
High Doses of D-Chiro-Inositol Alone Induce a PCO-Like Syndrome and Other Alterations in Mouse Ovaries

 , ,
, ,
Abstract
:1. Introduction
2. Results
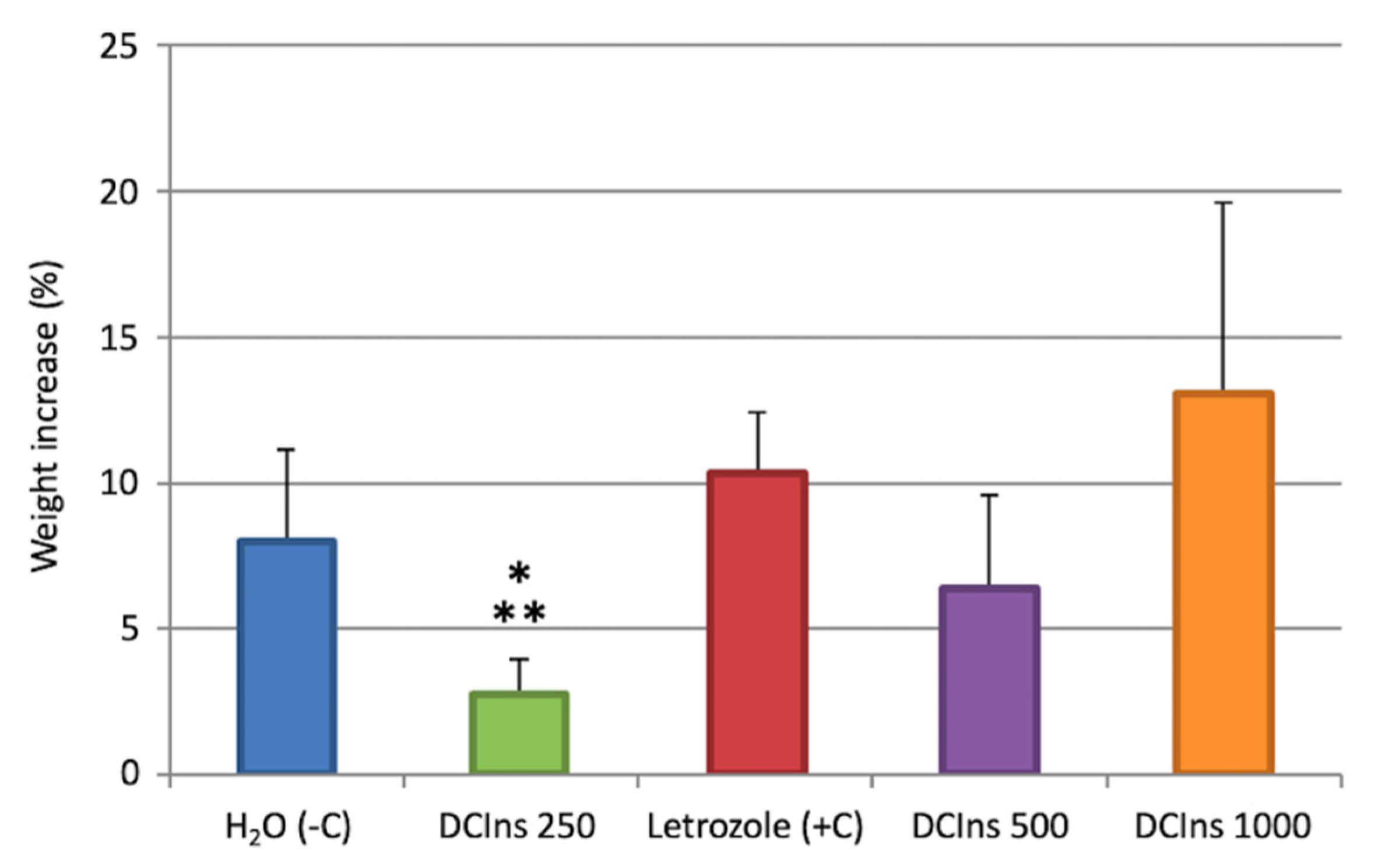
2.1. Increase in Mouse Weights during the Treatment
2.2. Assessment of Cycle Progression during the Treatment
2.3. Gross Morphology of Uteri/Ovaries and Histology of Ovaries at the End of the Treatment
2.4. Follicular Composition of Mouse Ovaries at the End of the Treatment
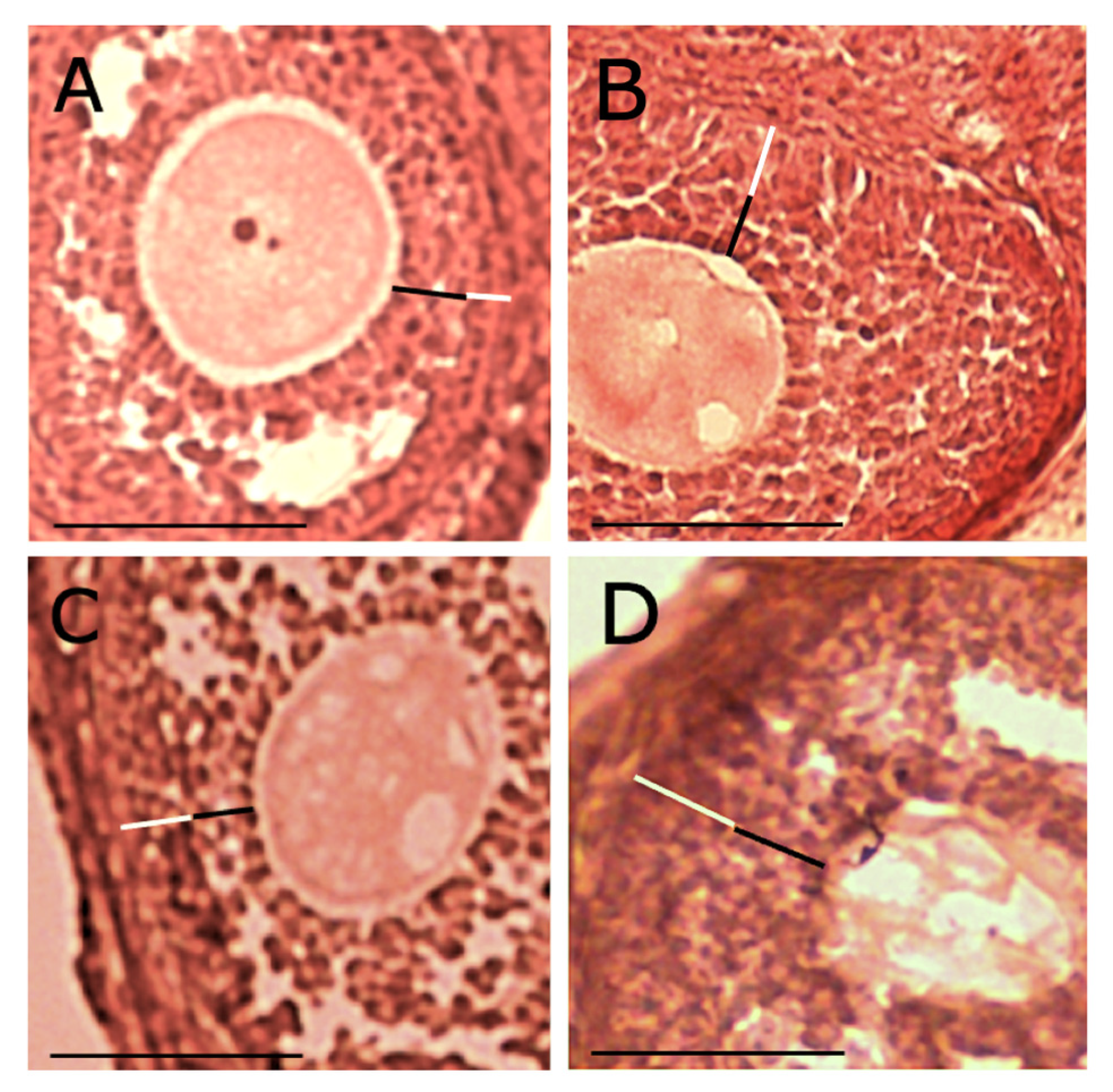
2.5. Theca/granulosa Cell Layer Measurements and Their Ratio
