Z.B.; Li, J.J. MicroRNA-211-5p suppresses tumour cell proliferation, invasion, migration and metastasis in triple-negative breast cancer by directly targeting SETBP1. Br. J. Cancer 2017, 117, 78–88. [Google Scholar] [CrossRef]
- Santolla, M.F.; Lappano, R.; Cirillo, F.; Rigiracciolo, D.C.; Sebastiani, A.; Abonante, S.; Tassone, P.; Tagliaferri, P.; Di Martino, M.T.; Maggiolini, M.V.; et al. miR-221 stimulates breast cancer cells and cancer-associated fibroblasts (CAFs) through selective interference with the A20/c-Rel/CTGF signaling. J. Exp. Clin. Cancer Res. 2018, 37, 1–12. [Google Scholar] [CrossRef]
- Deng, L.; Lei, Q.; Wang, Y.; Wang, Z.; Xie, G.; Zhong, X.; Wang, Y.; Chen, N.; Qiu, Y.; Pu, T.; et al. Downregulation of miR-221-3p and upregulation of its target gene PARP1 are prognostic biomarkers for triple negative breast cancer patients and associated with poor prognosis. Oncotarget 2017, 8, 108712–108725. [Google Scholar] [CrossRef]
- Muraki, K.; Nyhan, K.; Han, L.; Murnane, J.P. Mechanisms of telomere loss and their consequences for chromosome instability. Front. Oncol. 2012, 2. [Google Scholar] [CrossRef] [Green Version]
- Dong, Y.; Liu, Y.; Jiang, A.; Li, R.; Yin, M.; Wang, Y. MicroRNA-335 suppresses the proliferation, migration, and invasion of breast cancer cells by targeting EphA4. Mol. Cell. Biochem. 2018, 439, 95–104. [Google Scholar] [CrossRef]
- Tavazoie, S.F.; Alarcón, C.; Oskarsson, T.; Padua, D.; Wang, Q.; Bos, P.D.; Gerald, W.L.; Massagué, J. Endogenous human microRNAs that suppress breast cancer metastasis. Nature 2008, 451, 147–152. [Google Scholar] [CrossRef] [Green Version]
- Zhang, S.; Kim, K.H.; Jin, U.H.; Pfent, C.; Cao, H.; Amendt, B.; Liu, X.; Wilson-Robles, H.; Safe, S. Aryl hydrocarbon receptor agonists induce microRNA-335 expression and inhibit lung metastasis of estrogen receptor negative breast cancer cells. Mol. Cancer Ther. 2012, 11, 108–118. [Google Scholar] [CrossRef] [Green Version]
- He, J.; Wang, J.; Li, S.; Li, T.; Chen, K.; Zhang, S. Hypoxia-inhibited miR-338-3p suppresses breast cancer progression by directly targeting ZEB2. Cancer Sci. 2020, 111, 3550–3563. [Google Scholar] [CrossRef]
- Jin, Y.; Zhao, M.; Xie, Q.; Zhang, H.; Wang, Q.; Ma, Q. MicroRNA-338-3p functions as tumor suppressor in breast cancer by targeting SOX4. Int. J. Oncol. 2015, 47, 1594–1602. [Google Scholar] [CrossRef]
- Jiang, Q.; He, M.; Ma, M.T.; Wu, H.Z.; Yu, Z.J.; Guan, S.; Jiang, L.Y.; Wang, Y.; Di Zheng, D.; Jin, F.; et al. MicroRNA-148a inhibits breast cancer migration and invasion by directly targeting WNT-1. Oncol. Rep. 2016, 35, 1425–1432. [Google Scholar] [CrossRef]
- Zhang, J.; Zhang, Z.; Wang, Q.; Xing, X.-J.; Zhao, Y. Overexpression of microRNA-365 inhibits breast cancer cell growth and chemo-resistance through GALNT4. Eur. Rev. Med. Pharmacol. Sci. 2016, 20, 4710–4718. [Google Scholar]
- Huang, Q.; Gumireddy, K.; Schrier, M.; le Sage, C.; Nagel, R.; Nair, S.; Egan, D.A.; Li, A.; Huang, G.; Klein-Szanto, A.J.; et al. The microRNAs miR-373 and miR-520c promote tumour invasion and metastasis. Nat. Cell Biol. 2008, 10, 202–210. [Google Scholar] [CrossRef]
