
The Role of Fatty Acids in Ceramide Pathways and Their Influence on Hypothalamic Regulation of Energy Balance: A Systematic Review

 ,
,  and
and
Abstract
:1. Background
2. Materials and Methods
2.1. Data Sources Search Strategy
2.2. Eligibility Criteria
2.3. Data Extraction
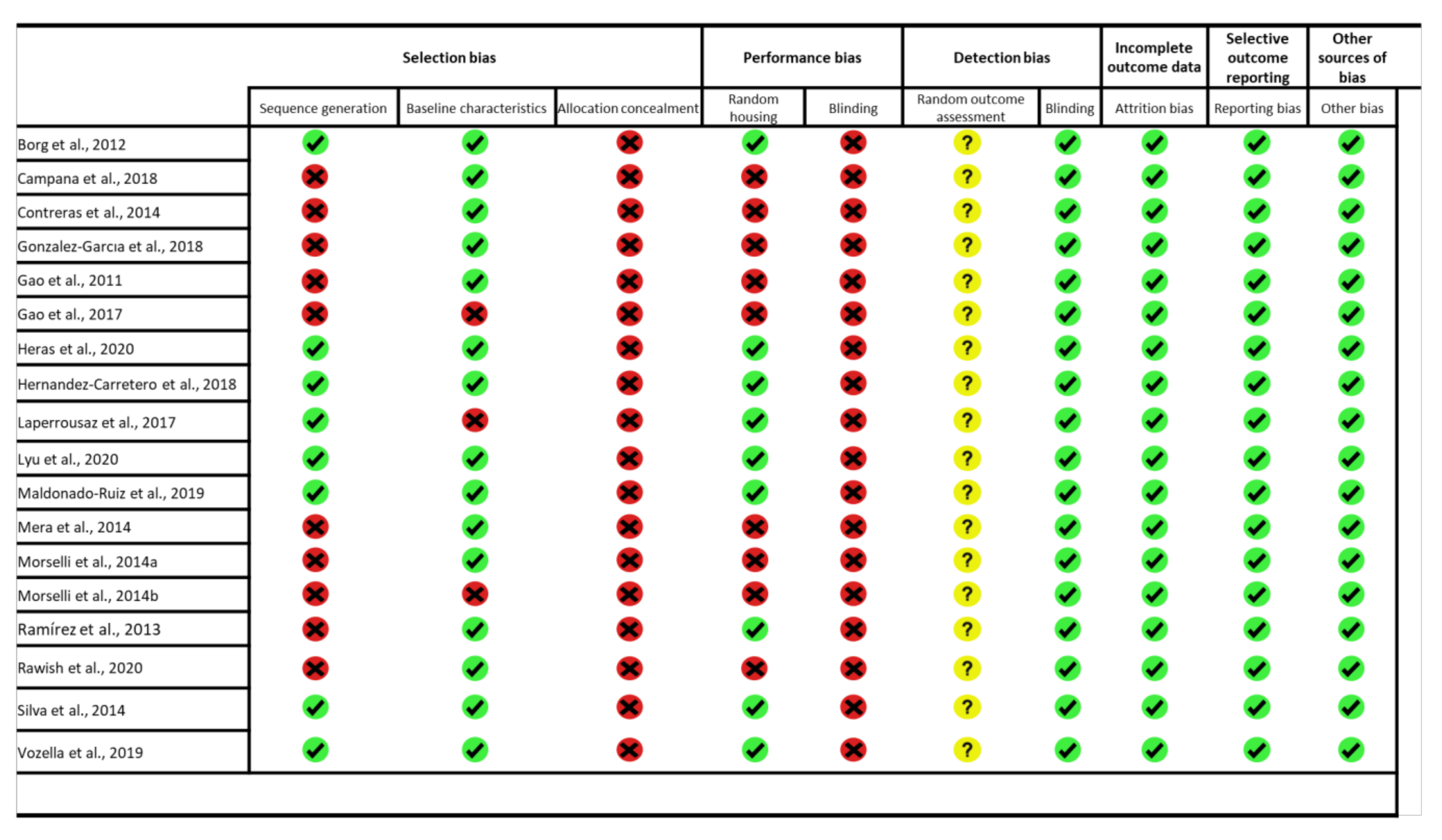
2.4. Assessment of Methodology Quality
3. Results
4. Methodological Approach to Assess Sphingolipids
5. Additional in Vitro Methodological Approach
6. Main Findings Regarding In Vitro Sphingolipid Levels and Their Role in the Hypothalamus
7. Additional In Vivo Methodological Approach
8. Main Findings Regarding In Vivo Sphingolipid Levels and Their Role in the Hypothalamus
9. Discussion
10. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Khaodhiar, L.; McCowen, K.C.; Blackburn, G.L. Obesity and Its Comorbid Conditions. Clin. Cornerstone 1999, 2, 17–31. [Google Scholar] [CrossRef]
- Ortega, F.B.; Lavie, C.J. Introduction and Update on Obesity and Cardiovascular Diseases 2018. Prog. Cardiovasc. Dis. 2018, 61, 87–88. [Google Scholar] [CrossRef]
- De Sales-Peres, S.H.C.; Azevedo-Silva, L.J.; Bonato, R.C.S.; Sales-Peres, M.D.C.; da Pinto, A.C.S.; Junior, J.F.S. Coronavirus (SARS-CoV-2) and the Risk of Obesity for Critically Illness and ICU Admitted: Meta-Analysis of the Epidemiological Evidence. Obes. Res. Clin. Pract. 2020, 14, 389–397. [Google Scholar] [CrossRef]
- Schwartz, M.W.; Woods, S.C.; Porte, D.; Seeley, R.J.; Baskin, D.G. Central Nervous System Control of Food Intake. Nature 2000, 404, 661–671. [Google Scholar] [CrossRef] [PubMed]
- Velloso, L.A. The Brain Is the Conductor: Diet-Induced Inflammation Overlapping Physiological Control of Body Mass and Metabolism. Arq. Bras. Endocrinol. Metabol. 2009, 53, 151–158. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yaqoob, P.; Shaikh, S.R. The Nutritional and Clinical Significance of Lipid Rafts. Curr. Opin. Clin. Nutr. Metab. Care 2010, 13, 156–166. [Google Scholar] [CrossRef] [PubMed]
- Milanski, M.; Degasperi, G.; Coope, A.; Morari, J.; Denis, R.; Cintra, D.E.; Tsukumo, D.M.L.; Anhe, G.; Amaral, M.E.; Takahashi, H.K.; et al. Saturated Fatty Acids Produce an Inflammatory Response Predominantly through the Activation of TLR4 Signaling in Hypothalamus: Implications for the Pathogenesis of Obesity. J. Neurosci. 2009, 29, 359–370. [Google Scholar] [CrossRef]
- Posey, K.A.; Clegg, D.J.; Printz, R.L.; Byun, J.; Morton, G.J.; Vivekanandan-Giri, A.; Pennathur, S.; Baskin, D.G.; Heinecke, J.W.; Woods, S.C.; et al. Hypothalamic Proinflammatory Lipid Accumulation, Inflammation, and Insulin Resistance in Rats Fed a High-Fat Diet. Am. J. Physiol. Endocrinol. Metab. 2009, 296, 1–21. [Google Scholar] [CrossRef] [PubMed]
- Valdearcos, M.; Xu, A.W.; Koliwad, S.K. Hypothalamic Inflammation in the Control of Metabolic Function. Annu. Rev. Physiol. 2015, 77, 131–160. [Google Scholar] [CrossRef]
- Thaler, J.P.; Yi, C.-X.; Schur, E.A.; Guyenet, S.J.; Hwang, B.H.; Dietrich, M.O.; Zhao, X.; Sarruf, D.A.; Izgur, V.; Maravilla, K.R.; et al. Obesity Is Associated with Hypothalamic Injury in Rodents and Humans. J. Clin. Investig. 2012, 122, 119–120. [Google Scholar] [CrossRef]
- Portovedo, M.; Reginato, A.; Miyamoto, J.; Simino, L.A.; Hakim, M.P.; Campana, M.; Leal, R.F.; Ignácio-Souza, L.M.; Torsoni, M.A.; Magnan, C.; et al. Lipid Excess Affects Chaperone-Mediated Autophagy in Hypothalamus. Biochimie 2020, 176, 110–116. [Google Scholar] [CrossRef]
- Reginato, A.; Siqueira, B.P.; Miyamoto, J.É.; Portovedo, M.; de Costa, S.O.; de Fante, T.; Rodrigues, H.G.; Ignácio-Souza, L.M.; Torsoni, M.A.; Torsoni, A.S.; et al. Acute Effects of Fatty Acids on Autophagy in NPY Neurones. J. Neuroendocrinol. 2020, 32, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Ye, W.; Ramos, E.H.; Wong, B.C.; Belsham, D.D. Beneficial Effects of Metformin and/or Salicylate on Palmitate- or TNFα-Induced Neuroinflammatory Marker and Neuropeptide Gene Regulation in Immortalized NPY/AgRP Neurons. PLoS ONE 2016, 11, e0166973. [Google Scholar] [CrossRef]
- Dalvi, P.S.; Chalmers, J.A.; Luo, V.; Han, D.Y.; Wellhauser, L.; Liu, Y.; Tran, D.Q.; Castel, J.; Luquet, S.; Wheeler, M.B.; et al. High Fat Induces Acute and Chronic Inflammation in the Hypothalamus: Effect of High-Fat Diet, Palmitate and TNF-α on Appetite-Regulating NPY Neurons. Int. J. Obes. 2017, 41, 149–158. [Google Scholar] [CrossRef] [PubMed]
- Tse, E.K.; Salehi, A.; Clemenzi, M.N.; Belsham, D.D. Role of the Saturated Fatty Acid Palmitate in the Interconnected Hypothalamic Control of Energy Homeostasis and Biological Rhythms. Am. J. Physiol. Endocrinol. Metab. 2018, 315, E133–E140. [Google Scholar] [CrossRef]
- Sergi, D.; Williams, L.M. Potential Relationship between Dietary Long-Chain Saturated Fatty Acids and Hypothalamic Dysfunction in Obesity. Nutr. Rev. 2020, 78, 261–277. [Google Scholar] [CrossRef] [PubMed]
- Kang, S.C.; Kim, B.R.; Lee, S.Y.; Park, T.S. Sphingolipid Metabolism and Obesity-Induced Inflammation. Front. Endocrinol. 2013, 4, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Dobrzyn, A.; Janikiewicz, J.; Tracz-Gaszewska, Z.; Filip, A.; Dobosz, A.M.; Krogulec, E.; Dobrzyn, P. Sphingolipid Mediators of Cell Signaling and Metabolism. Lipid Signal. Metab. 2020, 385–411. [Google Scholar] [CrossRef]
- Futerman, A.H.; Hannun, Y.A. The Complex Life of Simple Sphingolipids. EMBO Rep. 2004, 5, 777–782. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hannun, Y.A.; Obeid, L.M. Sphingolipids and Their Metabolism in Physiology and Disease. Nat. Rev. Mol. Cell Biol. 2018, 19, 175–191. [Google Scholar] [CrossRef]
- Morad, S.A.F.; Cabot, M.C. Ceramide-Orchestrated Signalling in Cancer Cells. Nat. Rev. Cancer 2013, 13, 51–65. [Google Scholar] [CrossRef]
- Guitton, J.; Bandet, C.L.; Mariko, M.L.; Tan-Chen, S.; Bourron, O.; Benomar, Y.; Hajduch, E.; le Stunff, H. Sphingosine-1-Phosphate Metabolism in the Regulation of Obesity/Type 2 Diabetes. Cells 2020, 9, 1682. [Google Scholar] [CrossRef] [PubMed]
- Sugiura, M.; Kono, K.; Liu, H.; Shimizugawa, T.; Minekura, H.; Spiegel, S.; Kohama, T. Ceramide Kinase, a Novel Lipid Kinase: Molecular Cloning and Functional Characterization. J. Biol. Chem. 2002, 277, 23294–23300. [Google Scholar] [CrossRef] [Green Version]
- Hoeferlin, L.A.; Wijesinghe, D.S.; Chalfant, C.E. The Role of Ceramide-1-Phosphate in Biological Functions. Handb. Exp. Pharmacol. 2013, 215, 153–166. [Google Scholar] [CrossRef] [Green Version]