2.6. Testosterone Levels at the End of the Treatment
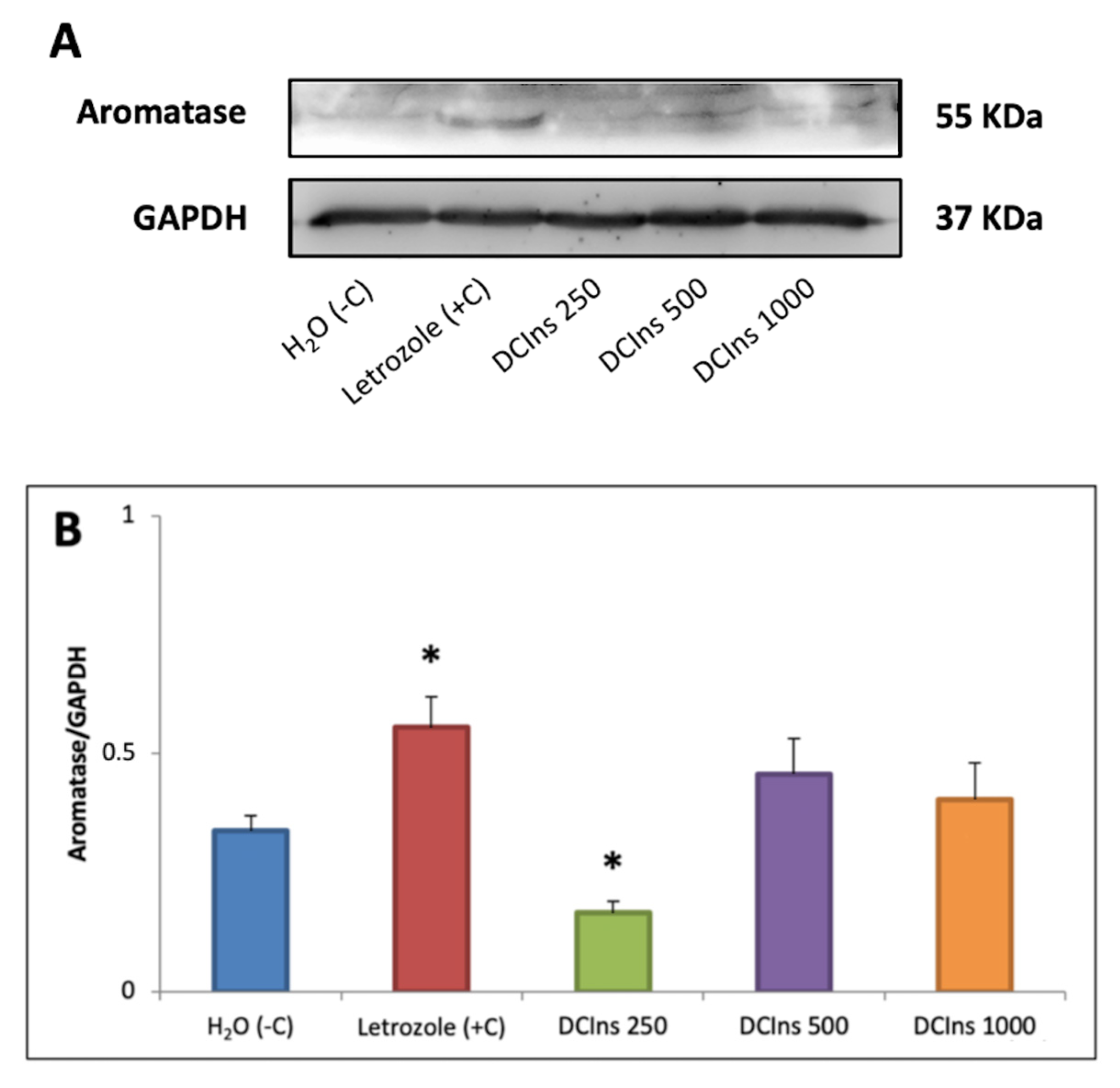
2.7. Presence and Relative Amounts of Aromatase at the End of the Treatment
3. Discussion
3.1. Effects of the Administration of DCIns 250 and Letrozole
3.2. Effects of the Administration of DCIns 500 and DCIns 1000
4. Materials and Methods
4.1. Animals
4.2. Administration of DCIns or Letrozole
4.3. Vaginal Smears
4.4. Blood and Organ Collection
4.5. Histological Analysis
4.6. Serum Testosterone Assay
4.7. Western Blot Analysis of Ovarian Aromatase
4.8. Chemicals
4.9. Statistical Analysis
5. Conclusions
- Exposure of mice to 5 mg/day DCIns for 21 days represents a novel procedure to obtain a useful experimental model of PCOS.
- Exposure of mice to higher daily amounts of DCIns for 21 days is toxic for ovarian histology and function, producing lesions different from those typical of PCOS but resembling a pre-menopausal/menopausal state.
- Serum testosterone levels are affected by administration of DCIns and letrozole. They are increased by 5 mg/day DCIns, but strongly decreased by higher DCIns amounts, probably due to a blockade in steroidogenesis produced by these doses.
