
Extracellular Vesicles: Potential Mediators of Psychosocial Stress Contribution to Osteoporosis?


Abstract
:1. Introduction
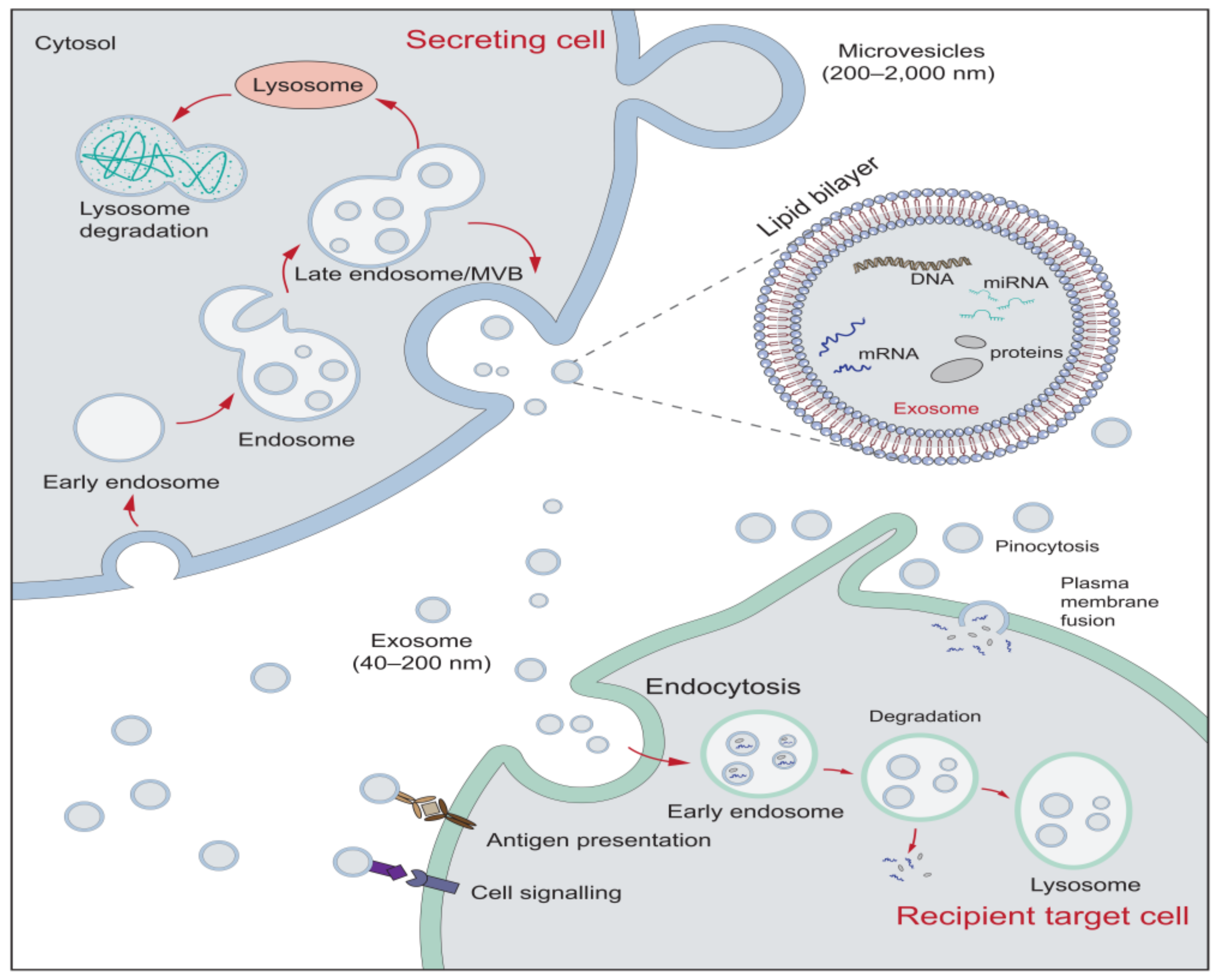
2. The Characteristics of Extracellular Vesicles
3. The Role of EVs in the Stress Response
3.1. EVs May Serve as Biomarkers for Psychosocial Stress
3.2. Stress Modifies miRNAs in EVs to Regulate the Immune Response
4. The Role of EVs in Osteoporosis
4.1. Overview of Osteoporosis and Bone Remodeling
4.2. EVs Regulate Osteoclasts Differentiation and Activity
4.3. EVs Affect Osteoblasts and Osteogenic Function
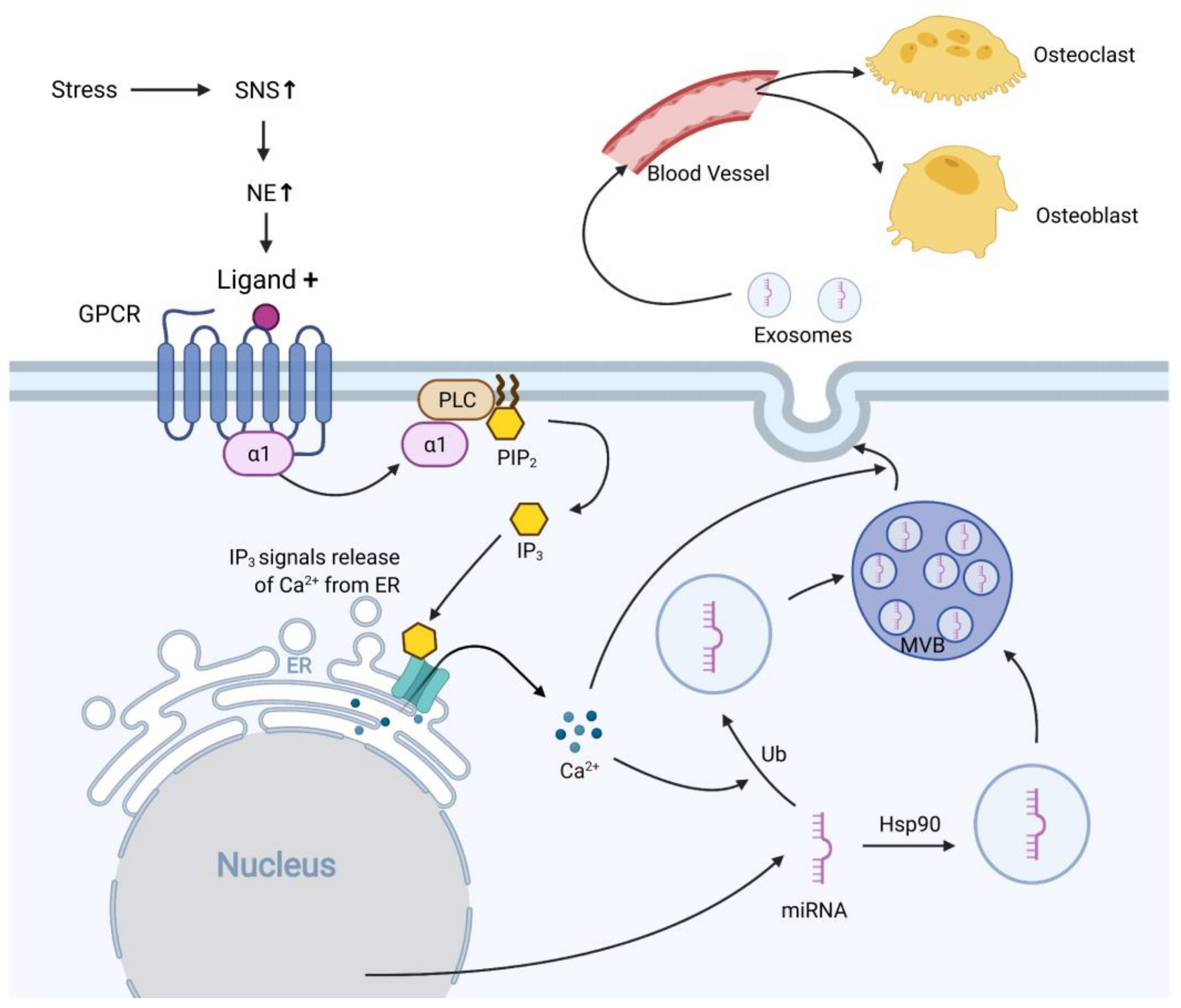
5. EVs as Potential Mediators of Psychosocial Stress and Osteoporosis
6. Conclusions and Perspective
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| BMSC | Bone marrow mesenchymal stem cell |
| CUMS | Chronic unpredictable mild stress |
| DAMPs | Danger-associated molecular patterns |
| ER | Endoplasmic reticulum |
| EVs | Extracellular vesicles |
| GPCR | G-protein coupled receptor |
| HSP | heat-shock protein |
| IP3 | Inositol trisphosphate |
| miRNA, miR | microRNA |
| MSCs | Mesenchymal stem cells |
| mtDNA | mitochondrial DNA |
| MVBs | Multivesicular bodies |
| PIP2 | Phosphatidylinositol bisphosphate |
| PLC | Phospholipase c |
| RANK | Receptor activator of nuclear factor κ-Β |
| RANKL | Receptor activator of nuclear factor κ-Β ligand |
| ROS | Reactive oxygen species |
| Runx2 | Runt-related transcription factor 2 |
| SNS | Sympathetic nervous system |
| SOCS-3 | Suppressor of cytokine signaling-3 |
| Ub | Ubiquitination |
| UTR | Untranslated region |
| Wnt | Wingless and Int-1 |
References
- Chrousos, G.P. Stress and Disorders of the Stress System. Nat. Rev. Endocrinol. 2009, 5, 374–381. [Google Scholar] [CrossRef] [PubMed]
- Nicolaides, N.C.; Kyratzi, E.; Lamprokostopoulou, A.; Chrousos, G.P.; Charmandari, E. Stress, the Stress System and the Role of Glucocorticoids. Neuroimmunomodulation 2015, 22, 6–19. [Google Scholar] [CrossRef] [PubMed]
- Chaudhuri, J.; Bains, Y.; Guha, S.; Kahn, A.; Hall, D.; Bose, N.; Gugliucci, A.; Kapahi, P. The Role of Advanced Glycation End Products in Aging and Metabolic Diseases: Bridging Association and Causality. Cell Metab. 2018, 28, 337–352. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fleming, T.H.; Humpert, P.M.; Nawroth, P.P.; Bierhaus, A. Reactive Metabolites and AGE/RAGE-Mediated Cellular Dysfunction Affect the Aging Process—A Mini-Review. Gerontology 2011, 57, 435–443. [Google Scholar] [CrossRef]
- McEwen, B.S.; Stellar, E. Stress and the Individual. Mechanisms Leading to Disease. Arch. Intern. Med. 1993, 153, 2093–2101. [Google Scholar] [CrossRef]
- NIH Consensus Development Panel on Osteoporosis Prevention, Diagnosis, and Therapy Osteoporosis Prevention, Diagnosis, and Therapy. JAMA 2001, 285, 785–795. [CrossRef]
- Riancho, J.A.; Brennan-Olsen, S.L. The Epigenome at the Crossroad between Social Factors, Inflammation, and Osteoporosis Risk. Clin. Rev. Bone Miner. Metab. 2017, 15, 59–68. [Google Scholar] [CrossRef] [Green Version]
- Gieseler, F.; Ender, F. Extracellular Vesicles and Cell–Cell Communication: New Insights and New Therapeutic Strategies Not Only in Oncology. Int. J. Mol. Sci. 2020, 21, 4331. [Google Scholar] [CrossRef]
- Bellavia, D.; Raimondi, L.; Costa, V.; De Luca, A.; Carina, V.; Maglio, M.; Fini, M.; Alessandro, R.; Giavaresi, G. Engineered Exosomes: A New Promise for the Management of Musculoskeletal Diseases. Biochim. Biophys. Acta BBA Gen. Subj. 2018, 1862, 1893–1901. [Google Scholar] [CrossRef]
- Fleshner, M.; Crane, C.R. Exosomes, DAMPs and MiRNA: Features of Stress Physiology and Immune Homeostasis. Trends Immunol. 2017, 38, 768–776. [Google Scholar] [CrossRef]
- Rubartelli, A.; Lotze, M.T.M.; Latz, E.; Manfredi, A.A. Mechanisms of Sterile Inflammation. Front. Immunol. 2013, 4. [Google Scholar] [CrossRef] [Green Version]
- Roh, J.S.; Sohn, D.H. Damage-Associated Molecular Patterns in Inflammatory Diseases. Immune Netw. 2018, 18. [Google Scholar] [CrossRef]
- Cristóvão, J.S.; Gomes, C.M. S100 Proteins in Alzheimer’s Disease. Front. Neurosci. 2019, 13. [Google Scholar] [CrossRef]