- Cai, J.; Guan, H.; Fang, L.; Yang, Y.; Zhu, X.; Yuan, J.; Li, M. MicroRNA-374a activates Wnt/β-catenin signaling to promote breast cancer metastasis. J. Clin. Investig. 2013, 123, 566–579. [Google Scholar] [CrossRef] [Green Version]
- Pan, Y.; Jiao, G.; Wang, C.; Yang, J.; Yang, W. MicroRNA-421 inhibits breast cancer metastasis by targeting metastasis associated 1. Biomed. Pharmacother. 2016, 83, 1398–1406. [Google Scholar] [CrossRef]
- Xie, D.; Song, H.; Wu, T.; Li, D.; Hua, K.; Xu, H.; Zhao, B.; Wu, C.; Hu, J.; Ji, C.; et al. MicroRNA-424 serves an anti-oncogenic role by targeting cyclin-dependent kinase 1 in breast cancer cells. Oncol. Rep. 2018, 40, 3416–3426. [Google Scholar] [CrossRef]
- Wang, B.; Zou, A.; Ma, L.; Chen, X.; Wang, L.; Zeng, X.; Tan, T. miR-455 inhibits breast cancer cell proliferation through targeting CDK14. Eur. J. Pharmacol. 2017, 807, 138–143. [Google Scholar] [CrossRef]
- Huang, X.; Lyu, J. Tumor suppressor function of miR-483-3p on breast cancer via targeting of the cyclin E1 gene. Exp. Ther. Med. 2018, 16, 2615–2620. [Google Scholar] [CrossRef] [Green Version]
- Zhan, M.N.; Yu, X.T.; Tang, J.; Zhou, C.X.; Wang, C.L.; Yin, Q.Q.; Gong, X.F.; He, M.; He, J.R.; Chen, G.Q.; et al. MicroRNA-494 inhibits breast cancer progression by directly targeting PAK1. Cell Death Dis. 2017, 8. [Google Scholar] [CrossRef] [Green Version]
- Luo, Q.; Li, X.; Gao, Y.; Long, Y.; Chen, L.; Huang, Y.; Fang, L. MiRNA-497 regulates cell growth and invasion by targeting cyclin E1 in breast cancer. Cancer Cell Int. 2013, 13, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Liu, J.; Zhou, Y.; Shi, Z.; Hu, Y.; Meng, T.; Zhang, X.; Zhang, S.; Zhang, J. microRNA-497 modulates breast cancer cell proliferation, invasion, and survival by targeting SMAD7. DNA Cell Biol. 2016, 35, 521–529. [Google Scholar] [CrossRef]
- Wu, Z.; Cai, X.; Huang, C.; Xu, J.; Liu, A. MiR-497 suppresses angiogenesis in breast carcinoma by targeting HIF-1α. Oncol. Rep. 2016, 35, 1696–1702. [Google Scholar] [CrossRef]
- Breunig, C.; Pahl, J.; Küblbeck, M.; Miller, M.; Antonelli, D.; Erdem, N.; Wirth, C.; Will, R.; Bott, A.; Cerwenka, A.; et al. MicroRNA-519a-3p mediates apoptosis resistance in breast cancer cells and their escape from recognition by natural killer cells. Cell Death Dis. 2017, 8. [Google Scholar] [CrossRef] [PubMed]
- Chen, P.; Xu, W.; Luo, Y.; Zhang, Y.; He, Y.; Yang, S.; Yuan, Z. MicroRNA 543 suppresses breast cancer cell proliferation, blocks cell cycle and induces cell apoptosis via direct targeting of ERK/MAPK. Onco. Targets Ther. 2017, 10, 1423–1431. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Senthil Kumar, K.J.; Gokila Vani, M.; Hsieh, H.W.; Lin, C.C.; Liao, J.W.; Chueh, P.J.; Wang, S.Y. MicroRNA-708 activation by glucocorticoid receptor agonists regulate breast cancer tumorigenesis and metastasis via downregulation of NF-κB signalling. Carcinogenesis 2019, 40, 335–348. [Google Scholar] [CrossRef] [PubMed]
- Yan, C.; Chen, Y.; Kong, W.; Fu, L.; Liu, Y.; Yao, Q.; Yuan, Y. PVT1-derived miR-1207-5p promotes breast cancer cell growth by targeting STAT6. Cancer Sci. 2017, 108, 868–876. [Google Scholar] [CrossRef] [Green Version]