- The amounts of ovarian aromatase are affected by administration of both 5 mg/day DCIns and letrozole but in opposite direction: DCIns downregulates and letrozole upregulates aromatase expression, confirming previous observations. Higher DCIns doses do not affect the amount of ovarian aromatase.
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Azziz, R. PCOS: Animal models for PCOS—Not the real thing. Nat. Rev. Endocrinol. 2017, 13, 382–384. [Google Scholar] [CrossRef]
- Vink, J.M.; Sadrzadeh, S.; Lambalk, C.B.; Boomsma, D.I. Heritability of polycystic ovary syndrome in a Dutch twin-family study. J. Clin. Endocrinol. Metab. 2006, 91, 2100–2104. [Google Scholar] [CrossRef] [Green Version]
- Diamanti-Kandarakis, E.; Kandarakis, H.; Legro, R.S. The role of genes and environment in the etiology of PCOS. Endocrine 2006, 30, 19–26. [Google Scholar] [CrossRef]
- Carmina, E.; Legro, R.S.; Stamets, K.; Lowell, J.; Lobo, R.A. Difference in body weight between American and Italian women with polycystic ovary syndrome: Influence of the diet. Hum. Reprod. 2003, 18, 2289–2293. [Google Scholar] [CrossRef] [Green Version]
- Oakley, O.; Lin, P.C.; Bridges, P.; Ko, C. Animal models for the study of polycystic ovarian syndrome. Endocrinol. Metab. 2011, 26, 193–202. [Google Scholar] [CrossRef]
- Walters, K.A.; Allan, C.M.; Handelsman, D.J. Rodent models for human polycystic ovary syndrome. Biol. Reprod. 2012, 86, 1–12. [Google Scholar]
- Shi, D.; Vine, D.F. Animal models of polycystic ovary syndrome: A focused review of rodent models in relationship to clinical phenotypes and cardiometabolic risk. Fertil. Steril. 2012, 98, 185–193. [Google Scholar] [CrossRef]
- Van Houten, E.L.; Visser, J.A. Mouse models to study polycystic ovary syndrome: A possible link between metabolism and ovarian function? Reprod. Biol. 2014, 14, 32–43. [Google Scholar] [CrossRef] [PubMed]
- Paixão, L.; Ramos, R.B.; Lavarda, A.; Morsh, D.M.; Spritzer, P.M. Animal models of hyperandrogenism and ovarian morphology changes as features of polycystic ovary syndrome: A systematic review. Reprod. Biol. Endocrinol. 2017, 15, 12. [Google Scholar] [CrossRef] [Green Version]
- Di Emidio, G.; Placidi, M.; Rea, F.; Rossi, G.; Falone, S.; Cristiano, L.; Nottola, S.; D’Alessandro, A.M.; Amicarelli, F.; Palmerini, M.G.; et al. Methylglyoxal-Dependent Glycative Stress and Deregulation of SIRT1 Functional Network in the Ovary of PCOS Mice. Cells 2020, 9, 209. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Di Emidio, G.; Rea, F.; Placidi, M.; Rossi, G.; Cocciolone, D.; Virmani, A.; Macchiarelli, G.; Palmerini, M.G.; D’Alessandro, A.M.; Artini, P.G.; et al. Regulatory Functions of L-Carnitine, Acetyl, and Propionyl L-Carnitine in a PCOS Mouse Model: Focus on Antioxidant/Antiglycative Molecular Pathways in the Ovarian Microenvironment. Antioxidants 2020, 9, 867. [Google Scholar] [CrossRef] [PubMed]
- Kafali, H.; Iriadam, M.; Ozardali, I.; Demir, N. Letrozole-induced polycystic ovaries in the rat: A new model for cystic ovarian disease. Arch. Med. Res. 2004, 35, 103–108. [Google Scholar] [CrossRef]
- Kauffman, A.S.; Thackray, V.G.; Ryan, G.E.; Tolson, K.P.; Glidewell-Kenney, C.A.; Semaan, S.J.; Poling, M.C.; Iwata, N.; Breen, K.M.; Duleba, A.J.; et al. A Novel Letrozole Model Recapitulates Both the Reproductive and Metabolic Phenotypes of Polycystic Ovary Syndrome in Female Mice. Biol. Reprod. 2015, 93, 69. [Google Scholar] [CrossRef] [PubMed]
