
Morphological and Metabolite Responses of Potatoes under Various Phosphorus Levels and Their Amelioration by Plant Growth-Promoting Rhizobacteria

 , , , and
, , , and
Abstract
:1. Introduction
2. Results
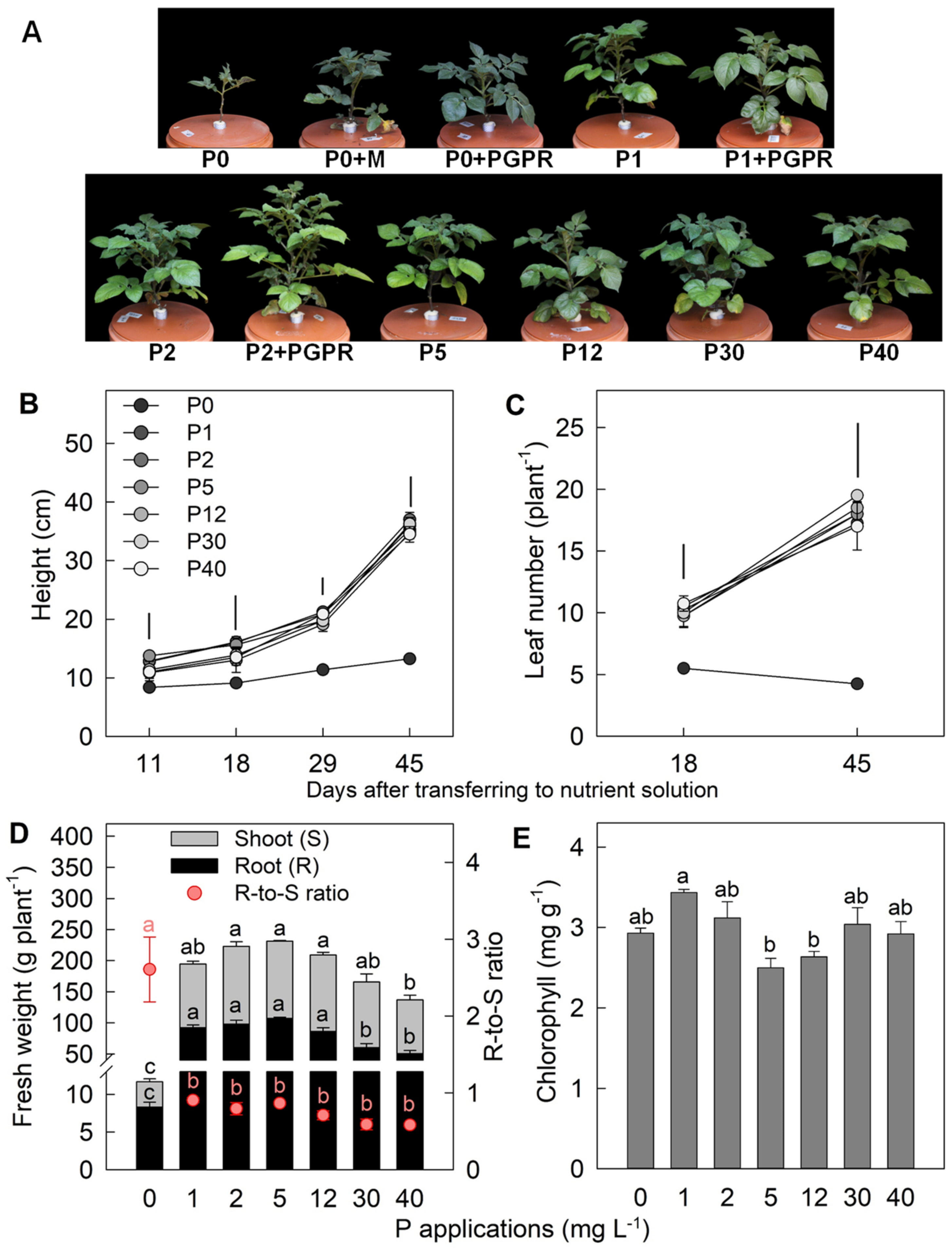
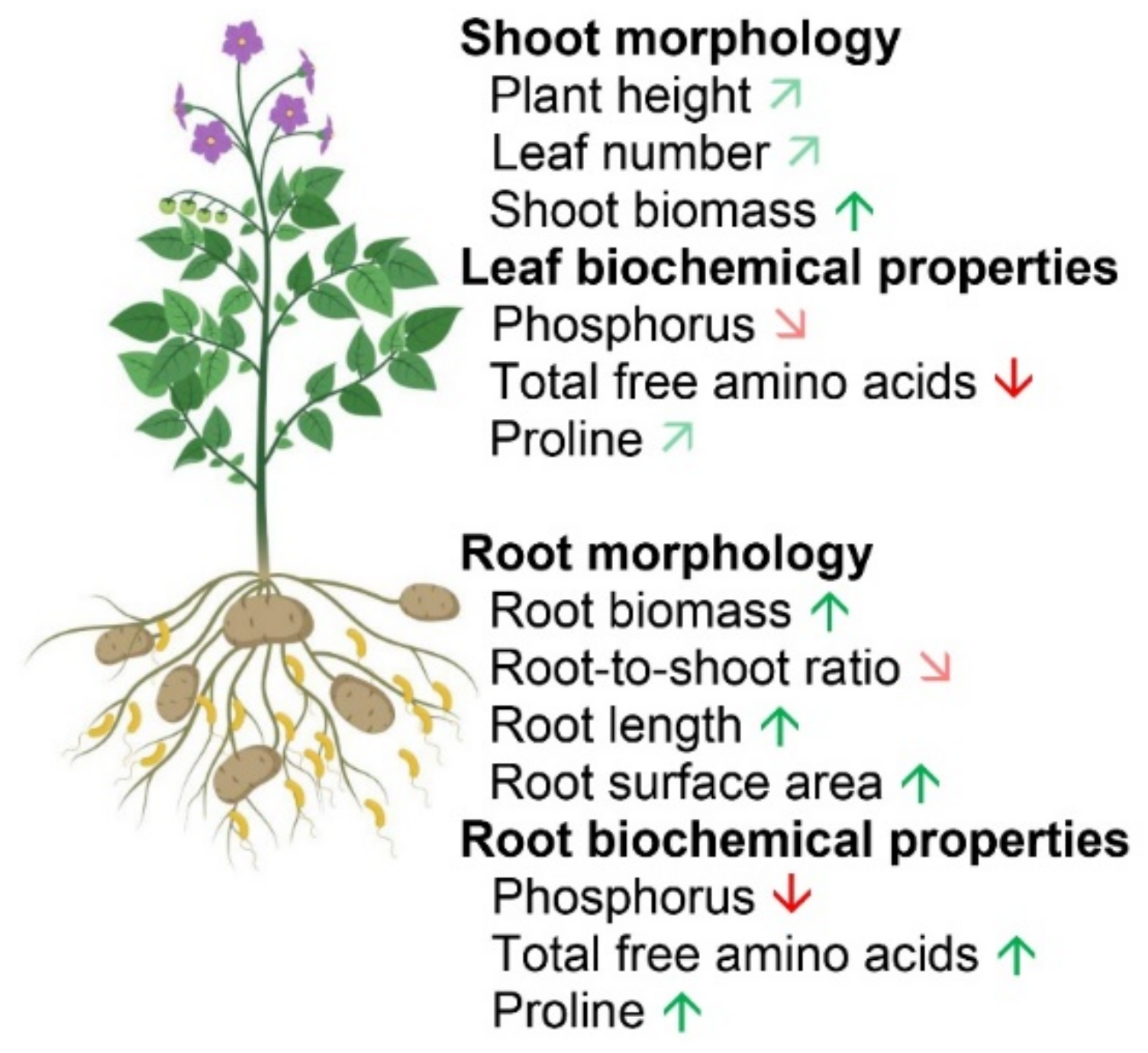
2.1. Effect of P Applications on Plant Morphology and Leaf Chlorophyll Concentration
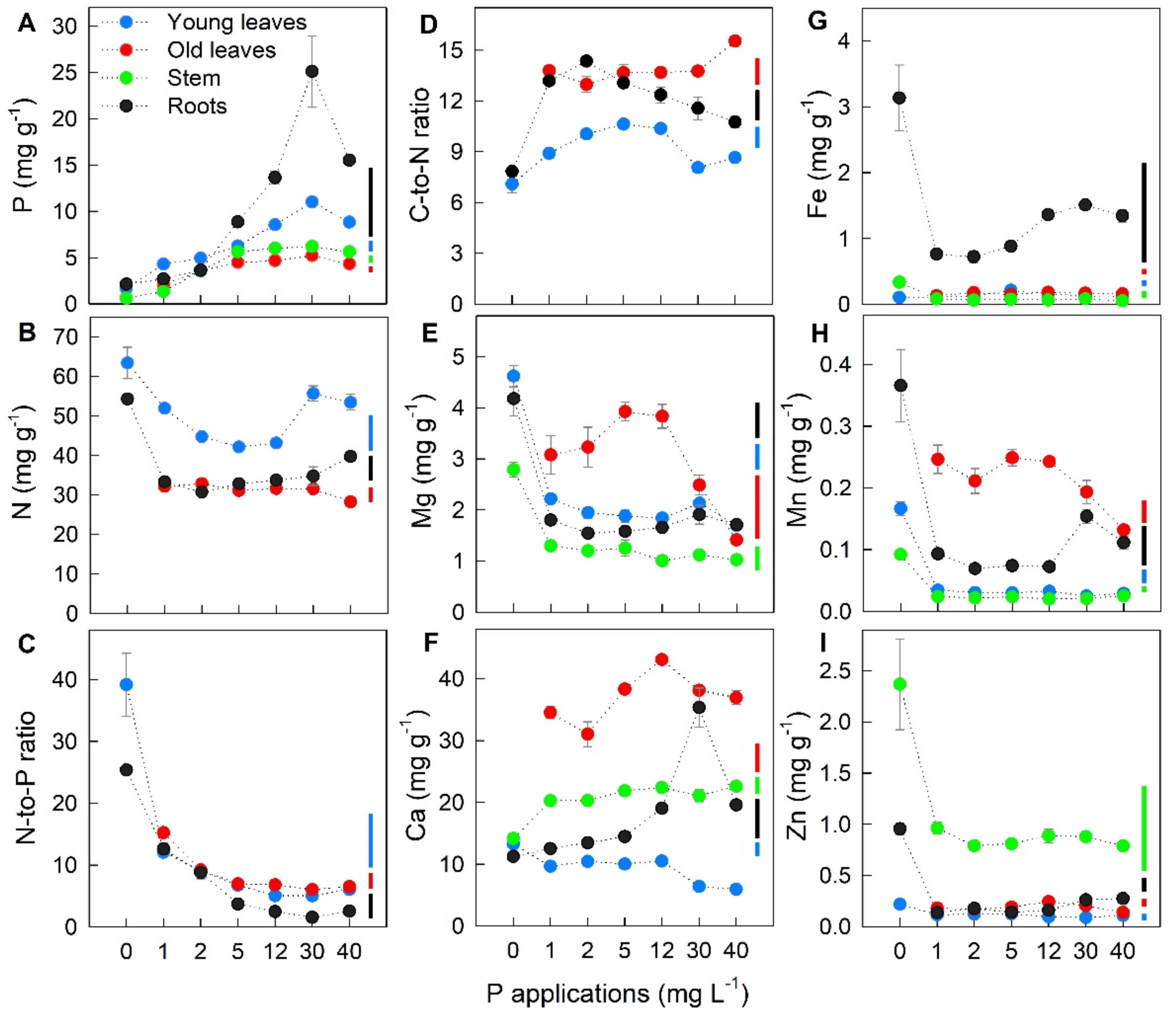
2.2. Effect of P Applications on Nutrient Concentration in Different Plant Parts
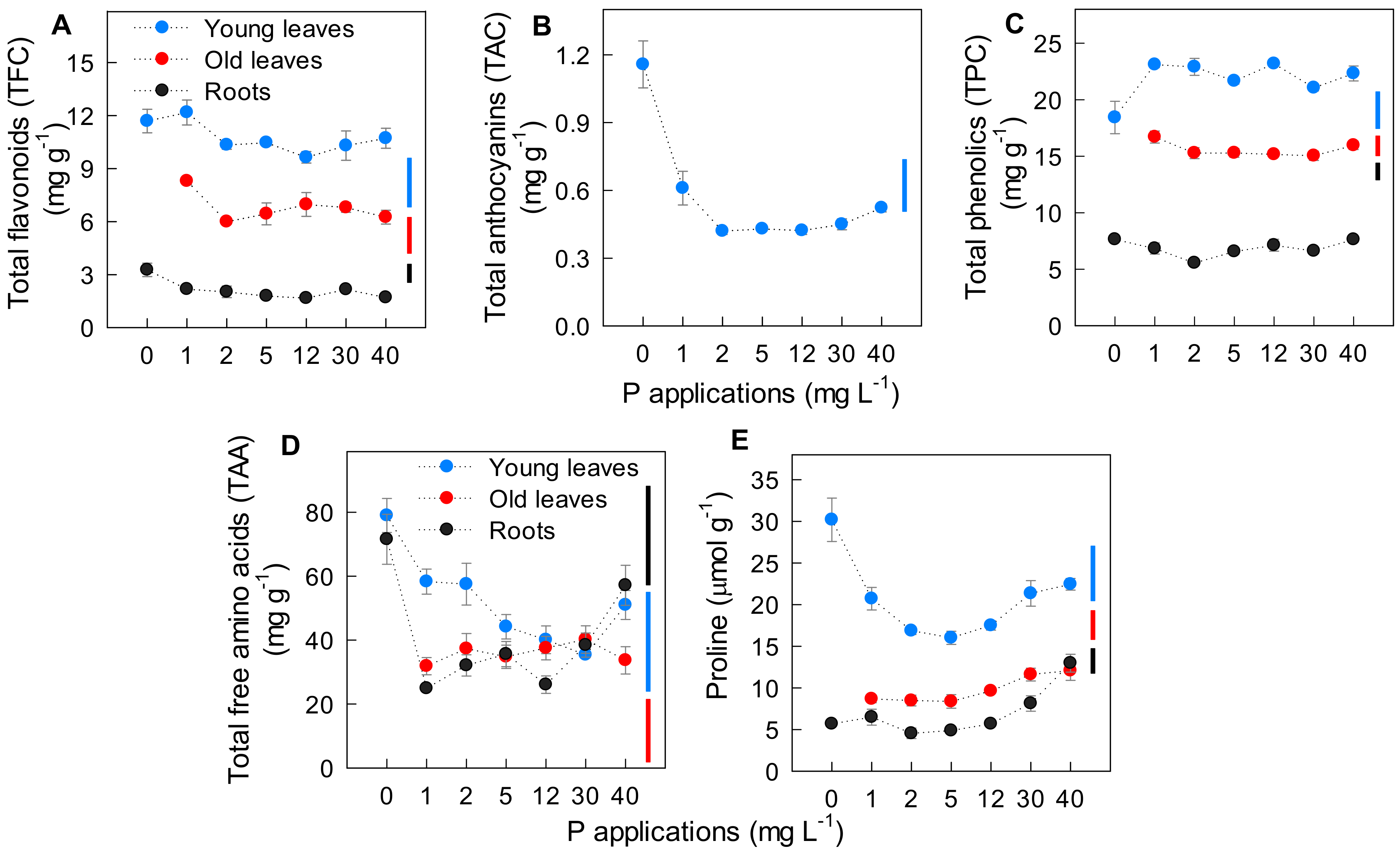
2.3. Effect of P Applications on Plant Secondary Metabolites and Amino Acids
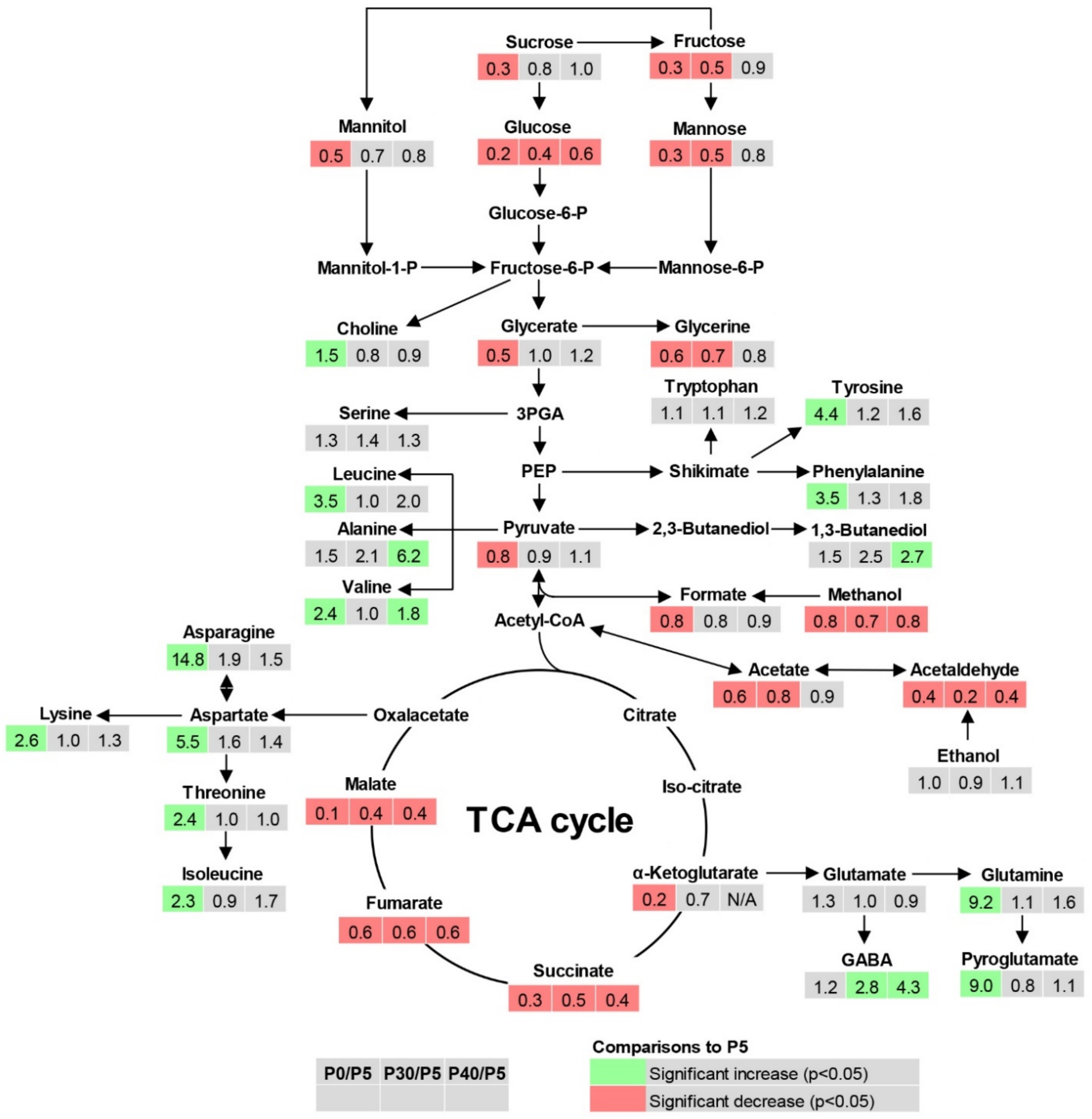
2.4. Root Metabolite Profiling under P Deficiency and Toxicity Conditions
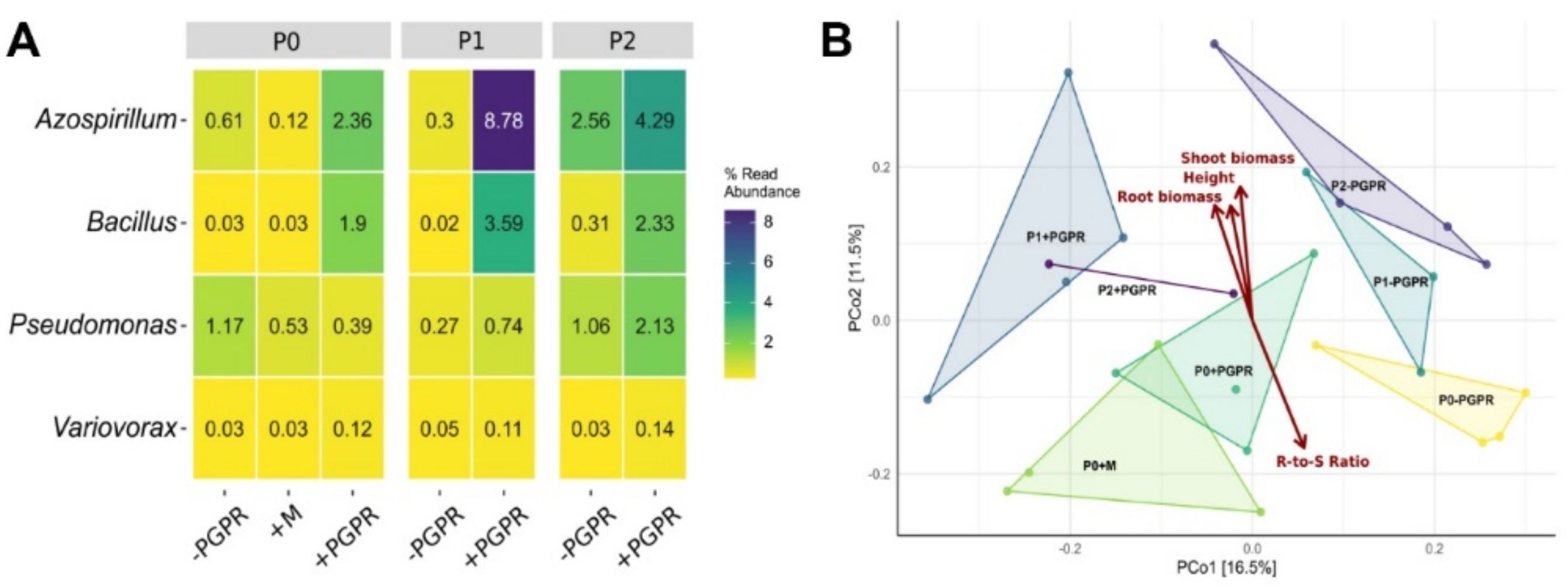
2.5. Effect of PGPR Inoculation on Plant Growth and Secondary Metabolites at Low P Supplies
2.6. Root-Associated Bacterial Community
3. Discussion
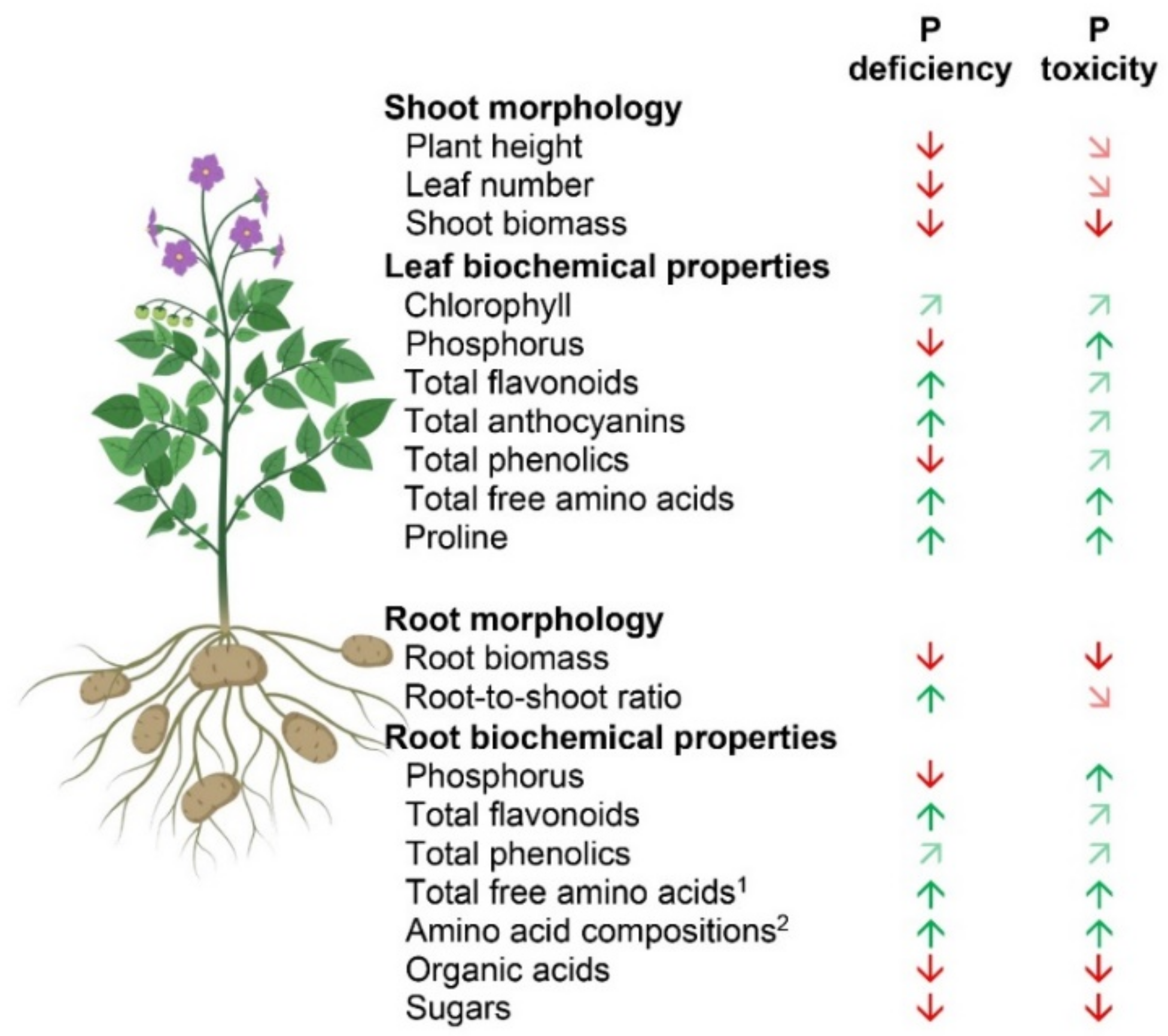
3.1. P Deficiency and Toxicity Conditions for Potato
3.2. Effect of P Deficiency and Toxicity on Plant Growth and Nutrient Assimilation
3.3. Effect of P Deficiency and Toxicity on Plant Secondary Metabolite and Root Metabolite Profiling
3.4. Plant Growth Promotion by PGPR under Low P Conditions
4. Materials and Methods
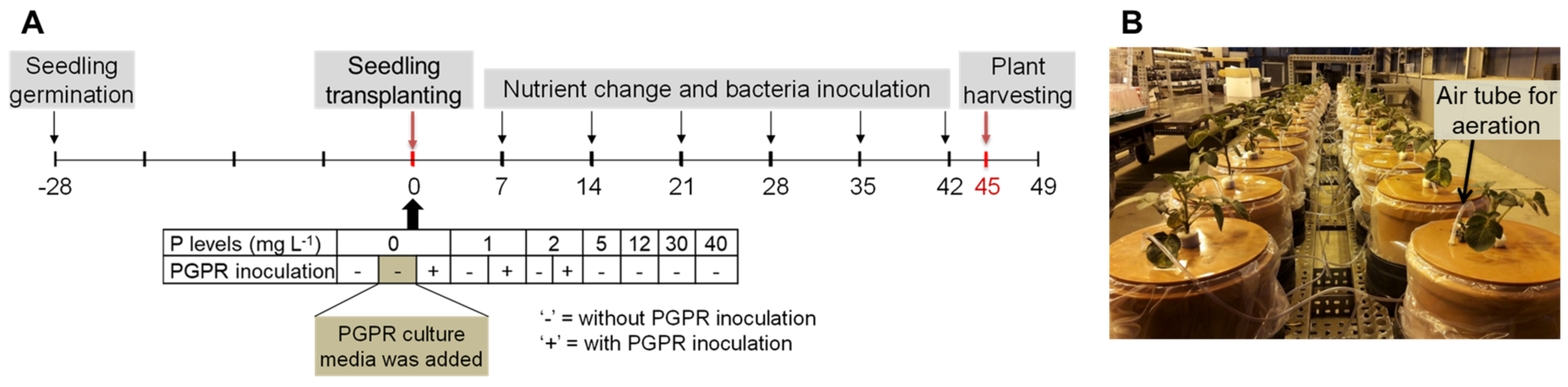
4.1. Experiment Setup and Plant Cultivation
4.2. Bacteria Culture and Inoculation
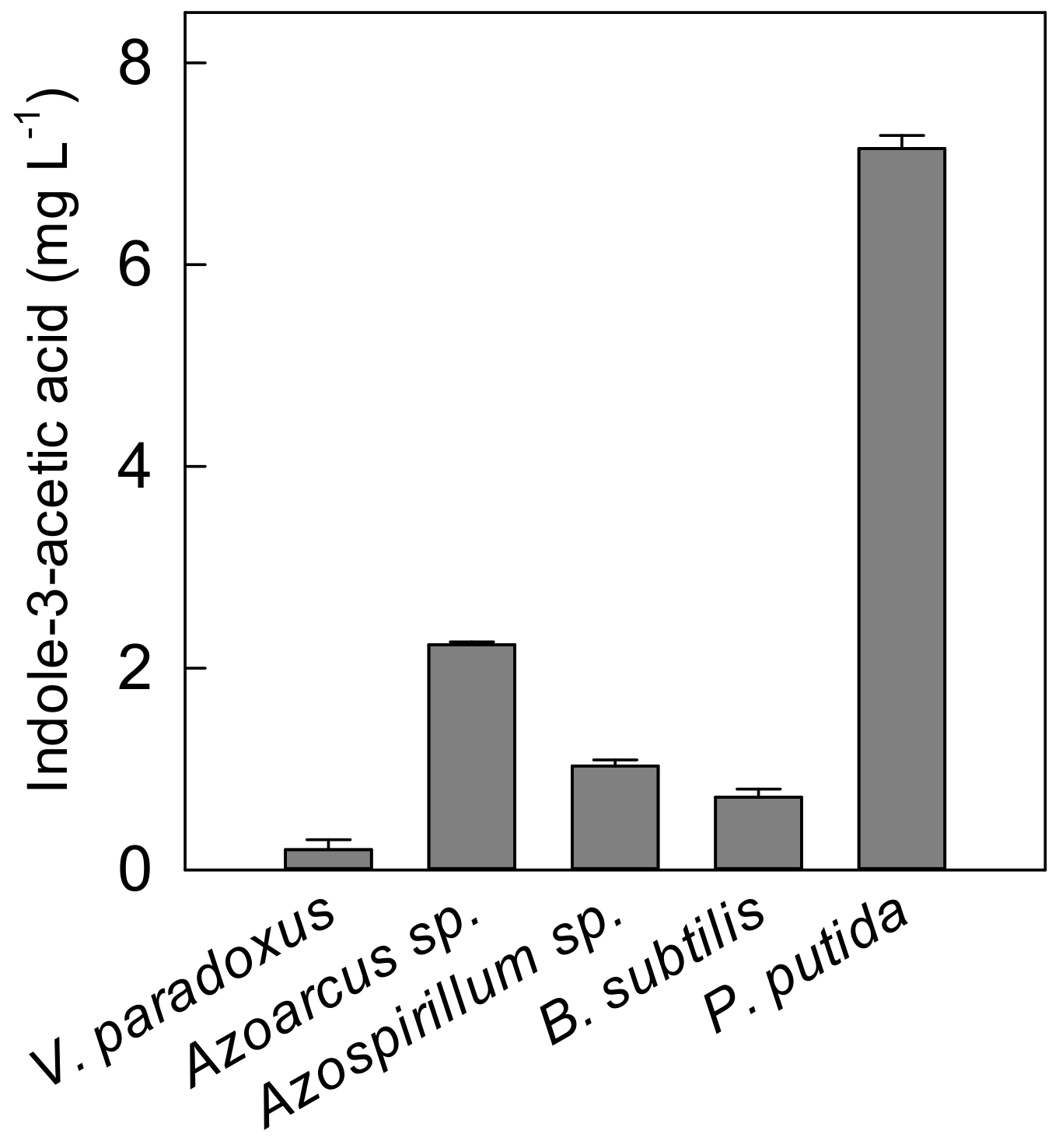
4.3. Quantification of Bacteria Derived Indole-3-Acetic acid
4.4. Plant Growth Measurements, Harvest, and Sample Processing
4.5. Mineral Nutrient Analyses
4.6. Chlorophyll, Proline, and Total Free Amino Acid Analyses
4.7. Secondary Metabolite Analyses
4.8. Root Metabolite Profiling
4.9. Root-Associated Bacterial Community
4.9.1. DNA Isolation and 16S rRNA Gene Amplification
4.9.2. Sequence Data Processing and Analyses
4.10. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hawkesford, M.; Horst, W.; Kichey, T.; Lambers, H.; Schjoerring, J.; Møller, I.S.; White, P. Functions of macronutrients. In Marschner’s Mineral Nutrition of Higher Plants, 3rd ed.; Marschner, P., Ed.; Elsevier: Amsterdam, NY, USA; Academic Press: Boston, MA, USA, 2012; ISBN 978-0-12-384905-2. [Google Scholar]
- Vance, C.P.; Uhde-Stone, C.; Allan, D.L. Phosphorus acquisition and use: Critical adaptations by plants for securing a nonrenewable resource. New Phytol. 2003, 157, 423–447. [Google Scholar] [CrossRef] [Green Version]
- MacDonald, G.K.; Bennett, E.M.; Potter, P.A.; Ramankutty, N. Agronomic phosphorus imbalances across the world’s croplands. Proc. Natl. Acad. Sci. USA 2011, 108, 3086–3091. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nawara, S.; van Dael, T.; Merckx, R.; Amery, F.; Elsen, A.; Odeurs, W.; Vandendriessche, H.; Mcgrath, S.; Roisin, C.; Jouany, C.; et al. A comparison of soil tests for available phosphorus in long-term field experiments in Europe. Eur. J. Soil. Sci. 2017, 68, 873–885. [Google Scholar] [CrossRef]
