
Molecular Elucidation of a Urate Oxidase from Deinococcus radiodurans for Hyperuricemia and Gout Therapy


Abstract
:1. Introduction
2. Results
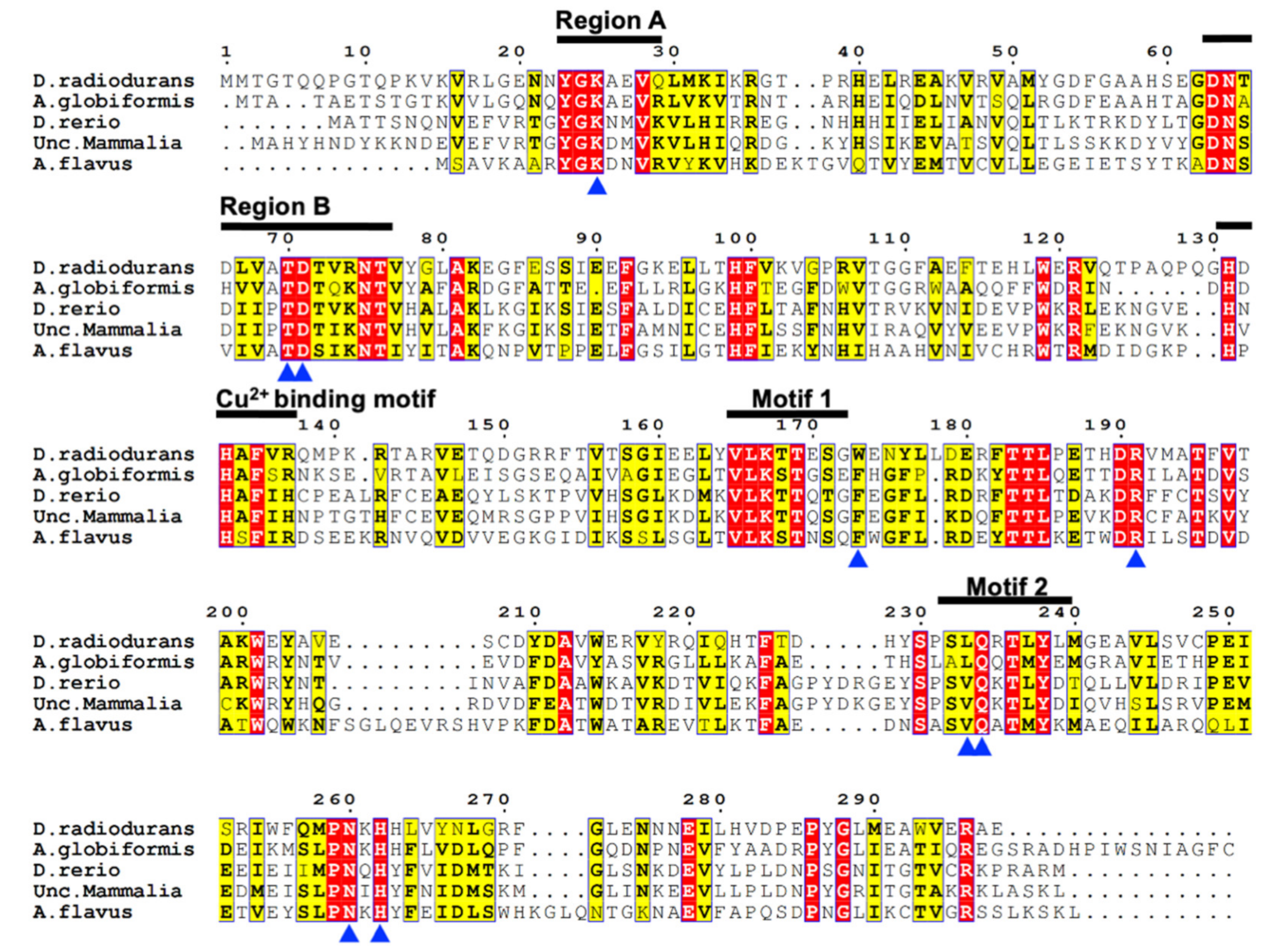
2.1. Amino Acid Sequence Analysis of DrUox
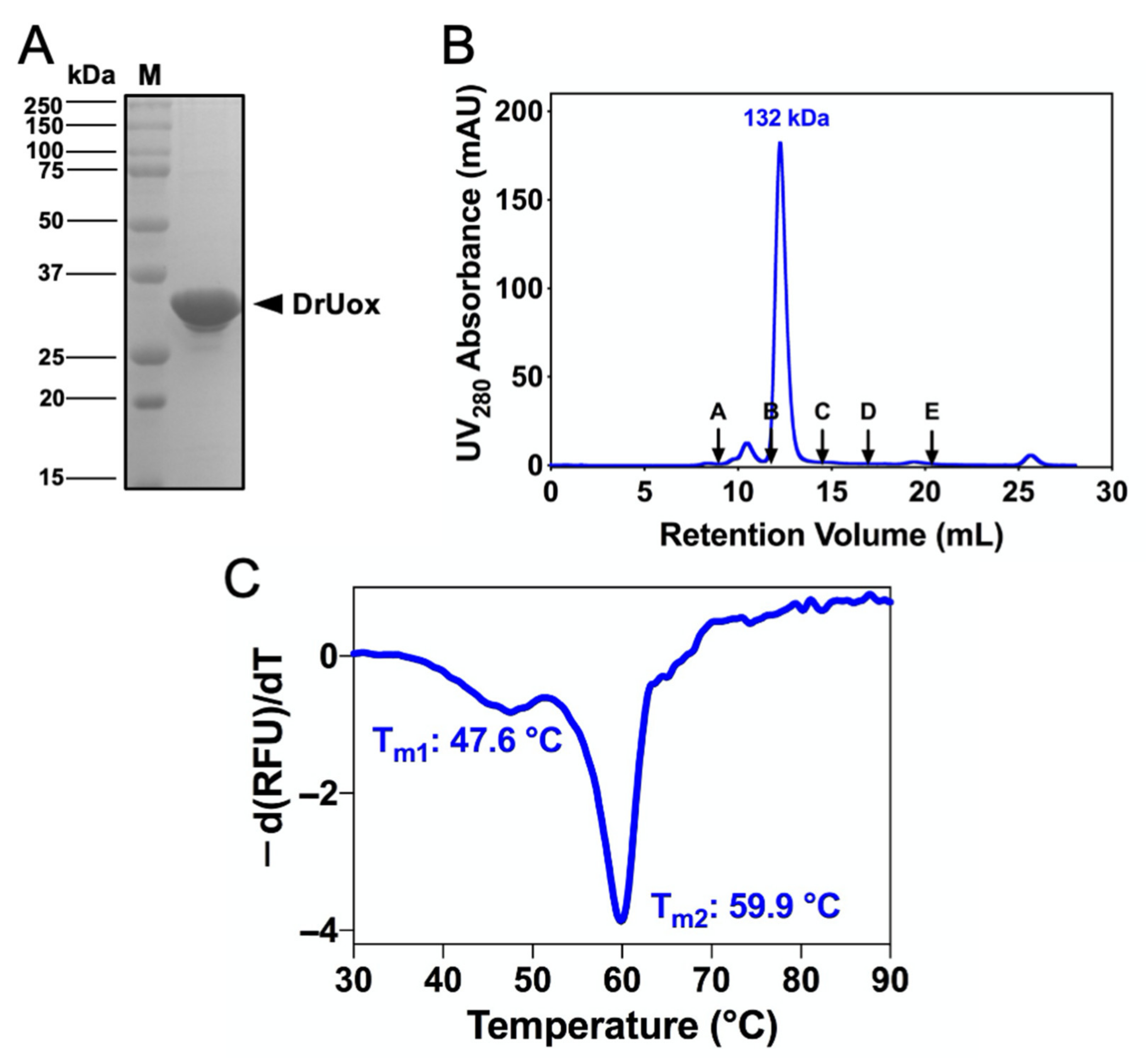
2.2. Cloning, Expression and Purification of Recombinant DrUox
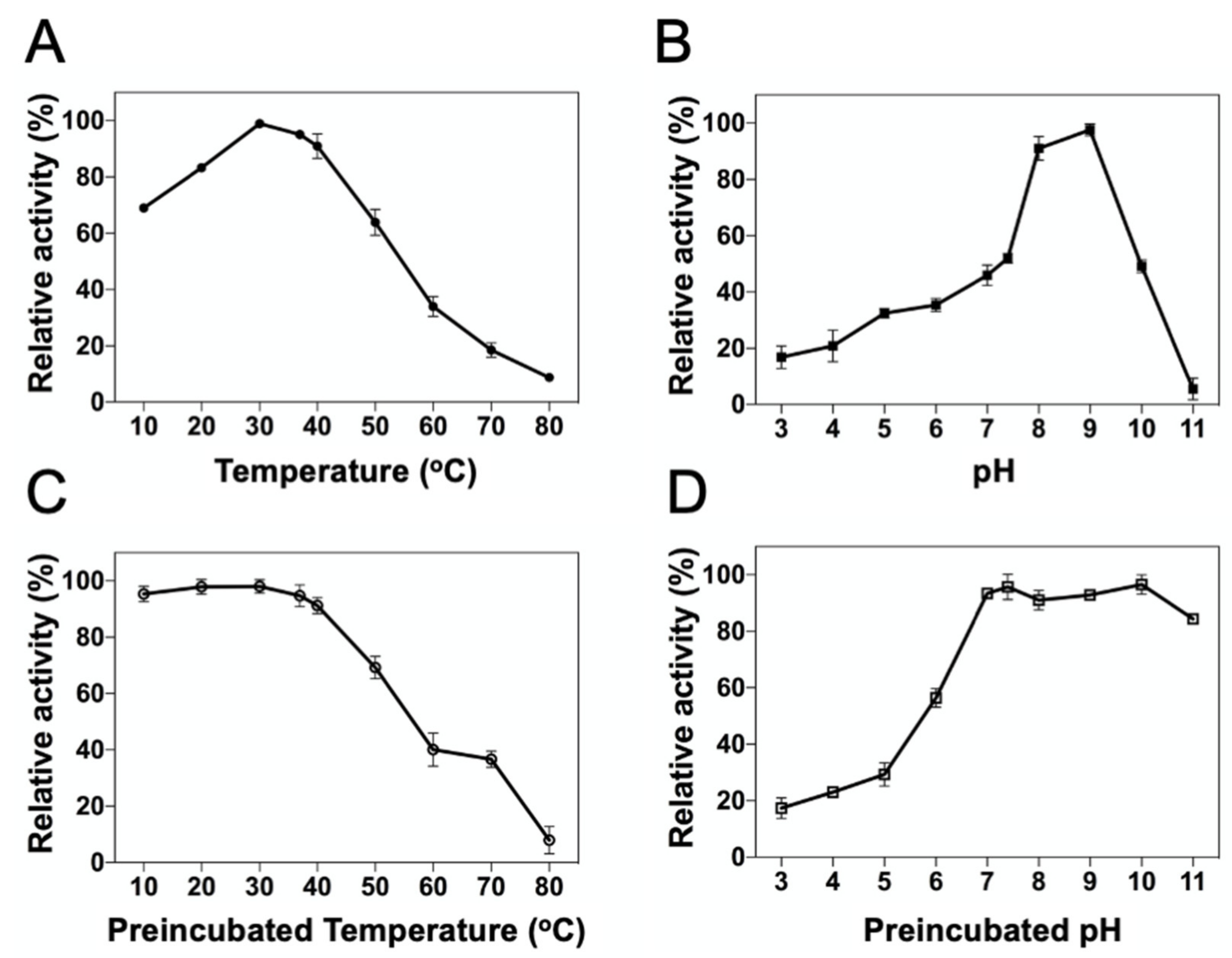
2.3. Influence of Temperature and PH on DrUox
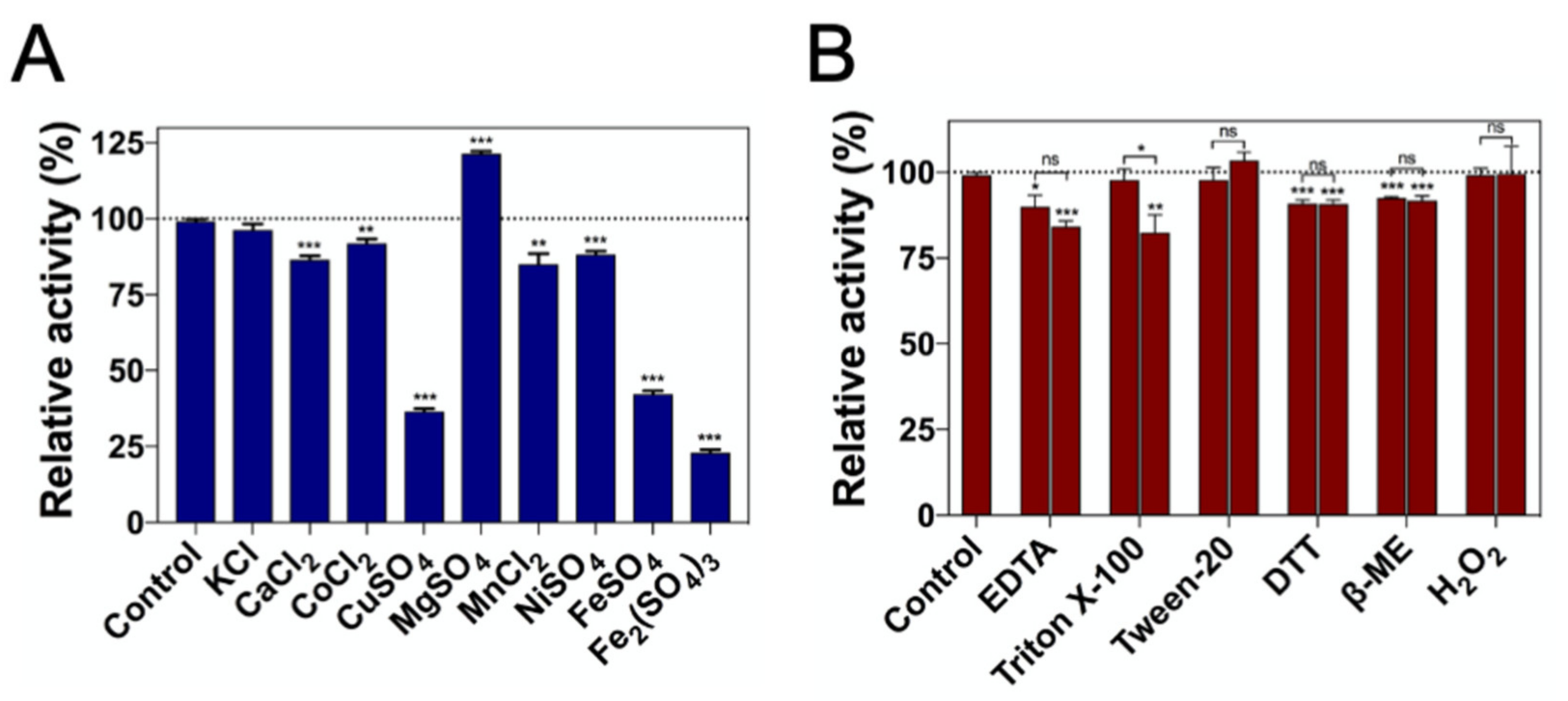
2.4. Effect of Metal Ions and Chemicals on DrUox Activity
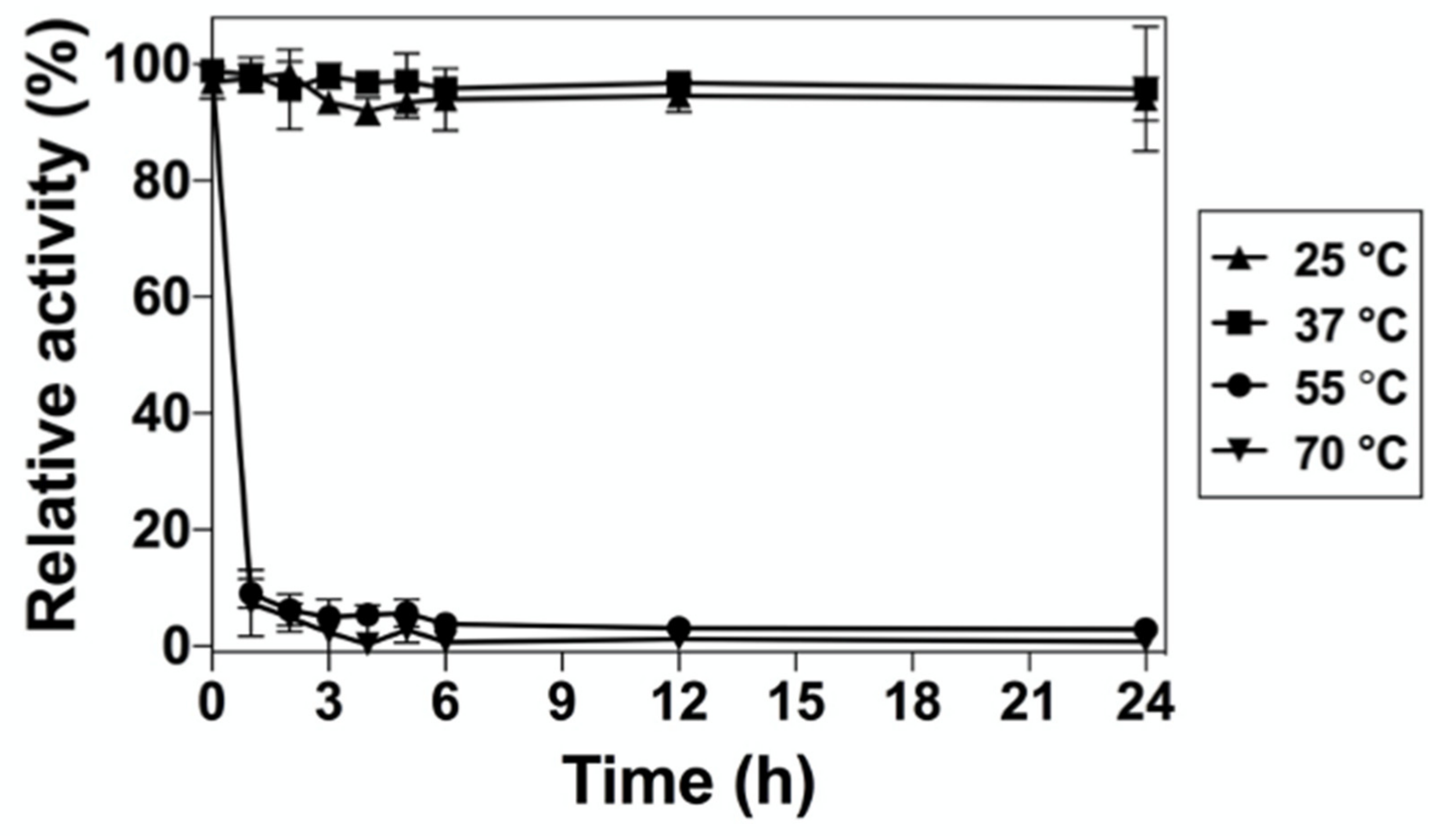
2.5. Effect of Temperature on the Long-Term Stability of DrUox
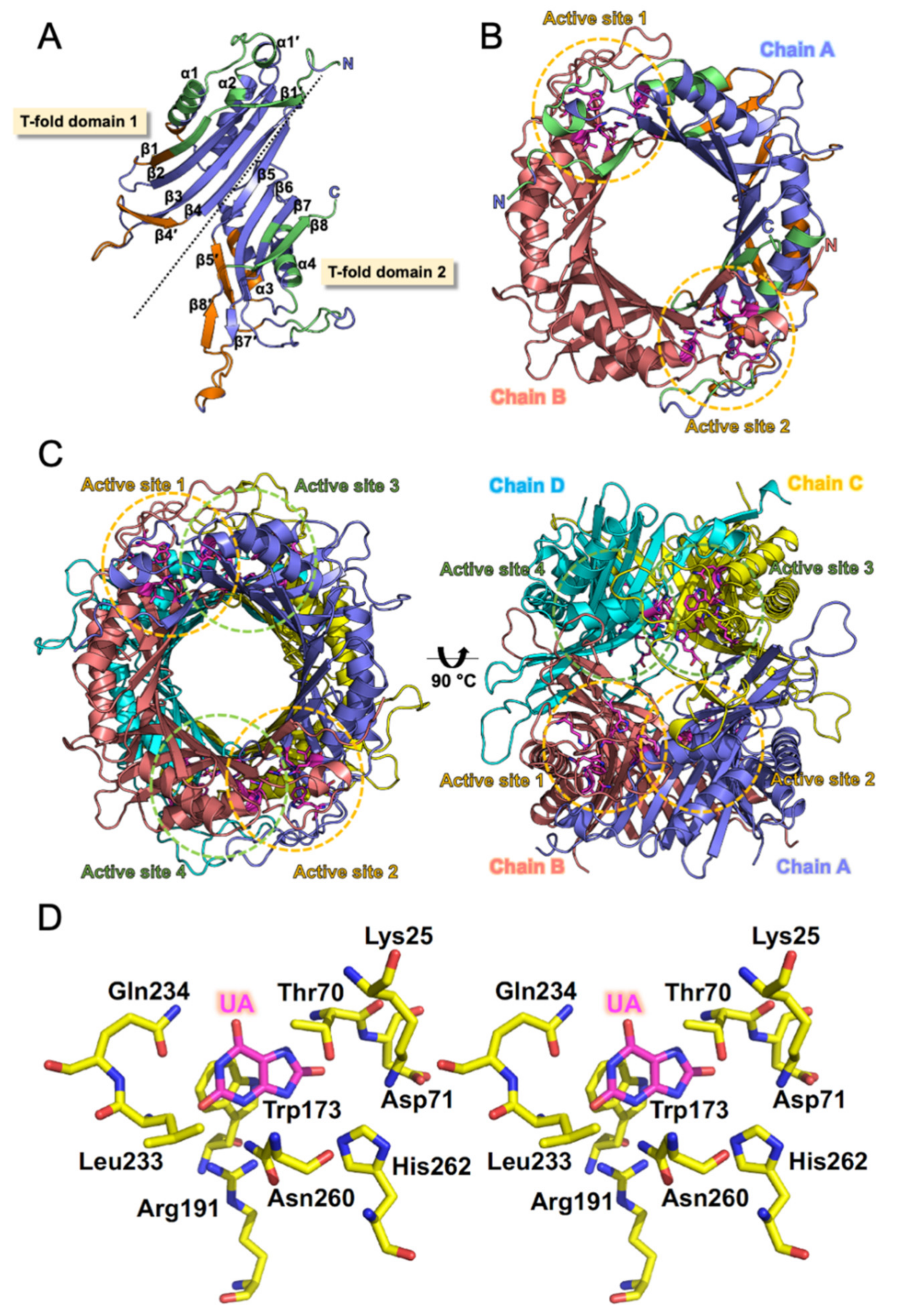
2.6. Analysis of Structural Properties of DrUox
3. Discussion
4. Materials and Methods
4.1. Materials
4.2. Genomic DNA Extraction
4.3. Plasmid Constructs, Protein Expression and Purification
4.4. Molecular Characterization by SDS-PAGE and Size Exclusion Chromatography
4.5. Circular Dichroism (CD) Spectroscopy
4.6. Thermal Shift Assay
4.7. Urate Oxidase Activity Assay
4.8. Effect of Metal Ions, Surfactants, Chelating Agent, Reducing Agents and Oxidative Agent
4.9. Long-Term Thermal Stability Measurement
4.10. Homology Modeling and Quality Evaluation of DrUox
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Colloc’h, N.; el Hajji, M.; Bachet, B.; L’Hermite, G.; Schiltz, M.; Prange, T.; Castro, B.; Mornon, J.P. Crystal structure of the protein drug urate oxidase-inhibitor complex at 2.05 A resolution. Nat. Struct. Biol. 1997, 4, 947–952. [Google Scholar] [CrossRef]
- Ramazzina, I.; Folli, C.; Secchi, A.; Berni, R.; Percudani, R. Completing the uric acid degradation pathway through phylogenetic comparison of whole genomes. Nat. Chem. Biol. 2006, 2, 144–148. [Google Scholar] [CrossRef] [PubMed]
- Kahn, K.; Tipton, P.A. Spectroscopic characterization of intermediates in the urate oxidase reaction. Biochemistry 1998, 37, 11651–11659. [Google Scholar] [CrossRef]
