
Identification of Three Type II Toxin-Antitoxin Systems in Model Bacterial Plant Pathogen Dickeya dadantii 3937

 , , ,
, , ,
Abstract
:1. Introduction
2. Results
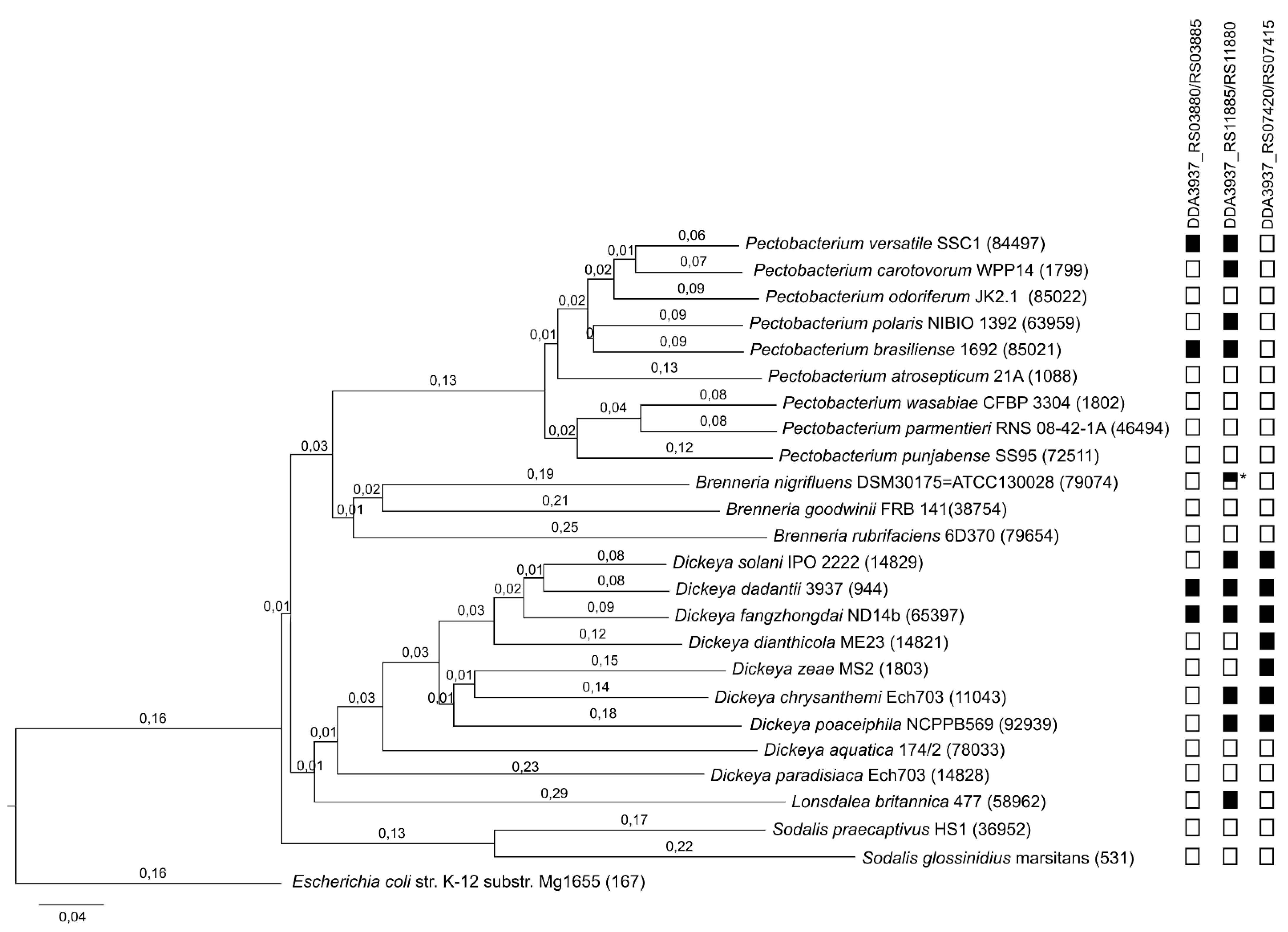
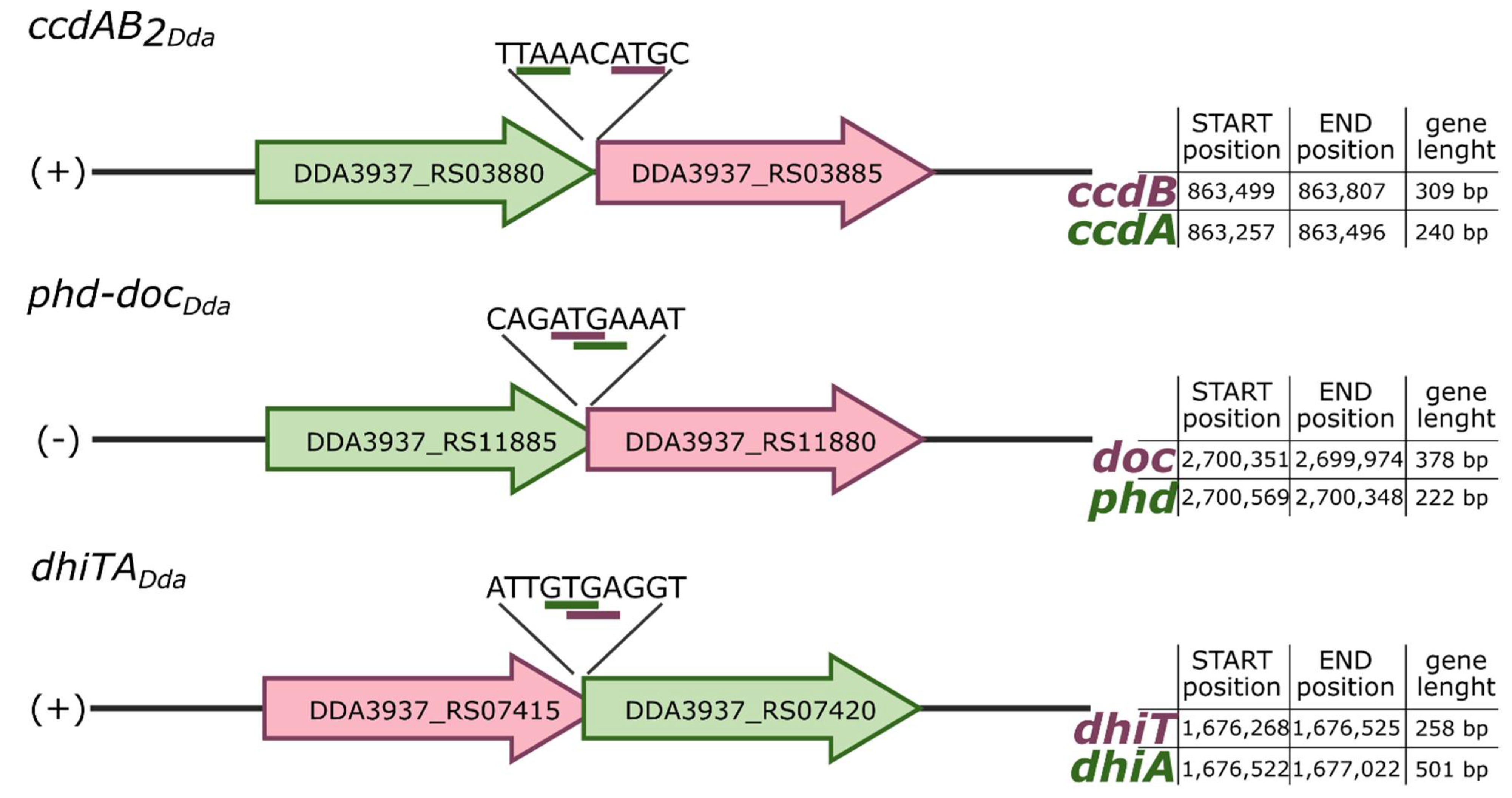
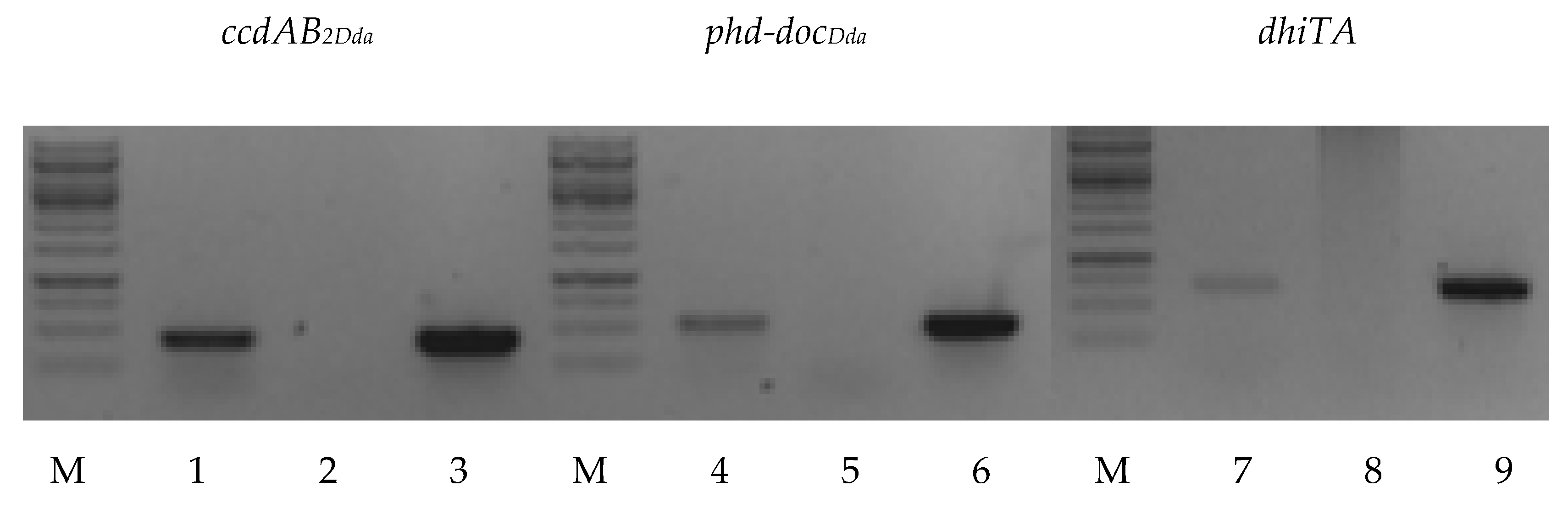
2.1. Putative TA Systems Identification
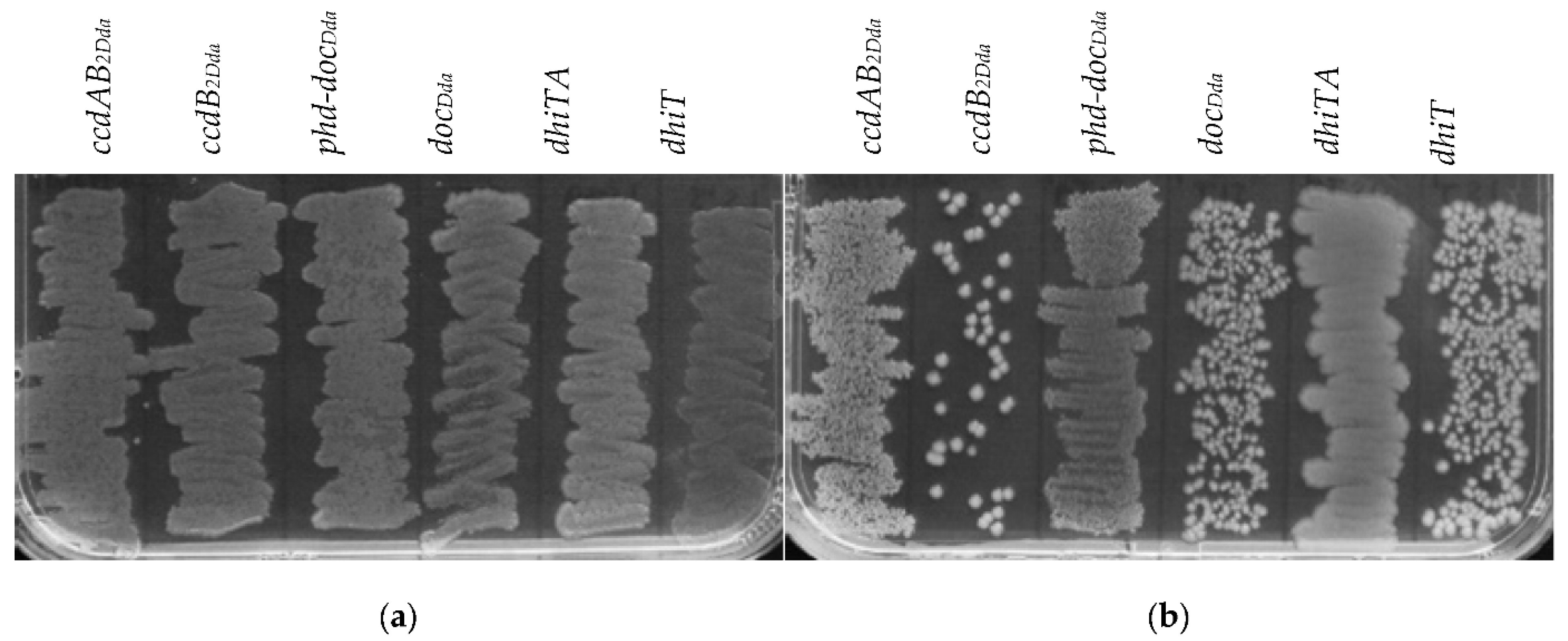
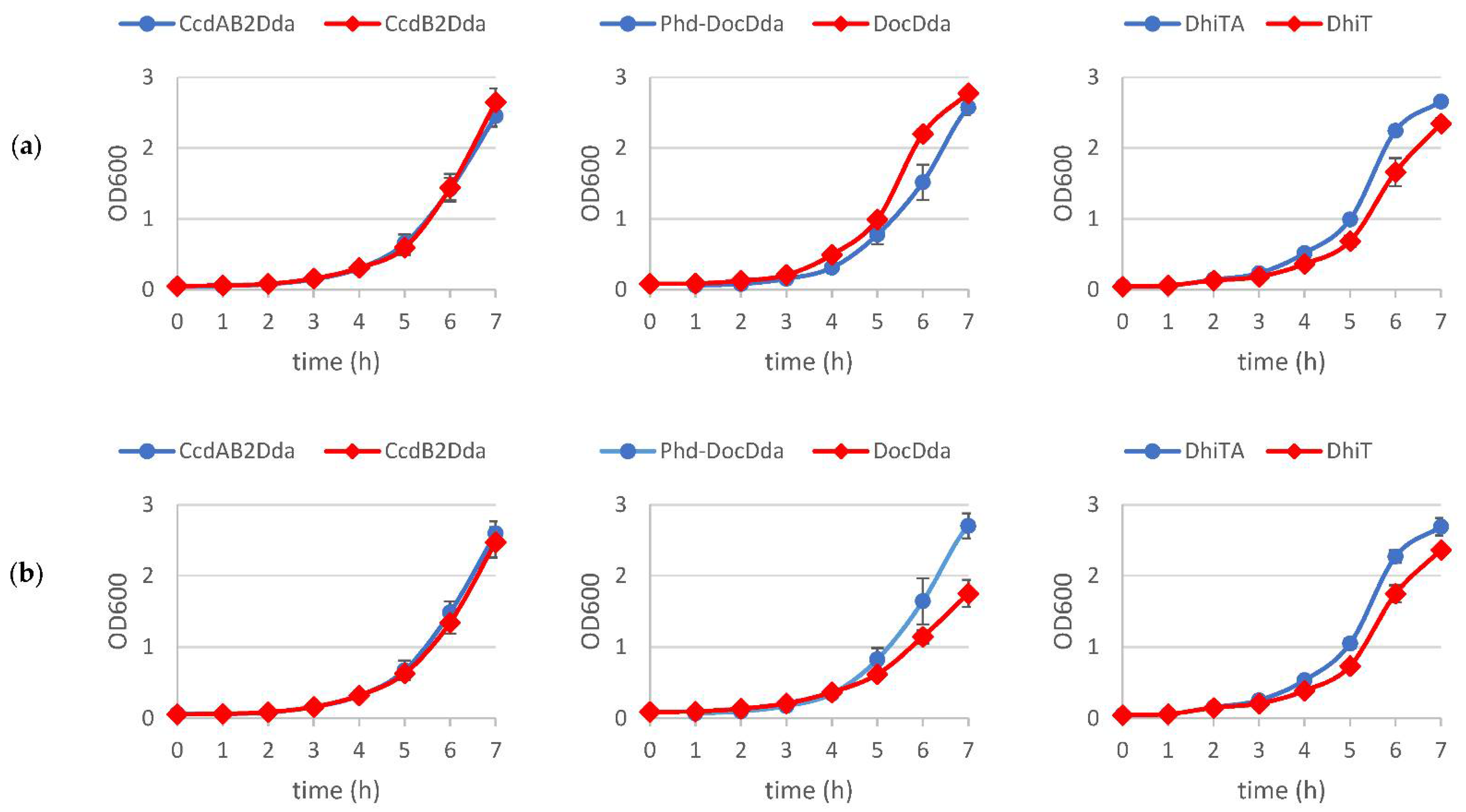
2.2. The Effect of Putative Toxins and Toxin-Antitoxin Complex Overproduction on Bacterial Growth
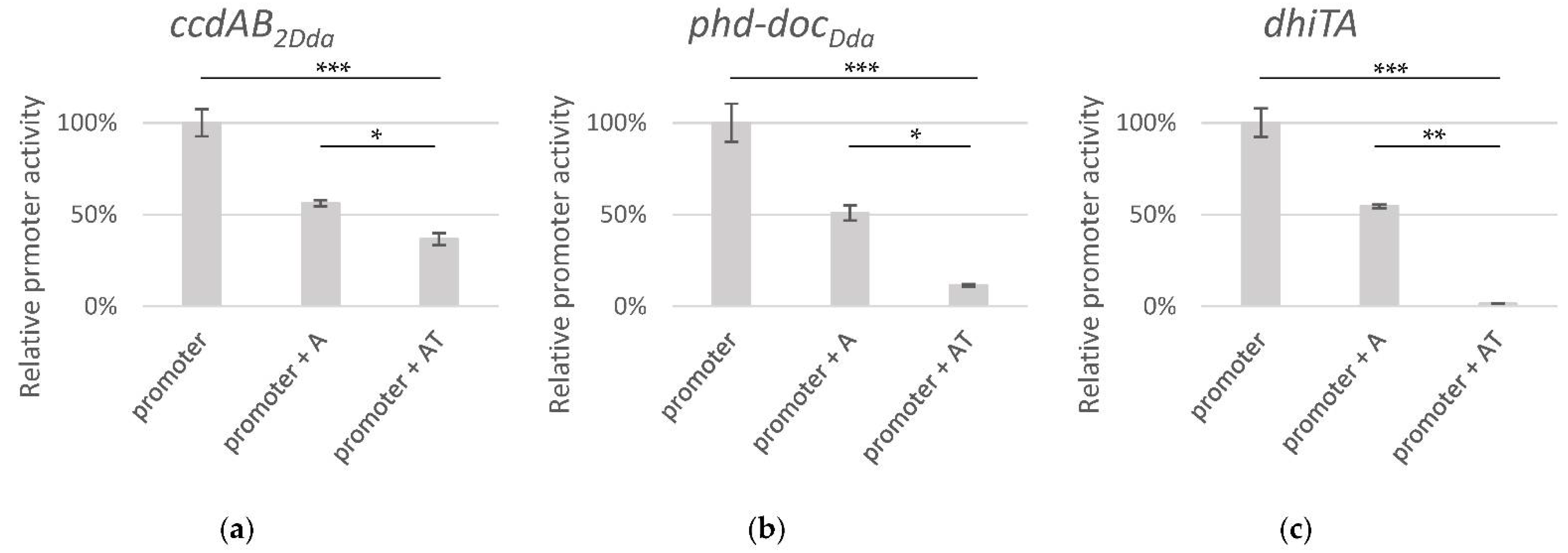
2.3. Transcriptional Regulation of the Putative TA Operons
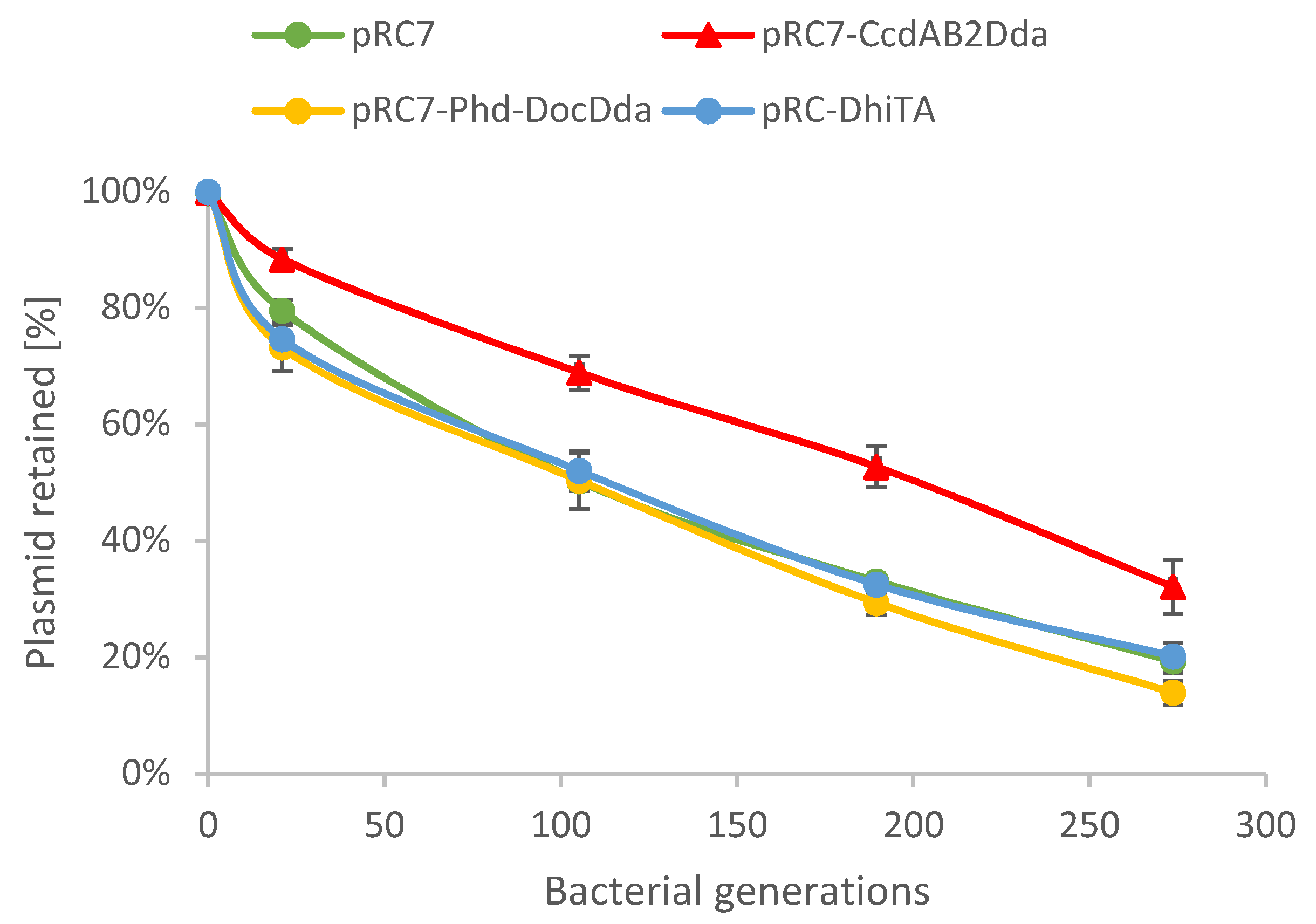
2.4. Plasmid Stability Is Affected by ccdAB2Dda System
3. Discussion
4. Materials and Methods
4.1. Bacterial Strains, Plasmids, Primers, and Growth Conditions
4.2. Bioinformatics and Statistical Analysis
4.3. Phylogenetic Analysis
4.4. TA Systems Toxicity Assay (Solid Medium)
4.5. TA Systems Toxicity Assay (Liquid Medium)
4.6. Promoter Activity Assay
4.7. Plasmid Stability Assay
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Reverchon, S.; Muskhelisvili, G.; Nasser, W. Virulence Program of a Bacterial Plant Pathogen: The Dickeya Model. In Progress in Molecular Biology and Translational Science; Elsevier: Amsterdam, The Netherlands, 2016; Volume 142, pp. 51–92. ISBN 9780128093856. [Google Scholar]
- Reverchon, S.; Nasser, W. Dickeya ecology, environment sensing and regulation of virulence programme. Environ. Microbiol. Rep. 2013, 5, 622–636. [Google Scholar] [CrossRef] [PubMed]
- Mansfield, J.; Genin, S.; Magori, S.; Citovsky, V.; Sriariyanum, M.; Ronald, P.; Dow, M.; Verdier, V.; Beer, S.V.; Machado, M.A.; et al. Top 10 plant pathogenic bacteria in molecular plant pathology. Mol. Plant Pathol. 2012. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sławiak, M.; van Beckhoven, J.R.C.M.; Speksnijder, A.G.C.L.; Czajkowski, R.; Grabe, G.; van der Wolf, J.M. Biochemical and genetical analysis reveal a new clade of biovar 3 Dickeya spp. strains isolated from potato in Europe. Eur. J. Plant Pathol. 2009, 125, 245–261. [Google Scholar] [CrossRef]
- Parkinson, N.; DeVos, P.; Pirhonen, M.; Elphinstone, J. Dickeya aquatica sp. nov., isolated from waterways. Int. J. Syst. Evol. Microbiol. 2014, 64, 2264–2266. [Google Scholar] [CrossRef] [PubMed]
