
Mesoglea Extracellular Matrix Reorganization during Regenerative Process in Anemonia viridis (Forskål, 1775)

 , , , ,
, , , ,
Abstract
:1. Introduction
2. Results
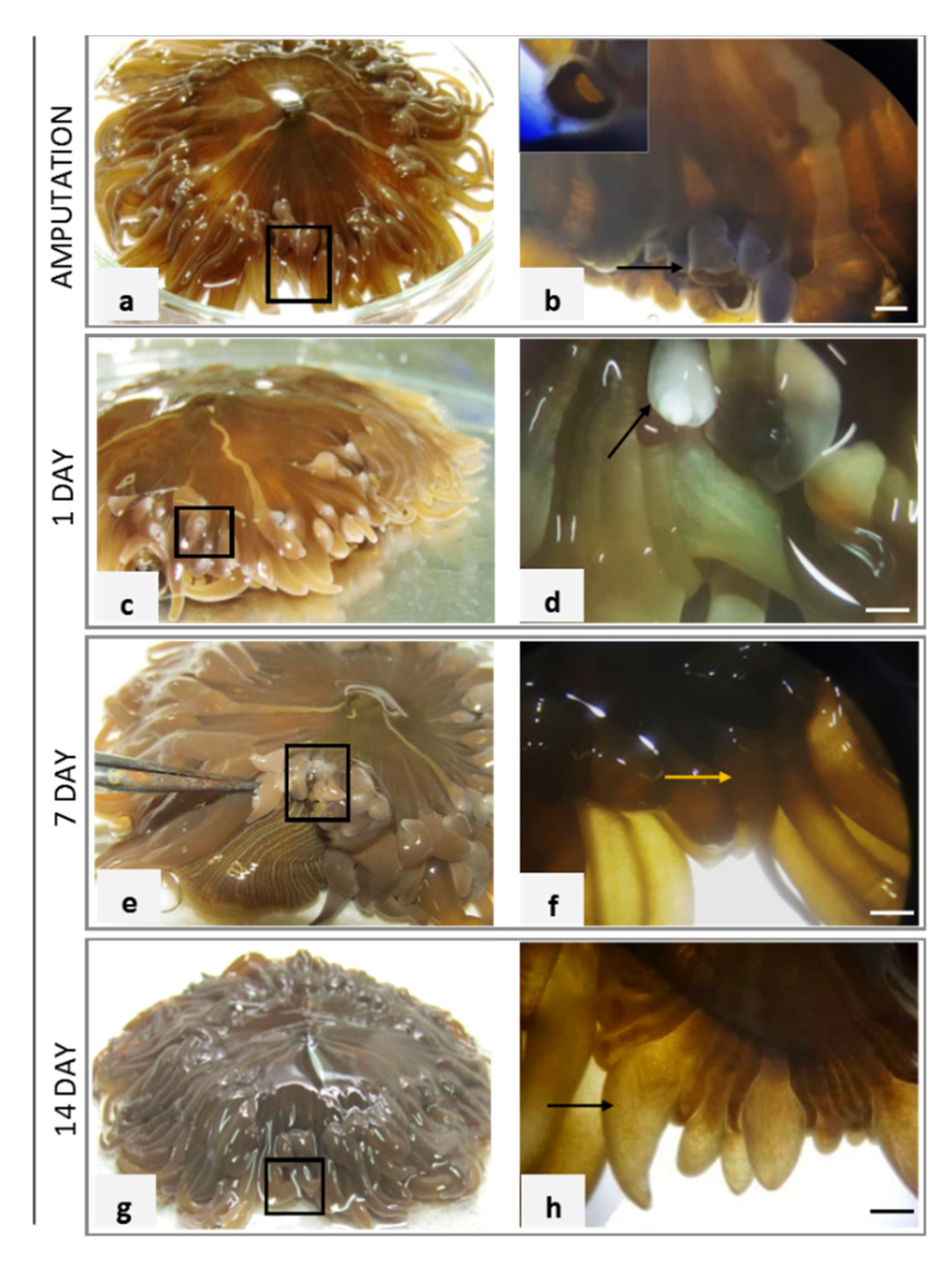
2.1. Macroscopic Assessment of A. viridis Recovery
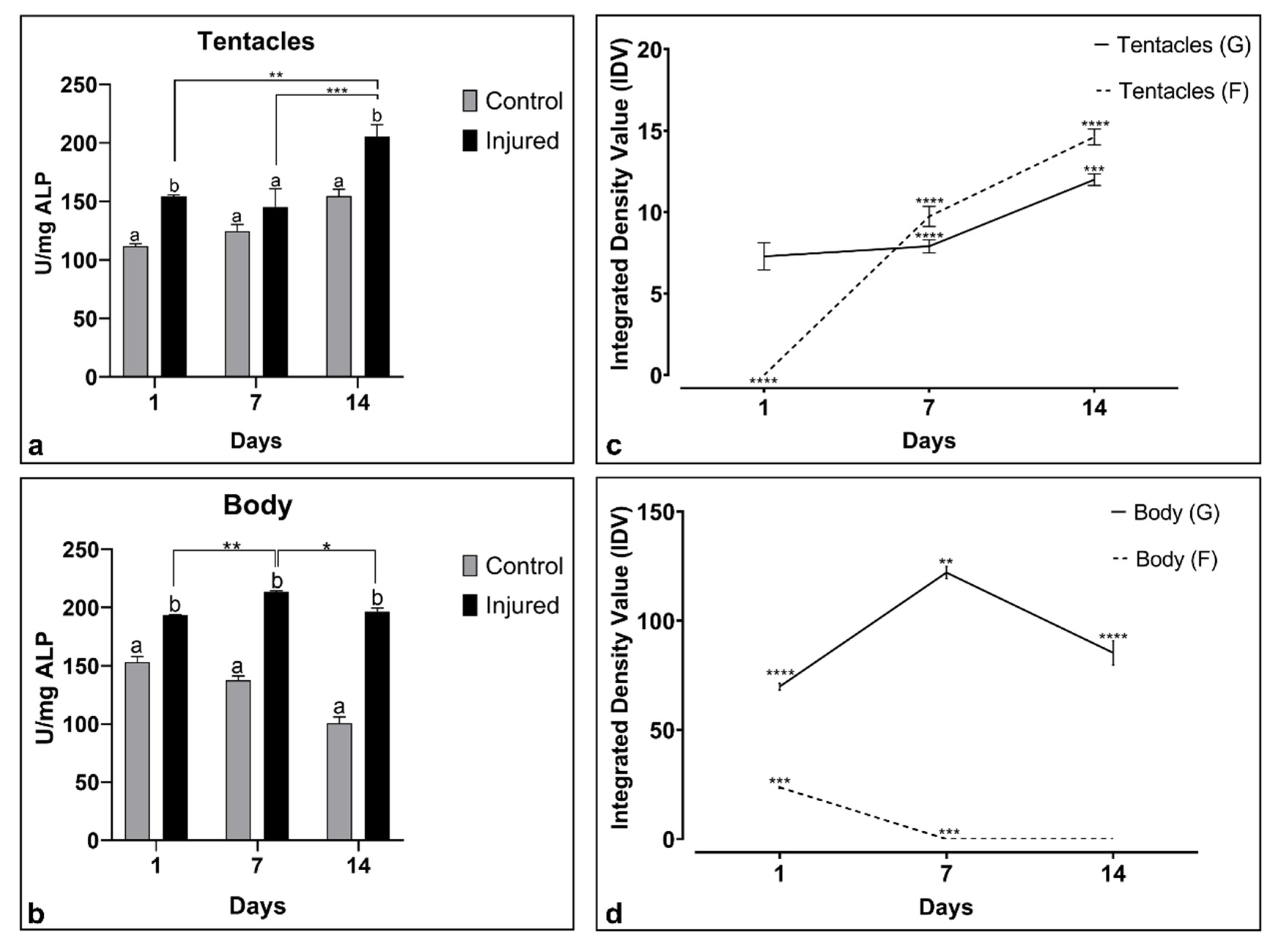
2.2. Biochemical Response
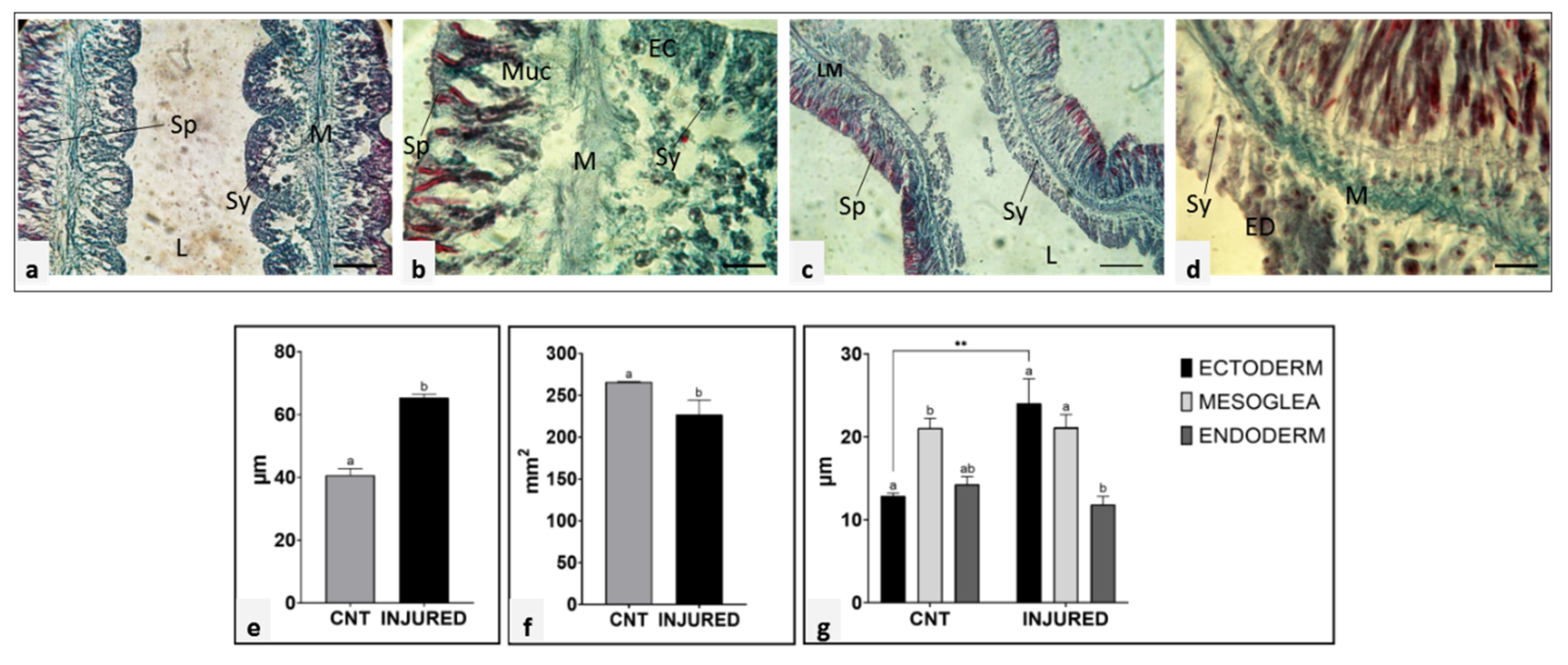
2.3. Biometry of Unamputated and Regenerating Tentacle Tissues
2.4. Morphological and Ultrastructural Analyses of Regenerating Tentacle
2.5. Mesoglea Matrix Reorganization and Collagen Neo-Synthesis
3. Discussion
4. Materials and Methods
4.1. Experimental Plan and Reagents
4.2. Tissues Extraction and Estimation of the Proteins Content
4.3. Alkaline Phosphatase Activity (ALP)
4.4. Protease Activity
4.5. Biometry of the Regenerative Area
4.6. Light and Transmission Electron Microscopy (TEM)
4.6.1. Embedding Tissue in Paraffin
4.6.2. Gomori’s Trichrome
4.6.3. Masson’s Trichrome
4.6.4. Embedding Tissues in Epoxy Resin
4.7. Immunofluorescence Analyses
4.8. Immunogold
4.9. Statistical Analyses
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Dubuc, T.Q.; Traylor-Knowles, N.; Martindale, M.Q. Initiating a regenerative response; cellular and molecular features of wound healing in the cnidarian Nematostella vectensis. BMC Biol. 2014, 12, 24. [Google Scholar] [CrossRef] [Green Version]
- Bosch, T.C. Why polyps regenerate and we don’t: Towards a cellular and molecular framework for Hydra regeneration. Dev. Biol. 2007, 303, 421–433. [Google Scholar] [CrossRef] [Green Version]
- Bely, A.E.; Nyberg, K.G. Evolution of animal regeneration: Re-emergence of a field. Trends Ecol. Evol. 2010, 25, 161–170. [Google Scholar] [CrossRef]
- Holstein, T.W.; Hobmayer, E.; Technau, U. Cnidarians: An Evolutionarily Conserved Model System for Regeneration? Dev. Dyn. 2003, 226, 257–267. [Google Scholar] [CrossRef] [PubMed]