- Wu, X.W.; Lee, C.C.; Muzny, D.M.; Caskey, C.T. Urate oxidase: Primary structure and evolutionary implications. Proc. Natl. Acad. Sci. USA 1989, 86, 9412–9416. [Google Scholar] [CrossRef] [Green Version]
- Varela-Echavarria, A.; Montes de Oca-Luna, R.; Barrera-Saldana, H.A. Uricase protein sequences: Conserved during vertebrate evolution but absent in humans. FASEB J. 1988, 2, 3092–3096. [Google Scholar] [CrossRef]
- Kratzer, J.T.; Lanaspa, M.A.; Murphy, M.N.; Cicerchi, C.; Graves, C.L.; Tipton, P.A.; Ortlund, E.A.; Johnson, R.J.; Gaucher, E.A. Evolutionary history and metabolic insights of ancient mammalian uricases. Proc. Natl. Acad. Sci. USA 2014, 111, 3763–3768. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, X.W.; Muzny, D.M.; Lee, C.C.; Caskey, C.T. Two independent mutational events in the loss of urate oxidase during hominoid evolution. J. Mol. Evol. 1992, 34, 78–84. [Google Scholar] [CrossRef]
- Neogi, T. Clinical practice. Gout. N. Engl. J. Med. 2011, 364, 443–452. [Google Scholar] [CrossRef]
- Feig, D.I.; Kang, D.H.; Johnson, R.J. Uric acid and cardiovascular risk. N. Engl. J. Med. 2008, 359, 1811–1821. [Google Scholar] [CrossRef] [PubMed]
- Abeles, A.M. Hyperuricemia, gout, and cardiovascular disease: An update. Curr. Rheumatol. Rep. 2015, 17, 13. [Google Scholar] [CrossRef]
- Jones, D.P.; Mahmoud, H.; Chesney, R.W. Tumor lysis syndrome: Pathogenesis and management. Pediatr. Nephrol. 1995, 9, 206–212. [Google Scholar] [CrossRef]
- Jordan, A.; Gresser, U. Side Effects and Interactions of the Xanthine Oxidase Inhibitor Febuxostat. Pharmaceuticals 2018, 11, 51. [Google Scholar] [CrossRef] [Green Version]
- Stamp, L.K.; Chapman, P.T. Urate-lowering therapy: Current options and future prospects for elderly patients with gout. Drugs Aging 2014, 31, 777–786. [Google Scholar] [CrossRef]
- Li, J.; Chen, Z.; Hou, L.; Fan, H.; Weng, S.; Xu, C.; Ren, J.; Li, B.; Chen, W. High-level expression, purification, and characterization of non-tagged Aspergillus flavus urate oxidase in Escherichia coli. Protein Expr. Purif. 2006, 49, 55–59. [Google Scholar] [CrossRef]