- Hugouvieux-Cotte-Pattat, N.; Jacot-Des-Combes, C.; Briolay, J. Dickeya lacustris sp. nov., a water-living pectinolytic bacterium isolated from lakes in France. Int. J. Syst. Evol. Microbiol. 2019, 69, 721–726. [Google Scholar] [CrossRef]
- Chakraborty, S. Migrate or evolve: Options for plant pathogens under climate change. Glob. Chang. Biol. 2013, 19, 1985–2000. [Google Scholar] [CrossRef]
- Czajkowski, R.; Fikowicz-Krosko, J.; Maciag, T.; Rabalski, L.; Czaplewska, P.; Jafra, S.; Richert, M.; Krychowiak-Maśnicka, M.; Hugouvieux-Cotte-Pattat, N. Genome-Wide Identification of Dickeya solani Transcriptional Units Up-Regulated in Response to Plant Tissues From a Crop-Host Solanum tuberosum and a Weed-Host Solanum dulcamara. Front. Plant Sci. 2020, 11, 1. [Google Scholar] [CrossRef]
- Glasner, J.D.; Yang, C.H.; Reverchon, S.; Hugouvieux-Cotte-Pattat, N.; Condemine, G.; Bohin, J.P.; van Gijsegem, F.; Yang, S.; Franza, T.; Expert, D.; et al. Genome sequence of the plant-pathogenic bacterium Dickeya dadantii 3937. J. Bacteriol. 2011, 193, 2076–2077. [Google Scholar] [CrossRef]
- Shidore, T.; Triplett, L.R. Toxin-Antitoxin Systems: Implications for Plant Disease. Annu. Rev. Phytopathol. 2017, 55, 161–179. [Google Scholar] [CrossRef]
- Zhang, S.-P.; Wang, Q.; Quan, S.-W.; Yu, X.-Q.; Wang, Y.; Guo, D.-D.; Peng, L.; Feng, H.-Y.; He, Y.-X. Type II toxin–antitoxin system in bacteria: Activation, function, and mode of action. Biophys. Rep. 2020, 6, 68–79. [Google Scholar] [CrossRef]
- Fraikin, N.; Goormaghtigh, F.; Melderen, L. Van Type II Toxin-Antitoxin Systems: Evolution and Revolutions. J. Bacteriol. 2020. [Google Scholar] [CrossRef] [Green Version]
- Jimmy, S.; Kumar Saha, C.; Kurata, T.; Stavropoulos, C.; Raquel Alves Oliveira, S.; Koh, A.; Cepauskas, A.; Takada, H.; Rejman, D.; Tenson, T.; et al. A widespread toxin−antitoxin system exploiting growth control via alarmone signaling. Proc. Natl. Acad. Sci. USA 2020, 117. [Google Scholar] [CrossRef] [PubMed]
- Fernandez-Garcia, L.; Kim, J.-S.; Tomas, M.; Wood, T.K. Toxins of toxin/antitoxin systems are inactivated primarily through promoter mutations. J. Appl. Microbiol. 2019, 127, 1859–1868. [Google Scholar] [CrossRef]