- Lawton, I.E.; Schwartz, N.B. Pituitary-ovarian function in rats exposed to constant light: A chronological study. Endocrinology 1967, 81, 497–508. [Google Scholar] [CrossRef]
- Takeo, Y. Influence of continuous illumination on estrous cycle of rats: Time course of changes in levels of gonadotropins and ovarian steroids until occurrence of persistent estrus. Neuroendocrinol 1984, 39, 97–104. [Google Scholar] [CrossRef] [PubMed]
- Bevilacqua, A.; Dragotto, J.; Giuliani, A.; Bizzarri, M. Myo-inositol and D-chiro-inositol (40:1) reverse histological and functional features of polycystic ovary syndrome in a mouse model. J. Cell. Physiol. 2019, 234, 9387–9398. [Google Scholar] [CrossRef]
- Azziz, R.; Carmina, E.; Dewailly, D.; Diamanti-Kandarakis, E.; Escobar-Morreale, H.F.; Futterweit, W.; Janssen, O.E.; Legro, R.S.; Norman, R.J.; Taylor, A.E.; et al. The androgen excess and PCOS Society criteria for the polycystic ovary syndrome: The complete task force report. Fertil. Steril. 2009, 91, 456–488. [Google Scholar] [CrossRef]
- Diamanti-Kandarakis, E.; Papavassiliou, A.G. Molecular mechanisms of insulin resistance in polycystic ovary syndrome. Trends Mol. Med. 2006, 12, 324–332. [Google Scholar] [CrossRef]
- Heimark, D.; McAllister, J.; Larner, J. Decreased myo-inositol to chiro-inositol (M/C) ratios and increased M/C epimerase activity in PCOS theca cells demonstrated increased insulin sensitivity compared to controls. Endocr. J. 2014, 61, 111–117. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mato, J.M.; Kelly, K.L.; Abler, A.; Jarett, L. Identification of a novel insulin-sensitive glycophospholipid from H35 hepatoma cells. J. Biol. Chem. 1987, 262, 2131–2137. [Google Scholar] [CrossRef]
- Low, M.G.; Saltiel, A.R. Structural and functional roles of glycosyl-phosphatidylinositol in membranes. Science 1988, 239, 268–275. [Google Scholar] [CrossRef] [PubMed]
- Baillargeon, J.P.; Nestler, J.E.; Ostlund, R.E.; Apridonidze, T.; Diamanti- Kandarakis, E. Greek hyperinsulinemic women, with or without polycystic ovary syndrome, display altered inositols metabolism. Hum. Reprod. 2008, 23, 1439–1446. [Google Scholar] [CrossRef] [PubMed]
- Colazingari, S.; Treglia, M.; Najjar, R.; Bevilacqua, A. The combined therapy myo-inositol plus D-chiro-inositol, rather than D- chiroinositol, is able to improve IVF outcomes: Results from a randomized controlled trial. Arch. Gynecol. Obst. 2013, 288, 1405–1411. [Google Scholar] [CrossRef]
- Dinicola, S.; Chiu, T.T.Y.; Unfer, V.; Carlomagno, G.; Bizzarri, M. The rationale of the myo-inositol and D-chiro-inositol combined treatment for polycystic ovary syndrome. J. Clin. Pharmacol. 2014, 54, 1079–1092. [Google Scholar] [CrossRef] [PubMed]
- Facchinetti, F.; Bizzarri, M.; Benvenga, S.; D’Anna, R.; Lanzone, A.; Soulage, C.; Di Renzo, G.C.; Hod, M.; Cavalli, P.; Chiu, T.T.; et al. Results from the International Consensus Conference on Myo-inositol and d-chiro-inositol in Obstetrics and Gynecology: The link between metabolic syndrome and PCOS. Eur. J. Obst. Gynecol. Reprod. Biol. 2015, 195, 72–76. [Google Scholar] [CrossRef]
- Facchinetti, F.; Appetecchia, M.; Aragona, C.; Bevilacqua, A.; Bezerra Espinola, M.S.; Bizzarri, M.; D’Anna, R.; Dewailly, D.; Diamanti-Kandarakis, E.; Hernández Marín, I.; et al. Experts’ opinion on inositols in treating polycystic ovary syndrome and non-insulin dependent diabetes mellitus: A further help for human reproduction and beyond. Expert Opin. Drug Metab. Toxicol. 2020, 16, 255–274. [Google Scholar] [CrossRef]