- Coiffier, B.; Mounier, N.; Bologna, S.; Ferme, C.; Tilly, H.; Sonet, A.; Christian, B.; Casasnovas, O.; Jourdan, E.; Belhadj, K.; et al. Efficacy and safety of rasburicase (recombinant urate oxidase) for the prevention and treatment of hyperuricemia during induction chemotherapy of aggressive non-Hodgkin’s lymphoma: Results of the GRAAL1 (Groupe d’Etude des Lymphomes de l’Adulte Trial on Rasburicase Activity in Adult Lymphoma) study. J. Clin. Oncol. 2003, 21, 4402–4406. [Google Scholar]
- Sundy, J.S.; Baraf, H.S.; Yood, R.A.; Edwards, N.L.; Gutierrez-Urena, S.R.; Treadwell, E.L.; Vazquez-Mellado, J.; White, W.B.; Lipsky, P.E.; Horowitz, Z.; et al. Efficacy and tolerability of pegloticase for the treatment of chronic gout in patients refractory to conventional treatment: Two randomized controlled trials. JAMA 2011, 306, 711–720. [Google Scholar]
- Sattui, S.E.; Gaffo, A.L. Treatment of hyperuricemia in gout: Current therapeutic options, latest developments and clinical implications. Ther. Adv. Musculoskelet. Dis. 2016, 8, 145–159. [Google Scholar] [CrossRef]
- Huang, Y.; Chen, Y.; Yang, X.; Zhao, H.; Hu, X.; Pu, J.; Liao, J.; Long, G.; Liao, F. Optimization of pH values to formulate the bireagent kit for serum uric acid assay. Biotechnol. Appl. Biochem. 2015, 62, 137–144. [Google Scholar] [CrossRef]
- Liu, J.; Li, G.; Liu, H.; Zhou, X. Purification and properties of uricase from Candida sp. and its application in uric acid analysis in serum. Appl. Biochem. Biotechnol. 1994, 47, 57–63. [Google Scholar] [CrossRef]
- Shi, Y.; Wang, T.; Zhou, X.E.; Liu, Q.F.; Jiang, Y.; Xu, H.E. Structure-based design of a hyperthermostable AgUricase for hyperuricemia and gout therapy. Acta Pharmacol. Sin. 2019, 40, 1364–1372. [Google Scholar] [CrossRef]
