
Human Indoleamine 2,3-dioxygenase 1 (IDO1) Expressed in Plant Cells Induces Kynurenine Production

 , , , , , , and
, , , , , , and 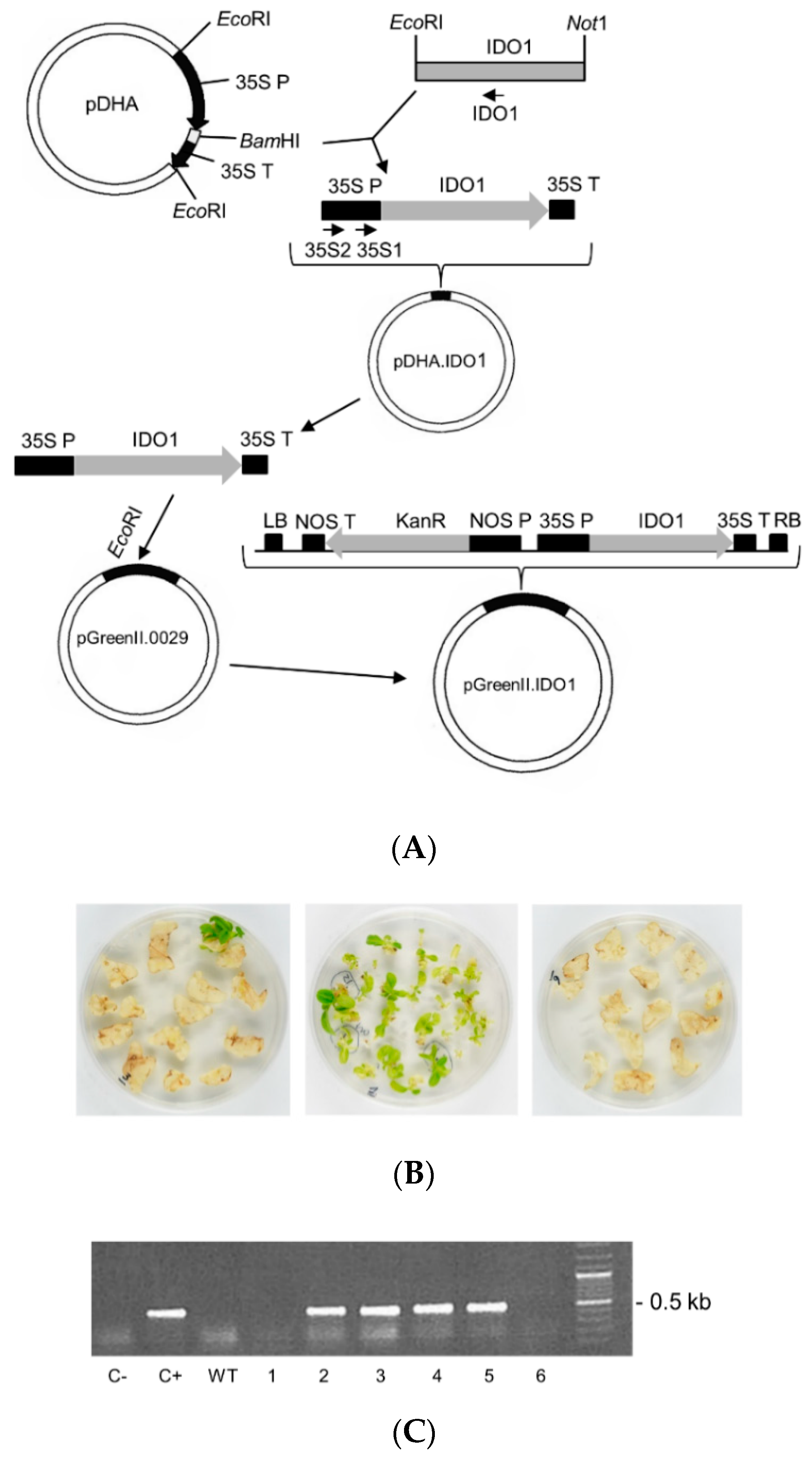{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Cloning of IDO1 cDNA in a Plant Expression Vector and Attempted Stably Expression in Tobacco Plants
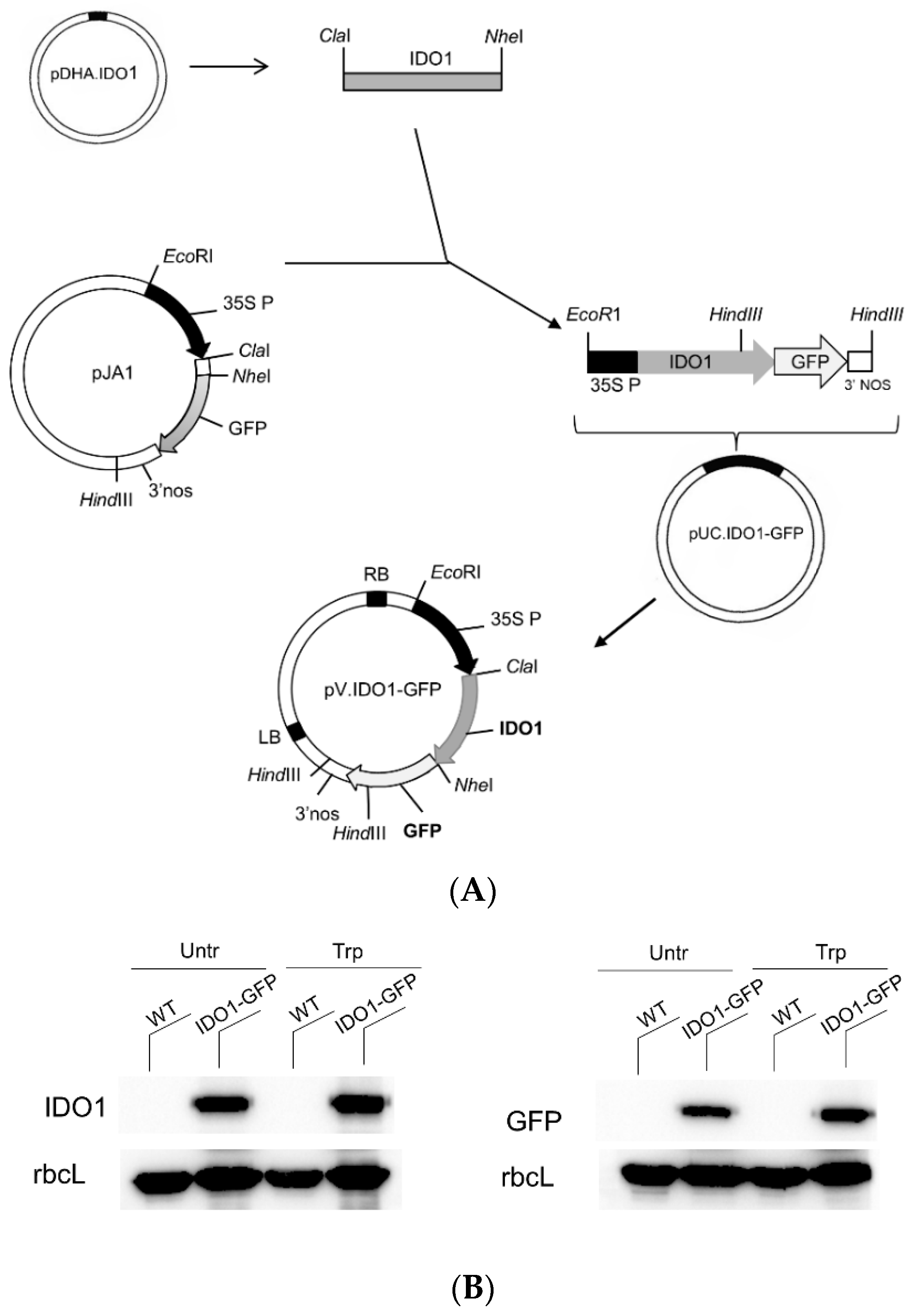
2.2. Human IDO1-GFP Can Be Transiently Expressed in Tobacco Protoplasts
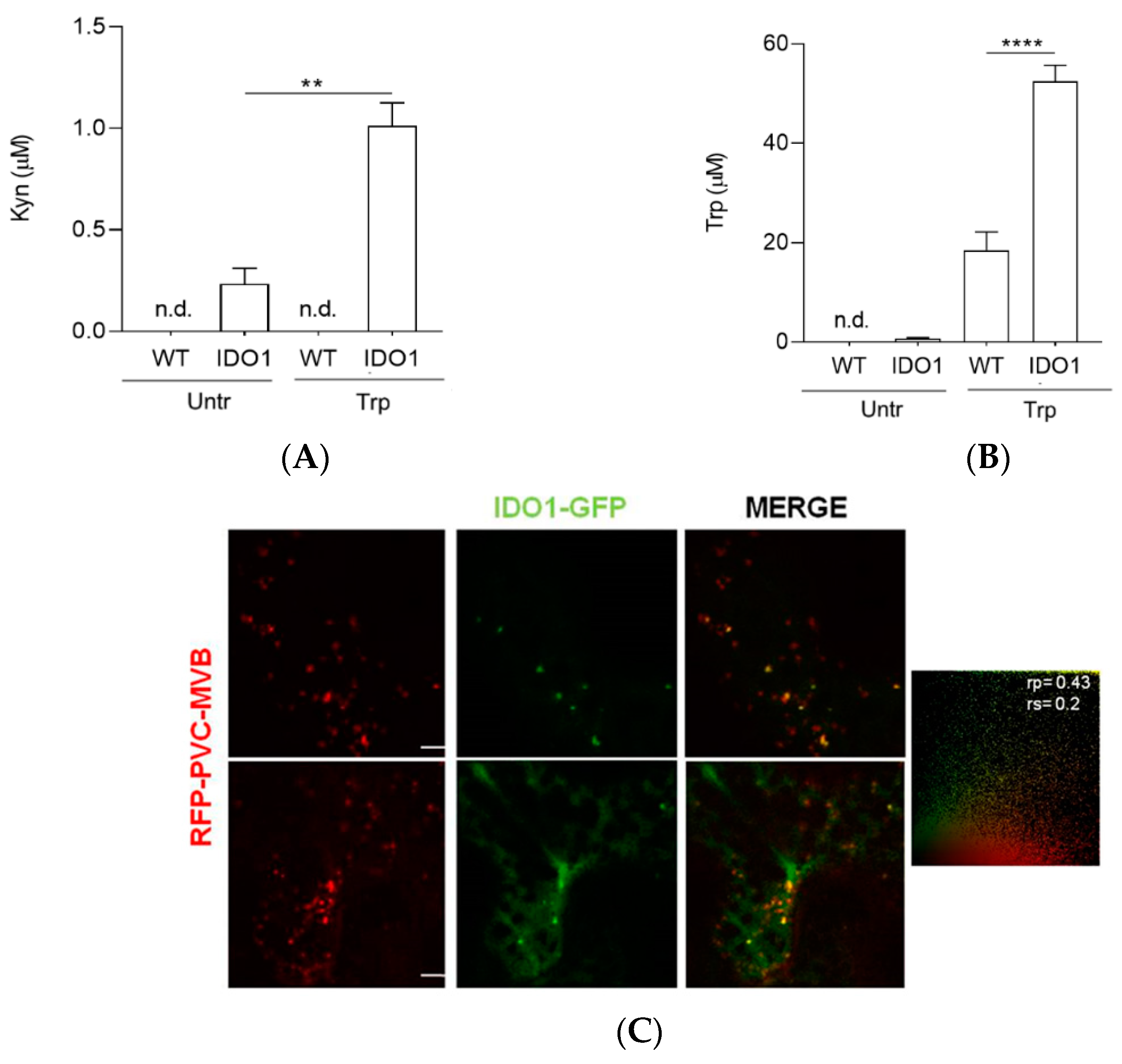
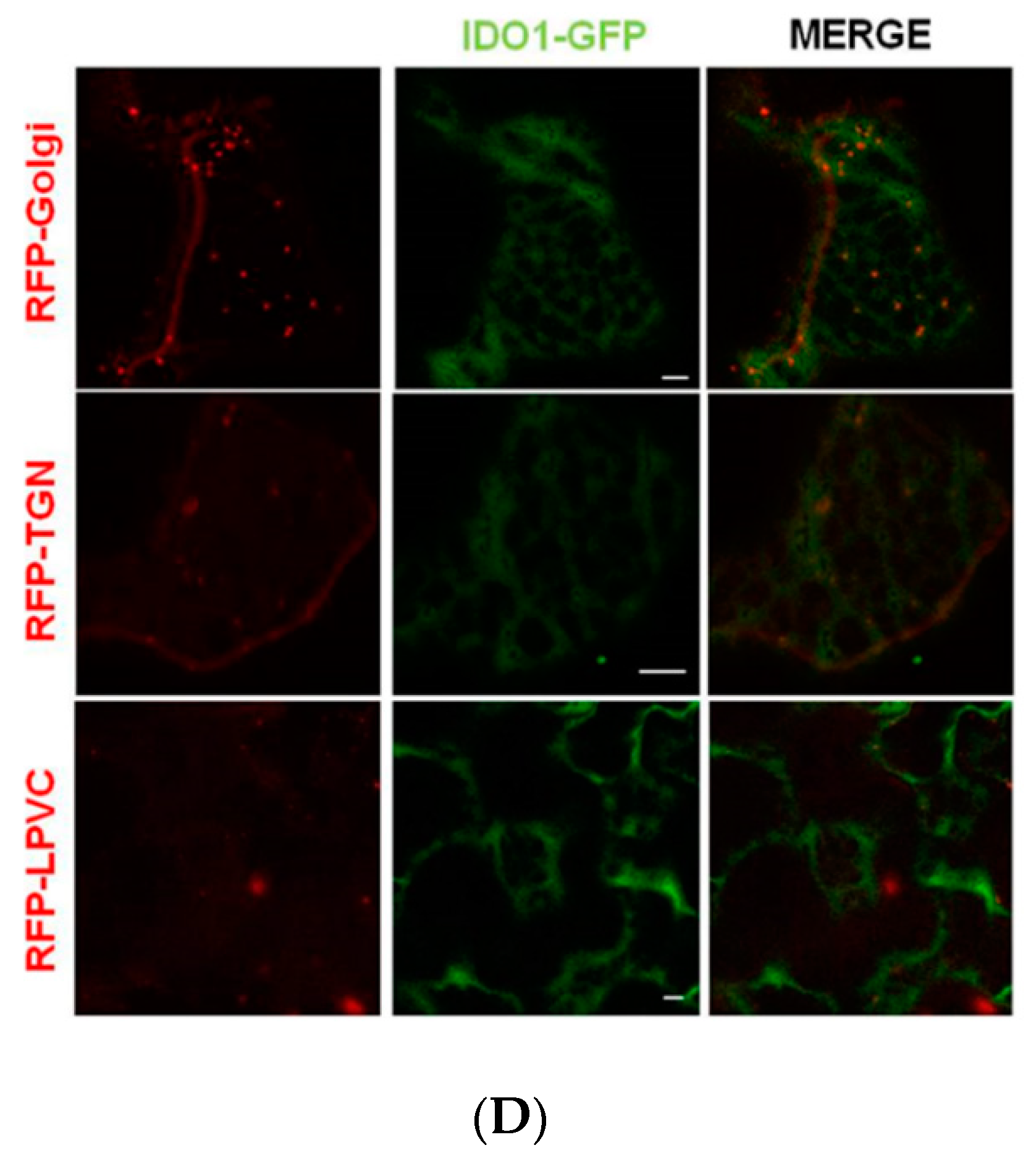
2.3. Transiently Expressed IDO1-GFP Had Catalytic Activity and Similar Subcellular Localisation of Mammal IDO1
3. Discussion
4. Materials and Methods
4.1. Construction of Plasmids for Plant Transformation
4.2. Tobacco Protoplast Isolation
4.3. Tobacco Protoplast Transformation
4.4. Plant Transformation by Agroinfiltration and Microscopy Analysis
4.5. DNA Isolation and Analysis
4.6. RNA Extraction and RT-PCR Analysis
4.7. Protein Extraction from Protoplasts and Western Blotting Analysis
4.8. Kynurenine Analysis
4.9. Statistics
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Hiatt, A.; Caffferkey, R.; Bowdish, K. Production of antibodies in transgenic plants. Nat. Cell Biol. 1989, 342, 76–78. [Google Scholar] [CrossRef]
- Fischer, R.; Drossard, J.; Commandeur, U.; Schillberg, S.; Emans, N. Towards molecular farming in the future: Moving from diagnostic protein and antibody production in microbes to plants. Biotechnol. Appl. Biochem. 1999, 30, 101–108. [Google Scholar]
- Schillberg, S.; Raven, N.; Spiegel, H.; Rasche, S.; Buntru, M. Critical Analysis of the Commercial Potential of Plants for the Production of Recombinant Proteins. Front. Plant. Sci. 2019, 10, 720. [Google Scholar] [CrossRef] [PubMed]