- Boyapati, R.K.; Dorward, D.A.; Tamborska, A.; Kalla, R.; Ventham, N.T.; Doherty, M.K.; Whitfield, P.D.; Gray, M.; Loane, J.; Rossi, A.G.; et al. Mitochondrial DNA Is a Pro-Inflammatory Damage-Associated Molecular Pattern Released during Active IBD. Inflamm. Bowel Dis. 2018, 24, 2113–2122. [Google Scholar] [CrossRef]
- Hernandez, C.; Huebener, P.; Pradere, J.-P.; Friedman, R.A.; Schwabe, R.F. HMGB1 Links Chronic Liver Injury to Progenitor Responses and Hepatocarcinogenesis. J. Clin. Investig. 2019, 128, 2436–2450. [Google Scholar] [CrossRef]
- Collett, G.P.; Redman, C.W.; Sargent, I.L.; Vatish, M. Endoplasmic Reticulum Stress Stimulates the Release of Extracellular Vesicles Carrying Danger-Associated Molecular Pattern (DAMP) Molecules. Oncotarget 2018, 9, 6707–6717. [Google Scholar] [CrossRef] [Green Version]
- Xie, X.; Xiong, Y.; Panayi, A.C.; Hu, L.; Zhou, W.; Xue, H.; Lin, Z.; Chen, L.; Yan, C.; Mi, B.; et al. Exosomes as a Novel Approach to Reverse Osteoporosis: A Review of the Literature. Front. Bioeng. Biotechnol. 2020, 8. [Google Scholar] [CrossRef]
- Colombo, M.; Raposo, G.; Théry, C. Biogenesis, Secretion, and Intercellular Interactions of Exosomes and Other Extracellular Vesicles. Annu. Rev. Cell Dev. Biol. 2014, 30, 255–289. [Google Scholar] [CrossRef]
- Théry, C.; Witwer, K.W.; Aikawa, E.; Alcaraz, M.J.; Anderson, J.D.; Andriantsitohaina, R.; Antoniou, A.; Arab, T.; Archer, F.; Atkin-Smith, G.K.; et al. Minimal Information for Studies of Extracellular Vesicles 2018 (MISEV2018): A Position Statement of the International Society for Extracellular Vesicles and Update of the MISEV2014 Guidelines. J. Extracell. Vesicles 2018, 7, 1535750. [Google Scholar] [CrossRef] [Green Version]
- Van Niel, G.; D’Angelo, G.; Raposo, G. Shedding Light on the Cell Biology of Extracellular Vesicles. Nat. Rev. Mol. Cell Biol. 2018, 19, 213–228. [Google Scholar] [CrossRef]
- Shao, H.; Im, H.; Castro, C.M.; Breakefield, X.; Weissleder, R.; Lee, H. New Technologies for Analysis of Extracellular Vesicles. Chem. Rev. 2018, 118, 1917–1950. [Google Scholar] [CrossRef]
- Alzhrani, G.N.; Alanazi, S.T.; Alsharif, S.Y.; Albalawi, A.M.; Alsharif, A.A.; Abdel-Maksoud, M.S.; Elsherbiny, N. Exosomes: Isolation, Characterization, and Biomedical Applications. Cell Biol. Int. 2021, 1–25. [Google Scholar] [CrossRef]
- Crescitelli, R.; Lässer, C.; Szabó, T.G.; Kittel, A.; Eldh, M.; Dianzani, I.; Buzás, E.I.; Lötvall, J. Distinct RNA Profiles in Subpopulations of Extracellular Vesicles: Apoptotic Bodies, Microvesicles and Exosomes. J. Extracell. Vesicles 2013, 2. [Google Scholar] [CrossRef]
- Mathieu, M.; Martin-Jaular, L.; Lavieu, G.; Théry, C. Specificities of Secretion and Uptake of Exosomes and Other Extracellular Vesicles for Cell-to-Cell Communication. Nat. Cell Biol. 2019, 21, 9–17. [Google Scholar] [CrossRef]
- Pegtel, D.M.; Gould, S.J. Exosomes. Annu. Rev. Biochem. 2019, 88, 487–514. [Google Scholar] [CrossRef]
- Gong, M.; Yu, B.; Wang, J.; Wang, Y.; Liu, M.; Paul, C.; Millard, R.W.; Xiao, D.-S.; Ashraf, M.; Xu, M. Mesenchymal Stem Cells Release Exosomes That Transfer MiRNAs to Endothelial Cells and Promote Angiogenesis. Oncotarget 2017, 8, 45200–45212. [Google Scholar] [CrossRef] [Green Version]
- Wu, R.; Gao, W.; Yao, K.; Ge, J. Roles of Exosomes Derived from Immune Cells in Cardiovascular Diseases. Front. Immunol. 2019, 10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ahmadi, M.; Rezaie, J. Tumor Cells Derived-Exosomes as Angiogenenic Agents: Possible Therapeutic Implications. J. Transl. Med. 2020, 18, 249. [Google Scholar] [CrossRef] [PubMed]
- Goetzl, E.J.; Goetzl, L.; Karliner, J.S.; Tang, N.; Pulliam, L. Human Plasma Platelet-Derived Exosomes: Effects of Aspirin. FASEB J. 2016, 30, 2058–2063. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, H.; Wang, Z. Cardiomyocyte-Derived Exosomes: Biological Functions and Potential Therapeutic Implications. Front. Physiol. 2019, 10. [Google Scholar] [CrossRef]
- Wang, J.; Zheng, Y.; Zhao, M. Exosome-Based Cancer Therapy: Implication for Targeting Cancer Stem Cells. Front. Pharmacol. 2017, 7. [Google Scholar] [CrossRef] [Green Version]
- Kalluri, R. The Biology and Function of Exosomes in Cancer. J. Clin. Investig. 2016, 126, 1208–1215. [Google Scholar] [CrossRef]
