
Small Molecule Compounds Identified from Mixture-Based Library Inhibit Binding between Plasmodium falciparum Infected Erythrocytes and Endothelial Receptor ICAM-1

 and
and 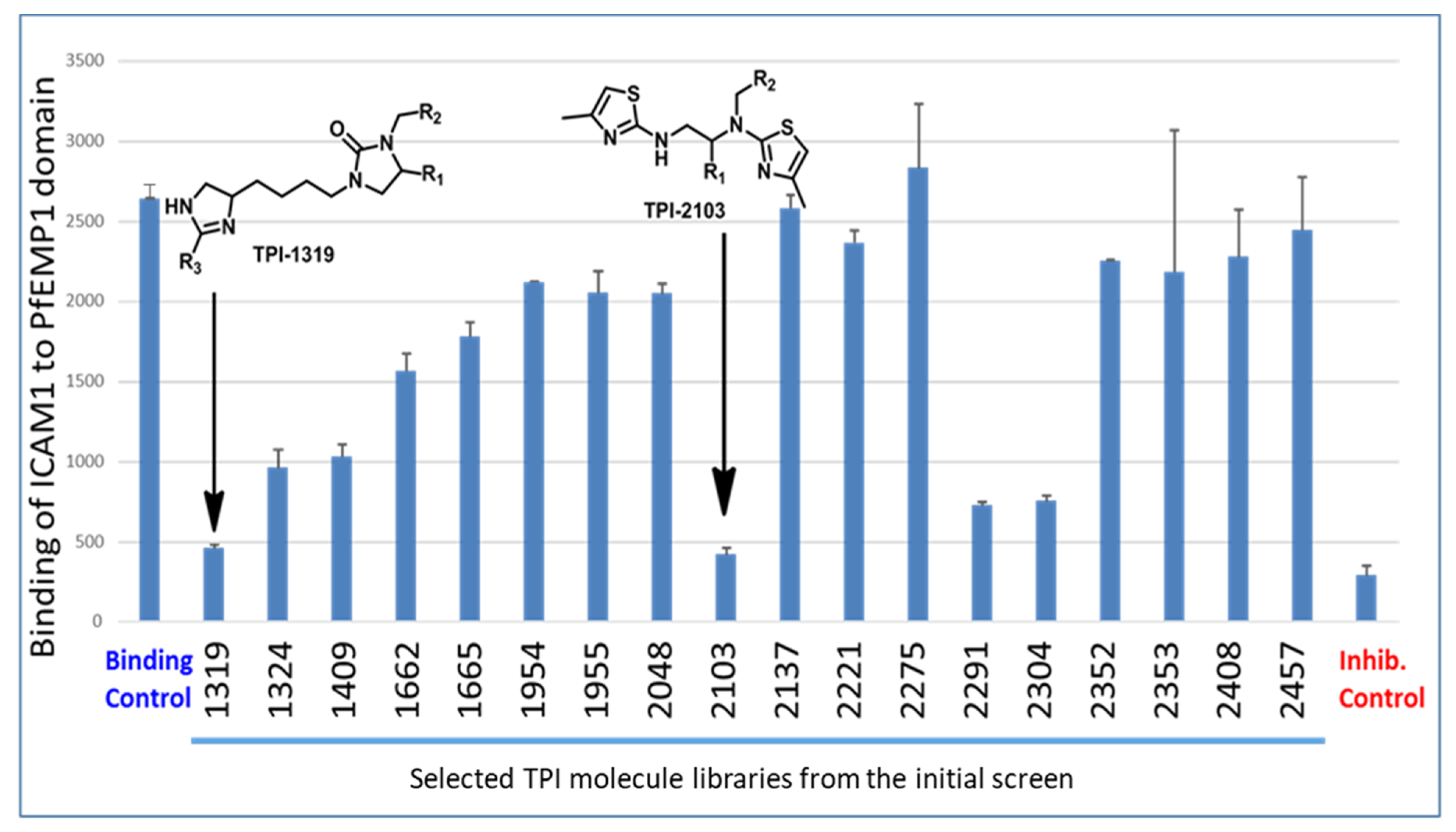{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
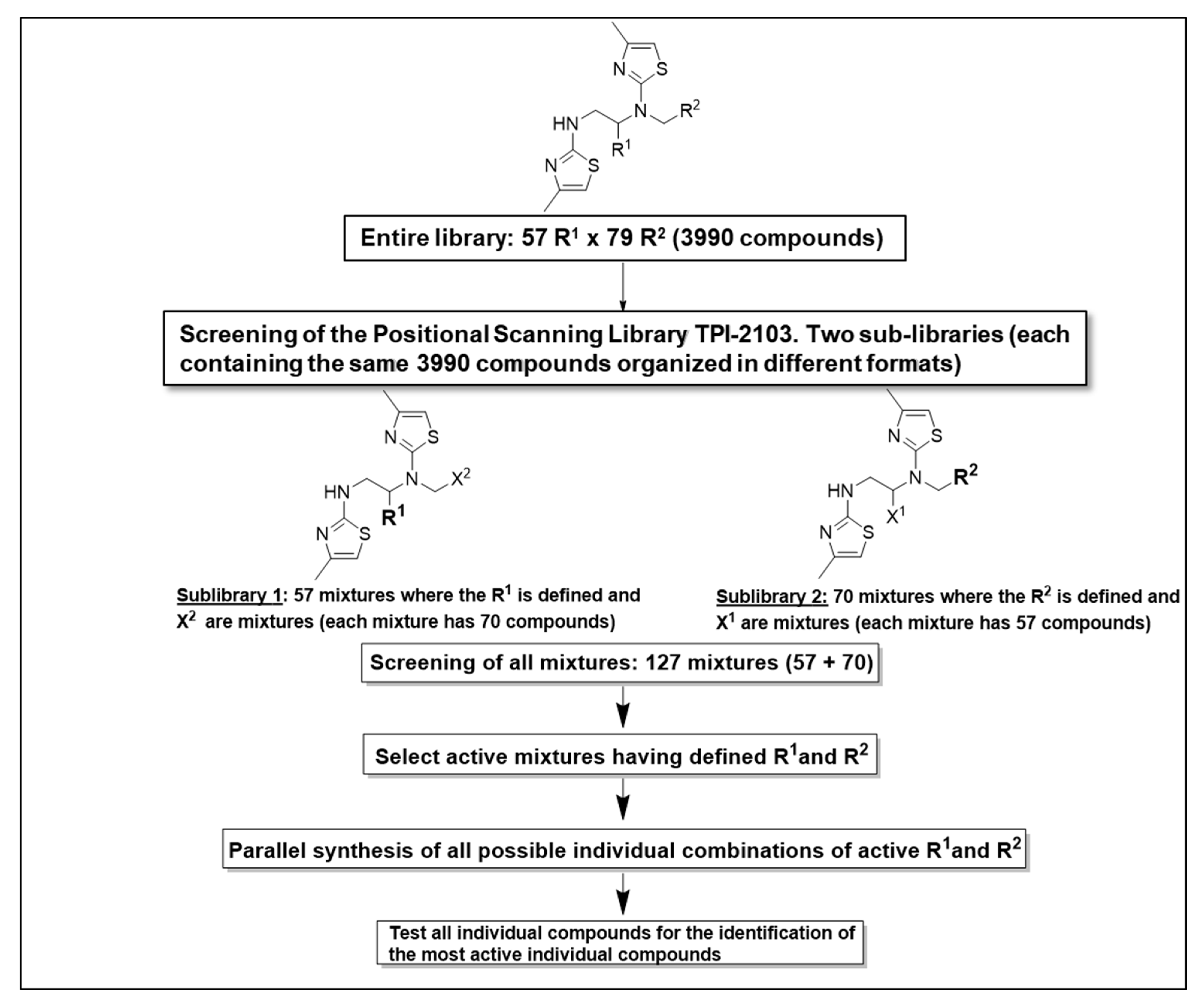
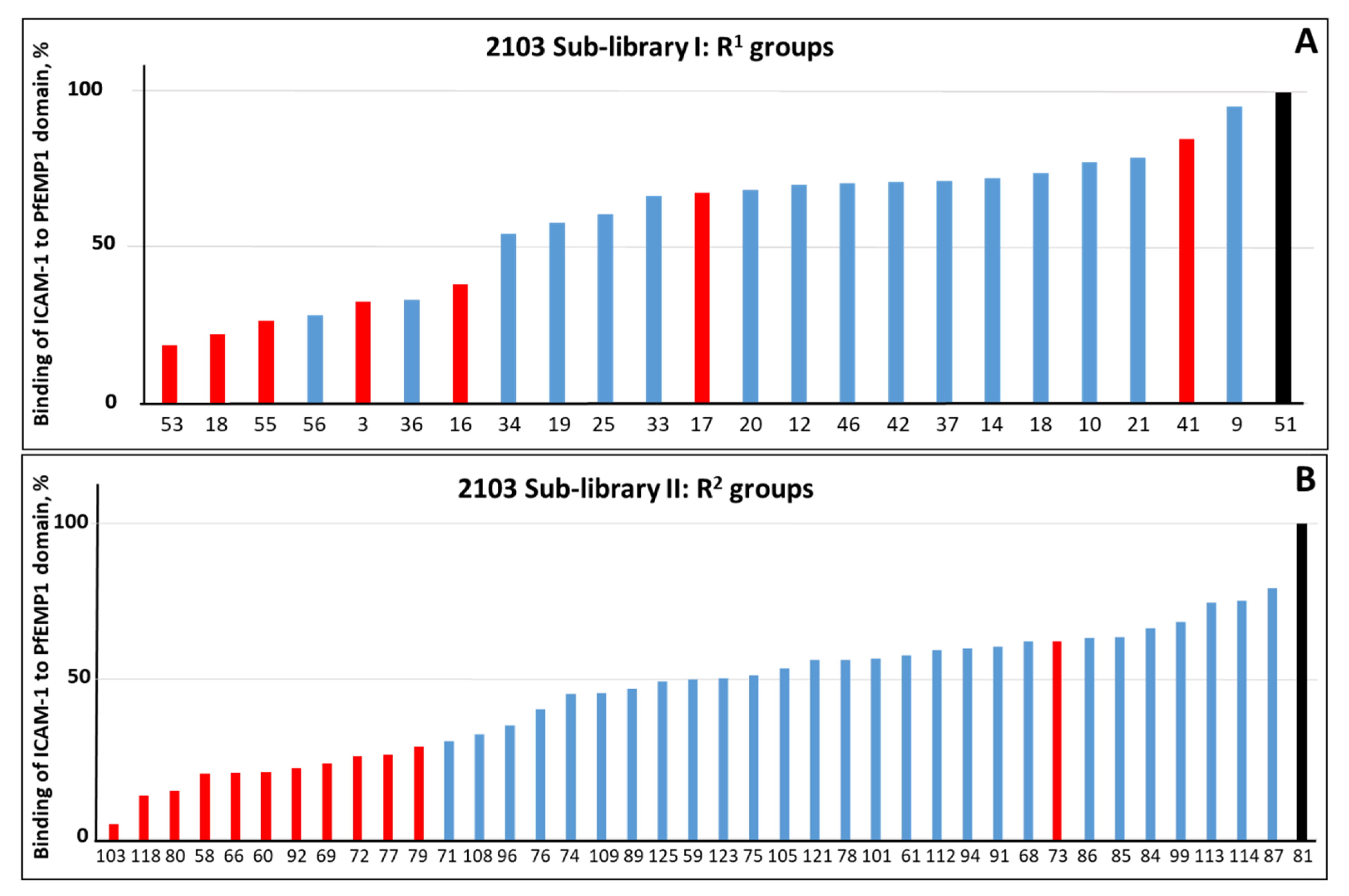
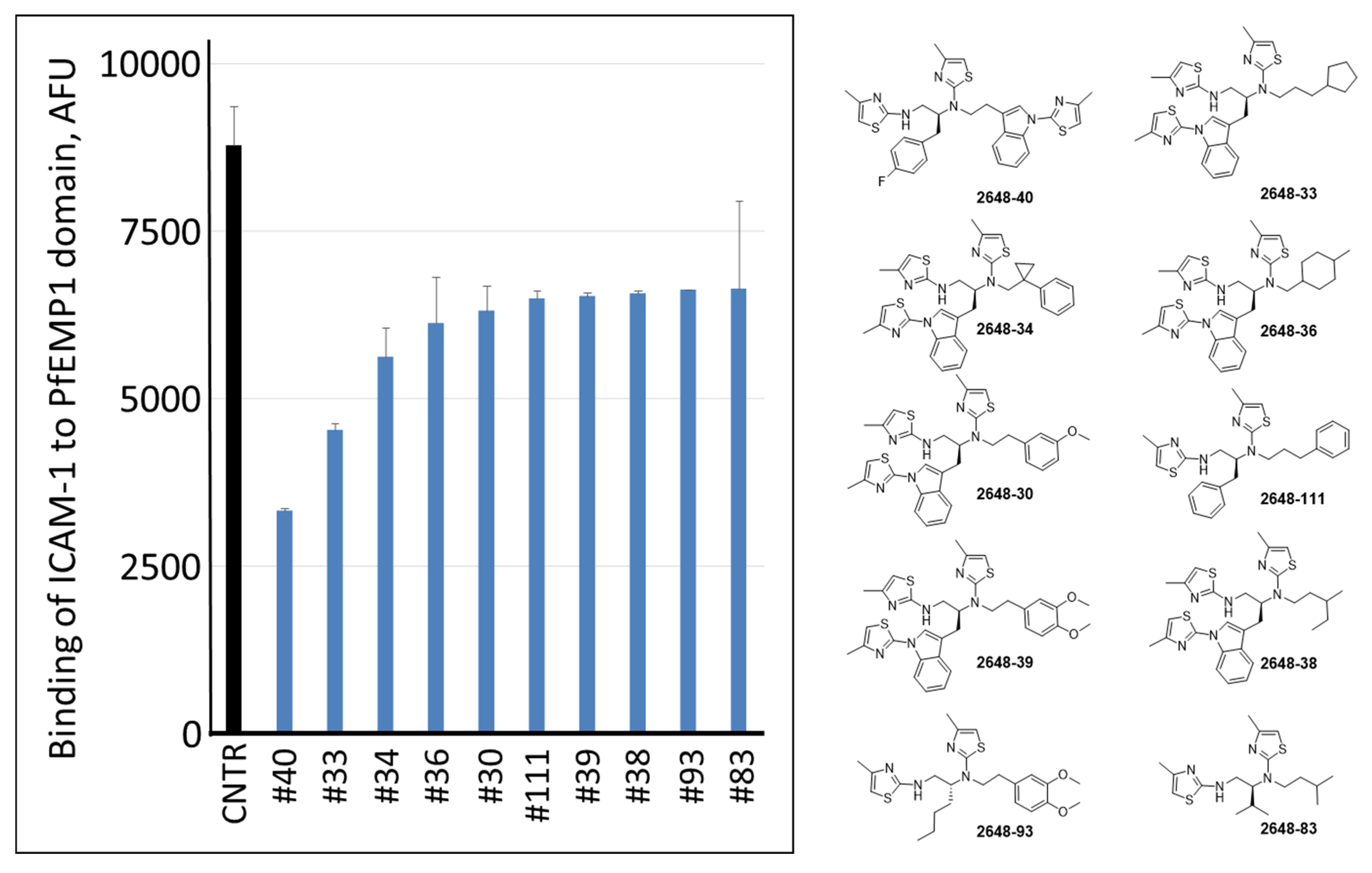
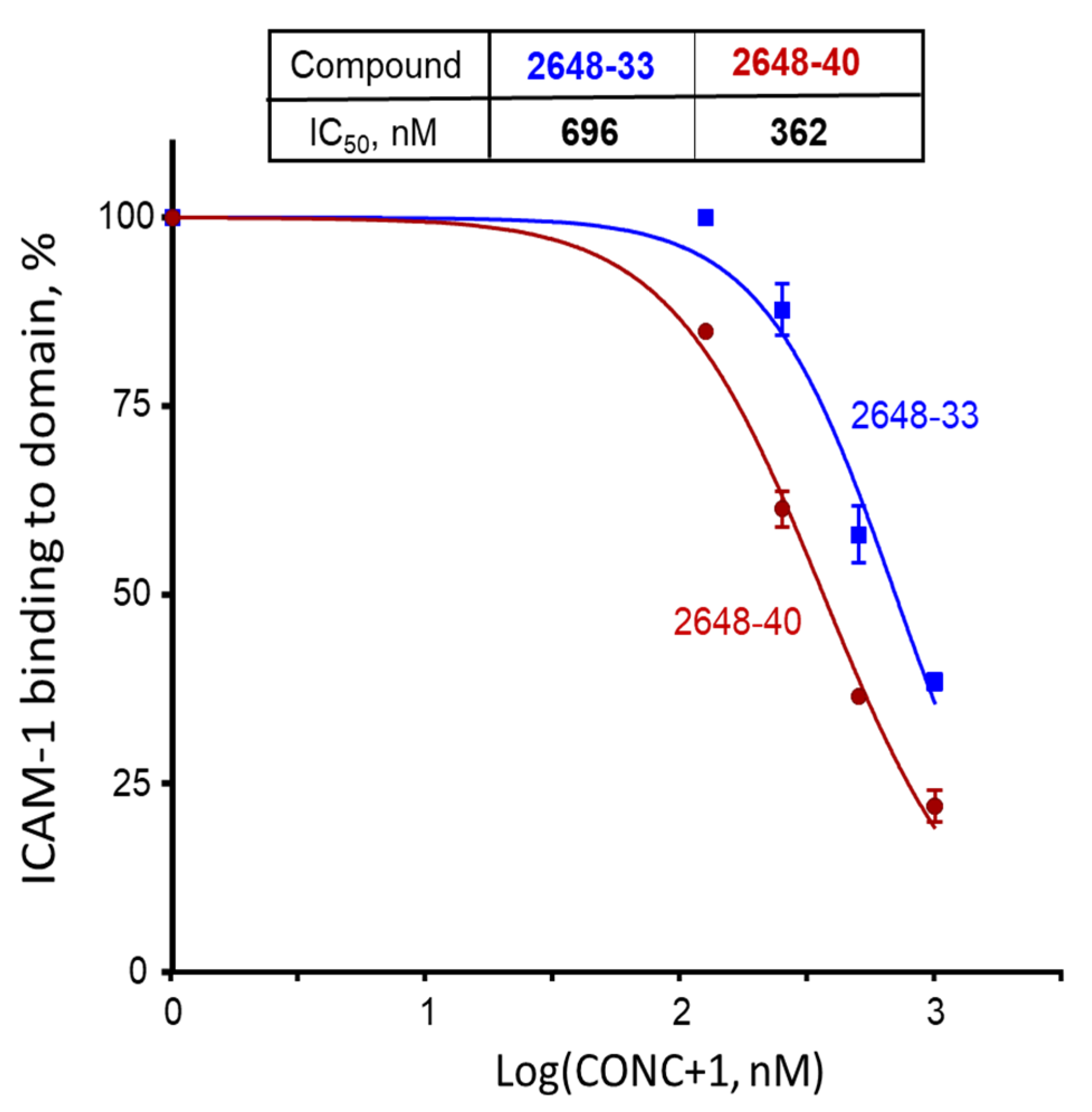
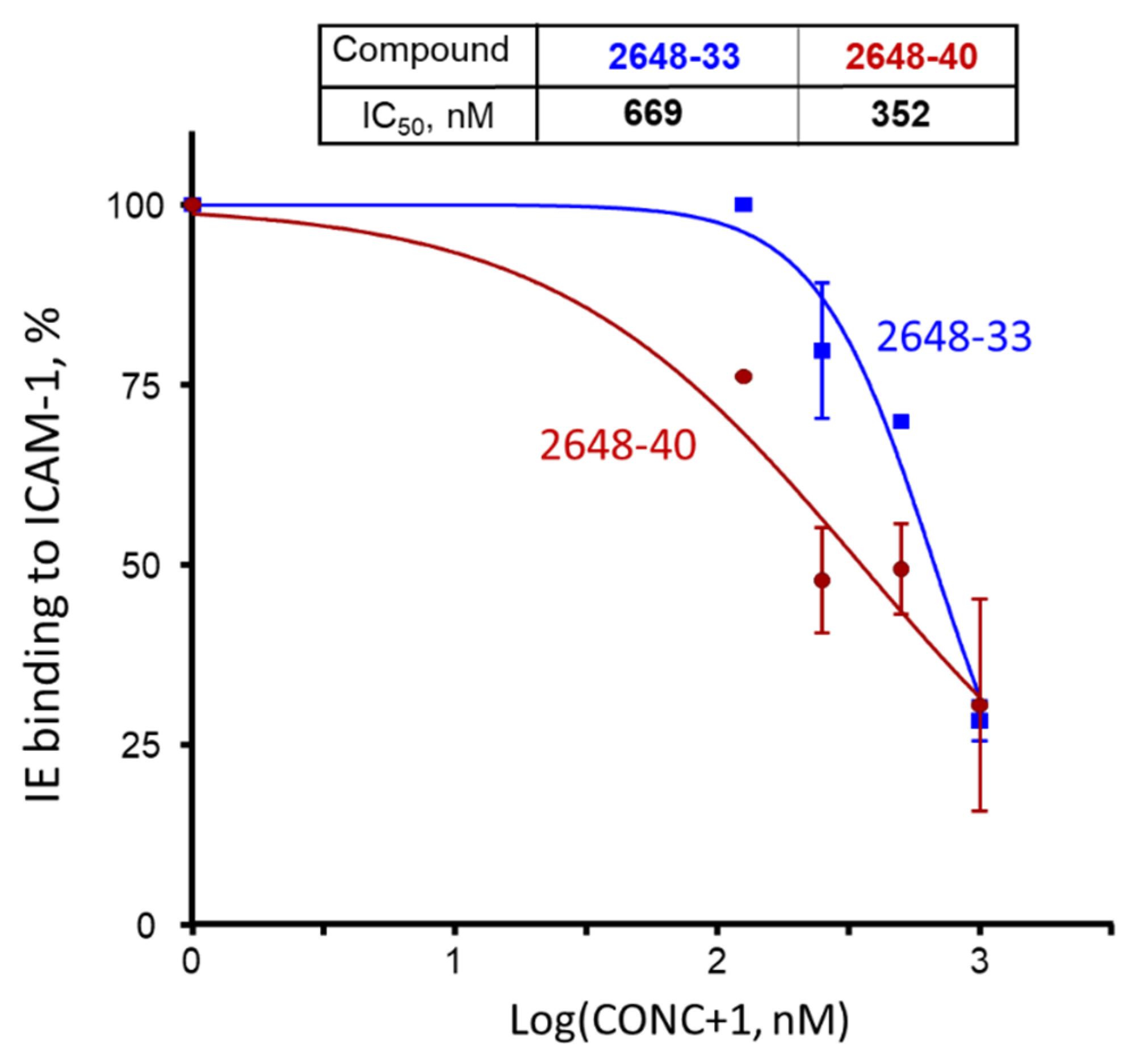
2.1. Screening of the Mixtures Library and Deconvolution of Hit Mixtures to Individual Compounds
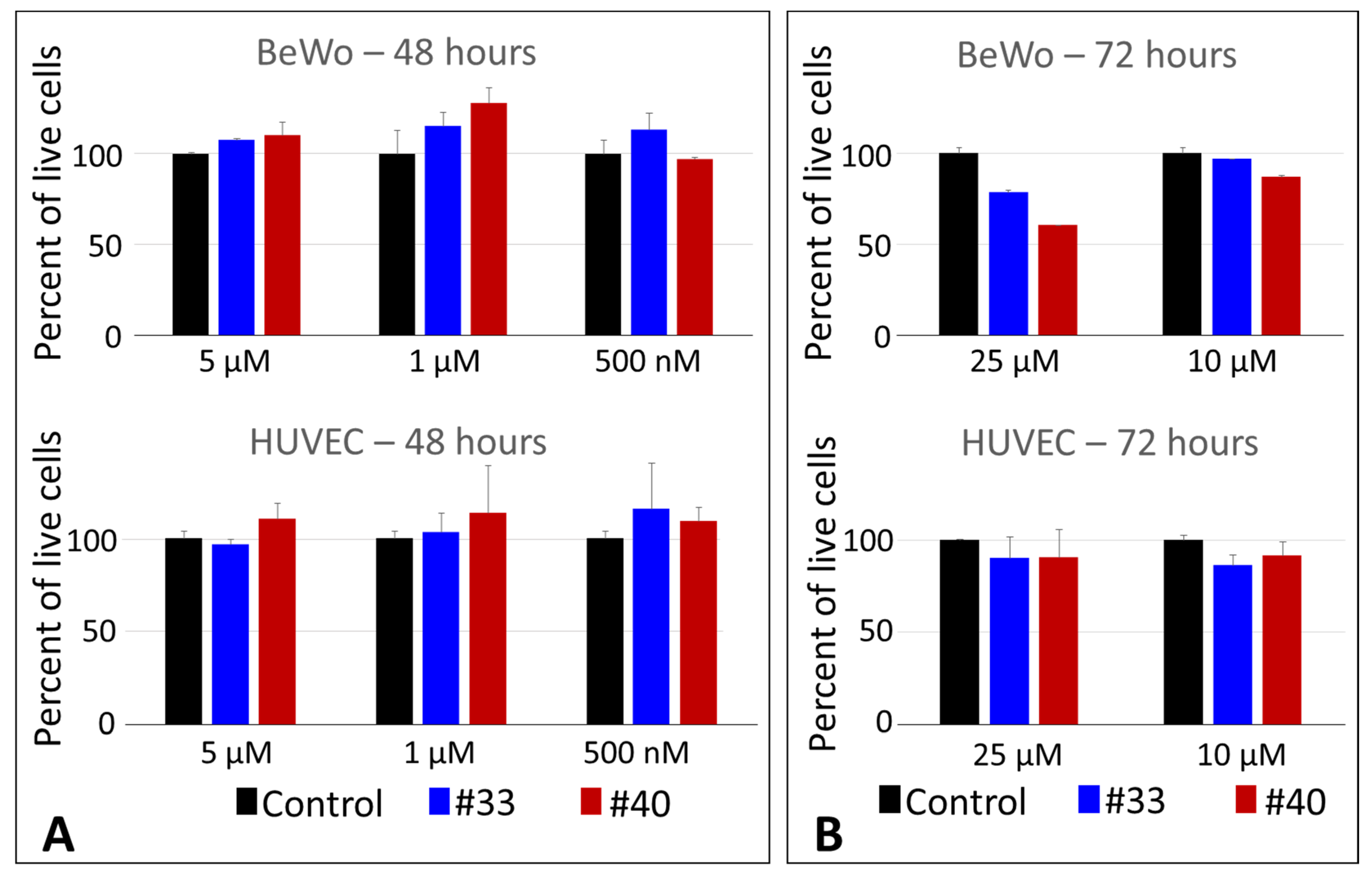
2.2. Cytotoxicity Studies of the Two Most Active Compounds
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. TP-FIU Library of Small Molecules
5.2. Chemistry General
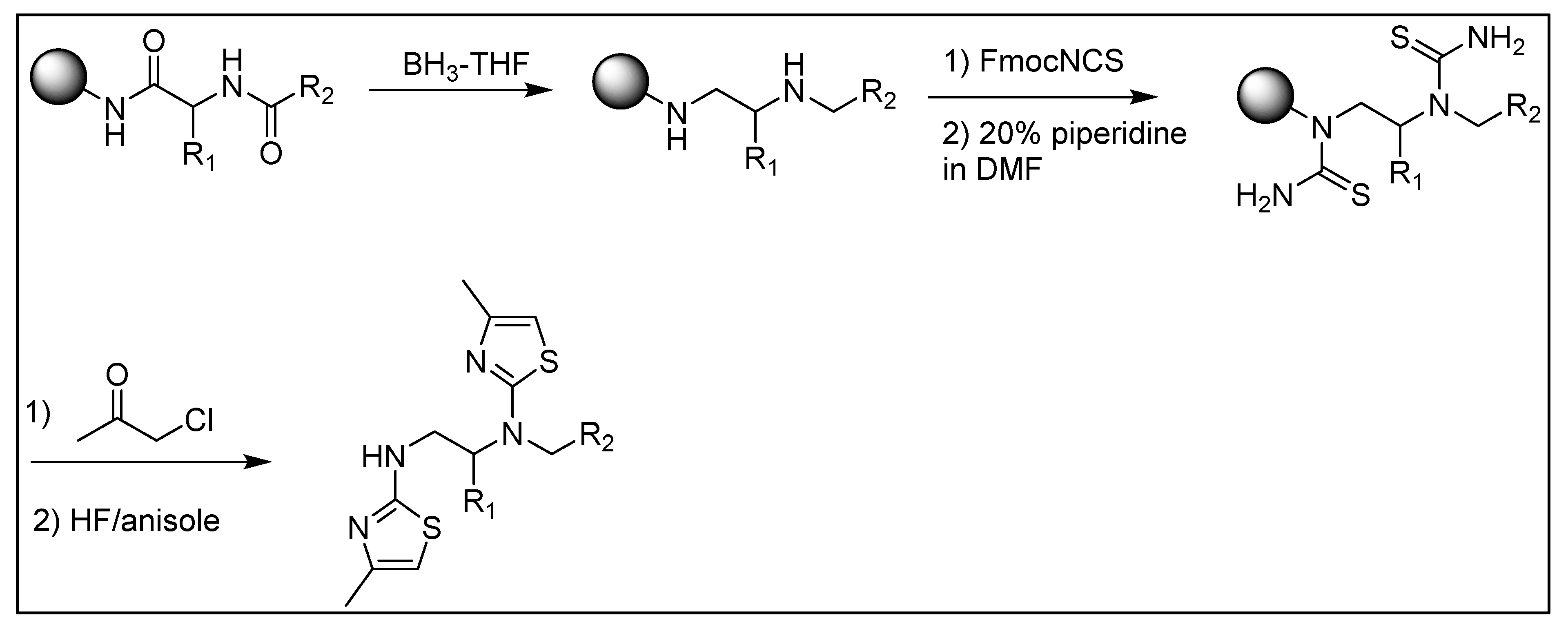
5.3. General Synthesis of the Dithiazole Compounds
5.4. Reversal of ICAM-1 Binding to PF11_0521 DBL2β3 Domain
5.5. Parasite Culture
5.6. Crystal Violet Cell Viability Assay
5.7. Statistical Analyses
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- WHO. World Malaria Report; WHO: Geneva, Switzerland, 2019. [Google Scholar]
- Woodrow, C.J.; White, N.J. The clinical impact of artemisinin resistance in Southeast Asia and the potential for future spread. FEMS Microbiol. Rev. 2017, 41, 34–48. [Google Scholar] [CrossRef] [Green Version]
- Hughes, K.R.; Biagini, G.A.; Craig, A.G. Continued cytoadherence of Plasmodium falciparum infected red blood cells after antimalarial treatment. Mol. Biochem. Parasitol. 2010, 169, 71–78. [Google Scholar] [CrossRef]
- Barnwell, J.W.; Asch, A.S.; Nachman, R.L.; Yamaya, M.; Aikawa, M.; Ingravallo, P. A human 88-kD membrane glycoprotein (CD36) functions in vitro as a receptor for a cytoadherence ligand on Plasmodium falciparum-infected erythrocytes. J. Clin. Investig. 1989, 84, 765–772. [Google Scholar] [CrossRef]
- Berendt, A.R.; Simmons, D.L.; Tansey, J.; Newbold, C.I.; Marsh, K. Intercellular adhesion molecule-1 is an endothelial cell adhesion receptor for Plasmodium falciparum. Nature 1989, 341, 57–59. [Google Scholar] [CrossRef]