- Song, S.; Wood, T.K. A Primary Physiological Role of Toxin/Antitoxin Systems Is Phage Inhibition. Front. Microbiol. 2020, 11, 1895. [Google Scholar] [CrossRef] [PubMed]
- Song, S.; Wood, T.K. Toxin/Antitoxin System Paradigms: Toxins Bound to Antitoxins Are Not Likely Activated by Preferential Antitoxin Degradation. Adv. Biosyst. 2020, 4, 1900290. [Google Scholar] [CrossRef]
- LeRoux, M.; Culviner, P.H.; Liu, Y.J.; Littlehale, M.L.; Laub, M.T. Stress Can Induce Transcription of Toxin-Antitoxin Systems without Activating Toxin. Mol. Cell 2020, 79, 280–292.e8. [Google Scholar] [CrossRef]
- Chistoserdova, L.; Kwak, Y.-S.; Meinhart, A.; De Souza, A.A.; Martins, P.M.M.; Machado, M.A.; Silva, N.V.; Takita, M.A. Type II Toxin-Antitoxin Distribution and Adaptive Aspects on Xanthomonas Genomes: Focus on Xanthomonas citri. Front. Microbiol. 2016, 1, 652. [Google Scholar] [CrossRef]
- Li, J.; Wang, N. Genome-Wide Mutagenesis of Xanthomonas axonopodis pv. citri Reveals Novel Genetic Determinants and Regulation Mechanisms of Biofilm Formation. PLoS ONE 2011, 6, e21804. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, X.; Wood, T.K. Toxin-Antitoxin Systems Influence Biofilm and Persister Cell Formation and the General Stress Response. Appl. Environ. Microbiol. 2011, 77, 5577–5583. [Google Scholar] [CrossRef] [Green Version]
- Riffaud, C.; Pinel-Marie, M.L.; Felden, B. Cross-Regulations between Bacterial Toxin–Antitoxin Systems: Evidence of an Interconnected Regulatory Network? Trends Microbiol. 2020, 28, 851–866. [Google Scholar] [CrossRef]
- Burbank, L.P.; Stenger, D.C. The DinJ/RelE toxin-antitoxin system suppresses bacterial proliferation and virulence of xylella fastidiosa in grapevine. Phytopathology 2017, 107, 388–394. [Google Scholar] [CrossRef] [Green Version]
- Granato, L.M.; Picchi, S.C.; De Oliveira Andrade, M.; Martins, P.M.M.; Takita, M.A.; Machado, M.A.; De Souza, A.A. The ECNA antitoxin is important not only for human pathogens: Evidence of Its Role in the Plant Pathogen Xanthomonas citri subsp. Citri. J. Bacteriol. 2019, 201. [Google Scholar] [CrossRef] [Green Version]