- Sanz-Rubio, D.; Martin-Burriel, I.; Gil, A.; Cubero, P.; Forner, M.; Khalyfa, A.; Marin, J.M. Stability of Circulating Exosomal MiRNAs in Healthy Subjects. Sci. Rep. 2018, 8, 10306. [Google Scholar] [CrossRef] [Green Version]
- Bartel, D.P. MicroRNAs: Genomics, Biogenesis, Mechanism, and Function. Cell 2004, 116, 281–297. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.; Li, S.; Li, L.; Li, M.; Guo, C.; Yao, J.; Mi, S. Exosome and Exosomal MicroRNA: Trafficking, Sorting, and Function. Genom. Proteom. Bioinform. 2015, 13, 17–24. [Google Scholar] [CrossRef] [Green Version]
- Bellavia, D.; De Luca, A.; Carina, V.; Costa, V.; Raimondi, L.; Salamanna, F.; Alessandro, R.; Fini, M.; Giavaresi, G. Deregulated MiRNAs in Bone Health: Epigenetic Roles in Osteoporosis. Bone 2019, 122, 52–75. [Google Scholar] [CrossRef]
- Pethő, A.; Chen, Y.; George, A. Exosomes in Extracellular Matrix Bone Biology. Curr. Osteoporos. Rep. 2018, 16, 58–64. [Google Scholar] [CrossRef]
- Wang, J.; Veirman, K.D.; Faict, S.; Frassanito, M.A.; Ribatti, D.; Vacca, A.; Menu, E. Multiple Myeloma Exosomes Establish a Favourable Bone Marrow Microenvironment with Enhanced Angiogenesis and Immunosuppression. J. Pathol. 2016, 239, 162–173. [Google Scholar] [CrossRef]
- Aslan, C.; Maralbashi, S.; Salari, F.; Kahroba, H.; Sigaroodi, F.; Kazemi, T.; Kharaziha, P. Tumor-Derived Exosomes: Implication in Angiogenesis and Antiangiogenesis Cancer Therapy. J. Cell. Physiol. 2019, 234, 16885–16903. [Google Scholar] [CrossRef]
- Foessl, I.; Kotzbeck, P.; Obermayer-Pietsch, B. MiRNAs as Novel Biomarkers for Bone Related Diseases. J. Lab. Precis. Med. 2019, 4. [Google Scholar] [CrossRef]
- Gao, M.; Gao, W.; Papadimitriou, J.M.; Zhang, C.; Gao, J.; Zheng, M. Exosomes—the Enigmatic Regulators of Bone Homeostasis. Bone Res. 2018, 6, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Raimondi, L.; De Luca, A.; Fontana, S.; Amodio, N.; Costa, V.; Carina, V.; Bellavia, D.; Raimondo, S.; Siragusa, S.; Monteleone, F.; et al. Multiple Myeloma-Derived Extracellular Vesicles Induce Osteoclastogenesis through the Activation of the XBP1/IRE1α Axis. Cancers 2020, 12, 2167. [Google Scholar] [CrossRef] [PubMed]
- Patil, M.; Henderson, J.; Luong, H.; Annamalai, D.; Sreejit, G.; Krishnamurthy, P. The Art of Intercellular Wireless Communications: Exosomes in Heart Disease and Therapy. Front. Cell Dev. Biol. 2019, 7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Milano, G.; Biemmi, V.; Lazzarini, E.; Balbi, C.; Ciullo, A.; Bolis, S.; Ameri, P.; Di Silvestre, D.; Mauri, P.; Barile, L.; et al. Intravenous Administration of Cardiac Progenitor Cell-Derived Exosomes Protects against Doxorubicin/Trastuzumab-Induced Cardiac Toxicity. Cardiovasc. Res. 2020, 116, 383–392. [Google Scholar] [CrossRef]
- Patel, N.A.; Moss, L.D.; Lee, J.-Y.; Tajiri, N.; Acosta, S.; Hudson, C.; Parag, S.; Cooper, D.R.; Borlongan, C.V.; Bickford, P.C. Long Noncoding RNA MALAT1 in Exosomes Drives Regenerative Function and Modulates Inflammation-Linked Networks Following Traumatic Brain Injury. J. Neuroinflamm. 2018, 15, 204. [Google Scholar] [CrossRef] [Green Version]
- Song, Y.; Li, Z.; He, T.; Qu, M.; Jiang, L.; Li, W.; Shi, X.; Pan, J.; Zhang, L.; Wang, Y.; et al. M2 Microglia-Derived Exosomes Protect the Mouse Brain from Ischemia-Reperfusion Injury via Exosomal MiR-124. Theranostics 2019, 9, 2910–2923. [Google Scholar] [CrossRef]
- Goetzl, L.; Merabova, N.; Darbinian, N.; Martirosyan, D.; Poletto, E.; Fugarolas, K.; Menkiti, O. Diagnostic Potential of Neural Exosome Cargo as Biomarkers for Acute Brain Injury. Ann. Clin. Transl. Neurol. 2018, 5, 4–10. [Google Scholar] [CrossRef] [Green Version]
- Lv, L.-L.; Feng, Y.; Tang, T.-T.; Liu, B.-C. New Insight into the Role of Extracellular Vesicles in Kidney Disease. J. Cell. Mol. Med. 2019, 23, 731–739. [Google Scholar] [CrossRef]
- Baghaei, K.; Tokhanbigli, S.; Asadzadeh, H.; Nmaki, S.; Zali, M.R.; Hashemi, S.M. Exosomes as a Novel Cell-Free Therapeutic Approach in Gastrointestinal Diseases. J. Cell. Physiol. 2019, 234, 9910–9926. [Google Scholar] [CrossRef]