- Kitatani, K.; Idkowiak-Baldys, J.; Hannun, Y.A. The Sphingolipid Salvage Pathway in Ceramide Metabolism and Signaling. Cell. Signal. 2008, 20, 1010–1018. [Google Scholar] [CrossRef] [Green Version]
- Chaurasia, B.; Summers, S.A. Ceramides–Lipotoxic Inducers of Metabolic Disorders. Trends Endocrinol. Metab. 2015, 26, 538–550. [Google Scholar] [CrossRef]
- Kim, M.Y.; Linardic, C.; Obeid, L.; Hannun, Y. Identification of Sphingomyelin Turnover as an Effector Mechanism for the Action of Tumor Necrosis Factor α and γ-Interferon. Specific Role in Cell Differentiation. J. Biol. Chem. 1991, 266, 484–489. [Google Scholar] [CrossRef]
- Cowart, L.A.; Okamoto, Y.; Pinto, F.R.; Gandy, J.L.; Almeida, J.S.; Hannun, Y.A. Roles for Sphingolipid Biosynthesis in Mediation of Specific Programs of the Heat Stress Response Determined through Gene Expression Profiling. J. Biol. Chem. 2003, 278, 30328–30338. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brenner, B.; Ferlinz, K.; Grassmé, H.; Weller, M.; Koppenhoefer, U.; Dichgans, J.; Sandhoff, K.; Lang, F.; Gulbins, E. Fas/CD95/Apo-I Activates the Acidic Sphingomyelinase via Caspases. Cell Death Differ. 1998, 5, 29–37. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goldkorn, T.; Balaban, N.; Shannon, M.; Chea, V.; Matsukuma, K.; Gilchrist, D.; Wang, H.; Chan, C. H2O2 Acts on Cellular Membranes to Generate Ceramide Signaling and Initiate Apoptosis in Tracheobronchial Epithelial Cells. J. Cell Sci. 1998, 111, 3209–3220. [Google Scholar] [CrossRef]
- Memon, R.A.; Holleran, W.M.; Moser, A.H.; Seki, T.; Uchida, Y.; Fuller, J.; Shigenaga, J.K.; Grunfeld, C.; Feingold, K.R. Endotoxin and Cytokines Increase Hepatic Sphingolipid Biosynthesis and Produce Lipoproteins Enriched in Ceramides and Sphingomyelin. Arterioscler. Thromb. Vasc. Biol. 1998, 18, 1257–1265. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Samad, F.; Hester, K.D.; Yang, G.; Hannun, Y.A.; Bielawski, J. Altered Adipose and Plasma Sphingolipid Metabolism in Obesity: A Potential Mechanism for Cardiovascular and Metabolic Risk. Diabetes 2006, 55, 2579–2587. [Google Scholar] [CrossRef] [Green Version]
- Schilling, J.D.; Machkovech, H.M.; He, L.; Sidhu, R.; Fujiwara, H.; Weber, K.; Ory, D.S.; Schaffer, J.E. Palmitate and Lipopolysaccharide Trigger Synergistic Ceramide Production in Primary Macrophages. J. Biol. Chem. 2013, 288, 2923–2932. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Truet, A.P.; Snyderman, R.; Murray, J.J. Stimulation of Phosphorylcholine Turnover and Diacylglycerol Production in Human Polymorphonuclear Leukocytes. Novel Assay for Phosphorylcholine. Biochem. J. 1989, 260, 909–913. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goñi, F.M.; Alonso, A. Sphingomyelinases: Enzymology and Membrane Activity. FEBS Lett. 2002, 531, 38–46. [Google Scholar] [CrossRef] [Green Version]
- Andrieu-Abadie, N.; Levade, T. Sphingomyelin Hydrolysis during Apoptosis. Biochim. Biophys. Acta Mol. Cell Biol. Lipids 2002, 1585, 126–134. [Google Scholar] [CrossRef]
- Mullen, T.D.; Hannun, Y.A.; Obeid, L.M. Ceramide Synthases at the Centre of Sphingolipid Metabolism and Biology. Biochem. J. 2012, 441, 789–802. [Google Scholar] [CrossRef] [Green Version]
- Levy, M.; Futerman, A.H. Mammalian Ceramide Synthases. Iubmb Life 2010, 62. [Google Scholar] [CrossRef]
- Park, M.; Kaddai, V.; Ching, J.; Fridianto, K.T.; Sieli, R.J.; Sugii, S.; Summers, S.A. A Role for Ceramides, but Not Sphingomyelins, as Antagonists of Insulin Signaling and Mitochondrial Metabolism in C2C12 Myotubes. J. Biol. Chem. 2016. [Google Scholar] [CrossRef] [Green Version]
- Turpin-Nolan, S.M.; Hammerschmidt, P.; Chen, W.; Jais, A.; Timper, K.; Awazawa, M.; Brodesser, S.; Brüning, J.C. CerS1-Derived C18:0 Ceramide in Skeletal Muscle Promotes Obesity-Induced Insulin Resistance. Cell Rep. 2019, 26, 1–10.e7. [Google Scholar] [CrossRef] [Green Version]
- Turpin, S.M.; Nicholls, H.T.; Willmes, D.M.; Mourier, A.; Brodesser, S.; Wunderlich, C.M.; Mauer, J.; Xu, E.; Hammerschmidt, P.; Brönneke, H.S.; et al. Obesity-Induced CerS6-Dependent C16:0 Ceramide Production Promotes Weight Gain and Glucose Intolerance. Cell Metab. 2014, 20, 678–686. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, J.-W.; Park, W.-J.; Kuperman, Y.; Boura-Halfon, S.; Pewzner-Jung, Y.; Futerman, A.H. Ablation of Very Long Acyl Chain Sphingolipids Causes Hepatic Insulin Resistance in Mice Due to Altered Detergent-Resistant Membranes. Hepatology 2013, 57, 525–532. [Google Scholar] [CrossRef] [PubMed]
- Raichur, S.; Wang, S.T.; Chan, P.W.; Li, Y.; Ching, J.; Chaurasia, B.; Dogra, S.; Öhman, M.K.; Takeda, K.; Sugii, S.; et al. CerS2 Haploinsufficiency Inhibits β-Oxidation and Confers Susceptibility to Diet-Induced Steatohepatitis and Insulin Resistance. Cell Metab. 2014, 20, 687–695. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, I.L.; Chou, H.L.; Lee, J.C.; Chen, F.W.; Fong, Y.; Chang, W.C.; Huang, H.W.; Wu, C.Y.; Chang, W.T.; Wang, H.M.D.; et al. The Antiproliferative Effect of C2-Ceramide on Lung Cancer Cells through Apoptosis by Inhibiting Akt and NFκB. Cancer Cell Int. 2014, 14. [Google Scholar] [CrossRef] [Green Version]
- Hou, Q.; Jin, J.; Zhou, H.; Novgorodov, S.A.; Bielawska, A.; Szulc, Z.M.; Hannun, Y.A.; Obeid, L.M.; Hsu, Y. te Mitochondrially Targeted Ceramides Preferentially Promote Autophagy, Retard Cell Growth, and Induce Apoptosis. J. Lipid Res. 2011, 52, 278–288. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mullen, T.D.; Obeid, L.M. Ceramide and Apoptosis: Exploring the Enigmatic Connections between Sphingolipid Metabolism and Programmed Cell Death. Anti Cancer Agents Med. Chem. 2012, 12, 340–363. [Google Scholar] [CrossRef]
- Ogretmen, B.; Hannun, Y.A. Biologically Active Sphingolipids in Cancer Pathogenesis and Treatment. Nat. Rev. Cancer 2004, 4, 604–616. [Google Scholar] [CrossRef]
- Reforgiato, M.R.; Milano, G.; Fabriàs, G.; Casas, J.; Gasco, P.; Paroni, R.; Samaja, M.; Ghidoni, R.; Caretti, A.; Signorelli, P. Inhibition of Ceramide de Novo Synthesis as a Postischemic Strategy to Reduce Myocardial Reperfusion Injury. Basic Res. Cardiol. 2016, 111. [Google Scholar] [CrossRef]
- Ji, R.; Akashi, H.; Drosatos, K.; Liao, X.; Jiang, H.; Kennel, P.J.; Brunjes, D.L.; Castillero, E.; Zhang, X.; Deng, L.Y.; et al. Increased de Novo Ceramide Synthesis and Accumulation in Failing Myocardium. JCI Insight 2017, 2, 1–19. [Google Scholar] [CrossRef]
