
Antitumor Activity of Nitazoxanide against Colon Cancers: Molecular Docking and Experimental Studies Based on Wnt/β-Catenin Signaling Inhibition

 ,
,  ,
,  ,
,  , , ,
, , ,
Abstract
:1. Introduction
2. Results
2.1. Experiment 1: In Vitro Cytotoxic Activity
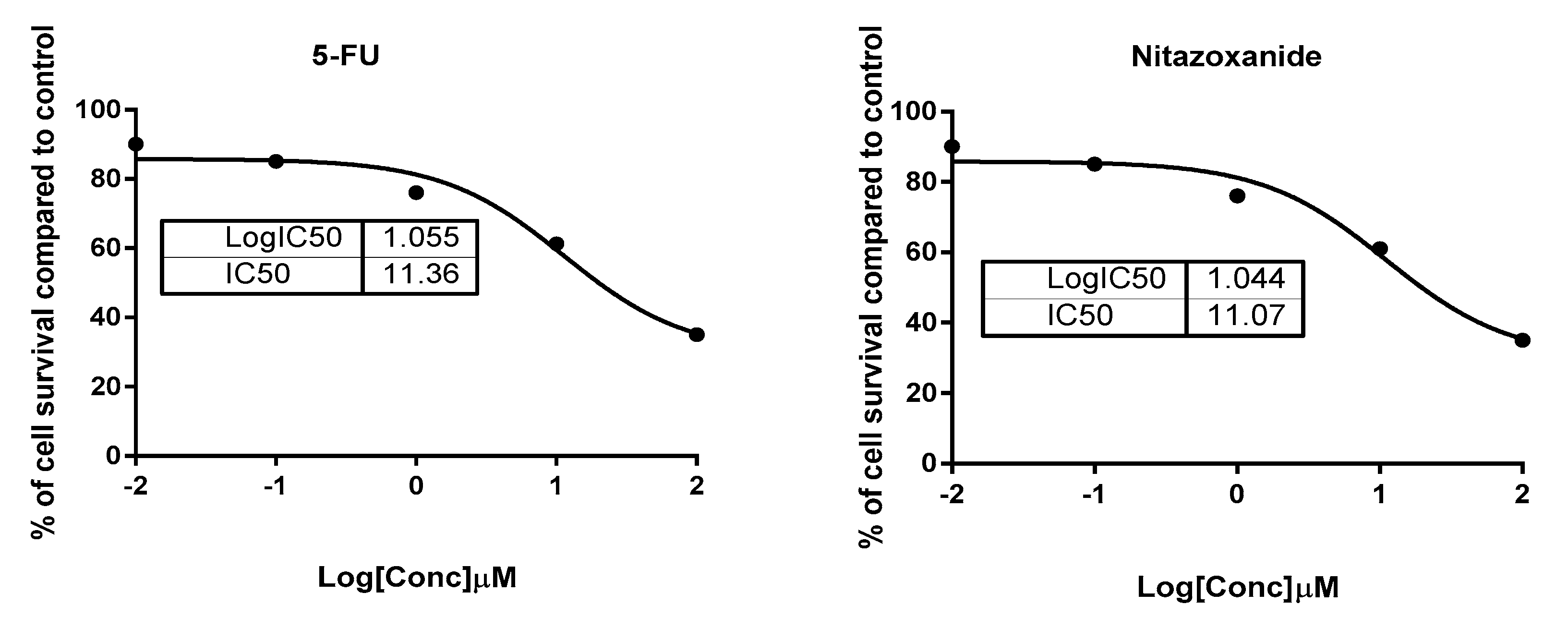
2.1.1. Cytotoxic Activity against Colon Cancer Cells and Normal Colon Cells
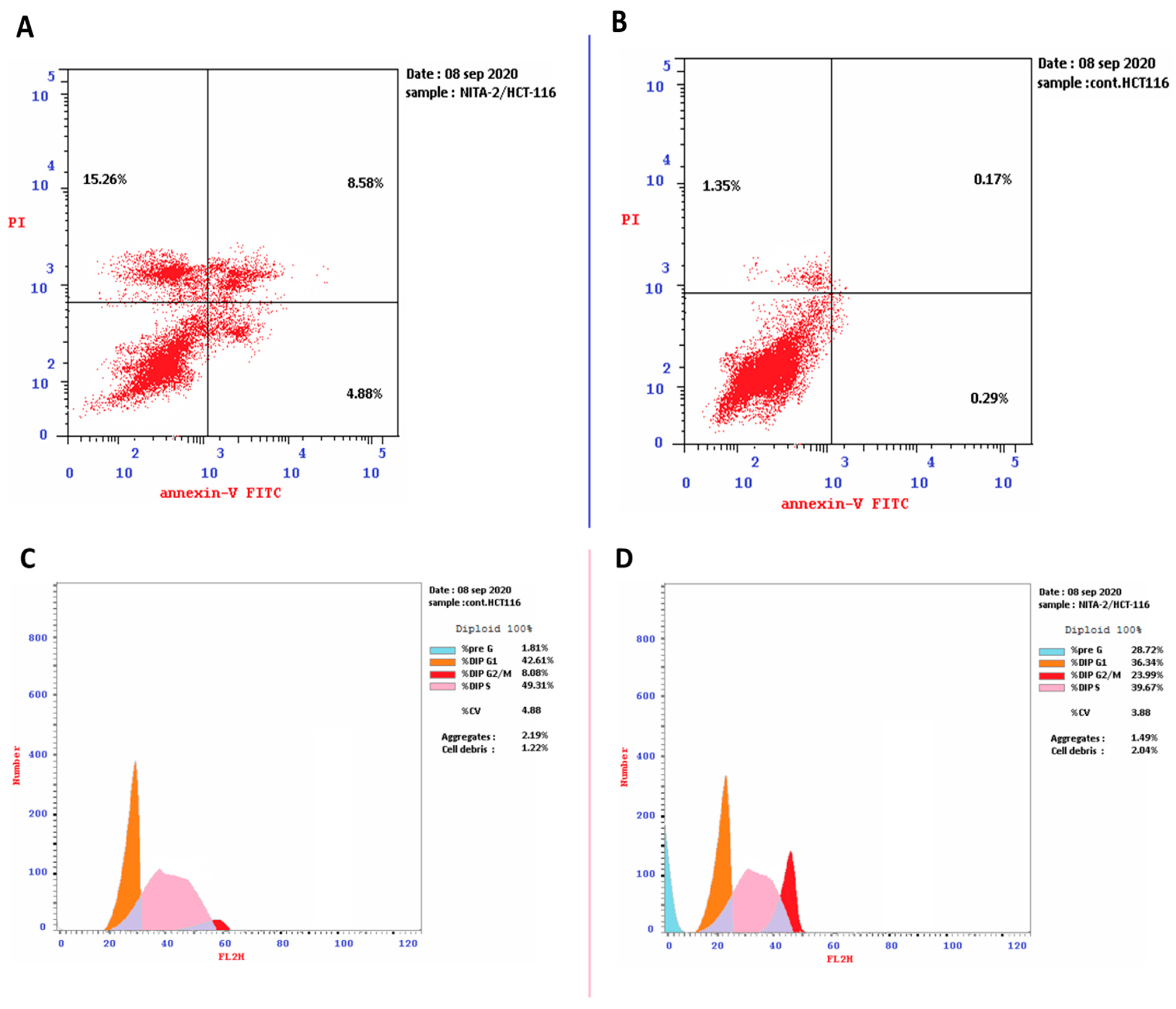
2.1.2. Exploration of Apoptosis Indicators
Annexin/Propidium Iodide (PI) Staining and Cell-Cycle Analysis
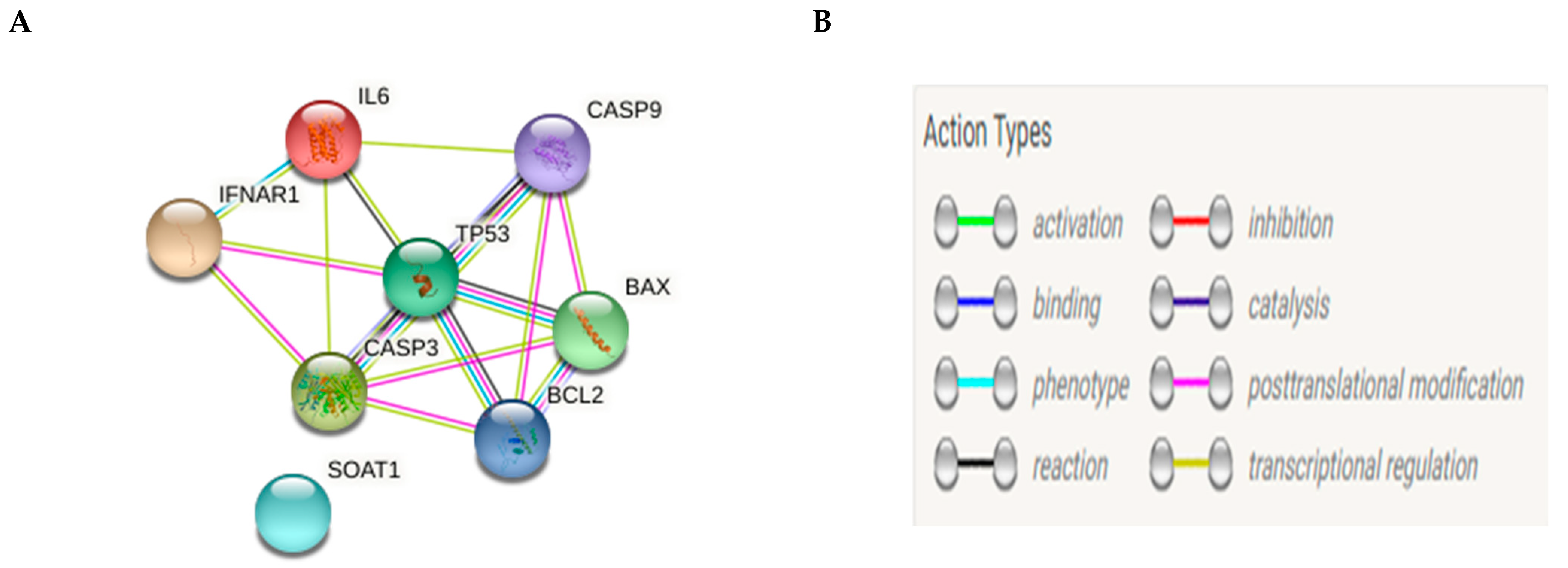
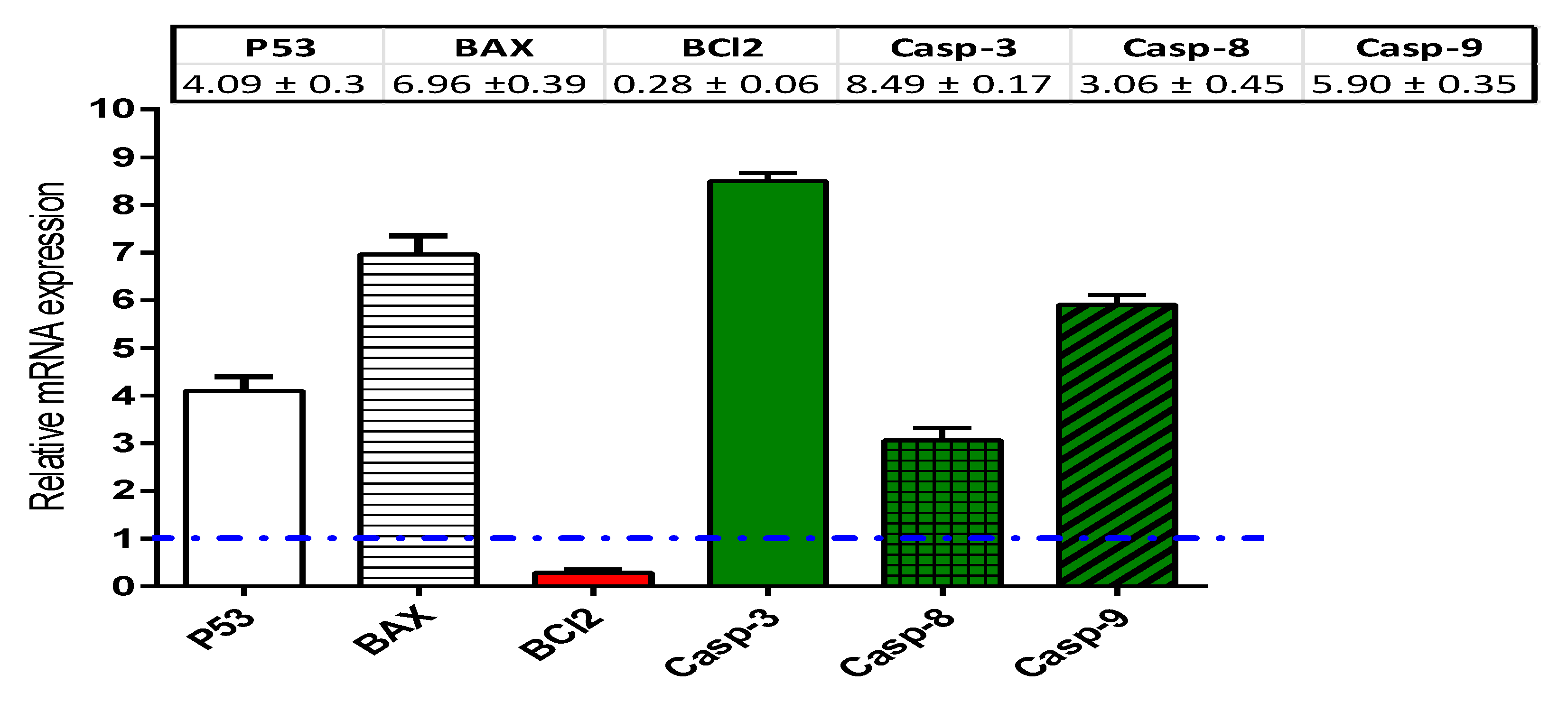
String Database Query Methods and Real-Time Polymerase Chain Reaction (RT-PCR)
2.2. Experiment 2: In Vivo Antitumor Activity
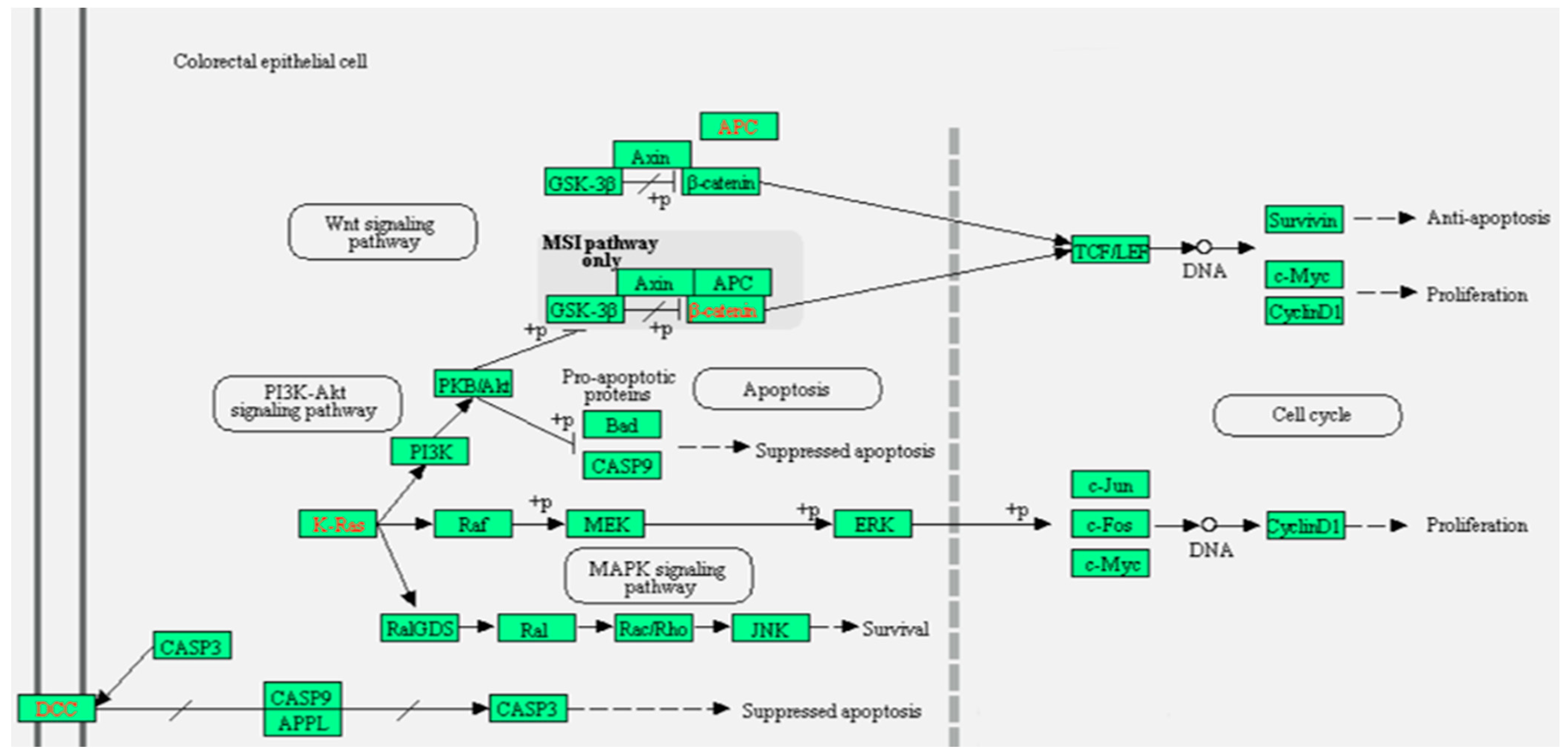
2.2.1. Pathway Enrichment Analysis
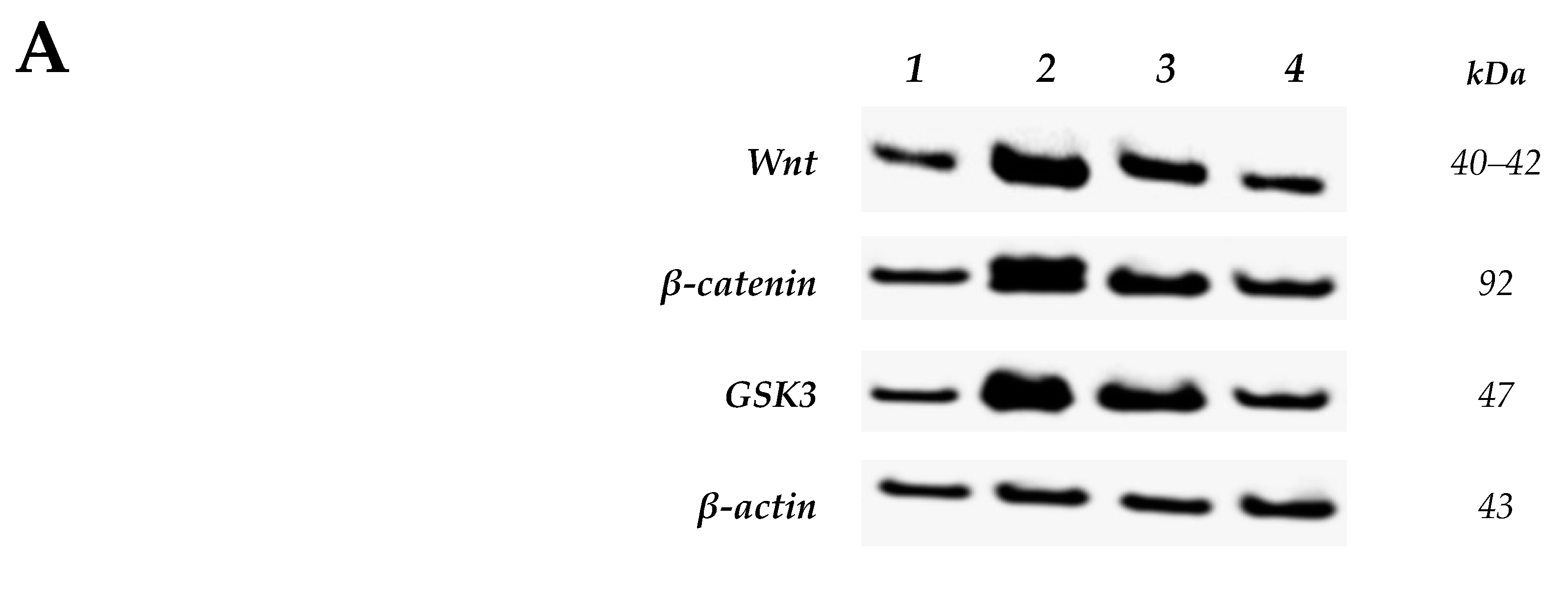
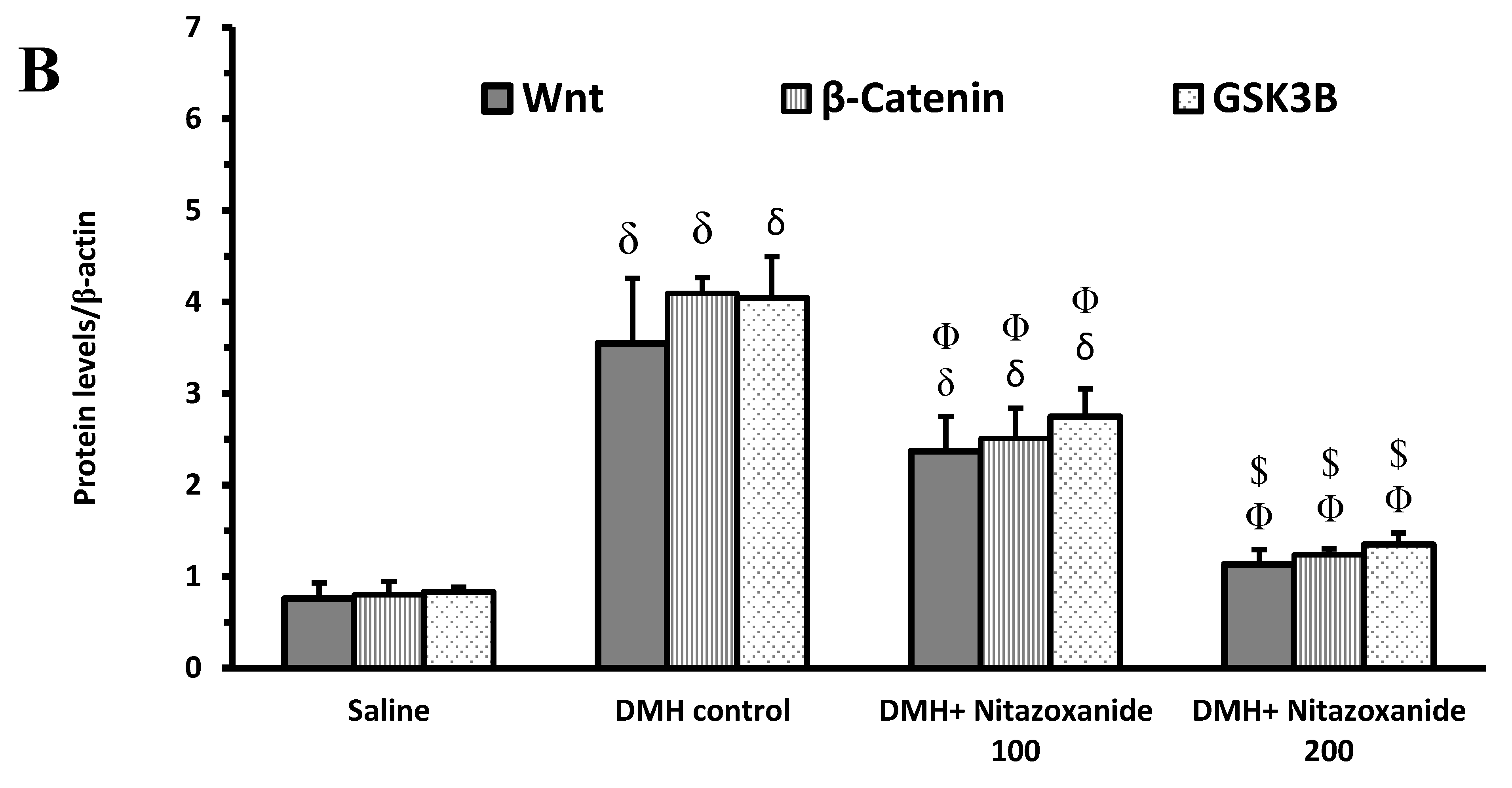
2.2.2. Effect of Nitazoxanide on the Levels of Wnt/Catenin/Glycogen Synthase Kinase-3β (GSK-3β) Proteins in Colon Cancer Model
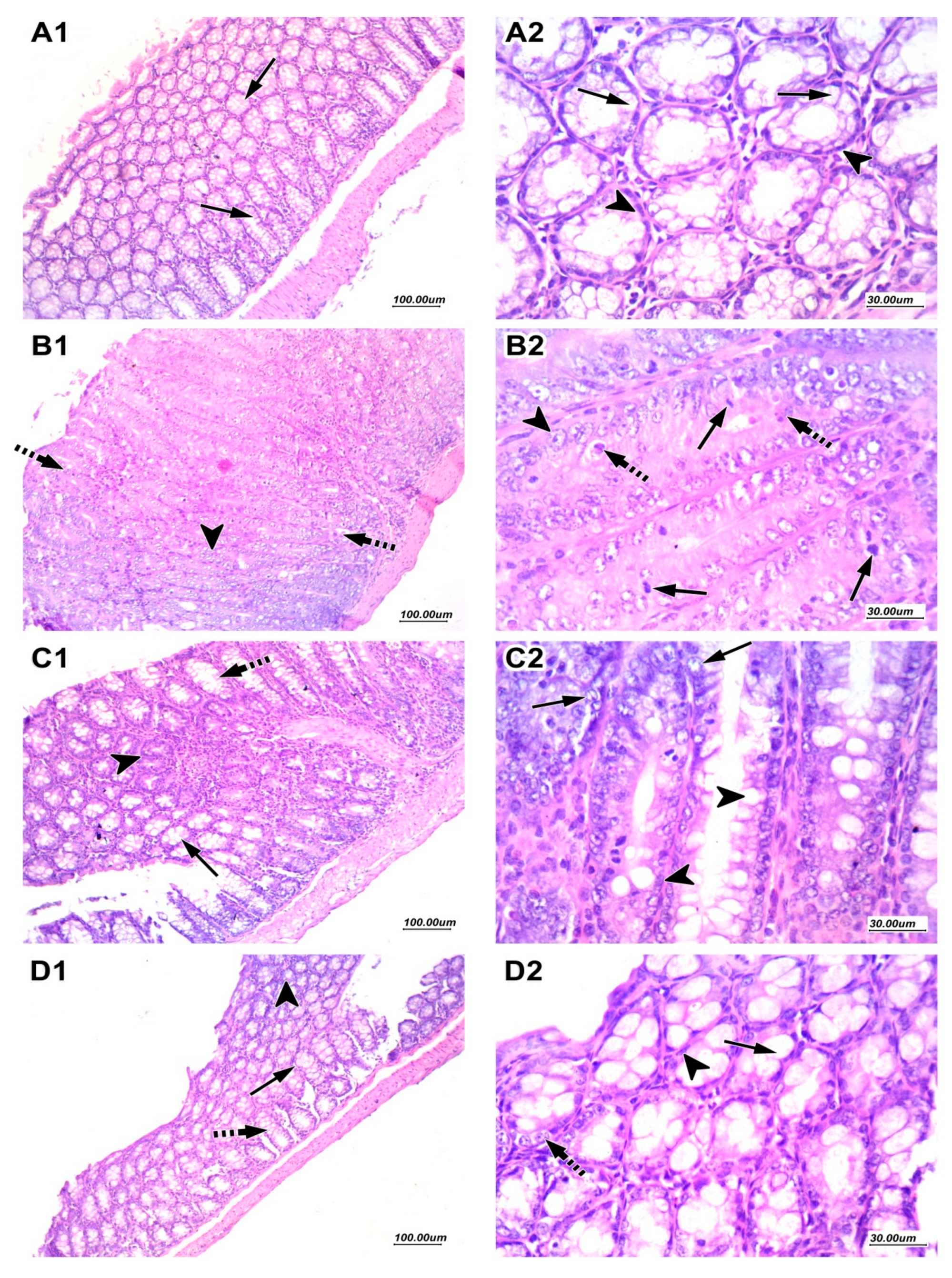
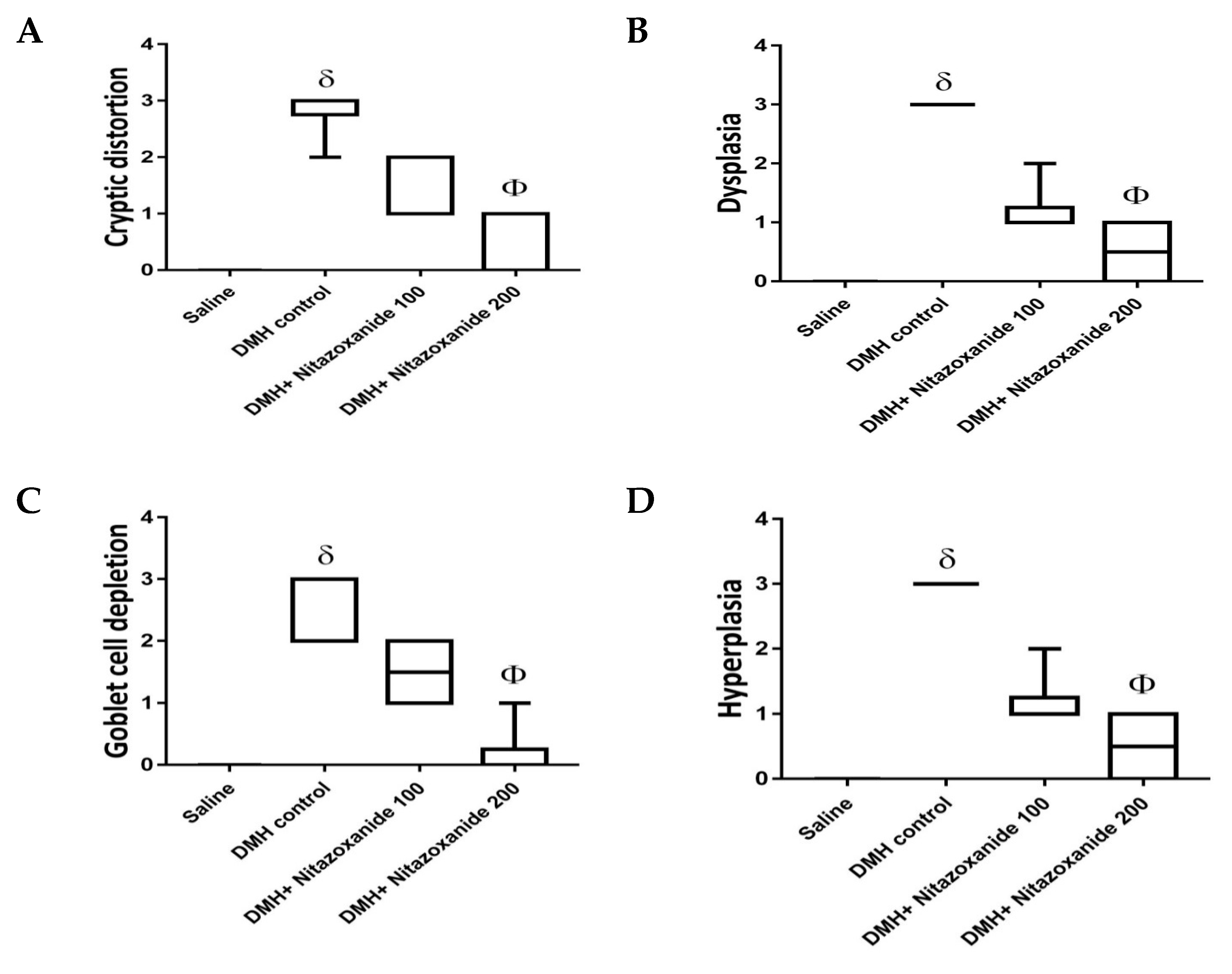
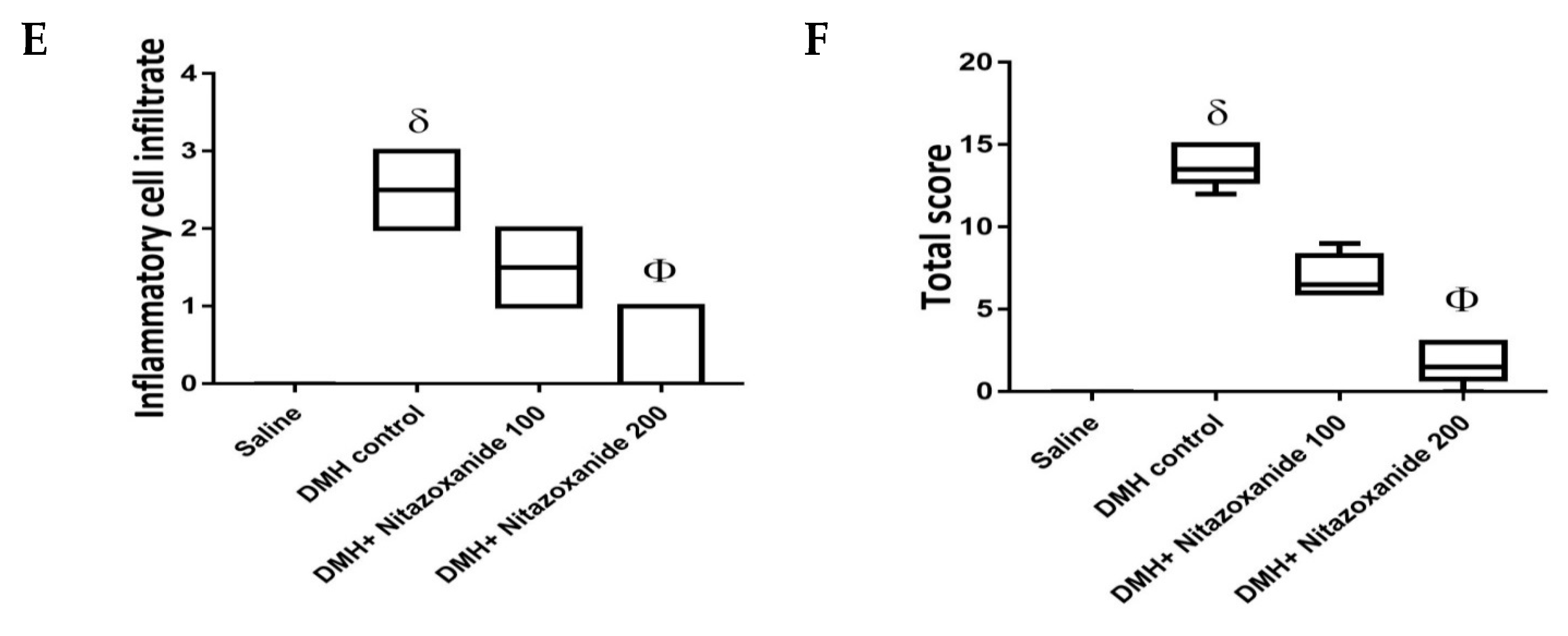
2.2.3. Effect of Nitazoxanide on Pathologic Picture in Mice with Colon Cancer
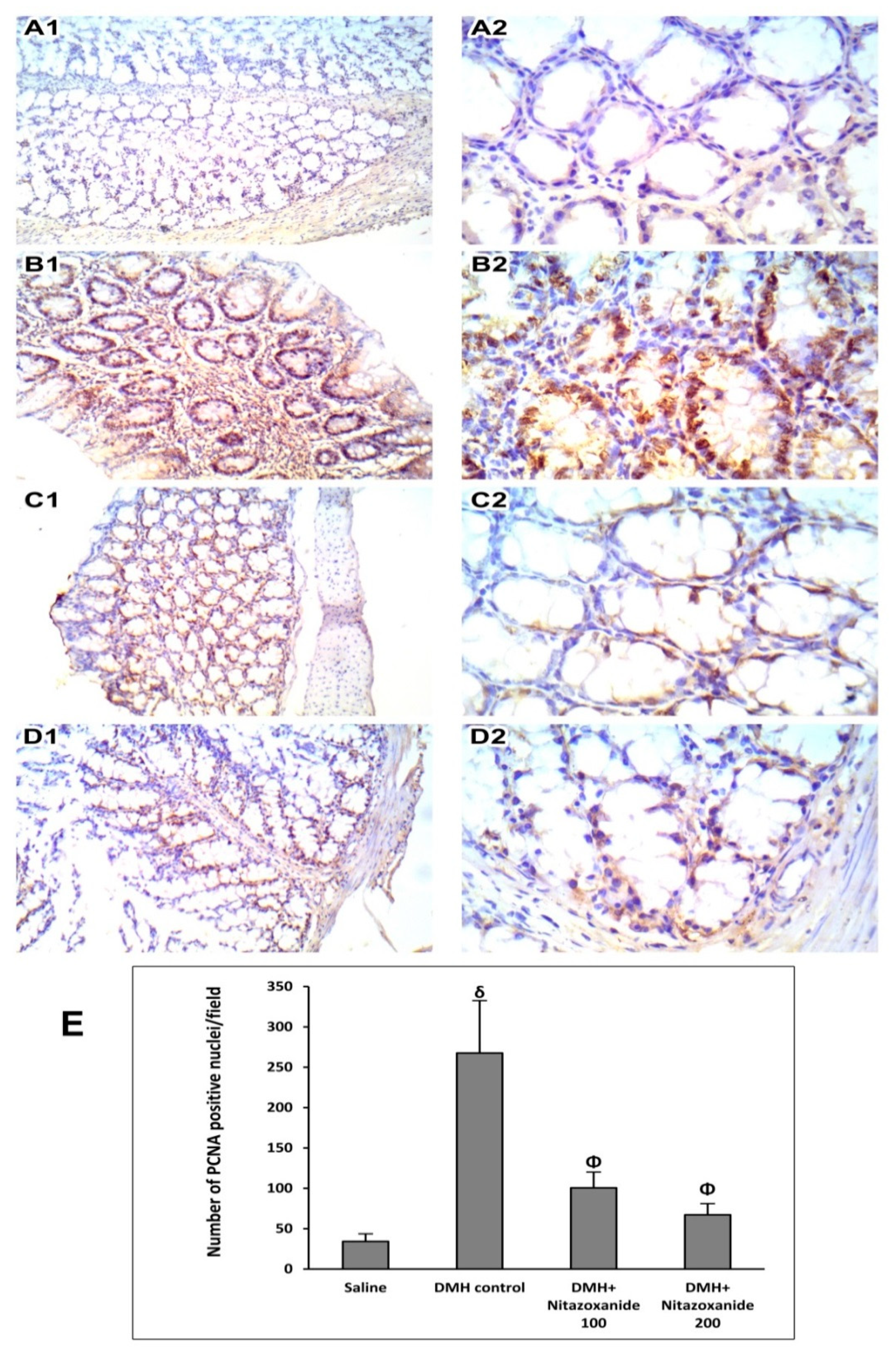
2.2.4. Effect of Nitazoxanide on Proliferating Cell Nuclear Antigen (PCNA) Immunoractivity in Mice with Colon Cancer
2.3. Experiment 3: In Silico Mechanism of Action Simulations
3. Discussion
4. Materials and Methods
4.1. Medications
4.2. Experiment 1: In Vitro Experiment
4.2.1. Cell Culturing and Cytotoxic Activity Using the MTT Assay
4.2.2. Investigation of Apoptosis in the Colon Cancer Cells
Annexin V/PI Staining and Cell-Cycle Analysi
4.2.3. Real Time-Polymerase Chain Reaction for the Selected Genes
4.3. Experiment 2: In Vivo Experiment
4.3.1. Ethics Statement
4.3.2. Animal Environment
4.3.3. Induction of Colonic Cancer and Experimental Groups
4.3.4. Signaling Pathway Enrichment Analysis
4.3.5. Western Blot Analysis for Wnt, β-Catenin and GSK-3β Proteins
4.3.6. Inspection of Hematoxylin and Eosin-Stained Colon Specimens
4.3.7. Immunohistochemistry for PCNA
4.4. Experiment 3: In Silico Molecular Docking Simulations for Nitazoxanide