- Facchinetti, F.; Dante, G.; Neri, I. The ratio of MI to DCI and its impact in the treatment of polycystic ovary syndrome: Experimental and literature evidences. Front. Gynecol. Endocrinol. 2015, 3, 103–109. [Google Scholar]
- Nordio, M.; Basciani, S.; Camajani, E. The 40:1 myo-inositol/D-chiro-inositol plasma ratio is able to restore ovulation in PCOS patients: Comparison with other ratios. Eur. Rev. Med. Pharmacol. Sci. 2019, 23, 5512–5521. [Google Scholar] [PubMed]
- Unfer, V.; Carlomagno, G.; Papaleo, E.; Vailati, S.; Candiani, M.; Baillargeon, J.P. Hyperinsulinemia alters myoinositol to d-chiroinositol ratio in the follicular fluid of patients with PCOS. Reprod. Sci. 2014, 21, 854–858. [Google Scholar] [CrossRef]
- Ravanos, K.; Monastra, G.; Pavlidou, T.; Goudakou, M.; Prapas, N. Can high levels of D-chiro- inositol in follicular fluid exert detrimental effects on blastocyst quality? Eur. Rev. Med. Pharmacol. Sci. 2017, 21, 5491–5498. [Google Scholar]
- Minini, M.; Monastra, G.; Dinicola, S. A double-edge sword: The role of D-chiro-inositol in oocyte and embryo quality. Int. J. Med. Dev. Adjust. Treat. 2019, 2, e208. [Google Scholar]
- Sacchi, S.; Marinaro, F.; Tondelli, D.; Lui, J.; Xella, S.; Marsella, T.; Tagliasacchi, D.; Argento, C.; Tirelli, A.; Giulini, S.; et al. Modulation of gonadotrophin induced steroidogenic enzymes in granulosa cells by d-chiroinositol. Reprod. Biol. Endocrinol. 2016, 14, 52. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nair, A.; Morsy, M.A.; Jacob, S. Dose translation between laboratory animals and human in preclinical and clinical phases of drug development. Drug Dev. Res. 2018, 79, 373–382. [Google Scholar] [CrossRef]
- La Marca, A.; Grisendi, V.; Dondi, G.; Sighinolfi, G.; Cianci, A. The menstrual cycle regularization following D-chiro-inositol treatment in PCOS women: A retrospective study. Gynecol. Endocrinol. 2015, 1, 52–56. [Google Scholar] [CrossRef] [PubMed]
- Gambioli, R.; Forte, G.; Aragona, C.; Bevilacqua, A.; Bizzarri, M.; Unfer, V. The use of D-chiro-Inositol in clinical practice. Eur. Rev. Med. Pharmacol. Sci. 2021, 25, 438–446. [Google Scholar]
- Veloz, A.; Reyes-Vázquez, L.; Patricio-Gómez, J.M.; Romano, M.C. Effect of mice Taenia crassiceps WFU cysticerci infection on the ovarian folliculogenesis, enzyme expression, and serum estradiol. Exp. Parasitol. 2019, 207, 107778. [Google Scholar] [CrossRef] [PubMed]
- Mori, T.; Ito, F.; Koshiba, A.; Kataoka, H.; Tanaka, Y.; Okimura, H.; Khan, K.N.; Kitawaki, J. Aromatase as a target for treating endometriosis. J. Obst. Gynaecol. Res. 2018, 44, 1673–1681. [Google Scholar] [CrossRef] [Green Version]
- Kakadia, N.; Patel, P.; Deshpande, S.; Shah, G. Effect of Vitex negundo L. seeds in letrozole induced polycystic ovarian syndrome. J. Tradit. Complement. Med. 2018, 9, 336–345. [Google Scholar] [CrossRef]
- Amini, L.; Tehranian, N.; Movahedin, M.; Ramezani Tehrani, F.; Soltanghoraee, H. Polycystic ovary morphology (PCOM) in estradiol valerate treated mouse model. Int. J. Womens Health Reprod. Sci. 2016, 4, 13–17. [Google Scholar] [CrossRef] [Green Version]
- Jeon, Y.; Aja, S.; Ronnett, G.V.; Kim, E.-K. D-chiro-inositol glycan reduces food intake by regulating hypothalamic neuropeptide expression via AKT-FoxO1 pathway. Biochem. Biophys. Res. Commun. 2016, 470, 818–823. [Google Scholar] [CrossRef] [PubMed]