- Santos-Gallego, C.G.; Vahl, T.P.; Goliasch, G.; Picatoste, B.; Arias, T.; Ishikawa, K.; Njerve, I.U.; Sanz, J.; Narula, J.; Sengupta, P.P.; et al. Sphingosine-1-Phosphate Receptor Agonist Fingolimod Increases Myocardial Salvage and Decreases Adverse Postinfarction Left Ventricular Remodeling in a Porcine Model of Ischemia/Reperfusion. Circulation 2016, 133, 954–966. [Google Scholar] [CrossRef]
- Contreras, C.; González-García, I.; Martínez-Sánchez, N.; Seoane-Collazo, P.; Jacas, J.; Morgan, D.A.; Serra, D.; Gallego, R.; Gonzalez, F.; Casals, N.; et al. Central Ceramide-Induced Hypothalamic Lipotoxicity and ER Stress Regulate Energy Balance. Cell Rep. 2014, 9, 366–377. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Picard, A.; Rouch, C.; Kassis, N.; Moullé, V.S.; Croizier, S.; Denis, R.G.; Castel, J.; Coant, N.; Davis, K.; Clegg, D.J.; et al. Hippocampal Lipoprotein Lipase Regulates Energy Balance in Rodents. Mol. Metab. 2014, 3, 167–176. [Google Scholar] [CrossRef]
- Cruciani-Guglielmacci, C.; López, M.; Campana, M.; le Stunff, H. Brain Ceramide Metabolism in the Control of Energy Balance. Front. Physiol. 2017, 8, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Saba, J.D.; Hla, T. Point-Counterpoint of Sphingosine 1-Phosphate Metabolism. Circ. Res. 2004, 94, 724–734. [Google Scholar] [CrossRef]
- Baumruker, T.; Bornancin, F.; Billich, A. The Role of Sphingosine and Ceramide Kinases in Inflammatory Responses. Immunol. Lett. 2005, 96, 175–185. [Google Scholar] [CrossRef] [PubMed]
- Hadas, Y.; Vincek, A.S.; Youssef, E.; Żak, M.M.; Chepurko, E.; Sultana, N.; Sharkar, M.T.K.; Guo, N.; Komargodski, R.; Kurian, A.A.; et al. Altering Sphingolipid Metabolism Attenuates Cell Death and Inflammatory Response after Myocardial Infarction. Circulation 2020, 916–930. [Google Scholar] [CrossRef]
- Silva, V.R.R.; Micheletti, T.O.; Pimentel, G.D.; Katashima, C.K.; Lenhare, L.; Morari, J.; Mendes, M.C.S.; Razolli, D.S.; Rocha, G.Z.; de Souza, C.T.; et al. Hypothalamic S1P/S1PR1 Axis Controls Energy Homeostasis. Nat. Commun. 2014, 5. [Google Scholar] [CrossRef] [Green Version]
- Vozella, V.; Realini, N.; Misto, A.; Piomelli, D. Feeding Stimulates Sphingosine-1-Phosphate Mobilization in Mouse Hypothalamus. Int. J. Mol. Sci. 2019, 20, 4008. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Chen, K.; Sloan, S.A.; Bennett, M.L.; Scholze, A.R.; O’Keeffe, S.; Phatnani, H.P.; Guarnieri, P.; Caneda, C.; Ruderisch, N.; et al. An RNA-Sequencing Transcriptome and Splicing Database of Glia, Neurons, and Vascular Cells of the Cerebral Cortex. J. Neurosci. 2014, 34, 11929–11947. [Google Scholar] [CrossRef] [PubMed]
- Hooijmans, C.R.; Rovers, M.M.; de Vries, R.B.; Leenaars, M.; Ritskes-Hoitinga, M.; Langendam, M.W. SYRCLE’s Risk of Bias Tool for Animal Studies. BMC Med. Res. Methodol. 2014, 14, 1–9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Golbach, L.A.; Portelli, L.A.; Savelkoul, H.F.J.; Terwel, S.R.; Kuster, N.; de Vries, R.B.M.; Verburg-van Kemenade, B.M.L. Calcium Homeostasis and Low-Frequency Magnetic and Electric Field Exposure: A Systematic Review and Meta-Analysis of in Vitro Studies. Environ. Int. 2016, 92–93, 695–706. [Google Scholar] [CrossRef] [PubMed]
- AlShwaimi, E.; Bogari, D.; Ajaj, R.; Al-Shahrani, S.; Almas, K.; Majeed, A. In Vitro Antimicrobial Effectiveness of Root Canal Sealers against Enterococcus Faecalis: A Systematic Review. J. Endod. 2016, 42, 1588–1597. [Google Scholar] [CrossRef] [PubMed]
- Campana, M.; Bellini, L.; Rouch, C.; Rachdi, L.; Coant, N.; Butin, N.; Bandet, C.L.; Philippe, E.; Meneyrol, K.; Kassis, N.; et al. Inhibition of Central de Novo Ceramide Synthesis Restores Insulin Signaling in Hypothalamus and Enhances β-Cell Function of Obese Zucker Rats. Mol. Metab. 2018, 8, 23–36. [Google Scholar] [CrossRef] [PubMed]
- Laperrousaz, E.; Moullé, V.S.; Denis, R.G.; Kassis, N.; Berland, C.; Colsch, B.; Fioramonti, X.; Philippe, E.; Lacombe, A.; Vanacker, C.; et al. Lipoprotein Lipase in Hypothalamus Is a Key Regulator of Body Weight Gain and Glucose Homeostasis in Mice. Diabetologia 2017, 60, 1314–1324. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- González-García, I.; Contreras, C.; Estévez-Salguero, Á.; Ruíz-Pino, F.; Colsh, B.; Pensado, I.; Liñares-Pose, L.; Rial-Pensado, E.; Martínez de Morentin, P.B.; Fernø, J.; et al. Estradiol Regulates Energy Balance by Ameliorating Hypothalamic Ceramide-Induced ER Stress. Cell Rep. 2018, 25, 413–423.e5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gao, Y.; Layritz, C.; Legutko, B.; Eichmann, T.O.; Laperrousaz, E.; Moullé, V.S.; Cruciani-Guglielmacci, C.; Magnan, C.; Luquet, S.; Woods, S.C.; et al. Disruption of Lipid Uptake in Astroglia Exacerbates Diet-Induced Obesity. Diabetes 2017, 66, 2555–2563. [Google Scholar] [CrossRef] [Green Version]
- Morselli, E.; Criollo, A.; Rodriguez-Navas, C.; Clegg, D.J. Chronic High Fat Diet Consumption Impairs Metabolic Health of Male Mice. Inflamm. Cell Sinal. 2014, 1, 1–15. [Google Scholar] [CrossRef]
- Morselli, E.; Fuente-Martin, E.; Finan, B.; Kim, M.; Frank, A.; Garcia-Caceres, C.; Navas, C.R.; Gordillo, R.; Neinast, M.; Kalainayakan, S.P.; et al. Hypothalamic PGC-1α Protects against High-Fat Diet Exposure by Regulating ERα. Cell Rep. 2014, 9, 633–645. [Google Scholar] [CrossRef] [Green Version]
- Mera, P.; Mir, J.F.; Fabriàs, G.; Casas, J.; Costa, A.S.H.; Malandrino, M.I.; Fernández-López, J.A.; Remesar, X.; Gao, S.; Chohnan, S.; et al. Long-Term Increased Carnitine Palmitoyltransferase 1A Expression in Ventromedial Hypotalamus Causes Hyperphagia and Alters the Hypothalamic Lipidomic Profile. PLoS ONE 2014, 9, e97195. [Google Scholar] [CrossRef] [Green Version]
- McFadden, J.W.; Aja, S.; Li, Q.; Bandaru, V.V.R.; Kim, E.K.; Haughey, N.J.; Kuhajda, F.P.; Ronnett, G.V. Increasing Fatty Acid Oxidation Remodels the Hypothalamic Neurometabolome to Mitigate Stress and Inflammation. PLoS ONE 2014, 9, e115642. [Google Scholar] [CrossRef]
- Gao, S.; Zhu, G.; Gao, X.; Wu, D.; Carrasco, P.; Casals, N.; Hegardt, F.G.; Moran, T.H.; Lopaschuk, G.D. Important Roles of Brain-Specific Carnitine Palmitoyltransferase and Ceramide Metabolism in Leptin Hypothalamic Control of Feeding. Proc. Natl. Acad. Sci. USA 2011, 108, 9691–9696. [Google Scholar] [CrossRef] [Green Version]
- Borg, M.L.; Omran, S.F.; Weir, J.; Meikle, P.J.; Watt, M.J. Consumption of a High-Fat Diet, but Not Regular Endurance Exercise Training, Regulates Hypothalamic Lipid Accumulation in Mice. J. Physiol. 2012, 590, 4377–4389. [Google Scholar] [CrossRef] [Green Version]
- Ramírez, S.; Martins, L.; Jacas, J.; Carrasco, P.; Pozo, M.; Clotet, J.; Serra, D.; Hegardt, F.G.; Diéguez, C.; López, M.; et al. Hypothalamic Ceramide Levels Regulated by Cpt1c Mediate the Orexigenic Effect of Ghrelin. Diabetes 2013, 62, 2329–2337. [Google Scholar] [CrossRef] [Green Version]