- Mason, B.N.; Califano, J.P.; Reinhart-King, C.A. Matrix Stiffness: A Regulator of Cellular Behavior and Tissue Formation. In Engineering Biomaterials for Regenerative Medicine: Novel Technologies for Clinical Applications; Bhatia, S.K., Ed.; Springer: New York, NY, USA, 2012; pp. 19–37. [Google Scholar]
- Frank, U.; Plickert, G.; Müller, W.A. Cnidarian Interstitial Cells: The Dawn of Stem Cell Research. In Stem Cells in Marine Organisms; Springer: Dordrecht, The Netherlands, 2009; pp. 33–59. [Google Scholar]
- Watanabe, H.; Hoang, V.T.; Mättner, R.; Holstein, T.W. Immortality and the base of multicellular life: Lessons from cnidarian stem cells. Semin. Cell Dev. Biol. 2009, 20, 1114–1125. [Google Scholar] [CrossRef] [PubMed]
- Bosch, T.C.G. Stem Cells in Immortal Hydra. In Stem Cells: From Hydra to Man; Bosch, T.C.G., Ed.; Springer: Dordrecht, The Netherlands, 2008; pp. 37–57. [Google Scholar]
- Bosch, T.C.G. Hydra and the evolution of stem cells. BioEssays 2009, 31, 478–486. [Google Scholar] [CrossRef] [PubMed]
- Galliot, B. Hydra, a fruitful model system for 270 years. Int. J. Dev. Biol. 2012, 56, 411–423. [Google Scholar] [CrossRef]
- Bradshaw, B.; Thompson, K.; Frank, U. Distinct mechanisms underlie oral vs aboral regeneration in the cnidarian Hydractinia echinata. Elife 2015, 4, e05506. [Google Scholar] [CrossRef]
- Dubuc, T.Q.; Schnitzler, C.E.; Chrysostomou, E.; McMahon, E.T.; Febrimarsa; Gahan, J.M.; Buggie, T.; Gornik, S.G.; Hanley, S.; Barreira, S.N.; et al. Transcription factor AP2 controls cnidarian germ cell induction. Science 2020, 367, 757–762. [Google Scholar] [CrossRef] [PubMed]
- Sinigaglia, C.; Peron, S.; Eichelbrenner, J.; Chevalier, S.; Steger, J.; Barreau, C.; Houliston, E.; Leclère, L. Pattern regulation in a regenerating jellyfish. Elife 2020, 9, e54868. [Google Scholar] [CrossRef]
- Fischer, A.B.; Hofmann, D.K. Budding, Bud Morphogenesis, and Regeneration in Carybdea Marsupialis Linnaeus, 1758 (Cnidaria: Cubozoa) In Coelenterate Biology 2003; Fautin, D.G., Westfall, J.A., Cartwrigh, P., Daly, M., Wyttenbach, C.R., Eds.; Springer: Dordrecht, The Netherlands, 2004; pp. 331–337. [Google Scholar]