- De Bast, M.S.; Mine, N.; Van Melderen, L. Chromosomal toxin-antitoxin systems may act as antiaddiction modules. J. Bacteriol. 2008, 190, 4603–4609. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kato, F.; Yoshizumi, S.; Yamaguchi, Y.; Inouye, M. Genome-wide screening for identification of novel toxin-antitoxin systems in Staphylococcus aureus. Appl. Environ. Microbiol. 2019, 85. [Google Scholar] [CrossRef]
- Xie, Y.; Wei, Y.; Shen, Y.; Li, X.; Zhou, H.; Tai, C.; Deng, Z.; Ou, H.Y. TADB 2.0: An updated database of bacterial type II toxin-antitoxin loci. Nucleic Acids Res. 2018, 46, D749–D753. [Google Scholar] [CrossRef]
- Sevin, E.W.; Barloy-Hubler, F. RASTA-Bacteria: A web-based tool for identifying toxin-antitoxin loci in prokaryotes. Genome Biol. 2007, 8, R155. [Google Scholar] [CrossRef] [Green Version]
- Arizala, D.; Arif, M. Genome-wide analyses revealed remarkable heterogeneity in pathogenicity determinants, antimicrobial compounds, and CRISPR-cas systems of complex phytopathogenic genus pectobacterium. Pathogens 2019, 8, 247. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brady, C.; Arnold, D.; McDonald, J.; Denman, S. Taxonomy and identification of bacteria associated with acute oak decline. World J. Microbiol. Biotechnol. 2017, 33, 143. [Google Scholar] [CrossRef]
- Marrero, G.; Schneider, K.L.; Jenkins, D.M.; Alvarez, A.M. Phylogeny and classification of Dickeya based on multilocus sequence analysis. Int. J. Syst. Evol. Microbiol. 2013, 63, 3524–3539. [Google Scholar] [CrossRef]
- Deter, H.S.; Jensen, R.V.; Mather, W.H.; Butzin, N.C. Mechanisms for differential protein production in toxin–antitoxin systems. Toxins 2017, 9, 211. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, J.; Zhang, N.; Cao, M.; Ren, S.; Zeng, T.; Qin, M.; Zhao, X.; Yuan, F.; Chen, H.; Bei, W. Identification of three type II toxin-antitoxin systems in Streptococcus suis serotype 2. Toxins 2018, 10, 467. [Google Scholar] [CrossRef] [Green Version]
- Abràmoff, M.D.; Magalhães, P.J.; Ram, S.J. Image processing with imageJ. Biophotonics Int. 2004, 11, 36–41. [Google Scholar] [CrossRef]
