
Effect of Salt Stress on the Expression and Promoter Methylation of the Genes Encoding the Mitochondrial and Cytosolic Forms of Aconitase and Fumarase in Maize


Abstract
:1. Introduction
2. Results
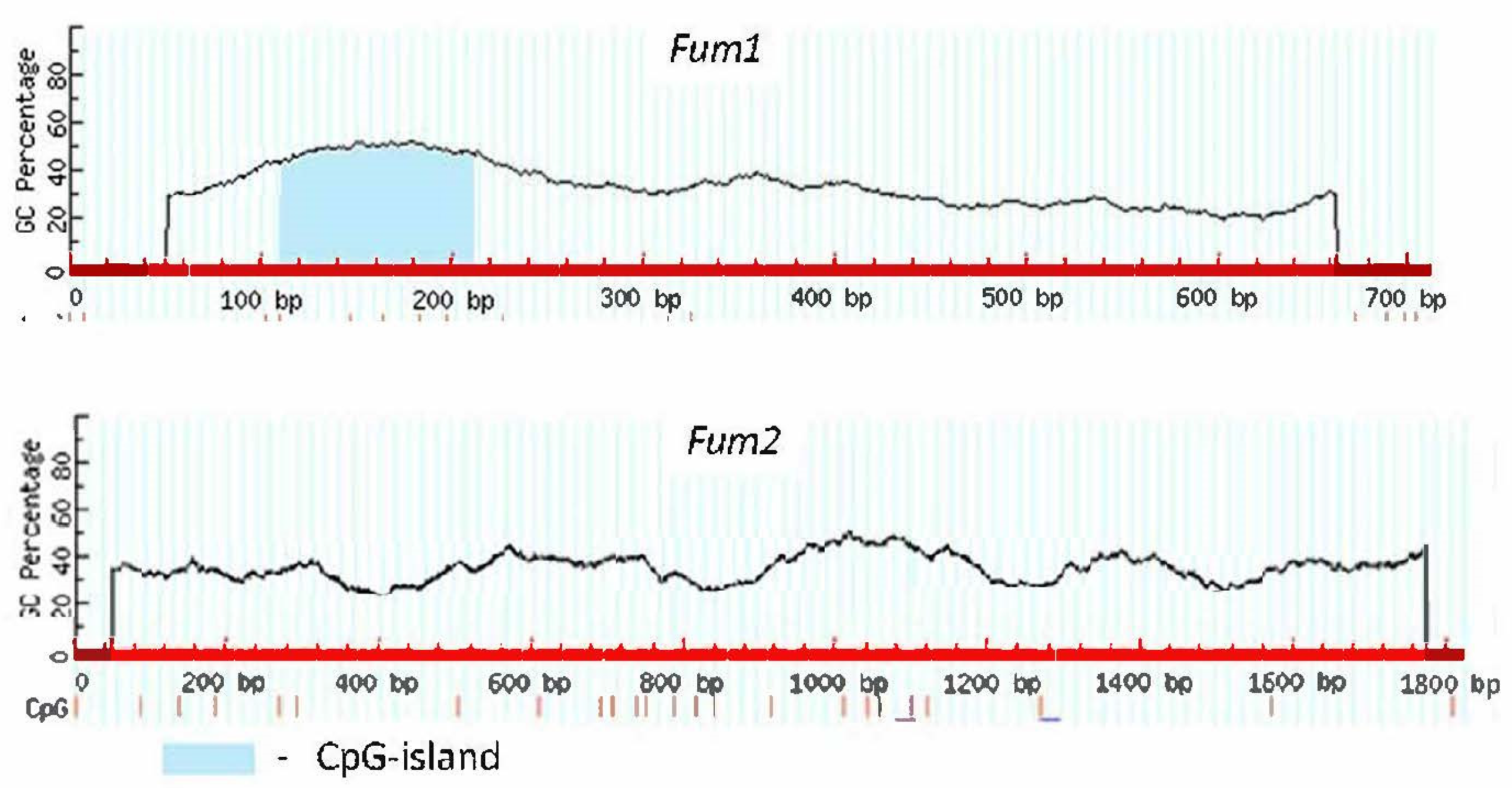
2.1. Analysis of CpG Islands
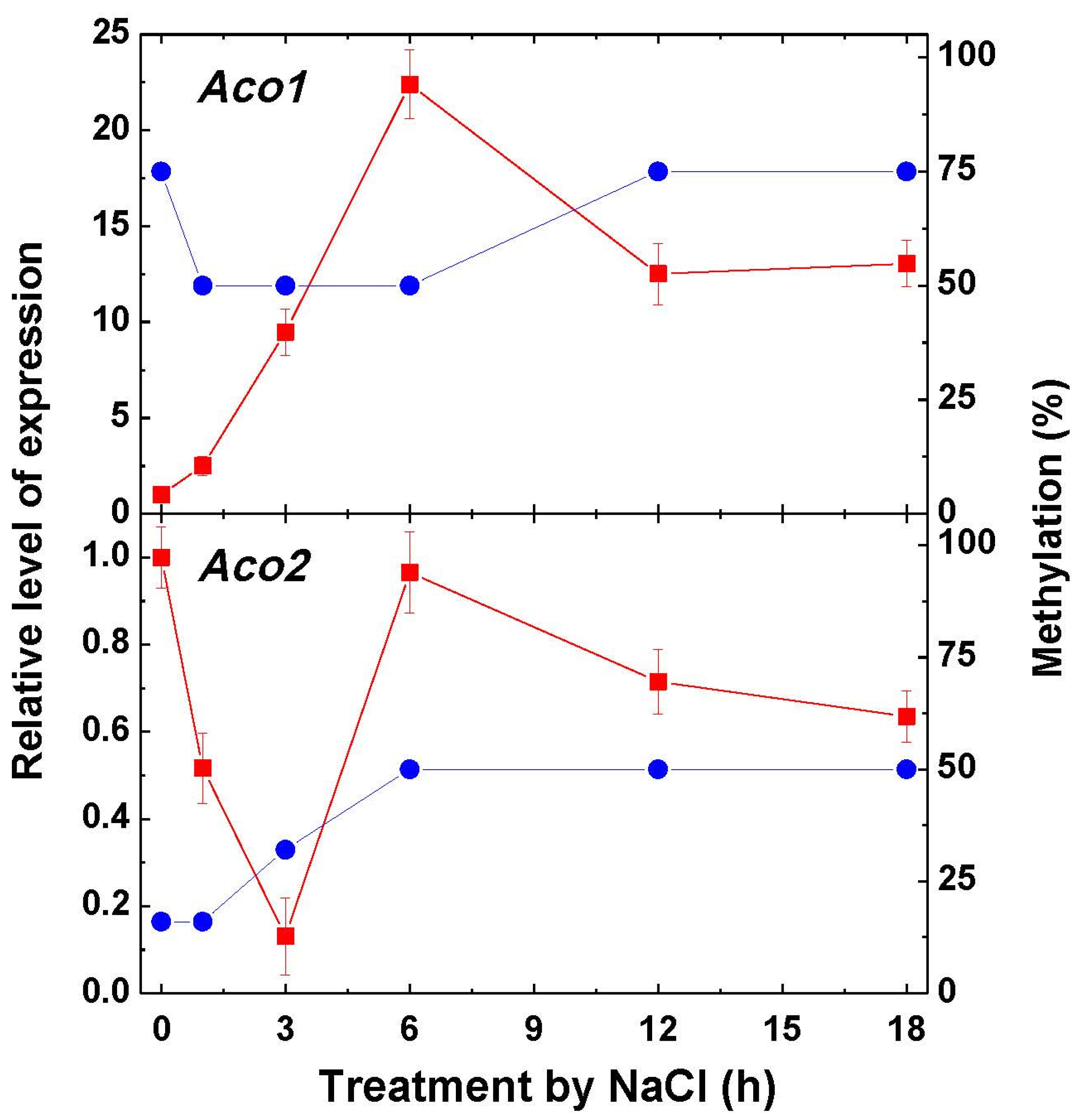
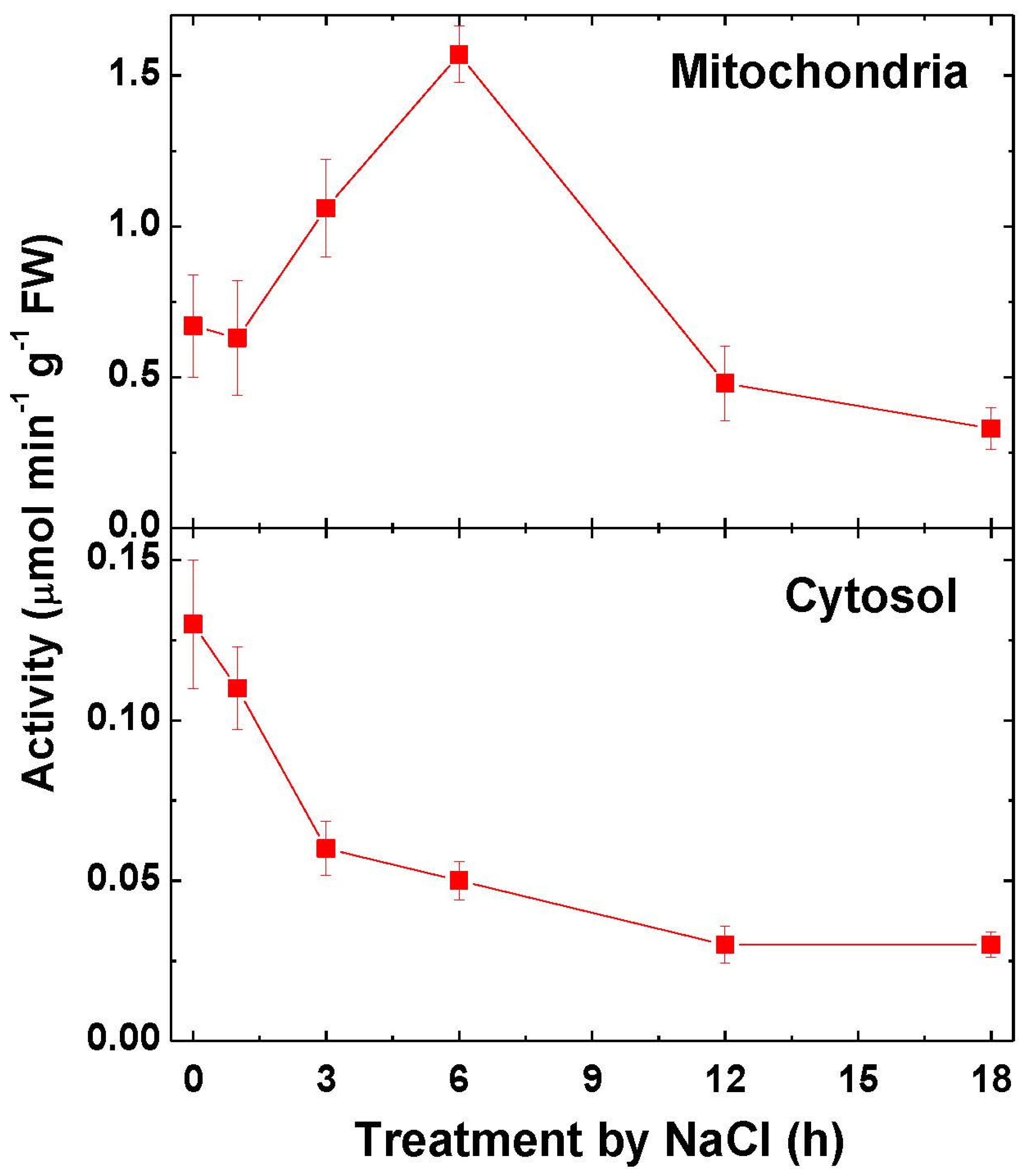
2.2. Aconitase Activity and Expression
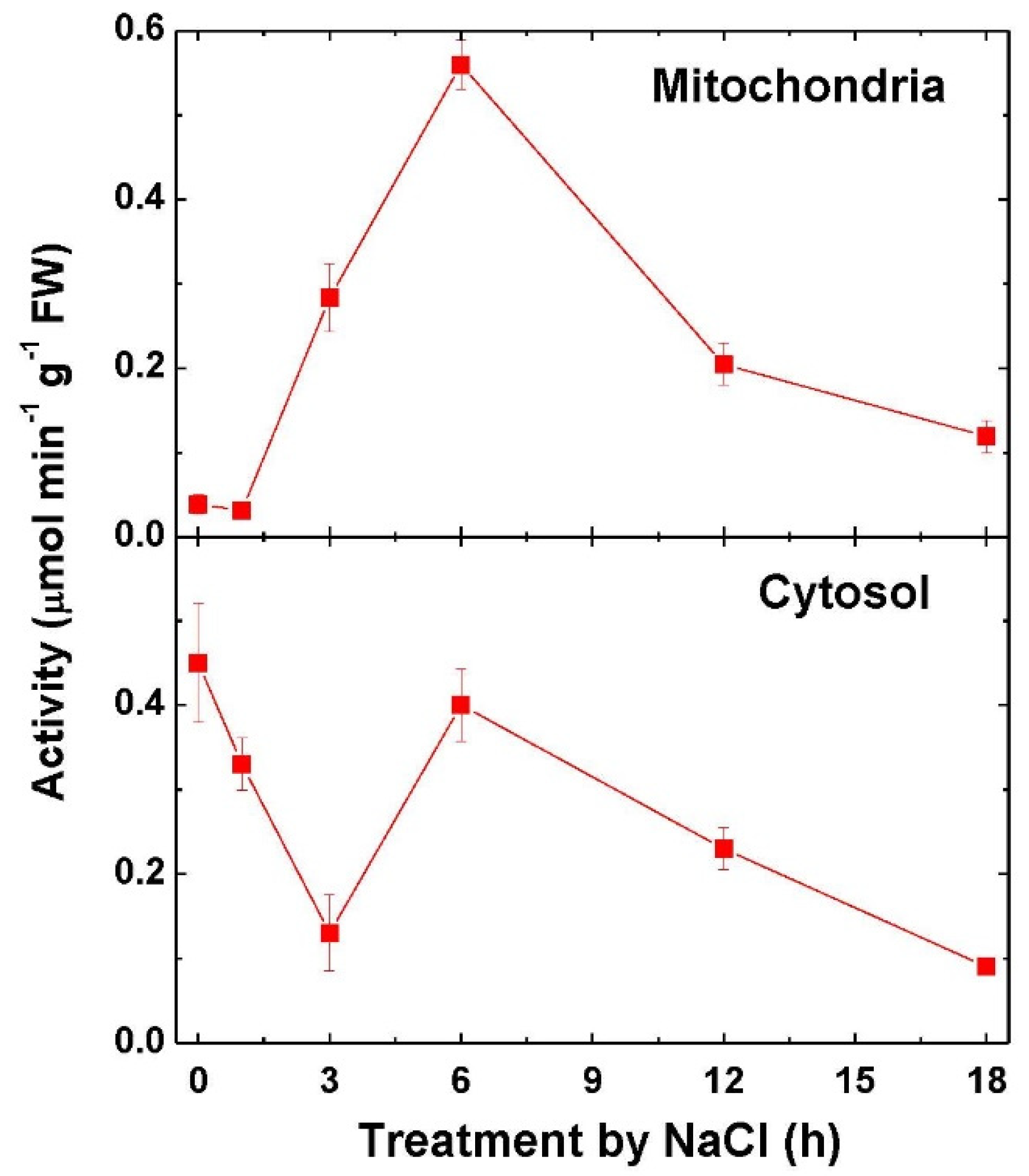
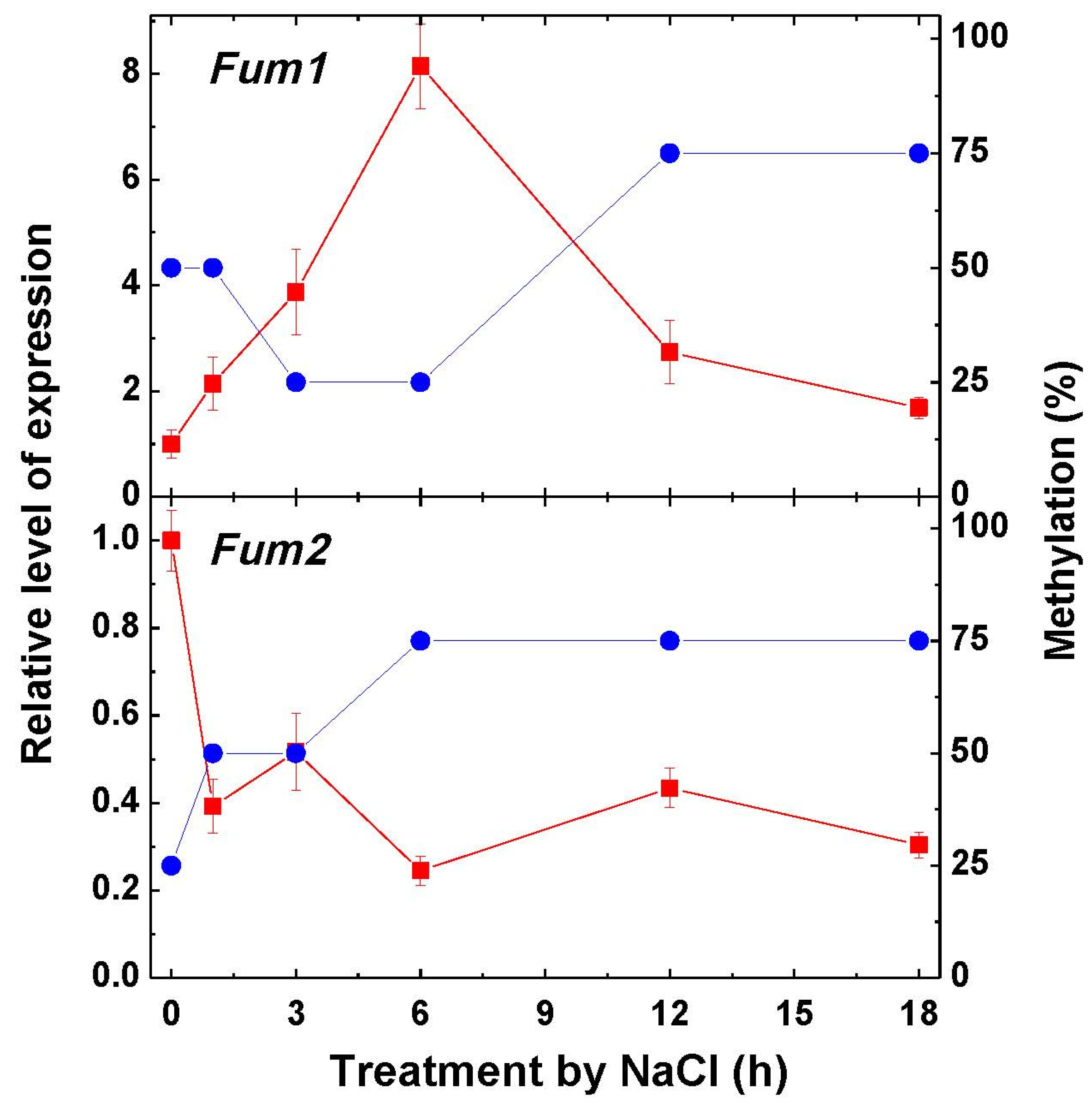
2.3. Fumarase Activity and Expression
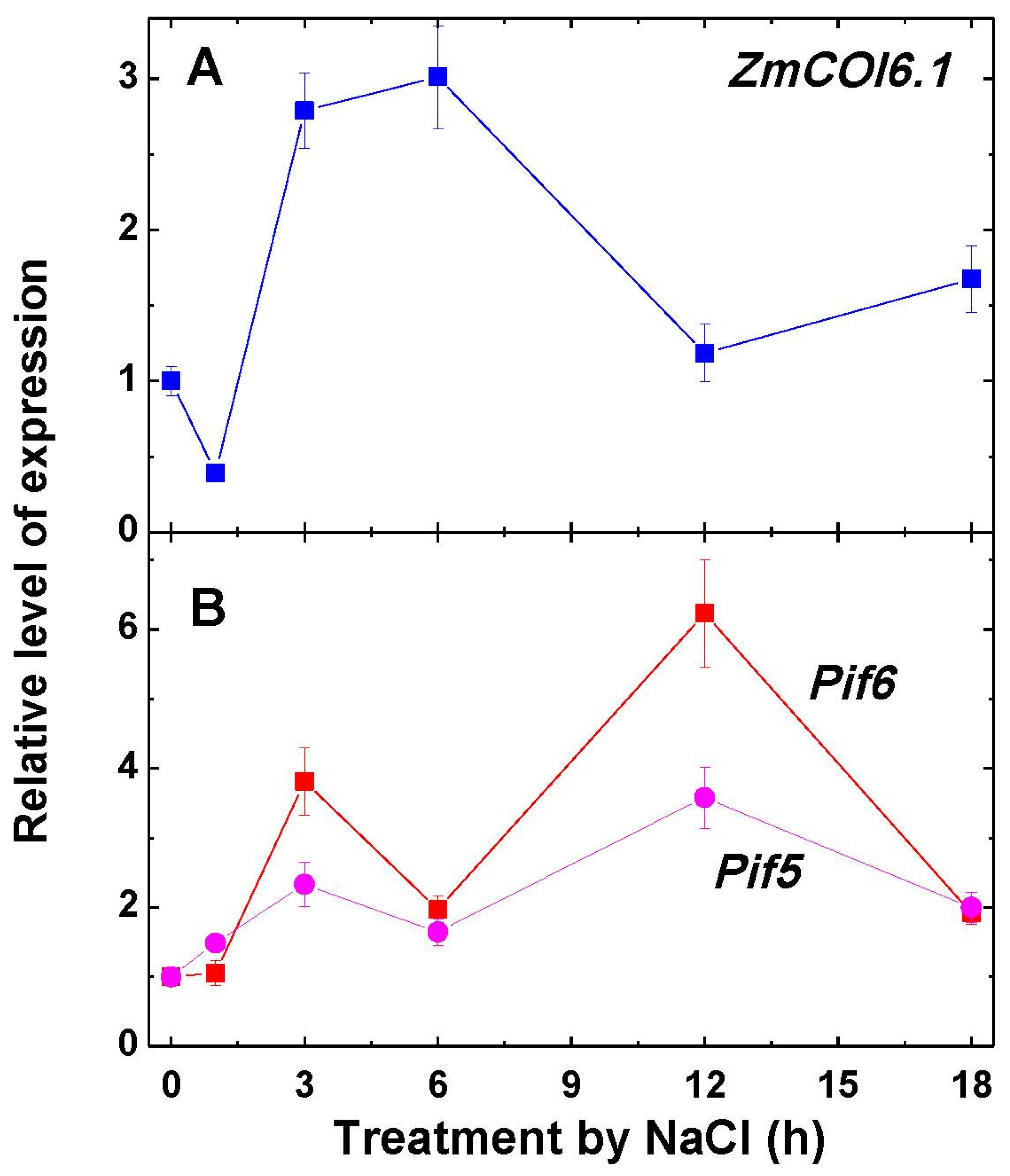
2.4. Expression of ZmCOI6.1, Pif5 and Pif6
3. Discussion
4. Materials and Methods
4.1. Object of Investigation
4.2. Determination of Aconitase and Fumarase Activities
4.3. Subcellular Distribution of Aconitase and Fumarase
4.4. RNA Isolation and PCR Analysis