- Hipps, P.P.; Sehgal, R.K.; Holland, W.H.; Sherman, W.R. Identification and partial characterization of inositol: NAD+ epimerase and inosose: NAD(P)H reductase from the fat body of the American cockroach, Periplaneta americana L. Biochemistry 1973, 12, 4507–4512. [Google Scholar] [CrossRef] [PubMed]
- Huang, L.C.; Fonteles, M.C.; Houston, D.B.; Zhang, C.; Larner, J. Chiroinositol deficiency and insulin resistance. III. Acute glycogenic and hypoglycemic effects of two inositol phosphoglycan insulin mediators in normal and streptozotocin-diabetic rats in vivo. Endocrinology 1993, 132, 652–657. [Google Scholar] [CrossRef] [PubMed]
- Pak, Y.; Huang, L.C.; Lilley, K.J.; Larner, J. In vivo conversion of [3H]myoinositol to [3H]chiroinositol in rat tissues. J. Biol. Chem. 1992, 267, 16904–16910. [Google Scholar] [CrossRef]
- Pak, Y.; Hong, Y.; Kim, S.; Piccariello, T.; Farese, R.V.; Larner, J. In vivo chiro-inositol metabolism in the rat: A defect in chiro-inositol synthesis from myo-inositol and an increased incorporation of chiro-[3H]inositol into phospholipid in the Goto-Kakizaki (G.K) rat. Mol. Cells 1998, 8, 301–309. [Google Scholar] [PubMed]
- Bizzarri, M.; Fuso, A.; Dinicola, S.; Cucina, A.; Bevilacqua, A. Pharmacodynamics and pharmacokinetics of inositol(s) in health and disease. Expert Opin. Drug Metab. Toxicol. 2016, 12, 1181–1196. [Google Scholar] [CrossRef] [PubMed]
- Lewin, L.M.; Yannai, Y.; Melmed, S.; Weiss, M. Myo-inositol in the reproductive tract of the female rat. Int. J. Biochem. 1982, 14, 147–150. [Google Scholar] [CrossRef]
- Chiu, T.T.; Rogers, M.S.; Briton-Jones, C.; Haines, C. Effects of myo-inositol on the in-vitro maturation and subsequent development of mouse oocytes. Hum. Reprod. 2003, 18, 408–416. [Google Scholar] [CrossRef] [Green Version]
- Colazingari, S.; Fiorenza, M.T.; Carlomagno, G.; Najjar, R.; Bevilacqua, A. Improvement of mouse embryo quality by myo-inositol supplementation of IVF media. J. Assist. Reprod. Genet. 2014, 31, 463–469. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kuşcu, N.; Bizzarri, M.; Bevilacqua, A. Myo-Inositol Safety in Pregnancy: From Preimplantation Development to Newborn Animals. Int. J. Endocrinol. 2016, 2016, 2413857. [Google Scholar] [CrossRef] [Green Version]
- Milewska, E.M.; Czyzyk, A.; Meczekalski, B.; Genazzani, A.D. Inositol and human reproduction. From cellular metabolism to clinical use. Gynecol. Endocrinol. 2016, 32, 690–695. [Google Scholar] [CrossRef] [PubMed]
- Matsuda, M.; Tsutsumi, K.; Kanematsu, T.; Fukami, K.; Terada, Y.; Takenawa, T.; Nakayama, K.I.; Hirata, M. Involvement of phospholipase C-related inactive protein in the mouse reproductive system through the regulation of gonadotropin levels. Biol. Reprod. 2009, 81, 681–689. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zacchè, M.M.; Caputo, L.; Filippis, S.; Zacchè, G.; Dindelli, M.; Ferrari, A. Efficacy of myo-inositol in the treatment of cutaneous disorders in young women with polycystic ovary syndrome. Gynecol. Endocrinol. 2009, 25, 508–513. [Google Scholar] [CrossRef]
- Lowther, K.M.; Weitzman, V.N.; Maier, D.; Mehlmann, L.M. Maturation, fertilization, and the structure and function of the endoplasmic reticulum in cryopreserved mouse oocytes. Biol. Reprod. 2009, 81, 147–154. [Google Scholar] [CrossRef]
- Nestler, J.E.; Jakubowicz, D.J.; Reamer, P.; Gunn, R.D.; Allan, G. Ovulatory and metabolic effects of D-chiro-inositol in the polycystic ovary syndrome. New Engl. J. Med. 1999, 340, 1314–1320. [Google Scholar] [CrossRef] [Green Version]