4.5. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bray, F.; Ferlay, J.; Soerjomataram, I.; Siegel, R.L.; Torre, L.A.; Jemal, A. Global cancer statistics 2018: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J. Clin. 2018, 68, 394–424. [Google Scholar] [CrossRef] [Green Version]
- Terzić, J.; Grivennikov, S.; Karin, E.; Karin, M. Inflammation and Colon Cancer. Gastroenterology 2010, 138, 2101–2114. [Google Scholar] [CrossRef]
- Network, C.G.A. Comprehensive molecular characterization of human colon and rectal cancer. Nature 2012, 487, 330. [Google Scholar] [CrossRef] [Green Version]
- Polakis, P.; Hart, M.; Rubinfeld, B. Defects in the regulation of β-catenin in colorectal cancer. In Colon Cancer Prevention; Springer: Berlin/Heidelberg, Germany, 1999; pp. 23–32. [Google Scholar]
- Baldus, S.E.; Mönig, S.P.; Huxel, S.; Landsberg, S.; Hanisch, F.-G.; Engelmann, K.; Schneider, P.M.; Thiele, J.; Hölscher, A.H.; Dienes, H.P. MUC1 and Nuclear β-Catenin Are Coexpressed at the Invasion Front of Colorectal Carcinomas and Are Both Correlated with Tumor Prognosis. Clin. Cancer Res. 2004, 10, 2790–2796. [Google Scholar] [CrossRef] [Green Version]
- Miyamoto, S.; Endoh, Y.; Hasebe, T.; Ishii, G.; Kodama, K.; Goya, M.; Ono, M.; Saitoh, N.; Chiba, T.; Ochiai, A. Nuclear β-catenin accumulation as a prognostic factor in Dukes’ D human colorectal cancers. Oncol. Rep. 2004, 12, 245–251. [Google Scholar] [CrossRef] [PubMed]
- Polakis, P. The oncogenic activation of beta-catenin. Curr. Opin. Genet. Dev. 1999, 9, 15–21. [Google Scholar] [CrossRef]
- Manoukian, A.S.; Woodgett, J.R. Role of glycogen synthase kinase-3 in cancer: Regulation by Wnts and other signaling pathways. Adv. Cancer Res. 2002, 84, 203–229. [Google Scholar] [CrossRef] [PubMed]
- Jope, R.S.; Johnson, G.V. The glamour and gloom of glycogen synthase kinase-3. Trends Biochem. Sci. 2004, 29, 95–102. [Google Scholar] [CrossRef] [PubMed]
- Lin, J.; Ye, Q.; Wang, Y.; Wang, Y.; Zeng, Y. Wnt/β-catenin signaling regulates pathogenesis of human middle ear cholestea-toma. Int. J. Clin. Exp. Pathol. 2019, 12, 1154–1162. [Google Scholar]
- White, A.C., Jr. Nitazoxanide: A new broad spectrum antiparasitic agent. Expert Rev. Anti-Infect. Ther. 2004, 2, 43–49. [Google Scholar] [CrossRef] [PubMed]