- Davis, C.N.; Tabarean, I.; Gaidarova, S.; Behrens, M.M.; Bartfai, T. IL-1β Induces a MyD88-Dependent and Ceramide-Mediated Activation of Src in Anterior Hypothalamic Neurons. J. Neurochem. 2006, 98, 1379–1389. [Google Scholar] [CrossRef]
- Sortino, M.A.; Condorelli, F.; Vancheri, C.; Canonico, P.L. Tumor Necrosis Factor-α Induces Apoptosis in Immortalized Hypothalamic Neurons: Involvement of Ceramide-Generating Pathways. Endocrinology 1999, 140, 4841–4849. [Google Scholar] [CrossRef] [PubMed]
- Rawish, E.; Nickel, L.; Schuster, F.; Stölting, I.; Frydrychowicz, A.; Saar, K.; Hübner, N.; Othman, A.; Kuerschner, L.; Raasch, W. Telmisartan Prevents Development of Obesity and Normalizes Hypothalamic Lipid Droplets. J. Endocrinol. 2020, 244, 95–110. [Google Scholar] [CrossRef] [PubMed]
- Maldonado-Ruiz, R.; Cárdenas-Tueme, M.; Montalvo-Martínez, L.; Vidaltamayo, R.; Garza-Ocañas, L.; Reséndez-Perez, D.; Camacho, A. Priming of Hypothalamic Ghrelin Signaling and Microglia Activation Exacerbate Feeding in Rats’ Offspring Following Maternal Overnutrition. Nutrients 2019, 11, 1241. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hernandez-Carretero, A.; Weber, N.; la Frano, M.R.; Ying, W.; Rodriguez, L.; Sears, D.D.; Wallenius, V.; Borgeson, E.; Newman, J.W.; Osborn, O. Long-Term Increased Carnitine Palmitoyltransferase 1A Expression in Ventromedial Hypotalamus Causes Hyperphagia and Alters the Hypothalamic Lipidomic Profile. Int. J. Obes. 2018, 42, 728–736. [Google Scholar] [CrossRef] [PubMed]
- Sergi, D.; Morris, A.C.; Kahn, D.E.; McLean, F.H.; Hay, E.A.; Kubitz, P.; MacKenzie, A.; Martinoli, M.G.; Drew, J.E.; Williams, L.M. Palmitic Acid Triggers Inflammatory Responses in N42 Cultured Hypothalamic Cells Partially via Ceramide Synthesis but Not via TLR4. Nutr. Neurosci. 2018, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Dusaban, S.S.; Chun, J.; Rosen, H.; Purcell, N.H.; Brown, J.H. Sphingosine 1-Phosphate Receptor 3 and RhoA Signaling Mediate Inflammatory Gene Expression in Astrocytes. J. Neuroinflamm. 2017, 14, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Heras, V.; Castellano, J.M.; Fernandois, D.; Velasco, I.; Rodríguez-Vazquez, E.; Roa, J.; Vazquez, M.J.; Ruiz-Pino, F.; Rubio, M.; Pineda, R.; et al. Central Ceramide Signaling Mediates Obesity-Induced Precocious Puberty. Cell Metab. 2020, 32, 951–966.e8. [Google Scholar] [CrossRef] [PubMed]
- Lyu, P.; Huang, Z.; Feng, Q.; Su, Y.; Zheng, M.; Hong, Y.; Cai, X.; Lu, Z. Unveiling the Transcriptome Alteration of POMC Neuron in Diet-Induced Obesity. Exp. Cell Res. 2020, 389, 111848. [Google Scholar] [CrossRef] [PubMed]
- Bellini, L.; Campana, M.; Mahfouz, R.; Carlier, A.; Veret, J.; Magnan, C.; Hajduch, E.; le Stunff, H. Targeting Sphingolipid Metabolism in the Treatment of Obesity/Type 2 Diabetes. Expert Opin. Ther. Targets 2015, 19, 1037–1050. [Google Scholar] [CrossRef]
- Le Stunff, H.; Coant, N.; Migrenne, S.; Magnan, C. Targeting Lipid Sensing in the Central Nervous System: New Therapy against the Development of Obesity and Type 2 Diabetes. Expert Opin. Ther. Targets 2013, 17, 545–555. [Google Scholar] [CrossRef]
- Goldenthal, M.J.; Marín-García, J. Mitochondrial Signaling Pathways: A Receiver/Integrator Organelle. Mol. Cell. Biochem. 2004, 262, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Schell, M.; Chudoba, C.; Leboucher, A.; Alfine, E.; Flore, T.; Ritter, K.; Weiper, K.; Wernitz, A.; Henkel, J.; Kleinridders, A. Interplay of Dietary Fatty Acids and Cholesterol Impacts Brain Mitochondria and Insulin Action. Nutrients 2020, 12, 1518. [Google Scholar] [CrossRef] [PubMed]
- Simopoulos, A.P. An Increase in the Omega-6/Omega-3 Fatty Acid Ratio Increases the Risk for Obesity. Nutrients 2016, 8, 128. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Henkel, J.; Coleman, C.D.; Schraplau, A.; Jöhrens, K.; Weber, D.; Castro, J.P.; Hugo, M.; Schulz, T.J.; Krämer, S.; Schürmann, A.; et al. Induction of Steatohepatitis (NASH) with Insulin Resistance in Wild-Type B6 Mice by a Western-Type Diet Containing Soybean Oil and Cholesterol. Mol. Med. 2017, 23, 70–82. [Google Scholar] [CrossRef] [PubMed]
- Bonaccio, M.; Di Castelnuovo, A.; Costanzo, S.; De Curtis, A.; Persichillo, M.; Sofi, F.; Cerletti, C.; Donati, M.B.; de Gaetano, G.; Iacoviello, L. Ultra-Processed Food Consumption Is Associated with Increased Risk of All-Cause and Cardiovascular Mortality in the Moli-Sani Study. Am. J. Clin. Nutr. 2021, 113, 446–455. [Google Scholar] [CrossRef]
- Leffa, P.S.; Hoffman, D.J.; Rauber, F.; Sangalli, C.N.; Valmórbida, J.L.; Vitolo, M.R. Longitudinal Associations between Ultra-Processed Foods and Blood Lipids in Childhood. Br. J. Nutr. 2020, 124, 341–348. [Google Scholar] [CrossRef] [Green Version]
- Haynes, C.A.; Allegood, J.C.; Park, H.; Sullards, M.C. Sphingolipidomics: Methods for the Comprehensive Analysis of Sphingolipids. J. Chromatogr. B 2009, 877, 2696–2708. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Han, X.; Jiang, X. A Review of Lipidomic Technologies Applicable to Sphingolipidomics and Their Relevant Applications. Eur. J. Lipid Sci. Technol. 2009, 111, 39–52. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Iqbal, J.; Walsh, M.T.; Hammad, S.M.; Hussain, M.M. Sphingolipids and Lipoproteins in Health and Metabolic Disorders. Trends Endocrinol. Metab. 2017, 28, 506–518. [Google Scholar] [CrossRef] [PubMed]
- Holland, W.L.; Brozinick, J.T.; Wang, L.P.; Hawkins, E.D.; Sargent, K.M.; Liu, Y.; Narra, K.; Hoehn, K.L.; Knotts, T.A.; Siesky, A.; et al. Inhibition of Ceramide Synthesis Ameliorates Glucocorticoid-, Saturated-Fat-, and Obesity-Induced Insulin Resistance. Cell Metab. 2007, 5, 167–179. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tettamanti, G. Ganglioside/Glycosphingolipid Turnover: New Concepts. Glycoconj. J. 2003, 20, 301–317. [Google Scholar] [CrossRef] [PubMed]
- Cuschieri, J.; Bulger, E.; Billgrin, J.; Garcia, I.; Maier, R.V. Acid Sphingomyelinase Is Required for Lipid Raft TLR4 Complex Formation. Surg. Infect. 2007, 8, 91–105. [Google Scholar] [CrossRef] [PubMed]
- Holland, W.L.; Bikman, B.T.; Wang, L.P.; Yuguang, G.; Sargent, K.M.; Bulchand, S.; Knotts, T.A.; Shui, G.; Clegg, D.J.; Wenk, M.R.; et al. Lipid-Induced Insulin Resistance Mediated by the Proinflammatory Receptor TLR4 Requires Saturated Fatty Acid-Induced Ceramide Biosynthesis in Mice. J. Clin. Investig. 2011, 121, 1858–1870. [Google Scholar] [CrossRef] [Green Version]