- Bossert, P.E.; Dunn, M.; Thomsen, G.H. A staging system for the regeneration of a polyp from the aboral physa of the anthozoan CnidarianNematostella vectensis. Dev. Dyn. 2013, 242, 1320–1331. [Google Scholar] [CrossRef] [PubMed]
- Schaffer, A.A.; Bazarsky, M.; Levy, K.; Chalifa-Caspi, V.; Gat, U. A transcriptional time-course analysis of oral vs. aboral whole-body regeneration in the Sea anemone Nematostella vectensis. BMC Genom. 2016, 17, 1–22. [Google Scholar] [CrossRef] [Green Version]
- Amiel, A.R.; Johnston, H.T.; Nedoncelle, K.; Warner, J.F.; Ferreira, S.; Röttinger, E. Characterization of Morphological and Cellular Events Underlying Oral Regeneration in the Sea Anemone, Nematostella vectensis. Int. J. Mol. Sci. 2015, 16, 28449–28471. [Google Scholar] [CrossRef] [PubMed]
- Amiel, A.R.; Foucher, K.; Ferreira, S.; Röttinger, E. Synergic Coordination of Stem Cells Is Required to Induce a Regenerative Response in Anthozoan Cnidarians. BioRxiv 2019. [Google Scholar] [CrossRef]
- Bosch, T.C.G.; Scheider, B.; David, C.N. Role of the cellular environment in interstitial stem cell proliferation in Hydra. Dev. Genes Evol. 1991, 200, 269–276. [Google Scholar] [CrossRef] [Green Version]
- Campbell, R.D.; Bode, H.R. Terminology for Morphology and Cell Types. In Hydra: Research Methods; Lenhoff, H.M., Ed.; Springer: Boston, MA, USA, 1983; pp. 5–14. [Google Scholar]
- Galliot, B.; Quiquand, M.; Ghila, L.; de Rosa, R.; Miljkovic-Licina, M.; Chera, S. Origins of neurogenesis, a cnidarian view. Dev. Biol. 2009, 332, 2–24. [Google Scholar] [CrossRef] [Green Version]
- Augustin, R.; Bosch, T.C.G. Cnidarian Immunity: A Tale of Two Barriers; Springer Science and Business Media LLC: Boston, MA, USA, 2010; pp. 1–16. [Google Scholar]