- Hu, G.; Gong, A.-Y.; Roth, A.L.; Huang, B.Q.; Ward, H.D.; Zhu, G.; LaRusso, N.F.; Hanson, N.D.; Chen, X.-M. Release of Luminal Exosomes Contributes to TLR4-Mediated Epithelial Antimicrobial Defense. PLoS Pathog. 2013, 9, e1003261. [Google Scholar] [CrossRef] [Green Version]
- Picard, M. Pathways to Aging: The Mitochondrion at the Intersection of Biological and Psychosocial Sciences. J. Aging Res. 2011, 2011, e814096. [Google Scholar] [CrossRef] [Green Version]
- Picard, M.; Juster, R.-P.; McEwen, B.S. Mitochondrial Allostatic Load Puts the “gluc” Back in Glucocorticoids. Nat. Rev. Endocrinol. 2014, 10, 303–310. [Google Scholar] [CrossRef]
- He, J.; Jiang, B.-H. Interplay between Reactive Oxygen Species and MicroRNAs in Cancer. Curr. Pharmacol. Rep. 2016, 2, 82–90. [Google Scholar] [CrossRef] [Green Version]
- Chettimada, S.; Lorenz, D.R.; Misra, V.; Wolinsky, S.M.; Gabuzda, D. Small RNA Sequencing of Extracellular Vesicles Identifies Circulating MiRNAs Related to Inflammation and Oxidative Stress in HIV Patients. BMC Immunol. 2020, 21, 57. [Google Scholar] [CrossRef]
- Picard, M.; McEwen, B.S. Psychological Stress and Mitochondria: A Conceptual Framework. Psychosom. Med. 2018, 80, 126–140. [Google Scholar] [CrossRef]
- Ho, P.W.-L.; Pang, W.-F.; Luk, C.C.-W.; Ng, J.K.-C.; Chow, K.-M.; Kwan, B.C.-H.; Li, P.K.-T.; Szeto, C.-C. Urinary Mitochondrial DNA Level as a Biomarker of Acute Kidney Injury Severity. Kidney Dis. 2017, 3, 78–83. [Google Scholar] [CrossRef]
- Pyle, A.; Brennan, R.; Kurzawa-Akanbi, M.; Yarnall, A.; Thouin, A.; Mollenhauer, B.; Burn, D.; Chinnery, P.F.; Hudson, G. Reduced Cerebrospinal Fluid Mitochondrial DNA Is a Biomarker for Early-Stage Parkinson’s Disease. Ann. Neurol. 2015, 78, 1000–1004. [Google Scholar] [CrossRef]
- Ellinger, J.; Müller, D.C.; Müller, S.C.; Hauser, S.; Heukamp, L.C.; von Ruecker, A.; Bastian, P.J.; Walgenbach-Brunagel, G. Circulating Mitochondrial DNA in Serum: A Universal Diagnostic Biomarker for Patients with Urological Malignancies. Urol. Oncol. Semin. Orig. Investig. 2012, 30, 509–515. [Google Scholar] [CrossRef]
- Riley, J.S.; Tait, S.W. Mitochondrial DNA in Inflammation and Immunity. EMBO Rep. 2020, 21, e49799. [Google Scholar] [CrossRef]
- Lazo, S.; Hooten, N.N.; Green, J.; Eitan, E.; Mode, N.A.; Liu, Q.-R.; Zonderman, A.B.; Ezike, N.; Mattson, M.P.; Ghosh, P.; et al. Mitochondrial DNA in Extracellular Vesicles Declines with Age. Aging Cell 2021, 20, e13283. [Google Scholar] [CrossRef]
- Lambertini, L.; Chen, J.; Nomura, Y. Mitochondrial Gene Expression Profiles Are Associated with Maternal Psychosocial Stress in Pregnancy and Infant Temperament. PLoS ONE 2015, 10, e0138929. [Google Scholar] [CrossRef]
- Mondal, T.K.; Emeny, R.T.; Gao, D.; Ault, J.G.; Kasten-Jolly, J.; Lawrence, D.A. A Physical/Psychological and Biological Stress Combine to Enhance Endoplasmic Reticulum Stress. Toxicol. Appl. Pharmacol. 2015, 289, 313–322. [Google Scholar] [CrossRef] [Green Version]
- Kouwaki, T.; Okamoto, M.; Tsukamoto, H.; Fukushima, Y.; Oshiumi, H. Extracellular Vesicles Deliver Host and Virus RNA and Regulate Innate Immune Response. Int. J. Mol. Sci. 2017, 18, 666. [Google Scholar] [CrossRef] [Green Version]
- Beninson, L.A.; Brown, P.N.; Loughridge, A.B.; Saludes, J.P.; Maslanik, T.; Hills, A.K.; Woodworth, T.; Craig, W.; Yin, H.; Fleshner, M. Acute Stressor Exposure Modifies Plasma Exosome-Associated Heat Shock Protein 72 (Hsp72) and MicroRNA (MiR-142-5p and MiR-203). PLoS ONE 2014, 9, e108748. [Google Scholar] [CrossRef] [Green Version]
- Ding, S.; Liang, Y.; Zhao, M.; Liang, G.; Long, H.; Zhao, S.; Wang, Y.; Yin, H.; Zhang, P.; Zhang, Q.; et al. Decreased MicroRNA-142-3p/5p Expression Causes CD4+ T Cell Activation and B Cell Hyperstimulation in Systemic Lupus Erythematosus. Arthritis Rheum. 2012, 64, 2953–2963. [Google Scholar] [CrossRef]
- Sonkoly, E.; Ståhle, M.; Pivarcsi, A. MicroRNAs and Immunity: Novel Players in the Regulation of Normal Immune Function and Inflammation. Semin. Cancer Biol. 2008, 18, 131–140. [Google Scholar] [CrossRef]
- Sonkoly, E.; Wei, T.; Janson, P.C.J.; Sääf, A.; Lundeberg, L.; Tengvall-Linder, M.; Norstedt, G.; Alenius, H.; Homey, B.; Scheynius, A.; et al. MicroRNAs: Novel Regulators Involved in the Pathogenesis of Psoriasis? PLoS ONE 2007, 2, e610. [Google Scholar] [CrossRef] [Green Version]