- Darwish, W.M.; Bayoumi, N.A.; El-Kolaly, M.T. Laser-responsive liposome for selective tumor targeting of nitazoxanide nanoparticles. Eur. J. Pharm. Sci. 2018, 111, 526–533. [Google Scholar] [CrossRef]
- Wang, X.; Shen, C.; Liu, Z.; Peng, F.; Chen, X.; Yang, G.; Zhang, D.; Yin, Z.; Ma, J.; Zheng, Z.; et al. Nitazoxanide, an antiprotozoal drug, inhibits late-stage autophagy and promotes ING1-induced cell cycle arrest in glioblastoma. Cell Death Dis. 2018, 9, 1–15. [Google Scholar] [CrossRef]
- Di Santo, N.; Ehrisman, J. Research Perspective: Potential Role of Nitazoxanide in Ovarian Cancer Treatment. Old Drug, New Purpose? Cancers 2013, 5, 1163–1176. [Google Scholar] [CrossRef] [PubMed]
- Fan-Minogue, H.; Bodapati, S.; Solow-Cordero, D.; Fan, A.; Paulmurugan, R.; Massoud, T.F.; Felsher, D.W.; Gambhir, S.S. A c-Myc Activation Sensor-Based High-Throughput Drug Screening Identifies an Antineoplastic Effect of Nitazoxanide. Mol. Cancer Ther. 2013, 12, 1896–1905. [Google Scholar] [CrossRef] [Green Version]
- Di Santo, N.; Ehrisman, J. A functional perspective of nitazoxanide as a potential anticancer drug. Mutat. Res. Mol. Mech. Mutagen. 2014, 768, 16–21. [Google Scholar] [CrossRef] [PubMed]
- Saito-Diaz, K.; Chen, T.W.; Wang, X.; Thorne, C.A.; Wallace, H.A.; Page-McCaw, A.; Lee, E. The way Wnt works: Components and mechanism. Growth Factors 2012, 31, 1–31. [Google Scholar] [CrossRef] [PubMed]
- MacDonald, B.T.; Tamai, K.; He, X. Wnt/β-catenin signaling: Components, mechanisms, and diseases. Dev. Cell 2009, 17, 9–26. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Choi, H.-J.; Huber, A.H.; Weis, W.I. Thermodynamics of β-catenin-ligand interactions the roles of the n-and c-terminal tails in modulating binding affinity. J. Biol. Chem. 2006, 281, 1027–1038. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schlessinger, K.; Hall, A.; Tolwinski, N. Wnt signaling pathways meet Rho GTPases. Genes Dev. 2009, 23, 265–277. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lai, S.-L.; Chien, A.J.; Moon, R.T. Wnt/Fz signaling and the cytoskeleton: Potential roles in tumorigenesis. Cell Res. 2009, 19, 532–545. [Google Scholar] [CrossRef]