- Technau, U.; Steele, R.E. Evolutionary crossroads in developmental biology: Cnidaria. Development 2011, 138, 1447–1458. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Palmer, C.V.; Traylor-Knowles, N.G.; Willis, B.L.; Bythell, J.C. Corals Use Similar Immune Cells and Wound-Healing Processes as Those of Higher Organisms. PLoS ONE 2011, 6, e23992. [Google Scholar] [CrossRef] [Green Version]
- Palmer, C.V. Traylor-Knowles Cnidaria: Anthozoans in the Hot Seat. In Advances in Comparative Immunology; Springer: Cham, Switzerland, 2018; pp. 773–791. [Google Scholar]
- Gold, D.A.; Jacobs, D.K. Stem cell dynamics in Cnidaria: Are there unifying principles? Dev. Genes Evol. 2012, 223, 53–66. [Google Scholar] [CrossRef]
- Parisi, M.G.; Parrinello, D.; Stabili, L.; Cammarata, M. Cnidarian Immunity and the Repertoire of Defense Mechanisms in Anthozoans. Biology 2020, 9, 283. [Google Scholar] [CrossRef] [PubMed]
- Davis, L.E.; Haynes, J.F. An ultrastructural examination of the mesoglea of Hydra. Cell Tissue Res. 1968, 92, 149–158. [Google Scholar] [CrossRef] [PubMed]
- Davis, L. Histological and ultrastructural studies of the basal disk of Hydra. Cell Tissue Res. 1975, 162, 107–118. [Google Scholar] [CrossRef] [PubMed]
- Muscatine, L.; Pool, R.R. Regulation of Numbers of Intracellular Algae. Proc. R. Soc. Lond. Ser. B Biol. Sci. 1979, 204, 131–139. [Google Scholar]
- Muller-Parker, G.; Cook, C.; D’Elia, C. Feeding affects phosphate fluxes in the symbiotic sea anemone Aiptasia pallida. Mar. Ecol. Prog. Ser. 1990, 60, 283–290. [Google Scholar] [CrossRef]
- Baranzini, N.; Weiss-Gayet, M.; Chazaud, B.; Monti, L.; De Eguileor, M.; Tettamanti, G.; Acquati, F.; Grimaldi, A. Recombinant HvRNASET2 protein induces marked connective tissue remodelling in the invertebrate model Hirudo verbana. Cell Tissue Res. 2020, 380, 565–579. [Google Scholar] [CrossRef] [PubMed]
- Sarras, M.P., Jr. Components, structure, biogenesis and function of the Hydra extracellular matrix in regeneration, pattern formation and cell differentiation. Int. J. Dev. Biol. 2012, 56, 567–576. [Google Scholar] [CrossRef] [Green Version]