- de Virgilio, M.; de Marchis, F.; Bellucci, M.; Mainieri, D.; Rossi, M.; Benvenuto, E.; Arcioni, S.; Vitale, A. The human immunodeficiency virus antigen Nef forms protein bodies in leaves of transgenic tobacco when fused to zeolin. J. Exp. Bot. 2008, 59, 2815–2829. [Google Scholar] [CrossRef] [PubMed]
- Bock, R. Engineering chloroplasts for high-level foreign protein expression. Methods Mol. Biol. 2014, 1132, 93–106. [Google Scholar] [PubMed]
- Krenek, P.; Samajova, O.; Luptovciak, I.; Doskocilova, A.; Komis, G.; Samaj, J. Transient plant transformation mediated by Agrobacterium tumefaciens: Principles, methods and applications. Biotechnol. Adv. 2015, 33, 1024–1042. [Google Scholar] [CrossRef] [PubMed]
- Hellens, R.P.; Edwards, E.A.; Leyland, N.R.; Bean, S.; Mullineaux, P.M. pGreen: A versatile and flexible binary Ti vector for Agrobacterium-mediated plant transformation. Plant. Mol. Biol. 2000, 42, 819–832. [Google Scholar] [CrossRef]
- Marillonnet, S.; Thoeringer, C.; Kandzia, R.; Klimyuk, V.; Gleba, Y. Systemic Agrobacterium tumefaciens–mediated transfection of viral replicons for efficient transient expression in plants. Nat. Biotechnol. 2005, 23, 718–723. [Google Scholar] [CrossRef] [PubMed]
- Craig, W.; Gargano, D.; Scotti, N.; Nguyen, T.T.; Lao, N.T.; Kavanagh, T.A.; Dix, P.J.; Cardi, T. Direct gene transfer in potato: A comparison of particle bombardment of leaf explants and PEG-mediated transformation of protoplasts. Plant. Cell Rep. 2005, 24, 603–611. [Google Scholar] [CrossRef]
- Wang, P.; Zhao, F.-J.; Kopittke, P.M. Engineering Crops without Genome Integration Using Nanotechnology. Trends Plant. Sci. 2019, 24, 574–577. [Google Scholar] [CrossRef]
- De Marchis, F.; Balducci, C.; Pompa, A.; Stensland, H.M.F.R.; Guaragno, M.; Pagiotti, R.; Menghini, A.R.; Persichetti, E.; Beccari, T.; Bellucci, M. Human α-mannosidase produced in transgenic tobacco plants is processed in human α-mannosidosis cell lines. Plant. Biotechnol. J. 2011, 9, 1061–1073. [Google Scholar] [CrossRef] [PubMed]