- Dechassa, N.; Schenk, M.K.; Claassen, N.; Steingrobe, B. Phosphorus efficiency of cabbage (Brassica oleraceae L. var capitata), carrot (Daucus carota L.), and potato (Solanum tuberosum L.). Plant Soil 2003, 250, 215–224. [Google Scholar] [CrossRef]
- White, P.J.; Bradshaw, J.E.; Brown, L.K.; Dale, M.F.B.; Dupuy, L.X.; George, T.S.; Hammond, J.P.; Subramanian, N.K.; Thompson, J.A.; Wishart, J.; et al. Juvenile root vigour improves phosphorus use efficiency of potato. Plant Soil 2018, 432, 45–63. [Google Scholar] [CrossRef]
- Barben, S.A.; Hopkins, B.G.; Jolley, V.D.; Webb, B.L.; Nichols, B.A. Optimizing phosphorus and zinc concentrations in hydroponic chelator-buffered nutrient solution for Russet Burbank potato. J. Plant Nutr. 2010, 33, 557–570. [Google Scholar] [CrossRef]
- Baron, D.; Amaro, A.C.E.; Campos, F.G.; Boaro, C.S.F.; Ferreira, G. Plant physiological responses to nutrient solution. In Plant Metabolites and Regulation under Environmental Stress; Ahmad, P., Abass Ahanger, M., Singh, V.P., Tripathi, D.K., Alam, P., Eds.; Academic Press: Amsterdam, The Netherlands, 2018; pp. 415–425. ISBN 9780128126899. [Google Scholar]
- Wang, Y.-L.; Almvik, M.; Clarke, N.; Eich-Greatorex, S.; Øgaard, A.F.; Krogstad, T.; Lambers, H.; Clarke, J.L. Contrasting responses of root morphology and root-exuded organic acids to low phosphorus availability in three important food crops with divergent root traits. AoB Plants 2015, 7. [Google Scholar] [CrossRef]
- Wissuwa, M.; Gamat, G.; Ismail, A.M. Is root growth under phosphorus deficiency affected by source or sink limitations? J. Exp. Bot. 2005, 56, 1943–1950. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fernandes, A.M.; Soratto, R.P. Nutrition, dry matter accumulation and partitioning and phosphorus use efficiency of potato grown at different phosphorus levels in nutrient solution. R. Bras. Ci. Solo 2012, 36, 1528–1537. [Google Scholar] [CrossRef] [Green Version]
- Stewart, A.J.; Chapman, W.; Jenkins, G.I.; Graham, I.; Martin, T.; Crozier, A. The effect of nitrogen and phosphorus deficiency on flavonol accumulation in plant tissues. Plant Cell Environ. 2001, 24, 1189–1197. [Google Scholar] [CrossRef]
- Signorelli, S.; Coitiño, E.L.; Borsani, O.; Monza, J. Molecular mechanisms for the reaction between (˙)OH radicals and proline: Insights on the role as reactive oxygen species scavenger in plant stress. J. Phys. Chem. B 2014, 118, 37–47. [Google Scholar] [CrossRef]