- Ferrier-Pagès, C.; Godinot, C.; D’Angelo, C.; Wiedenmann, J.; Grover, R. Phosphorus metabolism of reef organisms with algal symbionts. Ecol. Monogr. 2016, 86, 262–277. [Google Scholar] [CrossRef]
- Abe, T.; Abe, Y.; Aida, Y.; Hara, Y.; Maeda, K. Extracellular matrix regulates induction of alkaline phosphatase expression by ascorbic acid in human fibroblasts. J. Cell. Physiol. 2001, 189, 144–151. [Google Scholar] [CrossRef]
- De Petrocellis, L.; Orlando, P.; Pierobon, P.; De Falco, M.; Ruggiero, A.M.; Stefano, G.S.; Tino, A.; Grippo, P. Kelletinin A, from the marine mollusc Buccinulum corneum, promotes differentiation in Hydra vulgaris. Res. Commun. Mol. Pathol. Pharmacol. 1999, 103, 17–28. [Google Scholar]
- Parisi, M.G.; Lentini, A.; Cammarata, M. Seasonal changes in morpho-functional aspects of two Anemonia sulcata (Pennant, 1777) wild populations. Mar. Biodivers. 2017, 47, 561–573. [Google Scholar] [CrossRef]
- Stabili, L.; Parisi, M.G.; Parrinello, D.; Cammarata, M. Cnidarian Interaction with Microbial Communities: From Aid to Animal’s Health to Rejection Responses. Mar. Drugs 2018, 16, 296. [Google Scholar] [CrossRef] [Green Version]
- Rader, B.A. Alkaline Phosphatase, an Unconventional Immune Protein. Front. Immunol. 2017, 8, 897. [Google Scholar] [CrossRef] [Green Version]
- Annis, E.; Cook, C. Alkaline phosphatase activity in symbiotic dinoflagellates (zooxanthellae) as a biological indicator of environmental phosphate exposure. Mar. Ecol. Prog. Ser. 2002, 245, 11–20. [Google Scholar] [CrossRef] [Green Version]
- Brown, T.; Rodriguez-Lanetty, M. Defending against pathogens–immunological priming and its molecular basis in a sea anemone, cnidarian. Sci. Rep. 2015, 5, 17425. [Google Scholar] [CrossRef] [Green Version]
- Van De Water, J.A.J.M.; Ainsworth, T.; Leggat, W.; Bourne, D.G.; Willis, B.L.; Van Oppen, M.J.H. The coral immune response facilitates protection against microbes during tissue regeneration. Mol. Ecol. 2015, 24, 3390–3404. [Google Scholar] [CrossRef]