- Barragan, A.; Spillmann, D.; Carlson, J.; Wahlgren, M. Role of glycans in Plasmodium falciparum infection. Biochem. Soc. Trans. 1999, 27, 487–493. [Google Scholar]
- Fried, M.; Duffy, P.E. Adherence of Plasmodium falciparum to chondroitin sulfate A in the human placenta. Science 1996, 272, 1502–1504. [Google Scholar] [CrossRef]
- Miller, L.H.; Good, M.F.; Milon, G. Malaria pathogenesis. Science 1994, 264, 1878–1883. [Google Scholar] [CrossRef] [Green Version]
- Newbold, C.; Warn, P.; Black, G.; Berendt, A.; Craig, A.; Snow, B.; Msobo, M.; Peshu, N.; Marsh, K. Receptor-specific adhesion and clinical disease in Plasmodium falciparum. Am. J. Trop. Med. Hyg. 1997, 57, 389–398. [Google Scholar] [CrossRef]
- Turner, G.D.; Morrison, H.; Jones, M.; Davis, T.M.; Looareesuwan, S.; Buley, I.D.; Gatter, K.C.; Newbold, C.I.; Pukritayakamee, S.; Nagachinta, B.; et al. An immunohistochemical study of the pathology of fatal malaria. Evidence for widespread endothelial activation and a potential role for intercellular adhesion molecule-1 in cerebral sequestration. Am. J. Pathol. 1994, 145, 1057–1069. [Google Scholar]
- Adams, S.; Brown, H.; Turner, G. Breaking down the blood-brain barrier: Signaling a path to cerebral malaria? Trends Parasitol. 2002, 18, 360–366. [Google Scholar] [CrossRef]
- Lennartz, F.; Adams, Y.; Bengtsson, A.; Olsen, R.W.; Turner, L.; Ndam, N.T.; Ecklu-Mensah, G.; Moussiliou, A.; Ofori, M.F.; Gamain, B.; et al. Structure-Guided Identification of a Family of Dual Receptor-Binding PfEMP1 that Is Associated with Cerebral Malaria. Cell Host Microbe 2017, 21, 403–414. [Google Scholar] [CrossRef] [Green Version]
- Frank, M.; Deitsch, K. Activation, silencing and mutually exclusive expression within the var gene family of Plasmodium falciparum. Int. J. Parasitol. 2006, 36, 975–985. [Google Scholar] [CrossRef] [PubMed]