- Hibi, T.; Kume, A.; Kawamura, A.; Itoh, T.; Fukada, H.; Nishiya, Y. Hyperstabilization of Tetrameric Bacillus sp. TB-90 Urate Oxidase by Introducing Disulfide Bonds through Structural Plasticity. Biochemistry 2016, 55, 724–732. [Google Scholar] [CrossRef]
- Li, W.; Xu, S.; Zhang, B.; Zhu, Y.; Hua, Y.; Kong, X.; Sun, L.; Hong, J. Directed evolution to improve the catalytic efficiency of urate oxidase from Bacillus subtilis. PLoS ONE 2017, 12, e0177877. [Google Scholar] [CrossRef]
- Suzuki, K.; Sakasegawa, S.; Misaki, H.; Sugiyama, M. Molecular cloning and expression of uricase gene from Arthrobacter globiformis in Escherichia coli and characterization of the gene product. J. Biosci. Bioeng. 2004, 98, 153–158. [Google Scholar] [CrossRef]
- Koyama, Y.; Ichikawa, T.; Nakano, E. Cloning, sequence analysis, and expression in Escherichia coli of the gene encoding the Candida utilis urate oxidase (uricase). J. Biochem. 1996, 120, 969–973. [Google Scholar] [CrossRef] [Green Version]
- Chu, R.; Lin, Y.; Usuda, N.; Rao, M.S.; Reddy, J.K.; Yeldandi, A.V. Mutational analysis of the putative copper-binding site of rat urate oxidase. Ann. N. Y. Acad. Sci. 1996, 804, 781–786. [Google Scholar] [CrossRef]
- Nyborg, A.C.; Ward, C.; Zacco, A.; Chacko, B.; Grinberg, L.; Geoghegan, J.C.; Bean, R.; Wendeler, M.; Bartnik, F.; O’Connor, E.; et al. A Therapeutic Uricase with Reduced Immunogenicity Risk and Improved Development Properties. PLoS ONE 2016, 11, e0167935. [Google Scholar] [CrossRef]
- Liu, X.; Wen, M.; Li, J.; Zhai, F.; Ruan, J.; Zhang, L.; Li, S. High-yield expression, purification, characterization, and structure determination of tag-free Candida utilis uricase. Appl. Microbiol. Biotechnol. 2011, 92, 529–537. [Google Scholar] [CrossRef]
- Wang, B.; Luo, L.; Wang, D.; Ding, R.; Hong, J. Efficient purification of a recombinant tag-free thermostable Kluyveromyces marxianus uricase by pH-induced self-cleavage of intein and expression in Escherichia coli. 3 Biotech 2018, 8, 400. [Google Scholar] [CrossRef]