- Yan, L.X.; Huang, X.F.; Shao, Q.; Huang, M.Y.; Deng, L.; Wu, Q.L.; Zeng, Y.X.; Shao, J.Y. MicroRNA miRNA-21 overexpression in human breast cancer is associated with advanced clinical stage, lymph node metastasis and patient poor prognosis. RNA 2008, 14, 2348–2360. [Google Scholar] [CrossRef] [Green Version]
- Huang, G.L.; Zhang, X.H.; Guo, G.L.; Huang, K.T.; Yang, K.Y.; Shen, X.; You, J.; Hu, X.Q. Clinical significance of miR-21 expression in breast cancer: SYBR-Green I-based real-time RT-PCR study of invasive ductal carcinoma. Oncol. Rep. 2009, 21, 673–679. [Google Scholar]
- Zhu, S.; Si, M.L.; Wu, H.; Mo, Y.Y. MicroRNA-21 targets the tumor suppressor gene tropomyosin 1 (TPM1). J. Biol. Chem. 2007, 282, 14328–14336. [Google Scholar] [CrossRef] [Green Version]
- Frankel, L.B.; Christoffersen, N.R.; Jacobsen, A.; Lindow, M.; Krogh, A.; Lund, A.H. Programmed cell death 4 (PDCD4) is an important functional target of the microRNA miR-21 in breast cancer cells. J. Biol. Chem. 2008, 283, 1026–1033. [Google Scholar] [CrossRef] [Green Version]
- Iorio, M.V.; Casalini, P.; Piovan, C.; Di Leva, G.; Merlo, A.; Triulzi, T.; Ménard, S.; Croce, C.M.; Tagliabue, E. MicroRNA-205 regulates HER3 in human breast cancer. Cancer Res. 2009, 69, 2195–2200. [Google Scholar] [CrossRef] [Green Version]
- Di Leva, G.; Piovan, C.; Gasparini, P.; Ngankeu, A.; Taccioli, C.; Briskin, D.; Cheung, D.G.; Bolon, B.; Anderlucci, L.; Alder, H.; et al. Estrogen mediated-activation of miR-191/425 cluster modulates tumorigenicity of breast cancer cells depending on estrogen receptor status. PLoS Genet. 2013, 9, e1003311. [Google Scholar] [CrossRef]
- Miller, T.E.; Ghoshal, K.; Ramaswamy, B.; Roy, S.; Datta, J.; Shapiro, C.L.; Jacob, S.; Majumder, S. MicroRNA-221/222 confers tamoxifen resistance in breast cancer by targeting p27Kip1. J. Biol. Chem. 2008, 283, 29897–29903. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, J.J.; Lin, J.; Yang, H.; Kong, W.; He, L.; Ma, X.; Coppola, D.; Cheng, J.Q. MicroRNA-221/222 negatively regulates estrogen receptor alpha and is associated with tamoxifen resistance in breast cancer. J. Biol. Chem. 2008, 283, 31079–31086. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Klinge, C.M. miRNAs and estrogen action. Trends Endocrinol. Metab. 2012, 5, 223–233. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Klinge, C.M. miRNAs regulated by estrogens, tamoxifen, and endocrine disruptors and their downstream gene targets. Mol. Cell. Endocrinol. 2015, 418 Pt3, 273–297. [Google Scholar] [CrossRef] [Green Version]
- Tao, S.; He, H.; Chen, Q.; Yue, W. GPER mediated estradiol reduces miR-148a to promote HLA-G expression in breast cancer. Biochem. Biophys. Res. Commun. 2014, 451, 74–78. [Google Scholar] [CrossRef]
- Tao, S.; He, H.; Chen, Q. Estradiol induces HOTAIR levels via GPER-mediated miR-148a inhibition in breast cancer. J. Transl. Med. 2015, 13, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Ma, L. MicroRNA and Metastasis. Adv. Cancer Res. 2016, 132, 165–207. [Google Scholar] [CrossRef]
- Vivacqua, A. Personal Communication; University of Calabria: Rende, Italy, 2020. [Google Scholar]
Figure 1.