- Trapani, M.; Parisi, M.; Parrinello, D.; Sanfratello, M.; Benenati, G.; Palla, F.; Cammarata, M. Specific inflammatory response of Anemonia sulcata (Cnidaria) after bacterial injection causes tissue reaction and enzymatic activity alteration. J. Invertebr. Pathol. 2016, 135, 15–21. [Google Scholar] [CrossRef]
- Ricci, L.; Srivastava, M. Wound-induced cell proliferation during animal regeneration. Wiley Interdiscip. Rev. Dev. Biol. 2018, 7, e321. [Google Scholar] [CrossRef]
- Barzansky, B.; Lenhoff, H.M.; Bode, H. Hydra mesoglea: Similarity of its amino acid and neutral sugar composition to that of vertebrate basal lamina. Comp. Biochem. Physiol. Part B Comp. Biochem. 1975, 50, 419–424. [Google Scholar] [CrossRef]
- Barzansky, B.; Lenhoff, H.M. On the Chemical Composition and Developmental Role of the Mesoglea of Hydra. Am. Zool. 1974, 14, 575–581. [Google Scholar] [CrossRef]
- Miura, S.; Kimuras, S. The Evolution of Mesogloea Collagen in Coelenterates. J. Biol. Chem. 1985, 260, 15352–15356. [Google Scholar] [CrossRef]
- Schmid, V.; Bally, A. Species specificity in cell-substrate interactions in medusae. Dev. Biol. 1988, 129, 573–581. [Google Scholar] [CrossRef]
- Haller, M. Isolation and Partial Purification of Hydrozoan Collagen and Its Relevance for Species-Specific Cell Adhesion; University of Basel: Basel, Switzerland, 1989. [Google Scholar]
- Adams, E. Invertebrate collagens. Science 1978, 202, 591–598. [Google Scholar] [CrossRef]
- Lee, H.; Jung, E.-S.; Kang, C.; Yoon, W.D.; Kim, J.-S.; Kim, E. Scyphozoan jellyfish venom metalloproteinases and their role in the cytotoxicity. Toxicon 2011, 58, 277–284. [Google Scholar] [CrossRef]
- Li, R.; Yu, H.; Xue, W.; Yue, Y.; Liu, S.; Xing, R.; Li, P. Jellyfish venomics and venom gland transcriptomics analysis of Stomolophus meleagris to reveal the toxins associated with sting. J. Proteom. 2014, 106, 17–29. [Google Scholar] [CrossRef] [PubMed]