- Frangioudakis, G.; Garrard, J.; Raddatz, K.; Nadler, J.L.; Mitchell, T.W.; Schmitz-Peiffer, C. Saturated- and n-6 Polyunsaturated-Fat Diets Each Induce Ceramide Accumulation in Mouse Skeletal Muscle: Reversal and Improvement of Glucose Tolerance by Lipid Metabolism Inhibitors. Endocrinology 2010, 151, 4187–4196. [Google Scholar] [CrossRef] [Green Version]
- Thombare, K.; Ntika, S.; Wang, X.; Krizhanovskii, C. Long Chain Saturated and Unsaturated Fatty Acids Exert Opposing Effects on Viability and Function of GLP-1-Producing Cells: Mechanisms of Lipotoxicity. PLoS ONE 2017, 12, e0177605. [Google Scholar] [CrossRef] [Green Version]
- Tse, E.K.; Belsham, D.D. Palmitate Induces Neuroinflammation, ER Stress, and Pomc MRNA Expression in Hypothalamic MHypoA-POMC/GFP Neurons through Novel Mechanisms That Are Prevented by Oleate. Mol. Cell. Endocrinol. 2018, 472, 40–49. [Google Scholar] [CrossRef]
- Cintra, D.E.; Ropelle, E.R.; Moraes, J.C.; Pauli, J.R.; Morari, J.; de Souza, C.T.; Grimaldi, R.; Stahl, M.; Carvalheira, J.B.; Saad, M.J.; et al. Unsaturated Fatty Acids Revert Diet-Induced Hypothalamic Inflammation in Obesity. PLoS ONE 2012, 7. [Google Scholar] [CrossRef]
- Casals, N.; Zammit, V.; Herrero, L.; Fadó, R.; Rodríguez-Rodríguez, R.; Serra, D. Carnitine Palmitoyltransferase 1C: From Cognition to Cancer. Prog. Lipid Res. 2016, 61, 134–148. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wolfgang, M.J.; Kurama, T.; Dai, Y.; Suwa, A.; Asaumi, M.; Matsumoto, S.I.; Cha, S.H.; Shimokawa, T.; Lane, M.D. The Brain-Specific Carnitine Palmitoyltransferase-1c Regulates Energy Homeostasis. Proc. Natl. Acad. Sci. USA 2006, 103, 7282–7287. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jeng, J.Y.; Lee, W.H.; Tsai, Y.H.; Chen, C.Y.; Chao, S.Y.; Hsieh, R.H. Functional Modulation of Mitochondria by Eicosapentaenoic Acid Provides Protection against Ceramide Toxicity to C6 Glioma Cells. J. Agric. Food Chem. 2009, 57, 11455–11462. [Google Scholar] [CrossRef] [PubMed]
- Stoica, B.A.; Movsesyan, V.A.; Lea IV, P.M.; Faden, A.I. Ceramide-Induced Neuronal Apoptosis Is Associated with Dephosphorylation of Akt, BAD, FKHR, GSK-3β, and Induction of the Mitochondrial-Dependent Intrinsic Caspase Pathway. Mol. Cell. Neurosci. 2003, 22, 365–382. [Google Scholar] [CrossRef]
- Chavez, J.A.; Summers, S.A. A Ceramide-Centric View of Insulin Resistance. Cell Metab. 2012, 15, 585–594. [Google Scholar] [CrossRef] [Green Version]
- Adams, J.M.; Pratipanawatr, T.; Berria, R.; Wang, E.; DeFronzo, R.A.; Sullards, M.C.; Mandarino, L.J. Ceramide Content Is Increased in Skeletal Muscle from Obese Insulin-Resistant Humans. Diabetes 2004, 53, 25–31. [Google Scholar] [CrossRef] [Green Version]
- Kolak, M.; Westerbacka, J.; Velagapudi, V.R.; Wågsäter, D.; Yetukuri, L.; Makkonen, J.; Rissanen, A.; Häkkinen, A.M.; Lindell, M.; Bergholm, R.; et al. Adipose Tissue Inflammation and Increased Ceramide Content Characterize Subjects with High Liver Fat Content Independent of Obesity. Diabetes 2007, 56, 1960–1968. [Google Scholar] [CrossRef] [Green Version]
- Nowotny, B.; Zahiragic, L.; Krog, D.; Nowotny, P.J.; Herder, C.; Carstensen, M.; Yoshimura, T.; Szendroedi, J.; Phielix, E.; Schadewaldt, P.; et al. Mechanisms Underlying the Onset of Oral Lipid-Induced Skeletal Muscle Insulin Resistance in Humans. Diabetes 2013, 62, 2240–2248. [Google Scholar] [CrossRef] [Green Version]
- Haus, J.M.; Kashyap, S.R.; Kasumov, T.; Zhang, R.; Kelly, K.R.; Defronzo, R.A.; Kirwan, J.P. Plasma Ceramides Are Elevated in Obese Subjects with Type 2 Diabetes and Correlate with the Severity of Insulin Resistance. Diabetes 2009, 58, 337–343. [Google Scholar] [CrossRef] [Green Version]
- Galadari, S.; Rahman, A.; Pallichankandy, S.; Galadari, A.; Thayyullathil, F. Role of Ceramide in Diabetes Mellitus: Evidence and Mechanisms. Lipids Health Dis. 2013, 12. [Google Scholar] [CrossRef] [Green Version]
- Wang, M.Y.; Koyama, K.; Shimabukuro, M.; Newgard, C.B.; Uncer, R.H. OB-Rb Gene Transfer to Leptin-Resistant Islets Reverses Diabetogenic Phenotype. Proc. Natl. Acad. Sci. USA 1998, 95, 714–718. [Google Scholar] [CrossRef] [Green Version]
- Naik, M.U.; Benedikz, E.; Hernandez, I.; Libien, J.; Hrabe, J.; Valsamis, M.; Dow-edwards, D.; Osman, M.; Sacktor, T.C. Distribution of Protein Kinase Mζ and the Complete Protein Kinase C Isoform Family in Rat Brain. J. Comp. Neurol. 2000, 426, 243–258. [Google Scholar] [CrossRef]
- Contreras, C.; González-García, I.; Seoane-Collazo, P.; Martínez-Sánchez, N.; Liñares-Pose, L.; Rial-Pensado, E.; Fernø, J.; Tena-Sempere, M.; Casals, N.; Diéguez, C.; et al. Reduction of Hypothalamic Endoplasmic Reticulum Stress Activates Browning of White Fat and Ameliorates Obesity. Diabetes 2017, 66, 87–99. [Google Scholar] [CrossRef] [Green Version]
- Yatomi, Y. Plasma Sphingosine 1-Phosphate Metabolism and Analysis. Biochim. Biophys. Acta Gen. Subj. 2008, 1780, 606–611. [Google Scholar] [CrossRef] [PubMed]
- Christoffersen, C.; Obinata, H.; Kumaraswamy, S.B.; Galvani, S.; Ahnström, J.; Sevvana, M.; Egerer-Sieber, C.; Muller, Y.A.; Hla, T.; Nielsen, L.B.; et al. Endothelium-Protective Sphingosine-1-Phosphate Provided by HDL-Associated Apolipoprotein M. Proc. Natl. Acad. Sci. USA 2011, 108, 9613–9618. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tran-Dinh, A.; Diallo, D.; Delbosc, S.; Varela-Perez, L.M.; Dang, Q.B.; Lapergue, B.; Burillo, E.; Michel, J.B.; Levoye, A.; Martin-Ventura, J.L.; et al. HDL and Endothelial Protection. Br. J. Pharmacol. 2013, 169, 493–511. [Google Scholar] [CrossRef] [Green Version]
- Potì, F.; Simoni, M.; Nofer, J.R. Atheroprotective Role of High-Density Lipoprotein (HDL)-Associated Sphingosine-1-Phosphate (S1P). Cardiovasc. Res. 2014, 103, 395–404. [Google Scholar] [CrossRef] [Green Version]
- Fayyaz, S.; Henkel, J.; Japtok, L.; Krämer, S.; Damm, G.; Seehofer, D.; Püschel, G.P.; Kleuser, B. Involvement of Sphingosine 1-Phosphate in Palmitate-Induced Insulin Resistance of Hepatocytes via the S1P2 Receptor Subtype. Diabetologia 2014, 57, 373–382. [Google Scholar] [CrossRef] [Green Version]
- Véret, J.; Coant, N.; Gorshkova, I.A.; Giussani, P.; Fradet, M.; Riccitelli, E.; Skobeleva, A.; Goya, J.; Kassis, N.; Natarajan, V.; et al. Role of Palmitate-Induced Sphingoid Base-1-Phosphate Biosynthesis in INS-1 β-Cell Survival. Biochim. Biophys. Acta Mol. Cell Biol. Lipids 2013, 1831, 251–262. [Google Scholar] [CrossRef]

 = high reliability,
= high reliability,  = not clear,
= not clear,  = low reliability.