- Kühl, M.; Sheldahl, L.C.; Park, M.; Miller, J.R.; Moon, R.T. The wnt/ca2+ pathway: A new vertebrate wnt signaling pathway takes shape. Trends Genet. 2000, 16, 279–283. [Google Scholar] [CrossRef]
- Carpenter, A.C.; Rao, S.; Wells, J.M.; Campbell, K.; Lang, R.A. Generation of mice with a conditional null allele for Wntless. Genes 2010, 48, 554–558. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kohn, A.D.; Moon, R.T. Wnt and calcium signaling: β-Catenin-independent pathways. Cell Calcium 2005, 38, 439–446. [Google Scholar] [CrossRef] [PubMed]
- Anderson, V.R.; Curran, M.P. Nitazoxanide. Drugs 2007, 67, 1947–1967. [Google Scholar] [CrossRef]
- Du, W.; Hong, J.; Wang, Y.-C.; Zhang, Y.-J.; Wang, P.; Su, W.-Y.; Lin, Y.-W.; Lu, R.; Zou, W.-P.; Xiong, H.; et al. Inhibition of JAK2/STAT3 signalling induces colorectal cancer cell apoptosis via mitochondrial pathway. J. Cell. Mol. Med. 2012, 16, 1878–1888. [Google Scholar] [CrossRef] [PubMed]
- Abraha, A.M.; Ketema, E.B. Apoptotic pathways as a therapeutic target for colorectal cancer treatment. World J. Gastrointest. Oncol. 2016, 8, 583–591. [Google Scholar] [CrossRef] [Green Version]
- Müller, J.; Sidler, D.; Nachbur, U.; Wastling, J.; Brunner, T.; Hemphill, A. Thiazolides inhibit growth and induce glutathione-S-transferase Pi (GSTP1)-dependent cell death in human colon cancer cells. Int. J. Cancer 2008, 123, 1797–1806. [Google Scholar] [CrossRef]
- Hong, S.K.; Kim, H.J.; Song, C.S.; Choi, I.S.; Lee, J.B.; Park, S.Y. Nitazoxanide suppresses IL-6 production in LPS-stimulated mouse macrophages and TG-injected mice. Int. Immunopharmacol. 2012, 13, 23–27. [Google Scholar] [CrossRef]
- Qu, Y.; Olsen, J.R.; Yuan, X.; Cheng, P.F.; Levesque, M.P.; Brokstad, K.A.; Hoffman, P.S.; Oyan, A.M.; Zhang, W.; Kalland, K.-H.; et al. Small molecule promotes β-catenin citrullination and inhibits Wnt signaling in cancer. Nat. Chem. Biol. 2017, 14, 94–101. [Google Scholar] [CrossRef] [PubMed]
- Senkowski, W.; Zhang, X.; Olofsson, M.H.; Isacson, R.; Höglund, U.; Gustafsson, M.; Nygren, P.; Linder, S.; Larsson, R.; Fryknäs, M. Three-Dimensional Cell Culture-Based Screening Identifies the Anthelmintic Drug Nitazoxanide as a Candidate for Treatment of Colorectal Cancer. Mol. Cancer Ther. 2015, 14, 1504–1516. [Google Scholar] [CrossRef] [Green Version]