- Bartosik, A.A.; Glabski, K.; Kulinska, A.; Lewicka, E.; Godziszewska, J.; Markowska, A.; Jagura-Burdzy, G. Convenient broad-host-range unstable vectors for studying stabilization cassettes in diverse bacteria. BMC Microbiol. 2016, 16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hernández-Arriaga, A.M.; Chan, W.T.; Espinosa, M.; Díaz-Orejas, R. Conditional Activation of Toxin-Antitoxin Systems: Postsegregational Killing and Beyond. Microbiol. Spectr. 2014, 2. [Google Scholar] [CrossRef] [Green Version]
- Ogura, T.; Hiraga, S. Partition mechanism of F plasmid: Two plasmid gene-encoded products and a cis-acting region are involved in partition. Cell 1983, 32, 351–360. [Google Scholar] [CrossRef]
- Lehnherr, H.; Maguin, E.; Jafri, S.; Yarmolinsky, M.B. Plasmid addiction genes of bacteriophage P1: Doc, which causes cell death on curing of prophage, and phd, which prevents host death when prophage is retained. J. Mol. Biol. 1993, 233, 414–428. [Google Scholar] [CrossRef] [Green Version]
- Bernhardt, T.G.; De Boer, P.A.J. Screening for synthetic lethal mutants in Escherichia coli and identification of EnvC (YibP) as a periplasmic septal ring factor with murein hydrolase activity. Mol. Microbiol. 2004, 52, 1255–1269. [Google Scholar] [CrossRef]
- Taheri-Araghi, S.; Bradde, S.; Sauls, J.T.; Hill, N.S.; Levin, P.A.; Paulsson, J.; Vergassola, M.; Jun, S. Cell-size control and homeostasis in bacteria. Curr. Biol. 2015, 25, 385–391. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abu Bakar, F.; Chieng Yeo, C.; Ann Harikrishna, J. Neutralization of Bacterial YoeB Spn Toxicity and Enhanced Plant Growth in Arabidopsis thaliana via Co-Expression of the Toxin-Antitoxin Genes. Int. J. Mol. Sci. 2016, 17, 321. [Google Scholar] [CrossRef] [Green Version]
- Bakar, F.A.; Yeo, C.C.; Harikrishna, J.A. Expression of the Streptococcus pneumoniae yoeB Chromosomal toxin gene causes Cell Death in the model plant Arabidopsis thaliana. BMC Biotechnol. 2015, 15, 26. [Google Scholar] [CrossRef] [Green Version]
- Starkey, M.; Rahme, L.G. Modeling Pseudomonas aeruginosa pathogenesis in plant hosts. Nat. Protoc. 2009, 4, 117–124. [Google Scholar] [CrossRef]
- Rodríguez-Andrade, E.; Hernández-Ramírez, K.C.; Díaz-Peréz, S.P.; Díaz-Magaña, A.; Chávez-Moctezuma, M.P.; Meza-Carmen, V.; Ortíz-Alvarado, R.; Cervantes, C.; Ramírez-Díaz, M.I. Genes from pUM505 plasmid contribute to Pseudomonas aeruginosa virulence. Antonie Van Leeuwenhoek 2016, 109, 389–396. [Google Scholar] [CrossRef]