- Scherf, A.; Hernandez-Rivas, R.; Buffet, P.; Bottius, E.; Benatar, C.; Pouvelle, B.; Gysin, J.; Lanzer, M. Antigenic variation in malaria: In situ switching, relaxed and mutually exclusive transcription of var genes during intra-erythrocytic development in Plasmodium falciparum. EMBO J. 1998, 17, 5418–5426. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Howell, D.P.; Samudrala, R.; Smith, J.D. Disguising itself--insights into Plasmodium falciparum binding and immune evasion from the DBL crystal structure. Mol. Biochem. Parasitol. 2006, 148, 1–9. [Google Scholar] [CrossRef]
- Howell, D.P.; Levin, E.A.; Springer, A.L.; Kraemer, S.M.; Phippard, D.J.; Schief, W.R.; Smith, J.D. Mapping a common interaction site used by Plasmodium falciparum Duffy binding-like domains to bind diverse host receptors. Mol. Microbiol. 2008, 67, 78–87. [Google Scholar] [CrossRef]
- Oleinikov, A.V.; Amos, E.; Frye, I.T.; Rossnagle, E.; Mutabingwa, T.K.; Fried, M.; Duffy, P.E. High throughput functional assays of the variant antigen PfEMP1 reveal a single domain in the 3D7 Plasmodium falciparum genome that binds ICAM1 with high affinity and is targeted by naturally acquired neutralizing antibodies. PLoS Pathog. 2009, 5, e1000386. [Google Scholar] [CrossRef]
- Bengtsson, A.; Joergensen, L.; Rask, T.S.; Olsen, R.W.; Andersen, M.A.; Turner, L.; Theander, T.G.; Hviid, L.; Higgins, M.K.; Craig, A.; et al. A Novel Domain Cassette Identifies Plasmodium falciparum PfEMP1 Proteins Binding ICAM-1 and Is a Target of Cross-Reactive, Adhesion-Inhibitory Antibodies. J. Immunol. 2013, 190, 240–249. [Google Scholar] [CrossRef] [Green Version]
- Goncalves, B.P.; Huang, C.Y.; Morrison, R.; Holte, S.; Kabyemela, E.; Prevots, D.R.; Fried, M.; Duffy, P.E. Parasite burden and severity of malaria in Tanzanian children. N. Engl. J. Med. 2014, 370, 1799–1808. [Google Scholar] [CrossRef] [Green Version]
- Hogh, B. Clinical and parasitological studies on immunity to Plasmodium falciparum malaria in children. Scand. J. Infect. Dis. Suppl. 1996, 102, 1–53. [Google Scholar]
- Rask, T.S.; Hansen, D.A.; Theander, T.G.; Gorm Pedersen, A.; Lavstsen, T. Plasmodium falciparum erythrocyte membrane protein 1 diversity in seven genomes--divide and conquer. PLoS Comp. Biol. 2010, 6, e1000933. [Google Scholar] [CrossRef] [Green Version]