- Shaaltiel, Y.; Bartfeld, D.; Hashmueli, S.; Baum, G.; Brill-Almon, E.; Galili, G.; Dym, O.; Boldin-Adamsky, S.A.; Silman, I.; Sussman, J.L.; et al. Production of glucocerebrosidase with terminal mannose glycans for enzyme replacement therapy of Gaucher’s disease using a plant cell system. Plant Biotechnol. J. 2007, 5, 579–590. [Google Scholar] [CrossRef]
- Mor, T.S. Molecular pharming’s foot in the FDA’s door: Protalix’s trailblazing story. Biotechnol. Lett. 2015, 37, 2147–2150. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schwarcz, R. The kynurenine pathway of tryptophan degradation as a drug target. Curr. Opin. Pharm. 2004, 4, 12–17. [Google Scholar] [CrossRef] [PubMed]
- Munn, D.H.; Zhou, M.; Attwood, J.T.; Bondarev, I.; Conway, S.J.; Marshall, B.; Brown, C.; Mellor, A.L. Prevention of Allogeneic Fetal Rejection by Tryptophan Catabolism. Sciences 1998, 281, 1191–1193. [Google Scholar] [CrossRef]
- Orabona, C.; Mondanelli, G.; Pallotta, M.T.; Carvalho, A.; Albini, E.; Fallarino, F.; Vacca, C.; Volpi, C.; Belladonna, M.L.; Berioli, M.G.; et al. Deficiency of immunoregulatory indoleamine 2,3-dioxygenase 1in juvenile diabetes. JCI Insight 2018, 3. [Google Scholar] [CrossRef]
- Panfili, E.; Gerli, R.; Grohmann, U.; Pallotta, M.T. Amino Acid Metabolism in Rheumatoid Arthritis: Friend or Foe? Biomolecules 2020, 10, 1280. [Google Scholar] [CrossRef]
- Romani, L.; Fallarino, F.; De Luca, A.; Montagnoli, C.; D’Angelo, C.; Zelante, T.; Vacca, C.; Bistoni, F.; Fioretti, M.C.; Grohmann, U.; et al. Defective tryptophan catabolism underlies inflammation in mouse chronic granulomatous disease. Nat. Cell Biol. 2008, 451, 211–215. [Google Scholar] [CrossRef]