= high reliability, = not clear, = low reliability.
= low reliability.
= high reliability, = not clear, = low reliability.
 = high reliability,
= high reliability,  = not clear,
= not clear,  = low reliability.
= high reliability, = not clear, = low reliability.
= low reliability.
= high reliability, = not clear, = low reliability.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Inclusion Criteria | Exclusion Criteria | |
|---|---|---|
| Population | Rodents (offspring and/or adults); hypothalamic cell lines; or primary hypothalamic cell culture. | Other cells than hypothalamic; non-rodents and studies involving human subjects. |
| Intervention | High-fat feeding; genetic models of obesity; treatment with fatty acids; treatment with cytokines; treatment with sphingolipids (ceramide or sphingosine-1-phosphate). | Other genetic backgrounds; animals with protein restriction or receiving any other treatment. |
| Comparison | Sphingolipid levels (ceramide and sphingosine-1-phosphate); fasting. | Studies related to ganglioside metabolism. |
| Outcomes | Alterations in sphingolipid metabolism following disturbances in energy balance control. Primary outcomes: sphingolipid level (ceramide and sphingosine -1-phosphate). Secondary outcomes: Body weight, food intake, energy expenditure assessments, neuropeptides levels, and inflammatory markers. | Other outcomes not relevant to energy balance control. |
| Type of publication | Original article | Nonoriginal article (reviews, conference abstracts). |
| Other selection criteria | Studies following these main characteristics were included: animal and cell culture studies from 1990 until 2020. The articles needed to be published on these databases: PubMed, Embase, Web of Science, and Scopus. Keywords searched: sphingolipid and hypothalamus. | Non-English language. |
| Outcomes Type | ||||||
|---|---|---|---|---|---|---|
| Authors | Design | Treatment | Sphingolipid Extraction/Analysis Method | Sphingolipid Levels | Inflammatory Parameters | Neuropeptide Levels |
| Sortino et al., 1999 | Hypothalamic GT1-7 neuronal cells. |
| Diacylglycerol kinase assay (commercially available kit) | ↑ Cer-1-P (accumulation of ceramide) in GT1-7 cells stimulated with TNF-α. ↓ Cer-1-P in GT1-7 cells after pretreatment with D609. | ↑ TNFR1 and TNFR2 in GT1-7 cells after TNF-α exposure | Uninformed |
| Davis et al., 2006 | Wild-type mixed AH cultures (containing neurons and glia). |
| Uninformed | ↑ Src phosphorylation in the AH cultures after treatment with C2-Ceramide or IL-1β (concentration and time-dependent) | Uninformed | Uninformed |
| McFadden et al., 2014 | PHN, N38HN, or R7HN cells. |
| Lipidomics | ↑ Ceramide levels in PHD exposed to palmitate. ↓ Ceramide levels in PDH exposed to C75 | ↑ TNF-α, IL-1β, and IL-6 mRNA levels in PHN after exposed to C16:0 for 18 h. ↓ TNF-α, IL-1β mRNA levels in PHN exposed to C75 alone or in the presence of C16:0, FSG67, and C89b for 18 h. | Uninformed |
| Morselli et al., 2014a | N43 cells and BV2 cells. |
| Liquid chromatography/electrospray ionization/tandem mass spectrometry | ↑ Ceramide levels in N43 cells exposed to PA. | ↑ TNF-α and IL-6 mRNA expression in N43 and BV2 cells exposed to PA for 8 h. ↓ TNF-α and IL-6 in N43 cells transfected with siRNA for PGC-1a, followed by treatment with PA. ↑ TNF-α and IL-6 in N43 cells transfected with siRNA for PGC-1a, followed by infection with AdGFP-Era followed by infection with AdGFP-ERa, with or without exposed to PA; | Uninformed |
| Morselli et al., 2014b | N43 hypothalamic cell line and primary neuronal cell cultures. |
| Mass spectrometry | Uninformed | ↑ Inflammation in N43 cell line and primary neurons exposed to PA. ↓ Inflammation in N43 cell line and primary neurons were pretreatment with E2. | Uninformed |
| Silva et al., 2014 | GT1-7 cells. |
| Western blotting | ↑ S1PR1 protein levels in GT1-7 cells treated with leptin, in a time-dependent manner. ↓ S1PR1 protein levels in GT1-7 cells transfected with STAT3 siRNA. | Uninformed | Uninformed |
| Campana et al.,2017 | Hypothalamic GT1-7 neuronal cells. |
| Liquid chromatography coupled with high-resolution mass spectrometry. | ↑ Ceramide levels (C:16, C:18, C22, C24) in GT1-7 cells after treatment with palmitate. ↓ SPT2 blocked ceramide accumulation induced by palmitate in GT1-7 cells. | Uninformed | Uninformed |
| Dusaban et al., 2017 | Astrocytes were isolated from P1-P3 postnatal WT and S1P3 KO mice. |
| Quantitative-PCR | ↑ S1P3 in WT astrocytes after scratch injury. | ↑ COX2 and IL-6 levels in S1P3 KO astrocytes treated with S1P. | Uninformed |
| Tse, E. and Belsham, D., 2018 | mHypoA-POMC/GFP-1, -2, -3, and -4 neurons. |
| Uninformed | Uninformed | ↑ mRNA levels of IL-6, IL-1B, TLR4, TNF-α, and NFkB after treatment with 50 mM palmitate for 8 h. ↓ IL6 mRNA expression after inhibition of TLR4 and pretreatment with PS 1145. ↑ IL-6 mRNA expression after pretreatment with SP 600125 and SB 202190. ↓ IL-6 mRNA expression after Oleate co-treatment. ↓ JNK phosphorylation in mHypoAPOMC/GFP-2 neurons pretreated with 50 mM palmitate and insulin stimulation. | ↑ POMC mRNA expression in mHypoA-POMC/GFP-2 neurons with 50 mM palmitate for 8 h or 1 mM C16-ceramide for 8 h. ↓ POMC mRNA expression after pretreatments with 50mM SP 600125 (JNK inhibitor) or 10-mM PD 0325901 (ERK inhibitor). ↑ POMC mRNA expression in mHypoA-POMC/GFP-2 neurons with pretreatment with 100 mM myriocin or 50 mM L-cycloserine for 1 h. Cotreatment rescue PA induced POMC mRNA. |
| Sergi et al., 2018 | mHypoE-N42 hypothalamic cell line. Primary hypothalamic culture from Sprague Dawley rats. |
| LC- ESI-MS/MS | ↑ C16 ceramide after PA treatment. ↓ C16 ceramide when L-cycloserine was added in combination with PA. ↓ C16 ceramide in N42 neurons were treated with PA in the presence of OA or EPA. | ↑ IL-6 and TNF-α expression in N42, N49 cells and hypothalamic cultures, after treatment with PA or LPS. ↓ IL-6 e TNF-α expression after treatment with L-cycloserine, in N42 cells compared with LPS group. ↓ IL-6 expression after treatment with LA in N42 cells. ↓ IL-6 and TNF-α expression after cotreatment with OA and EPA in N42 cells. ↓ IL-6 expression after treatment with PA combined with cycloserine compared to PA alone in N42 cells | Uninformed |