- Gullingsrud, J.; Saveria, T.; Amos, E.; Duffy, P.E.; Oleinikov, A.V. Structure-function-immunogenicity studies of PfEMP1 domain DBL2betaPF11_0521, a malaria parasite ligand for ICAM-1. PLoS ONE 2013, 8, e61323. [Google Scholar] [CrossRef] [Green Version]
- Oleinikov, A.V.; Voronkova, V.V.; Frye, I.T.; Amos, E.; Morrison, R.; Fried, M.; Duffy, P.E. A Plasma Survey Using 38 PfEMP1 Domains Reveals Frequent Recognition of the Plasmodium falciparum Antigen VAR2CSA among Young Tanzanian Children. PLoS ONE 2012, 7, e31011. [Google Scholar] [CrossRef]
- Tessema, S.K.; Utama, D.; Chesnokov, O.; Hodder, A.N.; Lin, C.S.; Harrison, G.L.A.; Jespersen, J.S.; Petersen, B.; Tavul, L.; Siba, P.; et al. Antibodies to ICAM1-binding PfEMP1-DBLbeta are biomarkers of protective immunity to malaria in a cohort of young children from Papua New Guinea. Infect. Immun. 2018. [Google Scholar] [CrossRef] [Green Version]
- Tuikue Ndam, N.; Moussiliou, A.; Lavstsen, T.; Kamaliddin, C.; Jensen, A.T.R.; Mama, A.; Tahar, R.; Wang, C.W.; Jespersen, J.S.; Alao, J.M.; et al. Parasites Causing Cerebral Falciparum Malaria Bind Multiple Endothelial Receptors and Express EPCR and ICAM-1-Binding PfEMP1. J. Infect. Dis. 2017, 215, 1918–1925. [Google Scholar] [CrossRef] [Green Version]
- Moxon, C.A.; Wassmer, S.C.; Milner, D.A., Jr.; Chisala, N.V.; Taylor, T.E.; Seydel, K.B.; Molyneux, M.E.; Faragher, B.; Esmon, C.T.; Downey, C.; et al. Loss of endothelial protein C receptors links coagulation and inflammation to parasite sequestration in cerebral malaria in African children. Blood 2013, 122, 842–851. [Google Scholar] [CrossRef]
- Avril, M.; Bernabeu, M.; Benjamin, M.; Brazier, A.J.; Smith, J.D. Interaction between Endothelial Protein C Receptor and Intercellular Adhesion Molecule 1 to Mediate Binding of Plasmodium falciparum-Infected Erythrocytes to Endothelial Cells. MBio 2016, 7. [Google Scholar] [CrossRef] [Green Version]
- Azasi, Y.; Lindergard, G.; Ghumra, A.; Mu, J.; Miller, L.H.; Rowe, J.A. Infected erythrocytes expressing DC13 PfEMP1 differ from recombinant proteins in EPCR-binding function. Proc. Natl. Acad. Sci. USA 2018, 115, 1063–1068. [Google Scholar] [CrossRef] [Green Version]