- Hernández-Ramírez, K.C.; Valerio-Arellano, B.; Valle-Maldonado, M.I.; Ruíz-Herrera, L.F.; Meza-Carmen, V.; Ramírez-Díaz, M.I. Virulence Conferred by PumA Toxin from the Plasmid-Encoded PumAB Toxin-Antitoxin System is Regulated by Quorum System. Curr. Microbiol. 2020, 77, 2535–2543. [Google Scholar] [CrossRef]
- Ruhe, Z.C.; Low, D.A.; Hayes, C.S. Bacterial contact-dependent growth inhibition. Trends Microbiol. 2013, 21, 230–237. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Triplett, L.R.; Shidore, T.; Long, J.; Miao, J.; Wu, S.; Han, Q.; Zhou, C.; Ishihara, H.; Li, J.; Zhao, B.; et al. AvrRxo1 Is a Bifunctional Type III Secreted Effector and Toxin-Antitoxin System Component with Homologs in Diverse Environmental Contexts. PLoS ONE 2016, 11, e0158856. [Google Scholar] [CrossRef] [PubMed]
- Mruk, I.; Kobayashi, I. Survey and summary To be or not to be: Regulation of restriction-modification systems and other toxin-antitoxin systems. Nucleic Acids Res. 2014. [Google Scholar] [CrossRef]
- Nagornykh, M.O.; Bogdanova, E.S.; Protsenko, A.S.; Solonin, A.S.; Zakharova, M.V.; Severinov, K.V. Regulation of gene expression in a type II restriction-modification system. Russ. J. Genet. 2008, 44, 523–532. [Google Scholar] [CrossRef] [PubMed]
- Bernard, P.; Couturier, M. Cell killing by the F plasmid CcdB protein involves poisoning of DNA-topoisomerase II complexes. J. Mol. Biol. 1992, 226, 735–745. [Google Scholar] [CrossRef]
- Bahassi, E.M.; O’Dea, M.H.; Allali, N.; Messens, J.; Gellert, M.; Couturier, M. Interactions of CcdB with DNA: Gyrase in activation of GyrA, poisoning of the gyrase-DNA complex, and the antidote action of CdcA. J. Biol. Chem. 1999, 274, 10936–10944. [Google Scholar] [CrossRef] [Green Version]
- Bernard, P.; Kézdy, K.E.; Van Melderen, L.; Steyaert, J.; Wyns, L.; Pato, M.L.; Higgins, P.N.; Couturier, M. The F plasmid CcdB protein induces efficient ATP-dependent DNA cleavage by gyrase. J. Mol. Biol. 1993, 234, 534–541. [Google Scholar] [CrossRef]
- Couturier, M.; Bahassi, E.M.; Van Melderen, L. Bacterial death by DNA gyrase poisoning. Trends Microbiol. 1998, 6, 269–275. [Google Scholar] [CrossRef]