- Bock, F.; Rössner, S.; Onderka, J.; Lechmann, M.; Pallotta, M.T.; Fallarino, F.; Boon, L.; Nicolette, C.; Debenedette, M.A.; Tcherepanova, I.Y.; et al. Topical Application of Soluble CD83 Induces IDO-Mediated Immune Modulation, Increases Foxp3+ T Cells, and Prolongs Allogeneic Corneal Graft Survival. J. Immunol. 2013, 191, 1965–1975. [Google Scholar] [CrossRef] [Green Version]
- Mondanelli, G.; Ugel, S.; Grohmann, U.; Bronte, V. The immune regulation in cancer by the amino acid metabolizing enzymes ARG and IDO. Curr. Opin. Pharm. 2017, 35, 30–39. [Google Scholar] [CrossRef]
- Bessede, A.; Gargaro, M.; Pallotta, M.T.; Matino, D.; Servillo, G.; Brunacci, C.; Bicciato, S.; Mazza, E.M.C.; Macchiarulo, A.; Vacca, C.; et al. Aryl hydrocarbon receptor control of a disease tolerance defence pathway. Nature 2014, 511, 184–190. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grohmann, U.; Fallarino, F.; Puccetti, P. Tolerance, DCs and tryptophan: Much ado about IDO. Trends Immunol. 2003, 24, 242–248. [Google Scholar] [CrossRef]
- Pallotta, M.T.; Orabona, C.; Volpi, C.; Vacca, C.; Belladonna, M.L.L.; Bianchi, R.; Servillo, G.; Brunacci, C.; Calvitti, M.; Bicciato, S.; et al. Indoleamine 2,3-dioxygenase is a signaling protein in long-term tolerance by dendritic cells. Nat. Immunol. 2011, 12, 870–878. [Google Scholar] [CrossRef] [Green Version]
- Iacono, A.; Pompa, A.; De Marchis, F.; Panfili, E.; Greco, F.A.; Coletti, A.; Orabona, C.; Volpi, C.; Belladonna, M.L.; Mondanelli, G.; et al. Class IA PI3Ks regulate subcellular and functional dynamics of IDO1. EMBO Rep. 2020, 21, e49756. [Google Scholar] [CrossRef] [PubMed]
- Acuña-Castroviejo, D.; Tapias, V.; López, L.C.; Doerrier, C.; Camacho, E.; Carrión, M.D.; Mora, F.; Espinosa, A.; Escames, G. Protective effects of synthetic kynurenines on 1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine-induced parkinsonism in mice. Brain Res. Bull. 2011, 85, 133–140. [Google Scholar] [CrossRef] [PubMed]