- Pfrimer, P.; de Moraes, L.M.; Galdino, A.S.; Salles, L.P.; Reis, V.C.; De Marco, J.L.; Prates, M.V.; Bloch, C., Jr.; Torres, F.A. Cloning, purification, and partial characterization of Bacillus subtilis urate oxidase expressed in Escherichia coli. J. Biomed. Biotechnol. 2010, 2010, 674908. [Google Scholar] [CrossRef] [Green Version]
- Rubingh, D.N. The influence of surfactants on enzyme activity. Curr. Opin. Colloid Interface Sci. 1996, 1, 598–603. [Google Scholar] [CrossRef]
- Colloc’h, N.; Gabison, L.; Monard, G.; Altarsha, M.; Chiadmi, M.; Marassio, G.; Sopkova-de Oliveira Santos, J.; El Hajji, M.; Castro, B.; Abraini, J.H.; et al. Oxygen pressurized X-ray crystallography: Probing the dioxygen binding site in cofactorless urate oxidase and implications for its catalytic mechanism. Biophys. J. 2008, 95, 2415–2422. [Google Scholar] [CrossRef] [Green Version]
- Sreerama, N.; Woody, R.W. Estimation of protein secondary structure from circular dichroism spectra: Comparison of CONTIN, SELCON, and CDSSTR methods with an expanded reference set. Anal. Biochem. 2000, 287, 252–260. [Google Scholar] [CrossRef]
- Retailleau, P.; Colloc’h, N.; Vivares, D.; Bonnete, F.; Castro, B.; El-Hajji, M.; Mornon, J.P.; Monard, G.; Prange, T. Complexed and ligand-free high-resolution structures of urate oxidase (Uox) from Aspergillus flavus: A reassignment of the active-site binding mode. Acta. Crystallogr. D Biol. Crystallogr. 2004, 60, 453–462. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Imhoff, R.D.; Power, N.P.; Borrok, M.J.; Tipton, P.A. General base catalysis in the urate oxidase reaction: Evidence for a novel Thr-Lys catalytic diad. Biochemistry 2003, 42, 4094–4100. [Google Scholar] [CrossRef] [PubMed]
- Gabison, L.; Prange, T.; Colloc’h, N.; El Hajji, M.; Castro, B.; Chiadmi, M. Structural analysis of urate oxidase in complex with its natural substrate inhibited by cyanide: Mechanistic implications. BMC Struct. Biol. 2008, 8, 32. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goyal, R.N.; Mittal, A.; Agarwal, D. Electrochemical Oxidation and Kinetics of the Decay of Uv-Absorbing Intermediate of Uric-Acid Oxidation at Pyrolytic-Graphite Electrodes. Can. J. Chem. 1994, 72, 1668–1674. [Google Scholar] [CrossRef]