- Laszik, Z.; Mitro, A.; Taylor, F.B., Jr.; Ferrell, G.; Esmon, C.T. Human protein C receptor is present primarily on endothelium of large blood vessels: Implications for the control of the protein C pathway. Circulation 1997, 96, 3633–3640. [Google Scholar] [CrossRef]
- Kessler, A.; Dankwa, S.; Bernabeu, M.; Harawa, V.; Danziger, S.A.; Duffy, F.; Kampondeni, S.D.; Potchen, M.J.; Dambrauskas, N.; Vigdorovich, V.; et al. Linking EPCR-Binding PfEMP1 to Brain Swelling in Pediatric Cerebral Malaria. Cell Host Microbe 2017, 22, 601–614.e5. [Google Scholar] [CrossRef]
- Gullingsrud, J.; Milman, N.; Saveria, T.; Chesnokov, O.; Williamson, K.; Srivastava, A.; Gamain, B.; Duffy, P.E.; Oleinikov, A.V. High throughput screening platform identifies small molecules that prevent sequestration of Plasmodium falciparum-infected erythrocytes. J. Infect. Dis. 2015, 211, 1134–1143. [Google Scholar] [CrossRef]
- Houghten, R.A.; Pinilla, C.; Giulianotti, M.A.; Appel, J.R.; Dooley, C.T.; Nefzi, A.; Ostresh, J.M.; Yu, Y.; Maggiora, G.M.; Medina-Franco, J.L.; et al. Strategies for the use of mixture-based synthetic combinatorial libraries: Scaffold ranking, direct testing in vivo, and enhanced deconvolution by computational methods. J. Comb. Chem. 2008, 10, 3–19. [Google Scholar] [CrossRef]
- Janes, J.H.; Wang, C.P.; Levin-Edens, E.; Vigan-Womas, I.; Guillotte, M.; Melcher, M.; Mercereau-Puijalon, O.; Smith, J.D. Investigating the host binding signature on the Plasmodium falciparum PfEMP1 protein family. PLoS Pathog. 2011, 7, e1002032. [Google Scholar] [CrossRef] [Green Version]
- Hsieh, F.L.; Turner, L.; Bolla, J.R.; Robinson, C.V.; Lavstsen, T.; Higgins, M.K. The structural basis for CD36 binding by the malaria parasite. Nat. Commun. 2016, 7, 12837. [Google Scholar] [CrossRef] [PubMed]
- Fairhurst, R.M.; Wellems, T.E. Modulation of malaria virulence by determinants of Plasmodium falciparum erythrocyte membrane protein-1 display. Curr. Opin. Hematol. 2006, 13, 124–130. [Google Scholar] [CrossRef] [PubMed]