Schematic representation of miRNA synthesis, as explained in the text. Abbreviations: RNA Pol II or III: RNA Polimerase II or III; Drosha: DGCR8: DiGeorge syndrome critical region gene 8; XOP5: Exportin 5; RISC complex: RNA-induced silencing complex.
Figure 1.
Schematic representation of miRNA synthesis, as explained in the text. Abbreviations: RNA Pol II or III: RNA Polimerase II or III; Drosha: DGCR8: DiGeorge syndrome critical region gene 8; XOP5: Exportin 5; RISC complex: RNA-induced silencing complex.

Figure 2.
Schematic representation of GPER1 involvement in the regulation of miRNA expression. Ligand-binding to GPER1 leads to activation of SRC through the β and γ subunits of the G protein. The complex SRC tyrosine kinase-SHC adapter protein triggers the EGFR transactivation, which, in turn, stimulates downstream pathways, like PI3K, MAPKs and PKCδ. In addition, GPER activation stimulates the activity of AC through the Gα subunit, leading to the PKA-mediated increase of CREB. Both signalling may trigger the expression of specific TFs involved in miRNA transcription. In the nucleus miRNAs are processed and exported in cytoplasm, where they become mature miRNAs. SRC: steroid receptor coactivator; EGFR: epidermal growth factor receptor; PI3K: phosphatidylinositol 3-kinase; PKCδ: phospho-kinase C delta; AC: adenylyl cyclase; PKA: phospho-kinase A; CREB: cAMP response element-binding protein; TFs: transcription factors. Further abbreviations are addressed in the text.
Figure 2.
Schematic representation of GPER1 involvement in the regulation of miRNA expression. Ligand-binding to GPER1 leads to activation of SRC through the β and γ subunits of the G protein. The complex SRC tyrosine kinase-SHC adapter protein triggers the EGFR transactivation, which, in turn, stimulates downstream pathways, like PI3K, MAPKs and PKCδ. In addition, GPER activation stimulates the activity of AC through the Gα subunit, leading to the PKA-mediated increase of CREB. Both signalling may trigger the expression of specific TFs involved in miRNA transcription. In the nucleus miRNAs are processed and exported in cytoplasm, where they become mature miRNAs. SRC: steroid receptor coactivator; EGFR: epidermal growth factor receptor; PI3K: phosphatidylinositol 3-kinase; PKCδ: phospho-kinase C delta; AC: adenylyl cyclase; PKA: phospho-kinase A; CREB: cAMP response element-binding protein; TFs: transcription factors. Further abbreviations are addressed in the text.


{kind=link}
{kind=link}
Table 1.
Main miRNAs modulated in breast cancers.