4.5. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| GABA | gamma-aminobutyric acid |
| PIF | phytochrome interacting factor |
| RNS | reactive nitrogen species |
| ROS | reactive oxygen species |
| TCA cycle | tricarboxylic acid cycle |
References
- Pracharoenwattana, I.; Zhou, W.; Keech, O.; Francisco, P.B.; Udomchalothorn, T.; Tschoep, H.; Stitt, M.; Gibon, Y.; Smith, S.M. Arabidopsis has a cytosolic fumarase required for the massive allocation of photosynthate into fumaric acid and for rapid plant growth on high nitrogen. Plant J. 2010, 62, 785–795. [Google Scholar] [CrossRef] [PubMed]
- Hooks, M.A.; Allwood, J.W.; Harrison, J.K.D.; Kopka, J.; Erban, A.; Goodacre, R.; Balk, J. Selective induction and subcellular distribution of ACONITASE 3 reveal the importance of cytosolic citrate metabolism during lipid mobilization in Arabidopsis. Biochem. J. 2014, 463, 309–317. [Google Scholar] [CrossRef] [Green Version]
- Eprintsev, A.T.; Fedorin, D.N.; Starinina, E.V.; Igamberdiev, A.U. Expression and properties of the mitochondrial and cytosolic forms of fumarase in germinating maize seeds. Physiol. Plant. 2014, 152, 231–240. [Google Scholar] [CrossRef] [PubMed]
- Eprintsev, A.T.; Fedorin, D.N.; Nikitina, M.V.; Igamberdiev, A.U. Expression and properties of the mitochondrial and cytosolic forms of aconitase in maize scutellum. J. Plant Physiol. 2015, 181, 14–19. [Google Scholar] [CrossRef] [PubMed]
- Selinski, J.; Scheibe, R. Malate valves: Old shuttles with new perspectives. Plant Biol. 2019, 21, 21–30. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Igamberdiev, A.U.; Eprintsev, A.T. Organic Acids: The Pools of Fixed Carbon Involved in Redox Regulation and Energy Balance in Higher Plants. Front. Plant Sci. 2016, 7, 1042. [Google Scholar] [CrossRef] [Green Version]
- Igamberdiev, A.U. Citrate valve integrates mitochondria into photosynthetic metabolism. Mitochondrion 2020, 52, 218–230. [Google Scholar] [CrossRef] [PubMed]
- Gupta, K.J.; Shah, J.K.; Brotman, Y.; Jahnke, K.; Willmitzer, L.; Kaiser, W.M.; Bauwe, H.; Igamberdiev, A. Inhibition of aconitase by nitric oxide leads to induction of the alternative oxidase and to a shift of metabolism towards biosynthesis of amino acids. J. Exp. Bot. 2012, 63, 1773–1784. [Google Scholar] [CrossRef] [Green Version]
- Igamberdiev, A.U.; Gardeström, P. Regulation of NAD- and NADP-dependent isocitrate dehydrogenases by reduction levels of pyridine nucleotides in mitochondria and cytosol of pea leaves. Biochim. Biophys. Acta (BBA)-Bioenerg. 2003, 1606, 117–125. [Google Scholar] [CrossRef] [Green Version]
- Flint, D.; Tuminello, J.; Emptage, M. The inactivation of Fe-S cluster containing hydro-lyases by superoxide. J. Biol. Chem. 1993, 268, 22369–22376. [Google Scholar] [CrossRef]
- Navarre, D.A.; Wendehenne, D.; Durner, J.; Noad, R.; Klessig, D.F. Nitric Oxide Modulates the Activity of Tobacco Aconitase. Plant Physiol. 2000, 122, 573–582. [Google Scholar] [CrossRef] [Green Version]
- Schwarzländer, M.; Fricker, M.D.; Sweetlove, L.J. Monitoring the in vivo redox state of plant mitochondria: Effect of respiratory inhibitors, abiotic stress and assessment of recovery from oxidative challenge. Biochim. Biophys. Acta (BBA)-Bioenerg. 2009, 1787, 468–475. [Google Scholar] [CrossRef] [Green Version]