- Ch’ng, J.H.; Moll, K.; Quintana Mdel, P.; Chan, S.C.; Masters, E.; Moles, E.; Liu, J.; Eriksson, A.B.; Wahlgren, M. Rosette-Disrupting Effect of an Anti-Plasmodial Compound for the Potential Treatment of Plasmodium falciparum Malaria Complications. Sci. Rep. 2016, 6, 29317. [Google Scholar] [CrossRef] [Green Version]
- Wu, J.; Zhang, Y.; Maida, L.E.; Santos, R.G.; Welmaker, G.S.; LaVoi, T.M.; Nefzi, A.; Yu, Y.; Houghten, R.A.; Toll, L.; et al. Scaffold ranking and positional scanning utilized in the discovery of nAChR-selective compounds suitable for optimization studies. J. Med. Chem. 2013, 56, 10103–10117. [Google Scholar] [CrossRef] [Green Version]
- Singh, N.; Guha, R.; Giulianotti, M.A.; Pinilla, C.; Houghten, R.A.; Medina-Franco, J.L. Chemoinformatic analysis of combinatorial libraries, drugs, natural products, and molecular libraries small molecule repository. J. Chem. Inf. Model. 2009, 49, 1010–1024. [Google Scholar] [CrossRef] [Green Version]
- Martinez-Mayorga, K.; Medina-Franco, J.L.; Giulianotti, M.A.; Pinilla, C.; Dooley, C.T.; Appel, J.R.; Houghten, R.A. Conformation-opioid activity relationships of bicyclic guanidines from 3D similarity analysis. Bioorg. Med. Chem. 2008, 16, 5932–5938. [Google Scholar] [CrossRef] [Green Version]
- Medina-Franco, J.L.; Martinez-Mayorga, K.; Bender, A.; Marin, R.M.; Giulianotti, M.A.; Pinilla, C.; Houghten, R.A. Characterization of activity landscapes using 2D and 3D similarity methods: Consensus activity cliffs. J. Chem. Inf. Model. 2009, 49, 477–491. [Google Scholar] [CrossRef]
- Medina-Franco, J.L.; Yongye, A.B.; Perez-Villanueva, J.; Houghten, R.A.; Martinez-Mayorga, K. Multitarget structure-activity relationships characterized by activity-difference maps and consensus similarity measure. J. Chem. Inf. Model. 2011, 51, 2427–2439. [Google Scholar] [CrossRef]
- Burrows, J.N.; Duparc, S.; Gutteridge, W.E.; Hooft van Huijsduijnen, R.; Kaszubska, W.; Macintyre, F.; Mazzuri, S.; Möhrle, J.J.; Wells, T.N.C. New developments in anti-malarial target candidate and product profiles. Malar. J. 2017, 16, 26. [Google Scholar] [CrossRef] [Green Version]