- Goeders, N.; Van Melderen, L. Toxin-antitoxin systems as multilevel interaction systems. Toxins 2013, 6, 304–324. [Google Scholar] [CrossRef] [Green Version]
- Gibson, D.G.; Young, L.; Chuang, R.Y.; Venter, J.C.; Hutchison, C.A.; Smith, H.O. Enzymatic assembly of DNA molecules up to several hundred kilobases. Nat. Methods 2009, 6, 343–345. [Google Scholar] [CrossRef]
- Akiyama, J.; Ruiz, M.-J. Automat(h). In A Day’s Adventure in Math Wonderland; World Scientific: Singapore, 2008; pp. 101–117. [Google Scholar] [CrossRef]
- Lenz, D.H.; Mok, K.C.; Lilley, B.N.; Kulkarni, R.V.; Wingreen, N.S.; Bassler, B.L. The small RNA chaperone Hfq and multiple small RNAs control quorum sensing in Vibrio harveyi and Vibrio cholerae. Cell 2004, 118, 69–82. [Google Scholar] [CrossRef] [Green Version]
- Almagro Armenteros, J.J.; Tsirigos, K.D.; Sønderby, C.K.; Petersen, T.N.; Winther, O.; Brunak, S.; von Heijne, G.; Nielsen, H. SignalP 5.0 improves signal peptide predictions using deep neural networks. Nat. Biotechnol. 2019, 37, 420–423. [Google Scholar] [CrossRef]
- Katoh, K.; Rozewicki, J.; Yamada, K.D. MAFFT online service: Multiple sequence alignment, interactive sequence choice and visualization. Brief. Bioinform. 2018, 20, 1160–1166. [Google Scholar] [CrossRef] [Green Version]
- Cummings, M.P. PAUP* [Phylogenetic Analysis Using Parsimony (and Other Methods)]. In Dictionary of Bioinformatics and Computational Biology; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2004. [Google Scholar]
- Jones, D.T.; Taylor, W.R.; Thornton, J.M. The rapid generation of mutation data matrices from protein sequences. Bioinformatics 1992, 8, 275–282. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular evolutionary genetics analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Darling, A.C.E.; Mau, B.; Blattner, F.R.; Perna, N.T. Mauve: Multiple alignment of conserved genomic sequence with rearrangements. Genome Res. 2004, 14, 1394–1403. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maciag-Dorszyńska, M.; Ignatowska, M.; Jannière, L.; Wegrzyn, G.; Szalewska-Pałasz, A. Mutations in central carbon metabolism genes suppress defects in nucleoid position and cell division of replication mutants in Escherichia coli. Gene 2012, 503, 31–35. [Google Scholar] [CrossRef] [PubMed]