- Nestler, J.E.; Jakubowicz, D.J.; de Vargas, A.F.; Brik, C.; Quintero, N.; Medina, F. Insulin stimulates testosterone biosynthesis by human thecal cells from women with polycystic ovary syndrome by activating its own receptor and using inositolglycan mediators as the signal transduction system. J. Clin. Endocrinol. Metab. 1998, 83, 2001–2005. [Google Scholar]
- Smith, E.R.; Yeasky, T.; Wei, J.Q.; Miki, R.A.; Cai, K.Q.; Smedberg, J.L.; Yang, W.L.; Xu, X.X. White spotting variant mouse as an experimental model for ovarian aging and menopausal biology. Menopause 2012, 19, 588–596. [Google Scholar] [CrossRef] [Green Version]
- Pericuesta, E.; Laguna-Barraza, R.; Ramos-Ibeas, P.; Gutierrez-Arroyo, J.L.; Navarro, J.A.; Vera, K.; Sanjuan, C.; Baixeras, E.; de Fonseca, F.R.; Gutierrez-Adan, A. D-Chiro-Inositol Treatment Affects Oocyte and Embryo Quality and Improves Glucose Intolerance in Both Aged Mice and Mouse Models of Polycystic Ovarian Syndrome. Int. J. Mol. Sci. 2020, 21, 6049. [Google Scholar] [CrossRef]
- Laviolette, L.A.; Ethier, J.F.; Senterman, M.K.; Devine, P.J.; Vanderhyden, B.C. Induction of a menopausal state alters the growth and histology of ovarian tumors in a mouse model of ovarian cancer. Menopause 2011, 18, 549–557. [Google Scholar] [CrossRef] [PubMed]
- Long, G.G.; Cohen, I.R.; Gries, C.L.; Young, J.K.; Francis, P.C.; Capen, C.C. Proliferative lesions of ovarian granulosa cells and reversible hormonal changes induced in rats by a selective estrogen receptor modulator. Toxicol. Pathol. 2001, 29, 403–410, Erratum in 2001, 29, 719–726. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Caligioni, C.S. Assessing reproductive status/stages in mice. Curr. Protoc. Neurosci. 2009, 48, A.4I.1–A.4I.8. [Google Scholar] [CrossRef] [Green Version]
- Harlow, E.; Lane, D. Antibodies: A Laboratory Manual; Cold Spring Harbor Laboratory: Huntington, NY, USA, 1988; p. 449. [Google Scholar]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| H20 (−C) | Letrozole (+C) | DCIns 250 | DCIns 500 | ||||
|---|---|---|---|---|---|---|---|
| TC | GC | TC | GC | TC | GC | TC | GC |
| 0.101 | 0.23 | 0.106 | 0.109 | 0.1 | 0.121 | 0.115 | 0.083 |
| 0.116 | 0.22 | 0.1 | 0.081 | 0.11 | 0.093 | 0.096 | 0.078 |
| 0.097 | 0.21 | 0.098 | 0.088 | 0.097 | 0.103 | 0.105 | 0.088 |
| 0.11 | 0.255 | 0.09 | 0.075 | 0.131 | 0.12 | 0.11 | 0.081 |
| 0.12 | 0.26 | 0.095 | 0.071 | 0.124 | 0.129 | 0.114 | 0.093 |
| 0.135 | 0.25 | 0.102 | 0.108 | 0.107 | 0.097 | 0.1 | 0.086 |
| TGR | |||||||
| Mean | 0.48 | 1.13 * | 1.02 * | 1.26 *, ** | |||
| SD | 0.05 | 0.15 | 0.13 | 0.09 | |||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bevilacqua, A.; Dragotto, J.; Lucarelli, M.; Di Emidio, G.; Monastra, G.; Tatone, C. High Doses of D-Chiro-Inositol Alone Induce a PCO-Like Syndrome and Other Alterations in Mouse Ovaries. Int. J. Mol. Sci. 2021, 22, 5691. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22115691
Bevilacqua A, Dragotto J, Lucarelli M, Di Emidio G, Monastra G, Tatone C. High Doses of D-Chiro-Inositol Alone Induce a PCO-Like Syndrome and Other Alterations in Mouse Ovaries. International Journal of Molecular Sciences. 2021; 22(11):5691. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22115691
Chicago/Turabian StyleBevilacqua, Arturo, Jessica Dragotto, Micaela Lucarelli, Giovanna Di Emidio, Giovanni Monastra, and Carla Tatone. 2021. "High Doses of D-Chiro-Inositol Alone Induce a PCO-Like Syndrome and Other Alterations in Mouse Ovaries" International Journal of Molecular Sciences 22, no. 11: 5691. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22115691