- Turner, L.; Lavstsen, T.; Berger, S.S.; Wang, C.W.; Petersen, J.E.; Avril, M.; Brazier, A.J.; Freeth, J.; Jespersen, J.S.; Nielsen, M.A.; et al. Severe malaria is associated with parasite binding to endothelial protein C receptor. Nature 2013, 498, 502–505. [Google Scholar] [CrossRef] [Green Version]
- Olsen, R.W.; Ecklu-Mensah, G.; Bengtsson, A.; Ofori, M.F.; Lusingu, J.P.A.; Castberg, F.C.; Hviid, L.; Adams, Y.; Jensen, A.T.R. Natural and Vaccine-Induced Acquisition of Cross-Reactive IgG-Inhibiting ICAM-1-Specific Binding of a Plasmodium falciparum PfEMP1 Subtype Associated Specifically with Cerebral Malaria. Infect. Immun. 2018, 86. [Google Scholar] [CrossRef] [Green Version]
- Bernabeu, M.; Gunnarsson, C.; Vishnyakova, M.; Howard, C.C.; Nagao, R.J.; Avril, M.; Taylor, T.E.; Seydel, K.B.; Zheng, Y.; Smith, J.D. Binding Heterogeneity of Plasmodium falciparum to Engineered 3D Brain Microvessels Is Mediated by EPCR and ICAM-1. MBio 2019, 10. [Google Scholar] [CrossRef] [Green Version]
- Storm, J.; Jespersen, J.S.; Seydel, K.B.; Szestak, T.; Mbewe, M.; Chisala, N.V.; Phula, P.; Wang, C.W.; Taylor, T.E.; Moxon, C.A.; et al. Cerebral malaria is associated with differential cytoadherence to brain endothelial cells. EMBO Mol. Med. 2019, 11. [Google Scholar] [CrossRef]
- Dietrich, J.B. The adhesion molecule ICAM-1 and its regulation in relation with the blood-brain barrier. J. Neuroimmunol. 2002, 128, 58–68. [Google Scholar] [CrossRef]
- Gimenez, F.; Barraud de Lagerie, S.; Fernandez, C.; Pino, P.; Mazier, D. Tumor necrosis factor alpha in the pathogenesis of cerebral malaria. Cell. Mol. Life Sci. CMLS 2003, 60, 1623–1635. [Google Scholar] [CrossRef]
- Houghten, R.A. General method for the rapid solid-phase synthesis of large numbers of peptides: Specificity of antigen-antibody interaction at the level of individual amino acids. Proc. Natl. Acad. Sci. USA 1985, 82, 5131–5135. [Google Scholar] [CrossRef] [Green Version]
- Nefzi, A.; Giulianotti, M.A.; Houghten, R.A. Solid-phase synthesis of bis-heterocyclic compounds from resin-bound orthogonally protected lysine. J. Comb. Chem. 2001, 3, 68–70. [Google Scholar] [CrossRef]