- Aleksza, D.; Horváth, G.V.; Sándor, G.; Szabados, L. Proline accumulation is regulated by transcription factors associated with phosphate starvation. Plant Physiol. 2017, 175, 555–567. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nguyen, V.L.; Palmer, L.; Roessner, U.; Stangoulis, J. Genotypic variation in the root and shoot metabolite profiles of wheat (Triticum aestivum L.) indicate sustained, preferential carbon allocation as a potential mechanism in phosphorus efficiency. Front. Plant Sci. 2019, 10, 995. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vacheron, J.; Desbrosses, G.; Bouffaud, M.-L.; Touraine, B.; Moënne-Loccoz, Y.; Muller, D.; Legendre, L.; Wisniewski-Dyé, F.; Prigent-Combaret, C. Plant growth-promoting rhizobacteria and root system functioning. Front. Plant Sci. 2013, 4, 356. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, F.; Chen, L.; Belimov, A.A.; Shaposhnikov, A.I.; Gong, F.; Meng, X.; Hartung, W.; Jeschke, D.W.; Davies, W.J.; Dodd, I.C. Multiple impacts of the plant growth-promoting rhizobacterium Variovorax paradoxus 5C-2 on nutrient and ABA relations of Pisum sativum. J. Exp. Bot. 2012, 63, 6421–6430. [Google Scholar] [CrossRef]
- Naqqash, T.; Hameed, S.; Imran, A.; Hanif, M.K.; Majeed, A.; van Elsas, J.D. Differential Response of Potato Toward Inoculation with Taxonomically Diverse Plant Growth Promoting Rhizobacteria. Front. Plant Sci. 2016, 7, 144. [Google Scholar] [CrossRef] [Green Version]
- Andreote, F.D.; de Araújo, W.L.; de Azevedo, J.L.; van Elsas, J.D.; da Rocha, U.N.; van Overbeek, L.S. Endophytic colonization of potato (Solanum tuberosum L.) by a novel competent bacterial endophyte, Pseudomonas putida strain P9, and its effect on associated bacterial communities. Appl. Environ. Microbiol. 2009, 75, 3396–3406. [Google Scholar] [CrossRef] [Green Version]
- Menéndez, E.; Paço, A. Is the application of plant probiotic bacterial consortia always beneficial for plants? Exploring synergies between rhizobial and non-rhizobial bacteria and their effects on agro-economically valuable crops. Life 2020, 10, 24. [Google Scholar] [CrossRef] [Green Version]
- Huett, D.O.; Maier, N.A.; Sparrow, L.A.; Piggott, T.J. Vegetable crops. In Plant Analysis: An Interpretation Manual, 2nd ed.; Reuter, D.J., Robinson, J.B., Dutkiewicz, C., Eds.; CSIRO Publishing: Collingwood, VIC, Australia, 1997; pp. 383–464. ISBN 0643059385. [Google Scholar]
- Bhatti, A.S.; Loneragan, J.F. The effect of early superphosphate toxicity on the subsequent growth of wheat. Aust. J. Agric. Res. 1970, 21, 881–892. [Google Scholar] [CrossRef]
- Chiera, J.; Thomas, J.; Rufty, T. Leaf initiation and development in soybean under phosphorus stress. J. Exp. Bot. 2002, 53, 473–481. [Google Scholar] [CrossRef] [Green Version]
- Mengel, K.; Kirkby, E.A.; Kosegarten, H.; Appel, T. Principles of Plant Nutrition; Springer: Dordrecht, The Netherlands, 2001; ISBN 978-1-4020-0008-9. [Google Scholar]
- Schachtman, D.P.; Reid, R.J.; Ayling, S.M. Phosphorus uptake by plants: From soil to cell. Plant Physiol. 1998, 116, 447–453. [Google Scholar] [CrossRef] [Green Version]