- Cohney, S.J.; Sanden, D.; Cacalano, N.A.; Yoshimura, A.; Mui, A.; Migone, T.S.; Johnston, J.A. SOCS-3 Is Tyrosine Phosphorylated in Response to Interleukin-2 and Suppresses STAT5 Phosphorylation and Lymphocyte Proliferation. Mol. Cell. Biol. 1999, 19, 4980–4988. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamamoto, K.; Yamaguchi, M.; Miyasaka, N.; Miura, O. SOCS-3 Inhibits IL-12-Induced STAT4 Activation by Binding through Its SH2 Domain to the STAT4 Docking Site in the IL-12 Receptor Β2 Subunit. Biochem. Biophys. Res. Commun. 2003, 310, 1188–1193. [Google Scholar] [CrossRef] [PubMed]
- Fang, K.; Xu, J.-X.; Chen, X.-X.; Gao, X.-R.; Huang, L.-L.; Du, A.-Q.; Jiang, C.; Ge, J.-F. Differential Serum Exosome MicroRNA Profile in a Stress-Induced Depression Rat Model. J. Affect. Disord. 2020, 274, 144–158. [Google Scholar] [CrossRef] [PubMed]
- Shyamasundar, S.; Ong, C.; Yung, L.-Y.L.; Dheen, S.T.; Bay, B.H. MiR-128 Regulates Genes Associated with Inflammation and Fibrosis of Rat Kidney Cells In Vitro. Anat. Rec. 2018, 301, 913–921. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, H.; Yang, T.; Fei, Z. MiR-26a-5p Alleviates Lipopolysaccharide-induced Acute Lung Injury by Targeting the Connective Tissue Growth Factor. Mol. Med. Rep. 2021, 23, 5. [Google Scholar] [CrossRef] [PubMed]
- Chen, P.; Miao, Y.; Yan, P.; Wang, X.J.; Jiang, C.; Lei, Y. MiR-455-5p Ameliorates HG-Induced Apoptosis, Oxidative Stress and Inflammatory via Targeting SOCS3 in Retinal Pigment Epithelial Cells. J. Cell. Physiol. 2019, 234, 21915–21924. [Google Scholar] [CrossRef] [PubMed]
- Wei, Z.-X.; Xie, G.-J.; Mao, X.; Zou, X.-P.; Liao, Y.-J.; Liu, Q.-S.; Wang, H.; Cheng, Y. Exosomes from Patients with Major Depression Cause Depressive-like Behaviors in Mice with Involvement of MiR-139-5p-Regulated Neurogenesis. Neuropsychopharmacology 2020, 45, 1050–1058. [Google Scholar] [CrossRef]
- Xie, F.; Wen, G.; Sun, W.; Jiang, K.; Chen, T.; Chen, S.; Wen, J. Mechanical Stress Promotes Angiogenesis through Fibroblast Exosomes. Biochem. Biophys. Res. Commun. 2020, 533, 346–353. [Google Scholar] [CrossRef]
- Chew, C.K.; Clarke, B.L. Causes of Low Peak Bone Mass in Women. Maturitas 2018, 111, 61–68. [Google Scholar] [CrossRef]
- Raggatt, L.J.; Partridge, N.C. Cellular and Molecular Mechanisms of Bone Remodeling. J. Biol. Chem. 2010, 285, 25103–25108. [Google Scholar] [CrossRef] [Green Version]
- Eriksen, E.F. Cellular Mechanisms of Bone Remodeling. Rev. Endocr. Metab. Disord. 2010, 11, 219–227. [Google Scholar] [CrossRef] [Green Version]
- Behera, J.; Tyagi, N. Exosomes: Mediators of Bone Diseases, Protection, and Therapeutics Potential. Oncoscience 2018, 5, 181–195. [Google Scholar] [CrossRef] [Green Version]
- Chen, H.; Senda, T.; Kubo, K. The Osteocyte Plays Multiple Roles in Bone Remodeling and Mineral Homeostasis. Med. Mol. Morphol. 2015, 48, 61–68. [Google Scholar] [CrossRef]
- Han, Y.; Kim, Y.-M.; Kim, H.S.; Lee, K.Y. Melatonin Promotes Osteoblast Differentiation by Regulating Osterix Protein Stability and Expression. Sci. Rep. 2017, 7, 5716. [Google Scholar] [CrossRef]
- Komori, T. Runx2, an Inducer of Osteoblast and Chondrocyte Differentiation. Histochem. Cell Biol. 2018, 149, 313–323. [Google Scholar] [CrossRef]
- Martin, T.J.; Sims, N.A. RANKL/OPG; Critical Role in Bone Physiology. Rev. Endocr. Metab. Disord. 2015, 16, 131–139. [Google Scholar] [CrossRef]
- Cui, Y.; Luan, J.; Li, H.; Zhou, X.; Han, J. Exosomes Derived from Mineralizing Osteoblasts Promote ST2 Cell Osteogenic Differentiation by Alteration of MicroRNA Expression. FEBS Lett. 2016, 590, 185–192. [Google Scholar] [CrossRef] [Green Version]
- Chen, C.; Cheng, P.; Xie, H.; Zhou, H.-D.; Wu, X.-P.; Liao, E.-Y.; Luo, X.-H. MiR-503 Regulates Osteoclastogenesis via Targeting RANK. J. Bone Miner. Res. 2014, 29, 338–347. [Google Scholar] [CrossRef]
- Sivan, U.; De Angelis, J.; Kusumbe, A.P. Role of Angiocrine Signals in Bone Development, Homeostasis and Disease. Open Biol. 2019, 190144. [Google Scholar] [CrossRef] [Green Version]
- Song, H.; Li, X.; Zhao, Z.; Qian, J.; Wang, Y.; Cui, J.; Weng, W.; Cao, L.; Chen, X.; Hu, Y.; et al. Reversal of Osteoporotic Activity by Endothelial Cell-Secreted Bone Targeting and Biocompatible Exosomes. Nano Lett. 2019, 19, 3040–3048. [Google Scholar] [CrossRef]