| miRNA Name | Expression | Main Targets | Action | References |
|---|---|---|---|---|
| Let-7 | Downregulated | H-Ras, HMGA2, LIN28, PEBP1 | Tumor suppressor | [96,97,98,99] |
| miR-10b | Upregulated | HOXD10 | Oncogenic miRNA | [96,100] |
| miR-15a and | Downregulated | Cyclin E1, E2F7 | Tumor suppressor | [101,102,103] |
| miR-16 | ||||
| miR17-5p | Downregulated | A1B1 gene, Cyclin D1, E2F1 | Tumor suppressor | [96,104,105] |
| miR-21 | Upregulated | Akt, BCL-2, BAX | Oncogenic miRNA | [101,106] |
| miR-22 | Downregulated | KRAS | Tumor suppressor | [101,107] |
| miR-24 | Upregulated | Oct-3/4, HIF-1α, Snail, VEGFA | Oncogenic miRNA | [101,108] |
| miR-26a | Downregulated | Cyclin D1, CDK1, CDK6, p21, p27, p53, MYC, RFP6/ERα/BCL6-Xl, E2F7, Cyclin E2 | Tumor suppressor | [101,109,110,111] |
| miR-27a | Upregulated | Zinc Finger ZBTB10, MYT-1 | Oncogenic miRNA | [96,112] |
| mir-29b | Downregulated | Akt3, VEGF, c-MYC | Tumor suppressor | [101,113] |
| miR-30b | Downregulated | Cyclin E2 | Tumor suppressor | [101,111] |
| miR-30c-2–3p | Downregulated | Cyclin E1 | Tumor suppressor | [101,114] |
| miR-31 | Downregulated | WAVE3, RhoA | Tumor suppressor | [115,116] |
| miR-33b | Downregulated | HMGA2, SALL4, Twist 1 | Tumor suppressor | [101,117] |
| miR-34a | Downregulated | Tumor protein D52 gene, p53, E-cadherin, N-cadherin, TGFβ | Tumor suppressor | [101,118] |
| miR-100 | Downregulated | VEFG, mTOR/HIF-1α | Tumor suppressor | [101,119] |
| miR-101 | Downregulated | EYA1, jagged1, Hes1, Hey1, SOX2 | Tumor suppressor | [101,120,121] |
| miR-122 | Upregulated | Pyruvate Kinase, Citrate Synthase | Tumor suppressor | [101,122] |
| miR-124 | Downregulated | STAT3 | Tumor suppressor | [101,123] |
| miR-124a and miR-26b | Downregulated | SerpinB2 | Tumor suppressor | [101,124] |
| miR-125b | Downregulated | Eritropoietin receptor, ERBB2, ENPEP, casein kinase 2α, Cyclin J | Tumor suppressor | [116,122,125] |
| miR-126 | Downregulated | VEFGA, IGFBP2, MERTK, PITPNC1 | Tumor suppressor | [101,126] |
| miR-133a | Downregulated | LASP1 | Tumor suppressor | [101,127] |
| miR-135b | Upregulated | LATS2, CDK2, p-YAP | Oncogenic miRNA | [101,128] |
| miR-138 | Downregulated | E-cadherin, vimentin, N-cadherin, Snail | Tumor suppressor | [101,129] |
| miR-140-5p | Downregulated | Cluster of differentiation 31, MMP9, Ki-67, VEGFA, | Tumor suppressor | [101,130] |
| miR-143 | Downregulated | ERK5, MAP3K7, Cyclin D1 | Tumor suppressor | [101,131] |
| miR-144 | Upregulated | Runx-1 | Oncogenic miRNA | [9] |
| miR-145 | Downregulated | p53-mediated repression of c-myc | Tumor suppressor | [96,122,132] |
| miR-146 | Downregulated | NF-kB | Tumor suppressor | [96,133] |
| miR-148a | Downregulated | WNT-1, β-catenin, MMP7, TF4, B-cell lymphoma-2, caspase | Tumor suppressor | [134] |
| miR-155 | Upregulated | TRF1 | Oncogenic miRNA | [122,135,136] |
| miR-191 | Upregulated | HuR, TGFβ2, SMAD3, BMP4, JUN, FOS, PTGS2, CTGF, VEGFA | Oncogenic miRNA | [101,137] |
| miR-191-5p | Upregulated | SOX4, caspase-3, caspase-7, p53 | Oncogenic miRNA | [101,138] |
| miR-193a | Downregulated | WT1 | Tumor suppressor | [101,139] |