- Altarsha, M.; Castro, B.; Monard, G. Intrinsic reactivity of uric acid with dioxygen: Towards the elucidation of the catalytic mechanism of urate oxidase. Bioorg. Chem. 2009, 37, 111–125. [Google Scholar] [CrossRef]
- Simic, M.G.; Jovanovic, S.V. Antioxidation Mechanisms of Uric-Acid. J. Am. Chem. Soc. 1989, 111, 5778–5782. [Google Scholar] [CrossRef]
- Kahn, K.; Tipton, P.A. Kinetic mechanism and cofactor content of soybean root nodule urate oxidase. Biochemistry 1997, 36, 4731–4738. [Google Scholar] [CrossRef]
- Laboureur, P.; Langlois, C. Urate Oxidase from Aspergillus Flavus. 2. Metabolism Inhibitions Specificity. B Soc. Chim. Biol. 1968, 50, 827. [Google Scholar]
- Imani, M.; Shahmohamadnejad, S. Recombinant production of Aspergillus Flavus uricase and investigation of its thermal stability in the presence of raffinose and lactose. 3 Biotech 2017, 7, 201. [Google Scholar] [CrossRef]
- Kotb, E. Characterization of a Thermostable Uricase Isolated from Bacillus firmus DWD-33 and its Application for Uric Acid Quantification in Human Serum. Protein Pept. Lett. 2015, 22, 402–409. [Google Scholar] [CrossRef] [PubMed]
- Kai, L.; Ma, X.H.; Zhou, X.L.; Jia, X.M.; Li, X.; Guo, K.P. Purification and characterization of a thermostable uricase from Microbacterium sp strain ZZJ4-1. World J. Microb. Biot. 2008, 24, 401–406. [Google Scholar] [CrossRef]
- Retailleau, P.; Colloc’h, N.; Vivares, D.; Bonnete, F.; Castro, B.; El Hajji, M.; Prange, T. Urate oxidase from Aspergillus flavus: New crystal-packing contacts in relation to the content of the active site. Acta. Crystallogr. D Biol. Crystallogr. 2005, 61, 218–229. [Google Scholar] [CrossRef] [PubMed]