- Xu, Z.; Liu, X.; Wang, H.; Li, J.; Dai, L.; Li, J.; Dong, C. Lung Adenocarcinoma Cell-Derived Exosomal MiR-21 Facilitates Osteoclastogenesis. Gene 2018, 666, 116–122. [Google Scholar] [CrossRef]
- Guo, L.; Zhu, Y.; Li, L.; Zhou, S.; Yin, G.; Yu, G.; Cui, H. Breast Cancer Cell-Derived Exosomal MiR-20a-5p Promotes the Proliferation and Differentiation of Osteoclasts by Targeting SRCIN1. Cancer Med. 2019, 8, 5687–5701. [Google Scholar] [CrossRef] [Green Version]
- Huynh, N.; VonMoss, L.; Smith, D.; Rahman, I.; Felemban, M.F.; Zuo, J.; Rody, W.J.; McHugh, K.P.; Holliday, L.S. Characterization of Regulatory Extracellular Vesicles from Osteoclasts. J. Dent. Res. 2016, 95, 673–679. [Google Scholar] [CrossRef]
- Deng, L.; Wang, Y.; Peng, Y.; Wu, Y.; Ding, Y.; Jiang, Y.; Shen, Z.; Fu, Q. Osteoblast-Derived Microvesicles: A Novel Mechanism for Communication between Osteoblasts and Osteoclasts. Bone 2015, 79, 37–42. [Google Scholar] [CrossRef] [Green Version]
- Cappariello, A.; Loftus, A.; Muraca, M.; Maurizi, A.; Rucci, N.; Teti, A. Osteoblast-Derived Extracellular Vesicles Are Biological Tools for the Delivery of Active Molecules to Bone. J. Bone Miner. Res. 2018, 33, 517–533. [Google Scholar] [CrossRef] [Green Version]
- Lee, W.-C.; Guntur, A.R.; Long, F.; Rosen, C.J. Energy Metabolism of the Osteoblast: Implications for Osteoporosis. Endocr. Rev. 2017, 38, 255–266. [Google Scholar] [CrossRef]
- Sun, W.; Zhao, C.; Li, Y.; Wang, L.; Nie, G.; Peng, J.; Wang, A.; Zhang, P.; Tian, W.; Li, Q.; et al. Osteoclast-Derived MicroRNA-Containing Exosomes Selectively Inhibit Osteoblast Activity. Cell Discov. 2016, 2, 1–23. [Google Scholar] [CrossRef]
- Wang, X.; Guo, B.; Li, Q.; Peng, J.; Yang, Z.; Wang, A.; Li, D.; Hou, Z.; Lv, K.; Kan, G.; et al. MiR-214 Targets ATF4 to Inhibit Bone Formation. Nat. Med. 2013, 19, 93–100. [Google Scholar] [CrossRef]
- Yang, J.-X.; Xie, P.; Li, Y.-S.; Wen, T.; Yang, X.-C. Osteoclast-Derived MiR-23a-5p-Containing Exosomes Inhibit Osteogenic Differentiation by Regulating Runx2. Cell. Signal. 2020, 70, 109504. [Google Scholar] [CrossRef]
- Qin, Y.; Wang, L.; Gao, Z.; Chen, G.; Zhang, C. Bone Marrow Stromal/Stem Cell-Derived Extracellular Vesicles Regulate Osteoblast Activity and Differentiation In Vitro and Promote Bone Regeneration In Vivo. Sci. Rep. 2016, 6, 21961. [Google Scholar] [CrossRef]
- Wang, X.; Omar, O.; Vazirisani, F.; Thomsen, P.; Ekström, K. Mesenchymal Stem Cell-Derived Exosomes Have Altered MicroRNA Profiles and Induce Osteogenic Differentiation Depending on the Stage of Differentiation. PLoS ONE 2018, 13, e0193059. [Google Scholar] [CrossRef]
- Fang, S.; Li, Y.; Chen, P. Osteogenic Effect of Bone Marrow Mesenchymal Stem Cell-Derived Exosomes on Steroid-Induced Osteonecrosis of the Femoral Head. Drug Des. Devel. Ther. 2018, 13, 45–55. [Google Scholar] [CrossRef] [Green Version]
- Li, D.; Liu, J.; Guo, B.; Liang, C.; Dang, L.; Lu, C.; He, X.; Cheung, H.Y.-S.; Xu, L.; Lu, C.; et al. Osteoclast-Derived Exosomal MiR-214-3p Inhibits Osteoblastic Bone Formation. Nat. Commun. 2016, 7, 10872. [Google Scholar] [CrossRef] [PubMed]
- Ramachandran, A.; Ravindran, S.; Huang, C.-C.; George, A. TGF Beta Receptor II Interacting Protein-1, an Intracellular Protein Has an Extracellular Role as a Modulator of Matrix Mineralization. Sci. Rep. 2016, 6, 37885. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, J.-F.; Yang, G.; Pan, X.-H.; Zhang, S.-J.; Zhao, C.; Qiu, B.-S.; Gu, H.-F.; Hong, J.-F.; Cao, L.; Chen, Y.; et al. Altered MicroRNA Expression Profile in Exosomes during Osteogenic Differentiation of Human Bone Marrow-Derived Mesenchymal Stem Cells. PLoS ONE 2014, 9, e114627. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.; Wang, D.; Moshaverinia, A.; Liu, D.; Kou, X.; Yu, W.; Yang, R.; Sun, L.; Shi, S. Mesenchymal Stem Cell Transplantation in Tight-Skin Mice Identifies MiR-151-5p as a Therapeutic Target for Systemic Sclerosis. Cell Res. 2017, 27, 559–577. [Google Scholar] [CrossRef] [PubMed]