- Shukla, D.; Rinehart, C.A.; Sahi, S.V. Comprehensive study of excess phosphate response reveals ethylene mediated signaling that negatively regulates plant growth and development. Sci. Rep. 2017, 7, 3074. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kane, C.D.; Jasoni, R.L.; Peffley, E.P.; Thompson, L.D.; Green, C.J.; Pare, P.; Tissue, D. Nutrient solution and solution pH influences on onion growth and mineral content. J. Plant Nutr. 2006, 29, 375–390. [Google Scholar] [CrossRef]
- Pang, J.; Tibbett, M.; Denton, M.D.; Lambers, H.; Siddique, K.H.M.; Ryan, M.H. Soil phosphorus supply affects nodulation and N:P ratio in 11 perennial legume seedlings. Crop Pasture Sci. 2011, 62, 992. [Google Scholar] [CrossRef]
- Shi, Q.; Pang, J.; Yong, J.W.H.; Bai, C.; Pereira, C.G.; Song, Q.; Wu, D.; Dong, Q.; Cheng, X.; Wang, F.; et al. Phosphorus-fertilisation has differential effects on leaf growth and photosynthetic capacity of Arachis hypogaea L. Plant Soil 2019, 447, 99–116. [Google Scholar] [CrossRef]
- Misson, J.; Raghothama, K.G.; Jain, A.; Jouhet, J.; Block, M.A.; Bligny, R.; Ortet, P.; Creff, A.; Somerville, S.; Rolland, N.; et al. A genome-wide transcriptional analysis using Arabidopsis thaliana Affymetrix gene chips determined plant responses to phosphate deprivation. Proc. Natl. Acad. Sci. USA 2005, 102, 11934–11939. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zheng, H.; Wang, Y.; Zhao, J.; Shi, X.; Ma, Z.; Fan, M. Tuber formation as influenced by the C: N ratio in potato plants. J. Plant Nutr. Soil Sci. 2018, 181, 686–693. [Google Scholar] [CrossRef]
- Koch, M.; Busse, M.; Naumann, M.; Jákli, B.; Smit, I.; Cakmak, I.; Hermans, C.; Pawelzik, E. Differential effects of varied potassium and magnesium nutrition on production and partitioning of photoassimilates in potato plants. Physiol. Plant. 2019, 166, 921–935. [Google Scholar] [CrossRef] [Green Version]
- Hirsch, J.; Marin, E.; Floriani, M.; Chiarenza, S.; Richaud, P.; Nussaume, L.; Thibaud, M.C. Phosphate deficiency promotes modification of iron distribution in Arabidopsis plants. Biochimie 2006, 88, 1767–1771. [Google Scholar] [CrossRef] [PubMed]
- Gunes, A.; Inal, A. Phosphorus efficiency in sunflower cultivars and its relationships with phosphorus, calcium, iron, zinc and manganese Nutrition. J. Plant Nutr. 2009, 32, 1201–1218. [Google Scholar] [CrossRef]
- Criado, M.V.; Veliz, C.G.; Roberts, I.N.; Caputo, C. Phloem transport of amino acids is differentially altered by phosphorus deficiency according to the nitrogen availability in young barley plants. Plant Growth Regul. 2017, 82, 151–160. [Google Scholar] [CrossRef]
- Volf, I.; Ignat, I.; Neamtu, M.; Popa, V. Thermal stability, antioxidant activity, and photo-oxidation of natural polyphenols. Chem. Pap. 2014, 68, 349. [Google Scholar] [CrossRef]