| Maldonado-Ruiz et al., 2019 | Microglia primary culture from Wistar rats. |
| Uninformed | Uninformed | ↑ TNF-α, IL-6 and IL-1 after treatment with palmitate. ↑ TNF-α after treatment with stearic acid. ↑ IL-6 after treatment with palmitoleic acid. ↓ IL-6 after treatment with C6. | Uninformed |
| Outcome Type | ||||||
|---|---|---|---|---|---|---|
| Authors | Design | Treatment | Weight gain | Food Intake | Adiposity (%) | Energy Expenditure |
| Gao et al., 2011 | Male Sprague Dawley. |
| Body weights after infusing leptin into the Arc overexpressing CPT1-α were significantly attenuated compared with the null condition. ↓ Body weight after infused myriocin or pretreatment with N-hexanoyl-D-sphingosine. | ↑ Food intake in rats with overexpression of CPT1-α. ↓ Feeding after infusing leptin into the Arc overexpressing CPT1-α. ↑ Food intake in CPT1-α knockout mice after injections with leptin or cerulenin. ↓ Food intake after in infused myriocin or pretreatment with N-hexanoyl-D-sphingosine. | Uninformed | Uninformed |
| Borg et al., 2012 | 8 week old male mice of C57BL/6 and ob/ob lineage. |
| ↑ Body mass in HFD mice Exercise training did not affect body mass in the HFD mice. | Uninformed | ↑ Epididymal fat mass in HFD mice. Exercise training did not decrease epididymal fat mass. | Uninformed |
| Ramírez et al., 2013 | Adult male mice of wild type and CPT1-α knockout. |
| Uninformed | ↑ Food intake after ghrelin IP injection in wild type mice. ↓ Food intake after ICV injection of myriocin (1 h before ghrelin administration). ↑ Food intake after ceramide injection in CPT1 KO mice. | Uninformed | Uninformed |
| Contreras et al., 2014 | Male Sprague Dawley rats and lean and obese male Zucker rats (LZR, OZR). |
| ↑ Body weight after C6 injections for 5 days. ↓ Weight gain ceramide-induced after administration of GRP78 adenovirus. ↓ Body weight in OZR treated with GRP78 adenovirus. | Central treatment of C6 ceramide and GRP78 adenovirus did not affect feeding. | ↑ Weight of gonadal and inguinal white adipose tissue promoted by central ceramide injections. ↓ Weight of gonadal and inguinal white adipose tissue promoted by GRP78 adenovirus. ↓ Weight of gonadal and inguinal white adipose tissue in OZR, promoted by GRP78 adenovirus. | Uninformed |
| Mera et al., 2014 | Male Sprague Dawley. |
| ↑ Body weight in CPT1AM rats after 20 days of AAV injection | ↑ Food intake in CPT1AM rats after 20 days of AAV injection | ↑ Adiposity in CPT1AM rats | Uninformed |
| Morselli et a., 2014a | 8 week old male and female C57BL/6. |
| ↑ Body weight in males and females after consuming HFD for 16 weeks. | Uninformed | Uninformed | Uninformed |
| Morselli et al., 2014b | 4 week old male and female C57BL/6. |
| ↑ Body weight of males and females after consuming HFD for 4 weeks. | Uninformed | Uninformed | Uninformed |
| Silva et al., 2014 | 5 week old male Wistar rats and 10 week old male Swiss, C57BL/6J, ob/ob and db/db mice. |
| ↓ Body weight in obese rats with ICV injection of S1P. No difference in the total body weight was observed 48 h after S1PR1 siRNA injection. | ↓ Food intake in obese and lean rats after injection of S1P or SEW2871. The JSI124 pretreatment, 30 min before S1P ICV injection, was sufficient to block the effect of S1P injection. | ↓ Epididymal fat-pad weight in obese rats with ICV injection of S1P. | ↑ Energy expenditure in rats with ICV injection of S1P. ↓ Energy expenditure in rats, after 4 h, with S1PR1 siRNA. Did not observe difference in the energy expenditure after S1P ICV injection in obese rats. |
| Campana et al., 2017 | 10 week old male Zucker rats (obese or lean) and Wistar. |
| Central myriocin treatment did not affect body weight in either obese or lean Zucker rats. | Central myriocin treatment did not affect food intake in either obese or lean Zucker rats. | Central myriocin treatment did not affect lean and fat body mass in either obese or lean Zucker rats. | Uninformed |
| Gao et al., 2017 | 6–10 week old male mice of LPL-knockout mice (GFAP-LPL−/−) and C57BL/6. |
| ↑ Body weight in GFAP-LPL−/− when fed an HFD. | ↑ Food intake in GFAP-LPL−/− when fed an HFD. | ↑ Fat mass in GFAP-LPL−/− when fed an HFD. | ↓ Locomotor activity in LPL knockout with HFD. ↑ Energy expenditure in LPL knockout with HFD |
| Laperrousaz et al., 2017 | 8 week old male mice of MBHLpl, C57BL6/J, NexLpl−/−, Agrp−/−. |
| ↑ Body weight after LPL deletion. ↓ Body weight after LPL overexpression | There was no difference | ↑ Adiposity in mice with LPL deletion. | ↓ Locomotor activity and energy expenditure in mice with LPL deletion. ↑ RER in mice with LPL deletion. |
| Gonzalez-Garcıa et al., 2018 | Female Sprague Dawley |
| ↑ Body weight in OVX rats after 15 days of surgery. ↓ Body weight in OVX rats after treatment with estradiol or myriocin or TUDCA or shSPTLC1 adenovirus or GPR78 adenovirus. ↑ Body weight in OVX rats after treatment with SR59230A. | ↑ Food intake in OVX rats. ↓ Food intake in OVX rats after treatment with estradiol or myriocin. The other treatments did not affect dietary intake. | ↑ Adiposity in OVX rats. ↓ Adiposity in OVX rats after treatment with estradiol or myriocin. | ↑ Energy expenditure in OVX rats after treatment with estradiol. ↓ Respiratory quotient in OVX rats after treatment with estradiol. |
| Hernandez-Carretero et al., 2018 | 12 week old male mice of C57BL/6N. |
| ↑ Body weight in HFD mice. ↓ Body weight in SWD mice. ↓ Body weight in HFD mice after treatment with HQL-79. | ↓ Food intake in HFD fed mice treated with HQL-79 | Uninformed | Uninformed |
| Maldonado-Ruiz et al., 2019 | 8 week old male and female Wistar rats. |
| Uninformed | ↑ Food intake in offspring programmed by the CAF diet. ↑ Food intake in spring programmed by maternal CAF diet after subcutaneous ghrelin injection. ↑ Food intake in offspring programmed by maternal CAF diet after LPS or PAL by ACSF injections. | Uninformed | Uninformed |
| Vozella et al., 2019 | 8 week old male mice of C57BL/6J. | Mice were fed a standard diet.