- Wu, H.-Q.; Lee, S.-C.; Schwarcz, R. Systemic administration of 4-chlorokynurenine prevents quinolinate neurotoxicity in the rat hippocampus. Eur. J. Pharm. 2000, 390, 267–274. [Google Scholar] [CrossRef]
- Gellért, L.; Fuzik, J.; Göblös, A.; Sárközi, K.; Marosi, M.; Kis, Z.; Farkas, T.; Szatmari, I.; Fülöp, F.; Vécsei, L.; et al. Neuroprotection with a new kynurenic acid analog in the four-vessel occlusion model of ischemia. Eur. J. Pharm. 2011, 667, 182–187. [Google Scholar] [CrossRef] [PubMed]
- Zanos, P.; Piantadosi, S.C.; Wu, H.-Q.; Pribut, H.J.; Dell, M.J.; Can, A.; Snodgrass, H.R.; Zarate, C.A.; Schwarcz, R.; Gould, T.D. The Prodrug 4-Chlorokynurenine Causes Ketamine-Like Antidepressant Effects, but Not Side Effects, by NMDA/GlycineB-Site Inhibition. J. Pharm. Exp. Ther. 2015, 355, 76–85. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jones, S.P.; Guillemin, G.J.; Brew, B.J. The Kynurenine Pathway in Stem Cell Biology. Int. J. Tryptophan Res. 2013, 6, 57–66. [Google Scholar] [CrossRef] [PubMed]
- Sanchez, R. MODBASE, a database of annotated comparative protein structure models. Nucleic Acids Res. 2000, 28, 250–253. [Google Scholar] [CrossRef] [Green Version]
- Yang, H.; Liu, L.; Xu, F. The promises and challenges of fusion constructs in protein biochemistry and enzymology. Appl. Microbiol. Biotechnol. 2016, 100, 8273–8281. [Google Scholar] [CrossRef]
- Stoger, E.; Fischer, R.; Moloney, M.; Ma, J.K.-C. Plant Molecular Pharming for the Treatment of Chronic and Infectious Diseases. Annu. Rev. Plant. Biol. 2014, 65, 743–768. [Google Scholar] [CrossRef] [PubMed]
- Fischer, R.; Buyel, J.F. Molecular farming—The slope of enlightenment. Biotechnol. Adv. 2020, 40, 107519. [Google Scholar] [CrossRef] [PubMed]
- Tabata, H. Paclitaxel Production by Plant-Cell-Culture Technology. Adv. Biochem. Eng. Biotechnol. 2004, 87, 1–23. [Google Scholar]