- Hadjihannas, M.V.; Brückner, M.; Jerchow, B.; Birchmeier, W.; Dietmaier, W.; Behrens, J. Aberrant wnt/β-catenin signaling can induce chromosomal instability in colon cancer. Proc. Natl. Acad. Sci. USA 2006, 103, 10747–10752. [Google Scholar] [CrossRef] [Green Version]
- Jansson, E.Å.; Are, A.; Greicius, G.; Kuo, I.-C.; Kelly, D.; Arulampalam, V.; Pettersson, S. The wnt/β-catenin signaling path-way targets pparγ activity in colon cancer cells. Proc. Natl. Acad. Sci. USA 2005, 102, 1460–1465. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- González-Sancho, J.M.; Aguilera, O.; García, J.M.; Pendás-Franco, N.; Peña, C.; Cal, S.; De Herreros, A.G.; Bonilla, F.; Muñoz, A. The Wnt antagonist DICKKOPF-1 gene is a downstream target of β-catenin/TCF and is downregulated in human colon cancer. Oncogene 2004, 24, 1098–1103. [Google Scholar] [CrossRef] [Green Version]
- Yu, J.; Yang, K.; Zheng, J.; Zhao, W.; Sun, X. Synergistic tumor inhibition of colon cancer cells by nitazoxanide and obeticholic acid, a farnesoid X receptor ligand. Cancer Gene Ther. 2020, 1–12. [Google Scholar] [CrossRef]
- Bertolini, F.; Sukhatme, V.P.; Bouche, G. Drug repurposing in oncology—Patient and health systems opportunities. Nat. Rev. Clin. Oncol. 2015, 12, 732–742. [Google Scholar] [CrossRef]
- Cai, W.; Chen, G.; Luo, Q.; Liu, J.; Guo, X.; Zhang, T.; Ma, F.; Yuan, L.; Li, B.; Cai, J. PMP22 Regulates Self-Renewal and Chemoresistance of Gastric Cancer Cells. Mol. Cancer Ther. 2017, 16, 1187–1198. [Google Scholar] [CrossRef] [Green Version]
- Liao, X.; Zhang, L.; Thrasher, J.B.; Du, J.; Li, B. Glycogen synthase kinase-3beta suppression eliminates tumor necrosis fac-tor-related apoptosis-inducing ligand resistance in prostate cancer. Mol. Cancer Ther. 2003, 2, 1215–1222. [Google Scholar] [PubMed]
- Mazor, M.; Kawano, Y.; Zhu, H.; Waxman, J.; Kypta, R.M. Inhibition of glycogen synthase kinase-3 represses androgen receptor activity and prostate cancer cell growth. Oncogene 2004, 23, 7882–7892. [Google Scholar] [CrossRef] [Green Version]