- Gabison, L.; Chiadmi, M.; Colloc’h, N.; Castro, B.; El Hajji, M.; Prange, T. Recapture of [S]-allantoin, the product of the two-step degradation of uric acid, by urate oxidase. FEBS Lett. 2006, 580, 2087–2091. [Google Scholar] [CrossRef] [Green Version]
- Juan, E.C.; Hoque, M.M.; Shimizu, S.; Hossain, M.T.; Yamamoto, T.; Imamura, S.; Suzuki, K.; Tsunoda, M.; Amano, H.; Sekiguchi, T.; et al. Structures of Arthrobacter globiformis urate oxidase-ligand complexes. Acta. Crystallogr. D Biol. Crystallogr. 2008, D64, 815–822. [Google Scholar] [CrossRef]
- Fraisse, L.; Bonnet, M.C.; de Farcy, J.P.; Agut, C.; Dersigny, D.; Bayol, A. A colorimetric 96-well microtiter plate assay for the determination of urate oxidase activity and its kinetic parameters. Anal. Biochem. 2002, 309, 173–179. [Google Scholar] [CrossRef]
- Waterhouse, A.; Bertoni, M.; Bienert, S.; Studer, G.; Tauriello, G.; Gumienny, R.; Heer, F.T.; de Beer, T.A.P.; Rempfer, C.; Bordoli, L.; et al. SWISS-MODEL: Homology modelling of protein structures and complexes. Nucleic Acids Res 2018, 46, W296–W303. [Google Scholar] [CrossRef] [Green Version]
- Ko, J.; Park, H.; Heo, L.; Seok, C. GalaxyWEB server for protein structure prediction and refinement. Nucleic Acids Res. 2012, 40, W294–W297. [Google Scholar] [CrossRef] [PubMed]
- Wallner, B.; Elofsson, A. Can correct protein models be identified? Protein Sci. 2003, 12, 1073–1086. [Google Scholar] [CrossRef] [Green Version]
- Eisenberg, D.; Luthy, R.; Bowie, J.U. VERIFY3D: Assessment of protein models with three-dimensional profiles. Methods Enzymol. 1997, 277, 396–404. [Google Scholar]
- Wiederstein, M.; Sippl, M.J. ProSA-web: Interactive web service for the recognition of errors in three-dimensional structures of proteins. Nucleic Acids Res. 2007, 35, W407–W410. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Laskowski, R.A.; Rullmannn, J.A.; MacArthur, M.W.; Kaptein, R.; Thornton, J.M. AQUA and PROCHECK-NMR: Programs for checking the quality of protein structures solved by NMR. J. Biomol. NMR 1996, 8, 477–486. [Google Scholar] [CrossRef] [PubMed]