- Georgiev, M.I.; Eibl, R.; Zhong, J.-J. Hosting the plant cells in vitro: Recent trends in bioreactors. Appl. Microbiol. Biotechnol. 2013, 97, 3787–3800. [Google Scholar] [CrossRef]
- Huang, T.-K.; McDonald, K.A. Bioreactor systems for in vitro production of foreign proteins using plant cell cultures. Biotechnol. Adv. 2012, 30, 398–409. [Google Scholar] [CrossRef] [PubMed]
- Capell, T.; Twyman, R.M.; Armario-Najera, V.; Ma, J.K.-C.; Schillberg, S.; Christou, P. Potential Applications of Plant Biotechnology against SARS-CoV-2. Trends Plant Sci. 2020, 25, 635–643. [Google Scholar] [CrossRef]
- Ma, J.K.-C.; Drake, P.M.W.; Christou, P. The production of recombinant pharmaceutical proteins in plants. Nat. Rev. Genet. 2003, 4, 794–805. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, N.T.; Enakahama, T.; Le, D.H.; Van Son, L.; Chu, H.H.; Ekishimoto, T. Aryl Hydrocarbon Receptor and Kynurenine: Recent Advances in Autoimmune Disease Research. Front. Immunol. 2014, 5, 551. [Google Scholar] [CrossRef] [Green Version]
- Tzin, V.; Galili, G. The Biosynthetic Pathways for Shikimate and Aromatic Amino Acids inArabidopsis thaliana. Arab. Book 2010, 8, e0132. [Google Scholar] [CrossRef] [Green Version]
- Katoh, A.; Hashimoto, T. Molecular biology of pyridine nucleotide and nicotine biosynthesis. Front. Biosci. 2004, 9, 1577–1586. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Okazaki, M.; Higuchi, K.; Aouini, A.; Ezura, H. Lowering intercellular melatonin levels by transgenic analysis of indoleamine 2,3-dioxygenase from rice in tomato plants. J. Pineal Res. 2010, 49, 239–247. [Google Scholar] [CrossRef]
- He, W.; Brumos, J.; Li, H.; Ji, Y.; Ke, M.; Gong, X.; Zeng, Q.; Li, W.; Zhang, X.; An, F.; et al. A Small-Molecule Screen Identifies l-Kynurenine as a Competitive Inhibitor of TAA1/TAR Activity in Ethylene-Directed Auxin Biosynthesis and Root Growth in Arabidopsis. Plant. Cell 2011, 23, 3944–3960. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Steenbergen, L.; Jongkees, B.J.; Sellaro, R.; Colzato, L.S. Tryptophan supplementation modulates social behavior: A review. Neurosci. Biobehav. Rev. 2016, 64, 346–358. [Google Scholar] [CrossRef] [PubMed]