- Fini, A.; Brunetti, C.; Di Ferdinando, M.; Ferrini, F.; Tattini, M. Stress-induced flavonoid biosynthesis and the antioxidant machinery of plants. Plant Signal. Behav. 2011, 6, 709–711. [Google Scholar] [CrossRef] [PubMed]
- Signorelli, S.; Dans, P.D.; Coitiño, E.L.; Borsani, O.; Monza, J. Connecting proline and γ-aminobutyric acid in stressed plants through non-enzymatic reactions. PLoS ONE 2015, 10, e0115349. [Google Scholar] [CrossRef] [Green Version]
- Tränkner, M.; Tavakol, E.; Jákli, B. Functioning of potassium and magnesium in photosynthesis, photosynthate translocation and photoprotection. Physiol. Plant. 2018. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Silber, A.; Ben-Jaacov, J.; Ackerman, A.; Bar-Tal, A.; Levkovitch, I.; Matsevitz-Yosef, T.; Swartzberg, D.; Riov, J.; Granot, D. Interrelationship between phosphorus toxicity and sugar metabolism in Verticordia plumosa L. Plant Soil 2002, 245, 249–260. [Google Scholar] [CrossRef]
- Li, K.; Xu, C.; Zhang, K.; Yang, A.; Zhang, J. Proteomic analysis of roots growth and metabolic changes under phosphorus deficit in maize (Zea mays L.) plants. Proteomics 2007, 7, 1501–1512. [Google Scholar] [CrossRef] [PubMed]
- Huang, C.Y.; Roessner, U.; Eickmeier, I.; Genc, Y.; Callahan, D.L.; Shirley, N.; Langridge, P.; Bacic, A. Metabolite profiling reveals distinct changes in carbon and nitrogen metabolism in phosphate-deficient barley plants (Hordeum vulgare L.). Plant Cell Physiol. 2008, 49, 691–703. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Müller, J.; Gödde, V.; Niehaus, K.; Zörb, C. Metabolic adaptations of white lupin roots and shoots under phosphorus deficiency. Front. Plant Sci. 2015, 6, 1014. [Google Scholar] [CrossRef] [Green Version]
- Chia, D.W.; Yoder, T.J.; Reiter, W.D.; Gibson, S.I. Fumaric acid: An overlooked form of fixed carbon in Arabidopsis and other plant species. Planta 2000, 211, 743–751. [Google Scholar] [CrossRef] [Green Version]
- Singh, A.; Roychoudhury, A. Role of γ-aminobutyric acid in the mitigation of abiotic stress in plants. In Protective Chemical Agents in the Amelioration of Plant Abiotic Stress; Roychoudhury, A., Tripathi, D.K., Eds.; Wiley: Hoboken, NJ, USA, 2020; pp. 413–423. ISBN 9781119551638. [Google Scholar]
- Galili, G. The aspartate-family pathway of plants: Linking production of essential amino acids with energy and stress regulation. Plant Signal. Behav. 2011, 6, 192–195. [Google Scholar] [CrossRef] [Green Version]
- Fernández, H.; Prandoni, N.; Fernández-Pascual, M.; Fajardo, S.; Morcillo, C.; Díaz, E.; Carmona, M. Azoarcus sp. CIB, an anaerobic biodegrader of aromatic compounds shows an endophytic lifestyle. PLoS ONE 2014, 9, e110771. [Google Scholar] [CrossRef] [PubMed]