- Colovos, C.; Yeates, T.O. Verification of protein structures: Patterns of nonbonded atomic interactions. Protein Sci. 1993, 2, 1511–1519. [Google Scholar] [CrossRef] [PubMed] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| pH | Temperature (°C) | Km (μM) | Kcat (s−1) | Kcat/Km (s−1∙μM−1) |
|---|---|---|---|---|
| 7.4 | 37 | 760.81 ± 85.89 | 1.41 ± 0.12 | 0.20 × 10−2 ± 0.01 |
| 9.0 | 30 | 332.58 ± 57.94 | 17.49 ± 1.49 | 5.35 × 10−2 ± 0.01 |
| Original Microbes | Genbank Accession No. | Expression Hosts | Specific Activity (U mg−1) | Optimal Temperature (°C) | Temperature Tolerance |
|---|---|---|---|---|---|
| D. radiodurans [this study] | AAF10733 | E. coli | 38.06 | 30 | 37 °C, 24 h, 100% |
| A. flavus [14,41] | CAA43895 | E. coli | 27 | 30–37 | 40 °C, 1 h, 30% |
| B. firmus DWD-33 [42] | — | — | 9.58 | 50 | 60 °C, 1 h, 100% |
| C. utilis [27] | P78609 | E. coli | 38.4 | 37 | 37 °C, 24 h, 40% |
| K. marixianus [28] | BAP70065 | E. coli | 50.54 | 42 | 40 °C, 90 h, 79% |
| Microbacterium sp. strain ZZJ4-1 [43] | AEY68606 | — | 5.32 | 30 | 65 °C, 0.5 h, 100% |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chiu, Y.-C.; Hsu, T.-S.; Huang, C.-Y.; Hsu, C.-H. Molecular Elucidation of a Urate Oxidase from Deinococcus radiodurans for Hyperuricemia and Gout Therapy. Int. J. Mol. Sci. 2021, 22, 5611. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22115611
Chiu Y-C, Hsu T-S, Huang C-Y, Hsu C-H. Molecular Elucidation of a Urate Oxidase from Deinococcus radiodurans for Hyperuricemia and Gout Therapy. International Journal of Molecular Sciences. 2021; 22(11):5611. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22115611
Chicago/Turabian StyleChiu, Yi-Chih, Ting-Syuan Hsu, Chen-Yu Huang, and Chun-Hua Hsu. 2021. "Molecular Elucidation of a Urate Oxidase from Deinococcus radiodurans for Hyperuricemia and Gout Therapy" International Journal of Molecular Sciences 22, no. 11: 5611. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22115611