- Nikolaus, S.; Schulte, B.; Al-Massad, N.; Thieme, F.; Schulte, D.M.; Bethge, J.; Rehman, A.; Tran, F.; Aden, K.; Häsler, R.; et al. Increased Tryptophan Metabolism Is Associated With Activity of Inflammatory Bowel Diseases. Gastroenterology 2017, 153, 1504–1516.e2. [Google Scholar] [CrossRef] [Green Version]
- Krishnan, S.; Ding, Y.; Saedi, N.; Choi, M.; Sridharan, G.V.; Sherr, D.H.; Yarmush, M.L.; Alaniz, R.C.; Jayaraman, A.; Lee, K. Gut Microbiota-Derived Tryptophan Metabolites Modulate Inflammatory Response in Hepatocytes and Macrophages. Cell Rep. 2018, 23, 1099–1111. [Google Scholar] [CrossRef]
- Tabe, L.M.; Wardley-Richardson, T.; Ceriotti, A.; Aryan, A.; McNabb, W.; Moore, A.; Higgins, T.J. A biotechnological approach to improving the nutritive value of alfalfa. J. Anim. Sci. 1995, 73, 2752–2759. [Google Scholar] [CrossRef]
- Pompa, A.; De Marchis, F.; Vitale, A.; Arcioni, S.; Bellucci, M. An engineered C-terminal disulfide bond can partially replace the phaseolin vacuolar sorting signal. Plant. J. 2010, 61, 782–791. [Google Scholar] [CrossRef] [PubMed]
- Foresti, O.; Gershlick, D.C.; Bottanelli, F.; Hummel, E.; Hawes, C.; Denecke, J. A Recycling-Defective Vacuolar Sorting Receptor Reveals an Intermediate Compartment Situated between Prevacuoles and Vacuoles in Tobacco. Plant. Cell 2010, 22, 3992–4008. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Marchis, F.; Pompa, A.; Bellucci, M. Chemical Secretory Pathway Modulation in Plant Protoplasts. Adv. Struct. Saf. Stud. 2016, 1459, 67–79. [Google Scholar] [CrossRef]