- Kaneko, T.; Minamisawa, K.; Isawa, T.; Nakatsukasa, H.; Mitsui, H.; Kawaharada, Y.; Nakamura, Y.; Watanabe, A.; Kawashima, K.; Ono, A.; et al. Complete genomic structure of the cultivated rice endophyte Azospirillum sp. B510. DNA Res. 2010, 17, 37–50. [Google Scholar] [CrossRef] [Green Version]
- Pathak, D.; Lone, R.; Khan, S.; Koul, K.K. Isolation, screening and molecular characterization of free-living bacteria of potato (Solanum tuberosum L.) and their interplay impact on growth and production of potato plant under Mycorrhizal association. Sci. Hortic. 2019, 252, 388–397. [Google Scholar] [CrossRef]
- Patten, C.L.; Glick, B.R. Role of Pseudomonas putida indoleacetic acid in development of the host plant root system. Appl. Environ. Microbiol. 2002, 68, 3795–3801. [Google Scholar] [CrossRef] [Green Version]
- Kisand, V.; Tuvikene, L.; Nõges, T. Role of phosphorus and nitrogen for bacteria and phytopankton development in a large shallow lake. Hydrobiologia 2001, 457, 187–197. [Google Scholar] [CrossRef]
- Mhlongo, M.I.; Piater, L.A.; Steenkamp, P.A.; Labuschagne, N.; Dubery, I.A. Metabolic profiling of PGPR-treated tomato plants reveal priming-related adaptations of secondary metabolites and aromatic amino acids. Metabolites 2020, 10, 210. [Google Scholar] [CrossRef]
- Weston, D.J.; Pelletier, D.A.; Morrell-Falvey, J.L.; Tschaplinski, T.J.; Jawdy, S.S.; Lu, T.-Y.; Allen, S.M.; Melton, S.J.; Martin, M.Z.; Schadt, C.W.; et al. Pseudomonas fluorescens induces strain-dependent and strain-independent host plant responses in defense networks, primary metabolism, photosynthesis, and fitness. Mol. Plant Microbe Interact. 2012, 25, 765–778. [Google Scholar] [CrossRef] [Green Version]
- Smith, S.E.; Read, D. Growth and carbon economy of arbuscular mycorrhizal symbionts. In Mycorrhizal Symbiosis, 3rd ed.; Academic Press: Amsterdam, NY, USA; Boston, MA, USA, 2008; pp. 117–144. ISBN 9780123705266. [Google Scholar]
- Chen, L.; Hao, Z.; Li, K.; Sha, Y.; Wang, E.; Sui, X.; Mi, G.; Tian, C.; Chen, W. Effectsof growth-promoting rhizobacteria on maize growth and rhizosphere microbial community under conservation tillage in Northeast China. Microb. Biotechnol. 2020. [Google Scholar] [CrossRef]
- Pereira, S.I.A.; Abreu, D.; Moreira, H.; Vega, A.; Castro, P.M.L. Plant growth-promoting rhizobacteria (PGPR) improve the growth and nutrient use efficiency in maize (Zea mays L.) under water deficit conditions. Heliyon 2020, 6, e05106. [Google Scholar] [CrossRef] [PubMed]
- Pantigoso, H.A.; Manter, D.K.; Vivanco, J.M. Phosphorus addition shifts the microbial community in the rhizosphere of blueberry (Vaccinium corymbosum L.). Rhizosphere 2018, 7, 1–7. [Google Scholar] [CrossRef]
- Han, J.-I.; Choi, H.-K.; Lee, S.-W.; Orwin, P.M.; Kim, J.; Laroe, S.L.; Kim, T.-G.; O’Neil, J.; Leadbetter, J.R.; Lee, S.Y.; et al. Complete genome sequence of the metabolically versatile plant growth-promoting endophyte Variovorax paradoxus S110. J. Bacteriol. 2011, 193, 1183–1190. [Google Scholar] [CrossRef] [Green Version]