- Weilner, S.; Schraml, E.; Wieser, M.; Messner, P.; Schneider, K.; Wassermann, K.; Micutkova, L.; Fortschegger, K.; Maier, A.B.; Westendorp, R.; et al. Secreted Microvesicular MiR-31 Inhibits Osteogenic Differentiation of Mesenchymal Stem Cells. Aging Cell 2016, 15, 744–754. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tu, M.; Tang, J.; He, H.; Cheng, P.; Chen, C. MiR-142-5p Promotes Bone Repair by Maintaining Osteoblast Activity. J. Bone Miner. Metab. 2017, 35, 255–264. [Google Scholar] [CrossRef]
- Li, H.; Yue, L.; Xu, H.; Li, N.; Li, J.; Zhang, Z.; Zhao, R.C. Curcumin Suppresses Osteogenesis by Inducing MiR-126a-3p and Subsequently Suppressing the WNT/LRP6 Pathway. Aging 2019, 11, 6983–6998. [Google Scholar] [CrossRef]
- Amjadi-Moheb, F.; Akhavan-Niaki, H. Wnt Signaling Pathway in Osteoporosis: Epigenetic Regulation, Interaction with Other Signaling Pathways, and Therapeutic Promises. J. Cell. Physiol. 2019, 234, 14641–14650. [Google Scholar] [CrossRef]
- Xu, T.; Luo, Y.; Wang, J.; Zhang, N.; Gu, C.; Li, L.; Qian, D.; Cai, W.; Fan, J.; Yin, G. Exosomal MiRNA-128-3p from Mesenchymal Stem Cells of Aged Rats Regulates Osteogenesis and Bone Fracture Healing by Targeting Smad5. J. Nanobiotechnology 2020, 18, 47. [Google Scholar] [CrossRef] [Green Version]
- Zhang, S.; Wu, W.; Jiao, G.; Li, C.; Liu, H. MiR-455-3p Activates Nrf2/ARE Signaling via HDAC2 and Protects Osteoblasts from Oxidative Stress. Int. J. Biol. Macromol. 2018, 107, 2094–2101. [Google Scholar] [CrossRef]
- Sun, Y.; Wang, X.; Chen, G.; Song, C.; Ma, X.; Fu, Y.; Feng, C.; Yan, J. MiRNA-187-5p Regulates Osteoblastic Differentiation of Bone Marrow Mesenchymal Stem Cells in Mice by Targeting ICAM1. BioMed Res. Int. 2020, 2020, 1–12. [Google Scholar] [CrossRef]
- Dai, Y.; Zheng, C.; Li, H. Inhibition of MiR-23a-3p Promotes Osteoblast Proliferation and Differentiation. J. Cell. Biochem. 2019. [Google Scholar] [CrossRef]
- Sato, M.; Suzuki, T.; Kawano, M.; Tamura, M. Circulating Osteocyte-derived Exosomes Contain MiRNAs Which Are Enriched in Exosomes from MLO-Y4 Cells. Biomed. Rep. 2017, 6, 223–231. [Google Scholar] [CrossRef]
- Xiong, Y.; Tang, Y.; Fan, F.; Zeng, Y.; Li, C.; Zhou, G.; Hu, Z.; Zhang, L.; Liu, Z. Exosomal Hsa-MiR-21-5p Derived from Growth Hormone-Secreting Pituitary Adenoma Promotes Abnormal Bone Formation in Acromegaly. Transl. Res. 2020, 215, 1–16. [Google Scholar] [CrossRef] [Green Version]
- Li, S.; Hu, C.; Li, J.; Liu, L.; Jing, W.; Tang, W.; Tian, W.; Long, J. Effect of MiR-26a-5p on the Wnt/Ca2+ Pathway and Osteogenic Differentiation of Mouse Adipose-Derived Mesenchymal Stem Cells. Calcif. Tissue Int. 2016, 99, 174–186. [Google Scholar] [CrossRef]
- Long, H.; Sun, B.; Cheng, L.; Zhao, S.; Zhu, Y.; Zhao, R.; Zhu, J. MiR-139-5p Represses BMSC Osteogenesis via Targeting Wnt/β-Catenin Signaling Pathway. DNA Cell Biol. 2017, 36, 715–724. [Google Scholar] [CrossRef]
- Gu, H.; Shi, S.; Xiao, F.; Huang, Z.; Xu, J.; Chen, G.; Zhou, K.; Lu, L.; Yin, X. MiR-1-3p Regulates the Differentiation of Mesenchymal Stem Cells to Prevent Osteoporosis by Targeting Secreted Frizzled-Related Protein 1. Bone 2020, 137, 115444. [Google Scholar] [CrossRef]
- Afrisham, R.; Paknejad, M.; Soliemanifar, O.; Sadegh-Nejadi, S.; Meshkani, R.; Ashtary-Larky, D. The Influence of Psychological Stress on the Initiation and Progression of Diabetes and Cancer. Int. J. Endocrinol. Metab. 2019, 17. [Google Scholar] [CrossRef] [Green Version]
- Pegtel, D.M.; van de Garde, M.D.B.; Middeldorp, J.M. Viral MiRNAs Exploiting the Endosomal–Exosomal Pathway for Intercellular Cross-Talk and Immune Evasion. Biochim. Biophys. Acta BBA Gene Regul. Mech. 2011, 1809, 715–721. [Google Scholar] [CrossRef]
- Beninson, L.A.; Fleshner, M. Exosomes: An Emerging Factor in Stress-Induced Immunomodulation. Semin. Immunol. 2014, 26, 394–401. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Tissue | Functions | Reference |
|---|---|---|
| Tumor | Biomarker Alters tumor microenvironment Regulates tumor immune response Involved in tumor angiogenesis | [31,32,38,39] |
| Bone | Biomarker Regulates osteogenic differentiation of mesenchymal stem cells Regulates osteoblast proliferation and activity Affects osteoblast differentiation Regulates osteoclast function and induces osteoclast differentiation | [17,40,41,42] |