- Hoeflich, K.P.; Luo, J.; Rubie, E.A.; Tsao, M.S.; Jin, O.; Woodgett, J.R. Requirement for glycogen synthase kinase-3beta in cell survival and nf-kappab activation. Nature 2000, 406, 86–90. [Google Scholar] [CrossRef]
- Schwabe, R.F.; Brenner, D.A. Role of glycogen synthase kinase-3 in tnf-alpha-induced nf-kappab activation and apoptosis in hepatocytes. Am. J. Physiol. Gastrointest. Liver Physiol. 2002, 283, G204–G211. [Google Scholar] [CrossRef]
- Du, Q.; Geller, D.A. Cross-regulation between wnt and nf-κb signaling pathways. In Forum on Immunopathological Diseases and Therapeutics; Begel House Inc.: New York, NY, USA, 2010; Volume 1, pp. 155–181. [Google Scholar]
- Cohen, P.; Goedert, M. GSK3 inhibitors: Development and therapeutic potential. Nat. Rev. Drug Discov. 2004, 3, 479–487. [Google Scholar] [CrossRef]
- Meijer, L.; Flajolet, M.; Greengard, P. Pharmacological inhibitors of glycogen synthase kinase 3. Trends Pharmacol. Sci. 2004, 25, 471–480. [Google Scholar] [CrossRef] [PubMed]
- Shakoori, A.; Ougolkov, A.; Yu, Z.W.; Zhang, B.; Modarressi, M.H.; Billadeau, D.D.; Mai, M.; Takahashi, Y.; Minamoto, T. Deregulated GSK3β activity in colorectal cancer: Its association with tumor cell survival and proliferation. Biochem. Biophys. Res. Commun. 2005, 334, 1365–1373. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Freshney, R.I. Culture of Animal Cells: A Manual of Basic Technique and Specialized Applications; John Wiley & Sons: Hoboken, NJ, USA, 2005. [Google Scholar]
- Mosdam, T. Rapid colorimetric assay for cellular growth and survival: Application to proliferation and cytotoxic assay. J. Immunol. Methods 1983, 65, 55–63. [Google Scholar]
- Tantawy, E.S.; Amer, A.M.; Mohamed, E.K.; Alla, M.M.A.; Nafie, M.S. Synthesis, characterization of some pyrazine derivatives as anti-cancer agents: In vitro and in Silico approaches. J. Mol. Struct. 2020, 1210, 128013. [Google Scholar] [CrossRef]
- Sarhan, A.A.M.; Boraei, A.T.A.; Barakat, A.; Nafie, M.S. Discovery of hydrazide-based pyridazino[4,5-b]indole scaffold as a new phosphoinositide 3-kinase (PI3K) inhibitor for breast cancer therapy. RSC Adv. 2020, 10, 19534–19541. [Google Scholar] [CrossRef]