- Han, J.-I.; Spain, J.C.; Leadbetter, J.R.; Ovchinnikova, G.; Goodwin, L.A.; Han, C.S.; Woyke, T.; Davenport, K.W.; Orwin, P.M. Genome of the root-associated plant growth-promoting bacterium Variovorax paradoxus strain EPS. Genome Announc. 2013, 1. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gond, S.K.; Bergen, M.S.; Torres, M.S.; White, J.F., Jr. Endophytic Bacillus spp. produce antifungal lipopeptides and induce host defence gene expression in maize. Microbiol. Res. 2015, 172, 79–87. [Google Scholar] [CrossRef] [PubMed]
- Sarwar, M.; Kremer, R.J. Determination of bacterially derived auxins using a microplate method. Lett. Appl. Microbiol. 1995, 20, 282–285. [Google Scholar] [CrossRef]
- Bates, L.S.; Waldren, R.P.; Teare, I.D. Rapid determination of free proline for water-stress studies. Plant Soil 1973, 39, 205–207. [Google Scholar] [CrossRef]
- Hada, H.; Hidema, J.; Maekawa, M.; Kumagai, T. Higher amounts of anthocyanins and UV-absorbing compounds effectively lowered CPD photorepair in purple rice (Oryza sativa L.). Plant Cell Environ. 2003, 26, 1691–1701. [Google Scholar] [CrossRef]
- Chandra, S.; Khan, S.; Avula, B.; Lata, H.; Yang, M.H.; Elsohly, M.A.; Khan, I.A. Assessment of total phenolic and flavonoid content, antioxidant properties, and yield of aeroponically and conventionally grown leafy vegetables and fruit crops: A comparative study. Evid. Based Complement. Alternat. Med. 2014, 2014, 253875. [Google Scholar] [CrossRef]
- Klindworth, A.; Pruesse, E.; Schweer, T.; Peplies, J.; Quast, C.; Horn, M.; Glöckner, F.O. Evaluation of general 16S ribosomal RNA gene PCR primers for classical and next-generation sequencing-based diversity studies. Nucleic Acids Res. 2013, 41, e1. [Google Scholar] [CrossRef]
- Chen, S.; Zhou, Y.; Chen, Y.; Gu, J. fastp: An ultra-fast all-in-one FASTQ preprocessor. Bioinformatics 2018, 34, i884–i890. [Google Scholar] [CrossRef]
- Zhang, J.; Kobert, K.; Flouri, T.; Stamatakis, A. PEAR: A fast and accurate Illumina Paired-End reAd mergeR. Bioinformatics 2014, 30, 614–620. [Google Scholar] [CrossRef] [Green Version]
- Martin, M. Cutadapt removes adapter sequences from high-throughput sequencing reads. EMBnet. J. 2011, 17, 10–12. [Google Scholar] [CrossRef]
- Rognes, T.; Flouri, T.; Nichols, B.; Quince, C.; Mahé, F. VSEARCH: A versatile open source tool for metagenomics. PeerJ 2016, 4, e2584. [Google Scholar] [CrossRef]
- Quast, C.; Pruesse, E.; Yilmaz, P.; Gerken, J.; Schweer, T.; Yarza, P.; Peplies, J.; Glöckner, F.O. The SILVA ribosomal RNA gene database project: Improved data processing and web-based tools. Nucleic Acids Res. 2013, 41, 590–596. [Google Scholar] [CrossRef] [PubMed]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2019; Available online: https://www.R-project.org (accessed on 14 December 2020).