| Heart | Biomarker Promotes angiogenesis Cardioprotection and regeneration | [43,44] |
| Brain | Biomarker Influences inflammatory and regulatory pathways in the brain Neuroprotective effect | [45,46,47] |
| Kidney | Biomarker Involved in the development of renal fibrosis Contributing to kidney repair | [48] |
| Gastro-intestinal tract | Immunomodulation Response of anti-apoptotic, antioxidant stress Regulates the homeostasis of gut microbiota | [49,50] |
| Stressors | MiRNAs in EVs | Source of EVs | Stress-Induced Change | References |
|---|---|---|---|---|
| Chronic unpredictable mild stress | miR-139-5p | Blood and brain from mice | [74] | |
| Chronic unpredictable mild stress | miR-126a-3p, miR-128-3p, miR-26a-5p, miR-191a-5p | Serum from rats | [70] | |
| Mechanical stress | miR-1246 | Fibroblast | [75] | |
| Mechanical stress | miR-133a-3p, miR-203-3p | Fibroblast | [75] | |
| Chronic unpredictable mild stress | miR-455-3p, miR-187-5p, miR-206-3p, miR-455-5p | Serum from rats | [70] | |
| Inescapable tail shock | miR-142-5p, miR-203 | Plasma from rats | [64] |
| Source | Bioactive Factors Containing | Target | Function | References |
|---|---|---|---|---|
| Osteoclasts | RANK | Osteoclasts | Inhibits osteoclast formation | [90] |
| Osteoclasts | miR-214 | Osteoblasts | Inhibits the activity of osteoblasts through ephrina2/ephrin type-A receptor 2 interaction and targets activating transcription factor 4 to inhibit bone formation | [94,95] |
| Osteoclasts | miR-23a-5p | Osteoblasts | Inhibits the activity of osteoblasts by targeting Runx2 | [96] |
| Osteoclasts | miR-214-3p | Osteoblasts | Inhibits osteoblastic bone formation | [100] |
| Osteoblasts | RANKL | Osteoclast precursors | Facilitates osteoclast formation by binding RANK on the osteoclast precursor surface | [91] |
| Osteoblasts | RANKL | Osteoclasts | Induces the apoptosis of osteoclasts | [92] |
| Preosteoblasts | TRIP-1 | The extracellular matrix of bone | Promotes mineralization | [101] |
| BMSCs | miR-196a | Osteoblasts | Improves osteogenic function | [97] |
| BMSCs | miR-885-5p | BMSCs | Inhibits osteogenic differentiation by repressing Runx2 | [102] |
| BMSCs | miR-151-5p | BMSCs | Promotes osteogenic differentiation | [103] |
| Endothelial cells | miR-155 | Osteoclasts | Inhibits the activity and differentiation of osteoclasts | [87] |
| Endothelial cells | miR-31 | MSCs | Inhibits osteogenic differentiation by repressing Frizzled-3 | [104] |
| MiRNAs in EVs | Stress-Induced Change | The Effect of MiRNAs in EVs on Bone | References |
|---|---|---|---|
| miR-126a-3p | Inhibits the osteogenesis of human adipose-derived mesenchymal stem cells | [70,106] | |
| miR-128-3p | Inhibits the osteogenic differentiation of MSCs | [70,108] | |
| miR-26a-5p | Inhibits the osteogenic differentiation of mouse adipose-derived mesenchymal stem cells | [70,114] | |
| miR-139-5p | Inhibits BMSC osteogenesis by targeting Wnt/β-catenin signaling pathway | [74,115] | |
| miR-455-3p | Protection of osteoblasts from oxidative stress | [70,109] | |
| miR-187-5p | Promotes differentiation of BMSCs to osteoblasts | [70,110] | |
| miR-1-3p | Stimulates the osteogenesis of mouse MSCs and inhibits their adipogenesis | [70,116] | |
| miR-23a-3p | Inhibits the osteogenesis | [70,111] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
He, Y.; Wuertz-Kozak, K.; Kuehl, L.K.; Wippert, P.-M. Extracellular Vesicles: Potential Mediators of Psychosocial Stress Contribution to Osteoporosis? Int. J. Mol. Sci. 2021, 22, 5846. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22115846
He Y, Wuertz-Kozak K, Kuehl LK, Wippert P-M. Extracellular Vesicles: Potential Mediators of Psychosocial Stress Contribution to Osteoporosis? International Journal of Molecular Sciences. 2021; 22(11):5846. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22115846
Chicago/Turabian StyleHe, Yangyang, Karin Wuertz-Kozak, Linn K. Kuehl, and Pia-Maria Wippert. 2021. "Extracellular Vesicles: Potential Mediators of Psychosocial Stress Contribution to Osteoporosis?" International Journal of Molecular Sciences 22, no. 11: 5846. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22115846