- Cui, Z.; Yang, C.H.; Kharadi, R.R.; Yuan, X.; Sundin, G.W.; Triplett, L.R.; Wang, J.; Zeng, Q. Cell-length heterogeneity: A population-level solution to growth/virulence trade-offs in the plant pathogen Dickeya dadantii. bioRxiv 2019, 1–28. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boss, L.; Labudda, Ł.; Węgrzyn, G.; Hayes, F.; Kędzierska, B. The Axe-Txe Complex of Enterococcus faecium Presents a Multilayered Mode of Toxin-Antitoxin Gene Expression Regulation. PLoS ONE 2013, 8, e73569. [Google Scholar] [CrossRef] [PubMed] [Green Version]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| TA Name | Toxin (T) NCBI ID [9] | Antitoxin (A) NCBI ID [9] | Strand | Distance (bp) 1 | Domain Pair 2 | T/A Family |
|---|---|---|---|---|---|---|
| ccdAB2Dda | DDA3937_RS03885 | DDA3937_RS03880 | + | +3 | MazF-like domain/RHH-like domain | ccdB/ccdA |
| phd-docDda | DDA3937_RS11880 | DDA3937_RS11885 | − | −3 | Fic-like domain/PHD-like domain | Doc/Phd/YefM |
| dhiTA | DDA3937_RS07415 | DDA3937_RS07420 | + | −3 | DUF4160/DUF2442 | NH |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Boss, L.; Górniak, M.; Lewańczyk, A.; Morcinek-Orłowska, J.; Barańska, S.; Szalewska-Pałasz, A. Identification of Three Type II Toxin-Antitoxin Systems in Model Bacterial Plant Pathogen Dickeya dadantii 3937. Int. J. Mol. Sci. 2021, 22, 5932. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22115932
Boss L, Górniak M, Lewańczyk A, Morcinek-Orłowska J, Barańska S, Szalewska-Pałasz A. Identification of Three Type II Toxin-Antitoxin Systems in Model Bacterial Plant Pathogen Dickeya dadantii 3937. International Journal of Molecular Sciences. 2021; 22(11):5932. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22115932
Chicago/Turabian StyleBoss, Lidia, Marcin Górniak, Alicja Lewańczyk, Joanna Morcinek-Orłowska, Sylwia Barańska, and Agnieszka Szalewska-Pałasz. 2021. "Identification of Three Type II Toxin-Antitoxin Systems in Model Bacterial Plant Pathogen Dickeya dadantii 3937" International Journal of Molecular Sciences 22, no. 11: 5932. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22115932