- Andersen, K.S.; Kirkegaard, R.H.; Karst, S.M.; Albertsen, M. ampvis2: An R package to analyse and visualise 16S rRNA amplicon data. bioRxiv 2018. [Google Scholar] [CrossRef] [Green Version]
- Bray, J.R.; Curtis, J.T. An ordination of the upland forest communities of southern Wisconsin. Ecol. Monogr. 1957, 27, 325–349. [Google Scholar] [CrossRef]
- Oksanen, J.; Blanchet, F.G.; Friendly, M.; Kindt, R.; Legendre, P.; McGlinn, D.; Minchin, P.R.; O’Hara, R.B.; Simpson, G.L.; Solymos, P.; et al. Vegan: Community Ecology Package. R Package Version 2.5-6. 2019. Available online: https://mran.microsoft.com/snapshot/2020-01-11/web/packages/vegan/index.html (accessed on 14 December 2020).










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| P level (mg L−1) | PGPR | Height (cm) | Leaf Number (plant−1) | Root (g plant−1) | Shoot (g plant−1) | R-to-S Ratio | Total Root Length (104 cm) | Root Surface Area (103 cm2) |
|---|---|---|---|---|---|---|---|---|
| 0 | −PGPR | 13.3 ± 0.5 b | 4.3 ± 0.3 b | 8.3 ± 0.7 c | 3.4 ± 0.4 c | 2.6 ± 0.4 | n.d. | n.d. |
| +M | 20.5 ± 0.9 a | 14.3 ± 1.4 a | 52.0 ± 3.3 b | 26.3 ± 0.7 b | 2.0 ± 0.1 | 4.7 ± 0.0 b | 2.8 ± 0.0 b | |
| +PGPR | 20.6 ± 0.9 a | 15.3 ± 0.5 a | 76.1 ± 2.6 a | 35.1 ± 2.2 a | 2.2 ± 0.2 | 6.8 ± 1.0 a | 3.7 ± 0.4 a | |
| 1 | −PGPR | 35.3 ± 1.1 | 17.3 ± 0.3 | 92.3 ± 4.3 | 102.2 ± 4.7 | 0.9 ± 0.0 | 6.6 ± 0.0 | 3.6 ± 0.0 |
| +PGPR | 32.8 ± 2.1 | 17.3 ± 1.3 | 79.9 ± 6.8 | 103.9 ± 7.8 | 0.8 ± 0.0 | 6.2 ± 0.5 | 3.5 ± 0.2 | |
| 2 | −PGPR | 37.0 ± 1.2 | 18.0 ± 1.0 | 97.9 ± 6.3 | 125.2 ± 7.4 | 0.8 ± 0.1 | 7.6 ± 0.5 | 4.3 ± 0.3 |
| +PGPR | 38.0 ± 1.6 | 17.0 ± 0.9 | 85.7 ± 7.0 | 115.6 ± 10.8 | 0.8 ± 0.1 | 6.4 ± 0.2 | 3.4 ± 0.1 |
| P Level (mg L−1) | PGPR | P (mg g−1) | Root P Uptake (mg plant−1) | Specific P Uptake (µg P cm−2) | TAA (mg g−1) | Proline (µmol g−1) | |||
|---|---|---|---|---|---|---|---|---|---|
| Leaves | Roots | Leaves | Roots | Leaves | Roots | ||||
| 0 | −PGPR | 1.7 ± 0.2 b | 2.1 ± 0.1 a | 1.1 ± 0.1 b | n.d. | 79.0 ± 8.3 a | 71.6 ± 7.8 a | 30.2 ± 2.6 | 5.7 ± 0.3 |
| +M | 2.4 ± 0.1 a | 2.3 ± 0.1 a | 7.2 ± 0.4 a | 2.6 ± 0.2 | 90.1 ± 3.9 a | 37.6 ± 4.1 b | 23.1 ± 1.1 | 6.3 ± 0.8 | |
| +PGPR | 2.3 ± 0.1 a | 1.9 ± 0.1 b | 9.2 ± 0.9 a | 2.7 ± 0.6 | 54.0 ± 8.7 b | 67.0 ± 9.3 a | 23.2 ± 1.8 | 8.0 ± 0.5 | |
| 1 | −PGPR | 4.3 ± 0.1 a | 2.7 ± 0.2 | 15.6 ± 1.3 | 3.9 ± 0.3 | 58.3 ± 3.9 | 24.5 ± 0.7 b | 20.7 ± 1.4 | 6.5 ± 1.0 |
| +PGPR | 3.5 ± 0.1 b | 2.5 ± 0.2 | 12.4 ± 1.0 | 3.6 ± 0.3 | 58.2 ± 3.3 | 82.4 ± 2.3 a | 20.5 ± 1.1 | 6.5 ± 0.0 | |
| 2 | −PGPR | 4.9 ± 0.3 | 3.6 ± 0.4 | 22.7 ± 4.1 | 4.4 ± 0.5 | 57.5 ± 12.5 | 32.1 ± 3.3 | 16.9 ± 0.4 | 4.5 ± 0.6 |
| +PGPR | 4.6 ± 0.2 | 3.8 ± 0.2 | 20.3 ± 1.8 | 5.5 ± 0.6 | 42.5 ± 4.4 | 45.1 ± 7.7 | 19.7 ± 0.6 | 5.1 ± 0.3 | |
| Bacterium | DSMZ# Code | Biome | Plant Growth-Promoting Effects |
|---|---|---|---|
| Variovorax paradoxus | DSM 30034 | Soil | Rhizosphere and endosphere bacterium of potato [58,59] Producing indole-3-acetic acid (IAA); improving plant biomass and P uptake of pea [17] |
| Azoarcus sp. | DSM 9506 | Laboratory aquifer column | Endosphere bacterium, producing IAA, fixing N2, and enhancing P uptake of rice [47] |
| Azospirillum sp. | DSM 1842 | Maize roots | Endosphere bacterium and IAA synthesis in rice [48] Producing IAA and promoting plant biomass of potato [18] |
| Bacillus subtilis | DSM 21393 | Potato tubers | Endosphere bacterium maize [60] Producing IAA; improving root length and plant biomass of potato [49] |
| Pseudomonas putida | DSM 6125 | Unknown | Endosphere bacterium of potato [19] Producing IAA, improving root growth of canola [50] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chea, L.; Pfeiffer, B.; Schneider, D.; Daniel, R.; Pawelzik, E.; Naumann, M. Morphological and Metabolite Responses of Potatoes under Various Phosphorus Levels and Their Amelioration by Plant Growth-Promoting Rhizobacteria. Int. J. Mol. Sci. 2021, 22, 5162. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22105162
Chea L, Pfeiffer B, Schneider D, Daniel R, Pawelzik E, Naumann M. Morphological and Metabolite Responses of Potatoes under Various Phosphorus Levels and Their Amelioration by Plant Growth-Promoting Rhizobacteria. International Journal of Molecular Sciences. 2021; 22(10):5162. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22105162
Chicago/Turabian StyleChea, Leangsrun, Birgit Pfeiffer, Dominik Schneider, Rolf Daniel, Elke Pawelzik, and Marcel Naumann. 2021. "Morphological and Metabolite Responses of Potatoes under Various Phosphorus Levels and Their Amelioration by Plant Growth-Promoting Rhizobacteria" International Journal of Molecular Sciences 22, no. 10: 5162. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22105162