- Nefzi, A.; Giulianotti, M.A.; Ong, N.A.; Houghten, R.A. Solid-phase synthesis of bis-2-imidazolidinethiones from resin-bound tripeptides. Org. Lett. 2000, 2, 3349–3350. [Google Scholar] [CrossRef] [PubMed]
- Chesnokov, O.; Merritt, J.; Tcherniuk, S.O.; Milman, N.; Oleinikov, A.V. Plasmodium falciparum infected erythrocytes can bind to host receptors integrins alphaVbeta3 and alphaVbeta6 through DBLdelta1_D4 domain of PFL2665c PfEMP1 protein. Sci. Rep. 2018, 8, 17871. [Google Scholar] [CrossRef]
- Tcherniuk, S.O.; Chesnokova, O.; Oleinikov, I.V.; Oleinikov, A.V. Nicotinamide inhibits the growth of P. falciparum and enhances the antimalarial effect of artemisinin, chloroquine and pyrimethamine. Mol. Biochem. Parasitol. 2017, 216, 14–20. [Google Scholar] [CrossRef]
- Tcherniuk, S.O.; Chesnokova, O.; Oleinikov, I.V.; Potopalsky, A.I.; Oleinikov, A.V. Anti-malarial effect of semi-synthetic drug amitozyn. Malar. J. 2015, 14, 425. [Google Scholar] [CrossRef] [PubMed] [Green Version]








Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chesnokov, O.; Visitdesotrakul, P.; Kalani, K.; Nefzi, A.; Oleinikov, A.V. Small Molecule Compounds Identified from Mixture-Based Library Inhibit Binding between Plasmodium falciparum Infected Erythrocytes and Endothelial Receptor ICAM-1. Int. J. Mol. Sci. 2021, 22, 5659. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22115659
Chesnokov O, Visitdesotrakul P, Kalani K, Nefzi A, Oleinikov AV. Small Molecule Compounds Identified from Mixture-Based Library Inhibit Binding between Plasmodium falciparum Infected Erythrocytes and Endothelial Receptor ICAM-1. International Journal of Molecular Sciences. 2021; 22(11):5659. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22115659
Chicago/Turabian StyleChesnokov, Olga, Pimnitah Visitdesotrakul, Komal Kalani, Adel Nefzi, and Andrew V. Oleinikov. 2021. "Small Molecule Compounds Identified from Mixture-Based Library Inhibit Binding between Plasmodium falciparum Infected Erythrocytes and Endothelial Receptor ICAM-1" International Journal of Molecular Sciences 22, no. 11: 5659. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22115659