| miR-195 | Downregulated | FASN, HMGCR, ACACA, CYP27B1 | Tumor suppressor | [101,140,141] |
| miR-200b | Upregulated | Ezrin/Radixin/Moesin (ERM) | Oncogenic miRNA | [101,142] |
| miR-200c and miR-141 | Upregulated | SerpinB2, c-Jun, c-FosB, FOXP3, KAT2B | Oncogenic miRNA | [101,124,143] |
| miR-203 | Upregulated | PKC-ERK, SOCS3 | Oncogenic miRNA | [101,106] |
| miR-204 | Downregulated | JAK2, BCL-2, Survivin | Tumor suppressor | [101,144] |
| miR-204-5p | Downregulated | PIK3CB | Tumor suppressor | [101,145] |
| miR-205 | Downregulated | HMGB3 | Tumor suppressor | [116,146,147] |
| miR-206 | Downregulated | WBP2, p-21, CDK4, Cyclin D1 | Tumor suppressor | [101,148] |
| miR-210 | Upregulated | HIFs, GPD1L, Pax-5 | Oncogenic miRNA | [101,149,150] |
| miR-211-5p | Downregulated | SETBP1 | Tumor suppressor | [101,151] |
| miR-221 | Upregulated | A20, PARP1 | Oncogenic miRNA | [152,153] |
| miR-296-5p and miR-512-5p | Downregulated | hTERT | Tumor suppressor | [101,154] |
| miR-331 | Upregulated | HER2, HOTAIR, E2F1, DOHH, PHLPP | Oncogenic miRNA | [101,141] |
| miR-335 | Downregulated | SOX4, TNC | Tumor suppressor | [101,155,156,157,158] |
| miR-338-3p | Downregulated | c-FOS, ZEB2, SOX4 | Tumor suppressor | [8,158,159] |
| miR-340 | Downregulated | Rho associated coleid-coil containing protein kinase 1, CTNNB1, c-MYC | Tumor suppressor | [101,160] |
| miR-365 | Downregulated | GALNT4 | Tumor suppressor | [101,161] |
| miR-373/520c | Upregulated | CD44 | Oncogenic miRNA | [101,162] |
| miR-374a | Upregulated | e-cadherin, γ-catenin, CK18, vimentin, N-cadherin, β-catenin, WIF1, PTEN, WNT5A | Oncogenic miRNA | [101,163] |
| miR-421 | Downregulated | MTA1 | Tumor suppressor | [101,164] |
| miR-424 | Downregulated | CDK1, YAP, ERK1/2 | Tumor suppressor | [101,165] |
| miR-455 | Downregulated | CDK14, Cyclin D1, p21 | Tumor suppressor | [101,166] |
| miR-483-3p | Downregulated | Cyclin W1, p-NPAT, CDK21 | Tumor suppressor | [101,167] |
| miR-494p | Downregulated | PAK1, E-cadherin | Tumor suppressor | [101,168] |
| miR-497 | Downregulated | Cyclin E1, SMAD7, CD274, VEGF, HIF-1α, Cyclin E1, E2F7 | Tumor suppressor | [101,141,169,170,171] |
| miR-519a-3p | Upregulated | TRAIL-R2, caspase-8, caspase 7, MICA, ULBP2 | Oncogenic miRNA | [101,172] |
| miR-543 | Downregulated | ERK/MAPK | Tumor suppressor | [101,173] |
| miR-708 | Downregulated | INF-kB subunit β, COX2, c-MYC | Tumor suppressor | [101,174] |
| miR-1207-5p | Upregulated | STAT2, CDKN, CDK1B | Oncogenic miRNA | [101,175] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Vivacqua, A. GPER1 and microRNA: Two Players in Breast Cancer Progression. Int. J. Mol. Sci. 2021, 22, 98. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22010098
AMA Style
Vivacqua A. GPER1 and microRNA: Two Players in Breast Cancer Progression. International Journal of Molecular Sciences. 2021; 22(1):98. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22010098
Chicago/Turabian StyleVivacqua, Adele. 2021. "GPER1 and microRNA: Two Players in Breast Cancer Progression" International Journal of Molecular Sciences 22, no. 1: 98. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22010098
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.