- Maliga, P.; Sz-Breznovits, Á.; Márton, L. Streptomycin-resistant Plants from Callus Culture of Haploid Tobacco. Nat. New Biol. 1973, 244, 29–30. [Google Scholar] [CrossRef] [PubMed]
- Sparkes, I.A.; Runions, J.; Kearns, A.; Hawes, C. Rapid, transient expression of fluorescent fusion proteins in tobacco plants and generation of stably transformed plants. Nat. Protoc. 2006, 1, 2019–2025. [Google Scholar] [CrossRef] [PubMed]
- Gershlick, D.C.; Lousa, C.D.M.; Foresti, O.; Lee, A.J.; Pereira, E.A.; DaSilva, L.L.; Bottanelli, F.; Denecke, J. Golgi-Dependent Transport of Vacuolar Sorting Receptors is Regulated by COPII, AP1, and AP4 Protein Complexes in Tobacco. Plant. Cell 2014, 26, 1308–1329. [Google Scholar] [CrossRef] [PubMed] [Green Version]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bellucci, M.; Pompa, A.; De Marcos Lousa, C.; Panfili, E.; Orecchini, E.; Maricchiolo, E.; Fraternale, D.; Orabona, C.; De Marchis, F.; Pallotta, M.T. Human Indoleamine 2,3-dioxygenase 1 (IDO1) Expressed in Plant Cells Induces Kynurenine Production. Int. J. Mol. Sci. 2021, 22, 5102. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22105102
Bellucci M, Pompa A, De Marcos Lousa C, Panfili E, Orecchini E, Maricchiolo E, Fraternale D, Orabona C, De Marchis F, Pallotta MT. Human Indoleamine 2,3-dioxygenase 1 (IDO1) Expressed in Plant Cells Induces Kynurenine Production. International Journal of Molecular Sciences. 2021; 22(10):5102. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22105102
Chicago/Turabian StyleBellucci, Michele, Andrea Pompa, Carine De Marcos Lousa, Eleonora Panfili, Elena Orecchini, Elisa Maricchiolo, Daniele Fraternale, Ciriana Orabona, Francesca De Marchis, and Maria Teresa Pallotta. 2021. "Human Indoleamine 2,3-dioxygenase 1 (IDO1) Expressed in Plant Cells Induces Kynurenine Production" International Journal of Molecular Sciences 22, no. 10: 5102. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22105102