- Nafie, M.S.; Arafa, K.; Sedky, N.K.; Alakhdar, A.A.; Arafa, R.K. Triaryl dicationic DNA minor-groove binders with antioxidant activity display cytotoxicity and induce apoptosis in breast cancer. Chem. Interact. 2020, 324, 109087. [Google Scholar] [CrossRef]
- Gad, E.M.; Nafie, M.S.; Eltamany, E.H.; Hammad, M.S.A.G.; Barakat, A.; Boraei, A.T.A. Discovery of New Apoptosis-Inducing Agents for Breast Cancer Based on Ethyl 2-Amino-4,5,6,7-Tetra Hydrobenzo[b]Thiophene-3-Carboxylate: Synthesis, In Vitro, and In Vivo Activity Evaluation. Molecules 2020, 25, 2523. [Google Scholar] [CrossRef]
- Khodair, A.I.; Alsafi, M.A.; Nafie, M.S. Synthesis, molecular modeling and anti-cancer evaluation of a series of quinazoline derivatives. Carbohydr. Res. 2019, 486, 107832. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative pcr and the 2(-delta delta c(t)) method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Untereiner, A.A.; Pavlidou, A.; Druzhyna, N.; Papapetropoulos, A.; Hellmich, M.R.; Szabo, C. Drug resistance induces the upregulation of H2S-producing enzymes in HCT116 colon cancer cells. Biochem. Pharmacol. 2018, 149, 174–185. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Liu, H.; Yu, J.; Yu, H. Chemoresistance to doxorubicin induces epithelial-mesenchymal transition via upregulation of transforming growth factor β signaling in HCT116 colon cancer cells. Mol. Med. Rep. 2015, 12, 192–198. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bahr, H.I.; Ibrahiem, A.T.; Gabr, A.M.; Elbahaie, A.M.; Elmahdi, H.S.; Soliman, N.; Youssef, A.M.; El-Sherbiny, M.; Zaitone, S.A. Chemopreventive effect of α-hederin/carboplatin combination against experimental colon hyperplasia and impact on JNKsignaling. Toxicol. Mech. Methods 2021, 31, 138–149. [Google Scholar] [CrossRef] [PubMed]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compound | IC50 (µM) | |
|---|---|---|
| HCT-116 (ATCC® CCL-247™) Colon Cancer | FHC (ATCC® CRL-1831™) Normal Colon | |
| Nitazoxanide | 11.07 ± 0.89 | 48.4 ± 1.23 |
| 5-Flurouracil | 11.36 ± 0.76 | >50 |
| Docked Compound | Docking Energy (Kcal/mol) | Drug-Target Interactions | Interacting Moiety | ||||
|---|---|---|---|---|---|---|---|
| Nitazoxanide | −10.58 | 2 HB with Asn 290 | -C = O (as hydrogen bond acceptor) | ||||