- Yan, L.; Pollock, G.; Nagase, H.; Sarras, M. A 25.7 × 103 Mr hydra metalloproteinase (HMP1), a member of the astacin family, localizes to the extracellular matrix of Hydra vulgaris in a head-specific manner and has a developmental function. Development 1995, 121, 1591–1602. [Google Scholar] [CrossRef] [PubMed]
- Yan, L.; Leontovich, A.; Fei, K.; Sarras, M.P. Hydra Metalloproteinase 1: A Secreted Astacin Metalloproteinase Whose Apical Axis Expression Is Differentially Regulated during Head Regeneration. Dev. Biol. 2000, 219, 115–128. [Google Scholar] [CrossRef] [Green Version]
- Jouiaei, M.; Yanagihara, A.A.; Madio, B.; Nevalainen, T.J.; Alewood, P.F.; Fry, B.G. Ancient Venom Systems: A Review on Cnidaria Toxins. Toxins 2015, 7, 2251–2271. [Google Scholar] [CrossRef] [Green Version]
- Tomlin, H.; Piccinini, A.M. A complex interplay between the extracellular matrix and the innate immune response to microbial pathogens. Immunology 2018, 155, 186–201. [Google Scholar] [CrossRef] [PubMed]
- Grimaldi, A.; Tettamanti, G.; Rinaldi, L.; Perletti, G.; Valvassori, R.; De Eguileor, M. Role of Cathepsin B in Leech Wound Healing. Invertebr. Surviv. J. 2004, 1, 38–46. [Google Scholar]
- Tettamanti, G.; Grimaldi, A.; Congiu, T.; Perletti, G.; Raspanti, M.; Valvassori, R.; Eguileor, M. Collagen reorganization in leech wound healing. Biol. Cell 2005, 97, 557–568. [Google Scholar] [CrossRef]
- Godwin, J.; Kuraitis, D.; Rosenthal, N. Extracellular matrix considerations for scar-free repair and regeneration: Insights from regenerative diversity among vertebrates. Int. J. Biochem. Cell Biol. 2014, 56, 47–55. [Google Scholar] [CrossRef] [PubMed]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-Dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Ross, N.W.; Firth, K.J.; Wang, A.; Burka, J.F.; Johnson, S.C. Changes in hydrolytic enzyme activities of naïve Atlantic salmon Salmo salar skin mucus due to infection with the salmon louse Lepeophtheirus salmonis and cortisol implantation. Dis. Aquat. Org. 2000, 41, 43–51. [Google Scholar] [CrossRef] [PubMed]