The refeeding groups were food deprived for 12 h and then allowed to feed for 1 h or 6 h. | Uninformed | Uninformed | Uninformed | Uninformed |
| Heras et a., 2020 | Female pups (small litters fed with HFD 45%). |
| ↑ Overnutrition female rats displayed increased body weight. C6 or Myriocin do not alter body weight. ↓ Underfed and Myriocin reduced body weight. | Not altered in Myriocin or C6 group | Uninformed | Uninformed |
| Lyu et al., 2020 | Pomc-Cre mice (Stock No. 005965) and ROSA-tdTomato mice (Stock No. 007676). |
| ↑ Body weight in HFD-fed mice. ↓ Body weight in HFD-fed mice with telmisartan treatment. ↓ (pRb∆P) lentivirus body weight induced by HFD. | ↑ Food intake in HFD-fed mice. | ↑ Abdominal white fa in HFD-fed mice. ↓ Abdominal white fat in pRb∆P lentivirus rescue. | Uninformed |
| Rawish et al., 2020 | 6–8 week old male mice of C57BL/6N. |
| ↑ Body weight in HFD-fed mice. ↓ Body weight in HFD-fed mice with telmisartan treatment. | ↑ Food intake in HDF-fed mice. ↓ Food intake in HFD- Fed mice with telmisartan treatment. | ↑ Fat mass in HFD-fed mice. ↓ Fat mass in HFD-fed mice with telmisartan treatment. | ↓ Energy expenditure and locomotion in HDF-fed mice. ↑ Energy expenditure and locomotion in HDF-fed mice with telmisartan treatment, in dark periods. |
| Outcomes Type | ||||
|---|---|---|---|---|
| Authors | Sphingolipid Extraction/Analysis Method | Sphingolipid Levels | Inflammatory Parameters | Neuropeptide Levels |
| Gao et al., 2011 | HPLC coupled with mass spectrometry | ↑ Ceramide level in rats with overexpressing CPT-1a (under fasting condition). ↓ Ceramide level after leptin injection in rats with overexpression of CPT-1a. ↓ Ceramide level after myriocin injection in rats with overexpression of CPT-1a. ↓ Ceramide level in rats with CPT-1c deleted. | Uninformed | ↑ NPY levels in rats with overexpression of CPT-1a. ↓ NPY levels after leptin injection in rats with overexpression of CPT-1a/ ↓ NPY levels after infused with myriocin in rats with overexpression of CPT-1a. The levels of AgRP and POMC were not altered. |
| Borg et al., 2012 | Electrospray ionization-tandem mass spectrometry | ↑ Ceramide species (18:0, 22:0 and 24:0) in the hypothalamus of HFD mice. ↑ Dihydroceramide and dihexosylceramide contents in the hypothalamus of HFD. | ↓ IκBα expression in HFD mice. ↑ IκBα with exercise training. Hypothalamic JNK signaling was not affected by obesity or exercise training | Uninformed |
| Ramírez et al., 2013 | LC-ESI-MS/MS system | ↑ Total ceramides and C:18 ceramides in the wild type mice after ghrelin injection. | ICV injection of myriocin (1 h before ghrelin administration) did not change the levels of inflammatory markers (TLR4, pIKKB, IKKB, NFKB) | ↑ AgRP and NPY levels after ghrelin injection in wild-type mice. ↓ AgRP and NPY levels after pretreatment with myriocin (1 h before ghrelin administration). ↑ AgRP and NPY levels after ceramide injection in CPT1 KO mice. |
| Contreras et al., 2014 | Liquid chromatography-electrospray ionization/multistage mass spectrometry system | ↑ Concentration of C16 ceramide in the hypothalamus promoted by central ceramide injections. ↓ Concentration of ceramide C16 and C18 in the hypothalamus of OZR. Administration of the GRP78 adenovirus did not alter the levels of hypothalamic ceramide. | ↑ IL-6, TNF-, and pIKKα/β expression in the hypothalamus after treatment with ceramide. Administration of the GRP78 adenovirus did not alter the expression of inflammatory markers in the hypothalamus. | Uninformed |
| Mera et al., 2014 | Lipidomic analysis | Unaltered total ceramide levels. ↑ Ceramide species (C14:0 and C18:1), total lactosylceramides, total concentrations of sphingomyelin and dihydrosphingomyelin and lysophosphatidylcholine in CPT1AM animals. ↓ Total levels of lysophosphatidylethanolamine, plasmalogen-phosphatidylethanolamine, lysoplasmalogen, and lysophosphatidylserin in CPT1A animals. | Unaltered TNF-, IL-6, IL-1, and INOS mRNA expression. ↑ MCP1 mRNA expression in CPT1AM animals. | Unaltered POMC, CART, NPY, AgRP mRNA levels. ↑ NPY1R mRNA level in CPT1AM animals. |
| Morselli et a., 2014a | Liquid chromatography/electrospray ionization/tandem mass spectrometry | ↑ Accumulation of ceramides and sphingomielin in the hypothalamus of male mice when compared to females, after HFD feeding. ↑ Glucosylceramide levels in males and females, after HFD feeding. | ↑ TNF-α, IL-1β, and IL-6 levels in the hypothalamus of male, after HFD feeding. ↓ IL-10 in the hypothalamus of male, after HFD feeding | Uninformed |
| Morselli et al., 2014b | Mass spectrometry | ↑ Ceramide, glucosylceramide, and sphingomyelin levels in male compared to female, consuming HFD for 4 weeks. | ↑ TNF-α, IL-1β and IL-6 levels in male compared to female, after consuming HFD for 4 weeks. | Uninformed |
| Silva et al., 2014 | Kit Assay, Western blot, RT-PCR | ↓ S1PR1 levels in the hypothalamus of obese rats. | ↑ STAT3 phosphorylation in the hypothalamus of obese rats after injection of S1P. | ↑ POMC mRNA in obese rats after injection of S1P or SEW2871, but not alter NPY. |
| Campana et al., 2017 | Liquid chromatography/tandem mass spectrometry. | ↑ Total ceramide levels in the hypothalamus of obese Zucker rats. ↓ Total ceramide levels in the hypothalamus of obese Zucker rats after ICV injection of myriocin. | Uninformed | Uninformed |
| Gao et al., 2017 | Chromatographic separation and mass spectrometer | ↑ Total ceramide levels and ceramide species (C18:0, C18:1 and C22) in the hypothalamus of GFAP-LPL−/− mice. | ↑ IBA1 in the hypothalamus of GFAP-LPL−/− mice. | ↑ AGRP in the GFAP-LPL−/− mice. POMC cell number did not change |
| Laperrousaz et al., 2017 | Liquid chromatography coupled with LC | ↓ Ceramide total levels and ceramide species (dC18/C16, dC18/C:18; DC18:0/C:20; dC18:00/C:22; dC18:0/C:24) in the hypothalamus with LPL deletion at 10 days after injection. ↓ Cers1 mRNA in the hypothalamus with LPL deletion at 10 days and 12 weeks after injection. ↑ Spt3, Cers2, Cers3 mRNA in the hypothalamus with LPL deletion at 12 weeks after injection. | Uninformed | NPY and AGRP did not affect |
| Gonzalez-Garcıa et al., 2018 | Liquid chromatography/tandem mass spectrometry | ↑ Ceramide levels in the hypothalamus after OVX. ↓ Ceramide levels in the hypothalamus of OVX rats after treatment with estradiol or myriocin. | Uninformed | Uninformed |
| Hernandez-Carretero et al., 2018 | Electrospray ionization | ↑ Ceramide species (C18:1) in adipose, muscle, and plasma of HDF mice. ↑ Ceramide species (C:20) in adipose, liver, and plasma of HDF mice. ↑ C18 dihydroceramide in adipose and plasma of HDF mice. ↑ C18:1 sphingosine in adipose and liver of HDF mice. | ↑ F4/80 in HDF and SWD mice. | Uninformed |
| Maldonado-Ruiz et al., 2019 | Uninformed | Uninformed | ↑ IBA-1 in offspring programmed by a maternal CAF diet after ghrelin and palmitic acid injections. ↑ NFKB phosphorylation after LPS administration; | Uninformed |
| Vozella et al., 2019 | Liquid chromatography/tandem mass spectrometry. | ↓ Sphingosine, Sphingosine 1 Phosphate, Diidroxiceramide after 12 h of fasting. ↓ Hypothalamic levels of SA1P, S1P and SA after 12 h of fasting, which was partially (SA1P) or completely (S1P, SA) reversed after 6 h refeeding. ↓ Levels of dihydroceramide (d18:0/18:0) in the hypothalamus of food-deprived mice. ↓ S1pr1 transcription levels after 12 h of fasting an effect was rapidly and completely reversed by refeeding. ↓ Transcription levels of Sptlc2, Lass1, SphK2, Sphk1 after 12 h of fasting, which are partially (Sptlc2) or completely (Lass1, SphK2) reversed by refeeding. | Uninformed | Uninformed |
| Heras et a., 2020 | High-performance liquid chromatography (HPLC) | ↑ Hypothalamic total ceramide and ceramide species (CERC16, CERC18, CERC18:1, CERC:20, CERC(a)24:1, CERC14:0, CERC16:1, CERC20:1, CERC22:1, and CER24:2) in female rats subjected to early overnutrition. | Uninformed | Myriocin treatment did not change hypothalamic expression of Pomc, Cart, Npy, or Agrp. ↓ NPY levels after C6 treatment. |
| Lyu et al., 2020 | RNAseq; qRT-PCR | ↑Asah2, Cers2, and Elovl1 were in POMC neurons in the HFD group. | ↑ Chemokines in POMC neurons in the HFD group. | Uninformed |
| Rawish et al., 2020 | Liquid chromatography-mass | ↑ Ceramide, cholesteryl ester, phosphatidylcholine, phosphatidylethanolamine, and sphingomyelin levels in plasma of HFD-fed mice. ↓ Ceramide levels in plasma of HFD-fed mice treated with telmisartan. ↑ Hypothalamic levels of ceramide (particularly Cer d36:1) (d18:1/18:0)) in HFD-fed mice. ↓ Hypothalamic levels of ceramide (particularly Cer d36:1) (d18:1/18:0)) in HFD-fed mice treated with telmisartan. | ↑ TNF-α and Cxcl12 in HFD-fed mice. ↓ CXCL12 in HFD-fed mice after Telmisartan treatment. ↑ IL-4 and IL-6 levels in plasma of both HFD-fed mice and HFD-fed mice treated with telmisartan. ↑ IL-5 levels in plasma of HFD-fed mice with treatment of telmisartan. | Uninformed |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Reginato, A.; Veras, A.C.C.; Baqueiro, M.d.N.; Panzarin, C.; Siqueira, B.P.; Milanski, M.; Lisboa, P.C.; Torsoni, A.S. The Role of Fatty Acids in Ceramide Pathways and Their Influence on Hypothalamic Regulation of Energy Balance: A Systematic Review. Int. J. Mol. Sci. 2021, 22, 5357. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22105357
Reginato A, Veras ACC, Baqueiro MdN, Panzarin C, Siqueira BP, Milanski M, Lisboa PC, Torsoni AS. The Role of Fatty Acids in Ceramide Pathways and Their Influence on Hypothalamic Regulation of Energy Balance: A Systematic Review. International Journal of Molecular Sciences. 2021; 22(10):5357. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22105357
Chicago/Turabian StyleReginato, Andressa, Alana Carolina Costa Veras, Mayara da Nóbrega Baqueiro, Carolina Panzarin, Beatriz Piatezzi Siqueira, Marciane Milanski, Patrícia Cristina Lisboa, and Adriana Souza Torsoni. 2021. "The Role of Fatty Acids in Ceramide Pathways and Their Influence on Hypothalamic Regulation of Energy Balance: A Systematic Review" International Journal of Molecular Sciences 22, no. 10: 5357. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22105357