| 5-Flurouracil | −8.31 | 1 HB with Asn 290 | -C = O (as hydrogen bond acceptor) | ||||
| HB: hydrogen bond, mol: mole. | |||||||
| A |  | B |  | ||||
| C |  | ||||||
| Primer | Sequence | Annealing Temperature |
|---|---|---|
| P53 | FOR: 5′-CTTTGAGGTGCGTGTTTGTG-3′ REV: 5′-GTGGTTTCTTCTTTGGCTGG-3′ | 57 °C |
| BCL-2 | FOR: 5′-GAGGATTGTGGCCTTCTTTG-3′ REV: 5′-ACAGTTCCACAAAGGCATCC0-3′ | 56 °C |
| BAX | FOR: 5′-TTTGCTTCAGGGTTTCATCC-3′ REV: 5′-CAGTTGAAGTTGCCGTCAGA-3′ | 55 °C |
| Casp-3 | FOR: 5′- TGGCCCTGAAATACGAAGTC-3′ REV: 5′- GGCAGTAGTCGACTCTGAAG -3′ | 56 °C |
| Casp-8 | FOR: 5′- AATGTTGGAGGAAAGCAAT -3′ REV: 5′- CATAGTCGTTGATTATCTTCAGC -3′ | 51 °C |
| Casp-9 | FOR: 5′- CGAACTAACAGGCAAGCAGC -3′ REV: 5′- ACCTCACCAAATCCTCCAGAAC -3′ | 58 °C |
| β-actin | FOR: 5′-GCACTCTTCCAGCCTTCCTTCC-3′ REV: 5′-GAGCCGCCGATCCACACG-3′ | 60 °C |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Abd El-Fadeal, N.M.; Nafie, M.S.; K. El-kherbetawy, M.; El-mistekawy, A.; Mohammad, H.M.F.; Elbahaie, A.M.; Hashish, A.A.; Alomar, S.Y.; Aloyouni, S.Y.; El-dosoky, M.; et al. Antitumor Activity of Nitazoxanide against Colon Cancers: Molecular Docking and Experimental Studies Based on Wnt/β-Catenin Signaling Inhibition. Int. J. Mol. Sci. 2021, 22, 5213. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22105213
Abd El-Fadeal NM, Nafie MS, K. El-kherbetawy M, El-mistekawy A, Mohammad HMF, Elbahaie AM, Hashish AA, Alomar SY, Aloyouni SY, El-dosoky M, et al. Antitumor Activity of Nitazoxanide against Colon Cancers: Molecular Docking and Experimental Studies Based on Wnt/β-Catenin Signaling Inhibition. International Journal of Molecular Sciences. 2021; 22(10):5213. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22105213
Chicago/Turabian StyleAbd El-Fadeal, Noha M., Mohamed S. Nafie, Mohammed K. El-kherbetawy, Amr El-mistekawy, Hala M. F. Mohammad, Alaaeldeen M. Elbahaie, Abdullah A. Hashish, Suliman Y. Alomar, Sheka Yagub Aloyouni, Mohamed El-dosoky, and et al. 2021. "Antitumor Activity of Nitazoxanide against Colon Cancers: Molecular Docking and Experimental Studies Based on Wnt/β-Catenin Signaling Inhibition" International Journal of Molecular Sciences 22, no. 10: 5213. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22105213