- Causton, B. The choice of resins for electron immunocytochemistry. In Immunolabelling for Electron Microscopy; Polak, J., Varndell, I., Eds.; Elsevier Science Limited: Amsterdam, The Netherlands; New York, NY, USA; Oxford, UK, 1984; pp. 29–36. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Tentacles | Body | ||||
|---|---|---|---|---|---|
| Activity | Variables | F Value | p Value | F Value | p Value |
| ALP | T | 1.668 | 0.229 | 5.417 | 0.0089 |
| I | 2.716 | 0.125 | 39.83 | <0.0001 | |
| T x I | 4.996 | 0.026 | 6.625 | 0.003 | |
| Tentacles | Body | ||||
|---|---|---|---|---|---|
| Activity | Variables | F Value | p Value | F Value | p Value |
| PROTEOLYTIC | T | 173.5 | <0.0001 | 26.51 | <0.0001 |
| S | 4.984 | 0.00454 | 1527 | <0.0001 | |
| T x S | 56.22 | <0.0001 | 102.8 | <0.0001 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Parisi, M.G.; Grimaldi, A.; Baranzini, N.; La Corte, C.; Dara, M.; Parrinello, D.; Cammarata, M. Mesoglea Extracellular Matrix Reorganization during Regenerative Process in Anemonia viridis (Forskål, 1775). Int. J. Mol. Sci. 2021, 22, 5971. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22115971
Parisi MG, Grimaldi A, Baranzini N, La Corte C, Dara M, Parrinello D, Cammarata M. Mesoglea Extracellular Matrix Reorganization during Regenerative Process in Anemonia viridis (Forskål, 1775). International Journal of Molecular Sciences. 2021; 22(11):5971. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22115971
Chicago/Turabian StyleParisi, Maria Giovanna, Annalisa Grimaldi, Nicolò Baranzini, Claudia La Corte, Mariano Dara, Daniela Parrinello, and Matteo Cammarata. 2021. "Mesoglea Extracellular Matrix Reorganization during Regenerative Process in Anemonia viridis (Forskål, 1775)" International Journal of Molecular Sciences 22, no. 11: 5971. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22115971