- Bandehagh, A.; Taylor, N.L. Can Alternative Metabolic Pathways and Shunts Overcome Salinity Induced Inhibition of Central Carbon Metabolism in Crops? Front. Plant Sci. 2020, 11, 1072. [Google Scholar] [CrossRef]
- Robertson, R.N.; Wilkins, M. Quantitative Relation Between Salt Accumulation and Salt Respiration in Plant Cells. Nat. Cell Biol. 1948, 161, 101. [Google Scholar] [CrossRef]
- Van Zelm, E.; Zhang, Y.; Testerink, C. Salt Tolerance Mechanisms of Plants. Annu. Rev. Plant Biol. 2020, 71, 403–433. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Igamberdiev, A.U.; Hill, R.D. Elevation of cytosolic Ca2+ in response to energy deficiency in plants: The general mechanism of adaptation to low oxygen stress. Biochem. J. 2018, 475, 1411–1425. [Google Scholar] [CrossRef] [PubMed]
- Hill, C.B.; Jha, D.; Bacic, A.; Tester, M.; Roessner, U. Characterization of Ion Contents and Metabolic Responses to Salt Stress of Different Arabidopsis AtHKT1;1 Genotypes and Their Parental Strains. Mol. Plant 2013, 6, 350–368. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chojak-Koźniewska, J.; Kuźniak, E.; Linkiewicz, A.; Sowa, S. Primary carbon metabolism-related changes in cucumber exposed to single and sequential treatments with salt stress and bacterial infection. Plant Physiol. Biochem. 2018, 123, 160–169. [Google Scholar] [CrossRef] [PubMed]
- Che-Othman, M.H.; Jacoby, R.P.; Millar, A.H.; Taylor, N.L. Wheat mitochondrial respiration shifts from the tricarboxylic acid cycle to the GABA shunt under salt stress. New Phytol. 2020, 225, 1166–1180. [Google Scholar] [CrossRef]
- Del-Saz, N.F.; Florez-Sarasa, I.; Clemente-Moreno, M.J.; Mhadhbi, H.; Flexas, J.; Fernie, A.R.; Ribas-Carbó, M. Salinity tolerance is related to cyanide-resistant alternative respiration in Medicago truncatula under sudden severe stress. Plant Cell Environ. 2016, 39, 2361–2369. [Google Scholar] [CrossRef]
- Sweetman, C.; Miller, T.K.; Booth, N.J.; Shavrukov, Y.; Jenkins, C.L.; Soole, K.L.; Day, D.A. Identification of Alternative Mitochondrial Electron Transport Pathway Components in Chickpea Indicates a Differential Response to Salinity Stress between Cultivars. Int. J. Mol. Sci. 2020, 21, 3844. [Google Scholar] [CrossRef]
- Beyrne, C.C.; Iusem, N.D.; González, R.M. Effect of Salt Stress on Cytosine Methylation within GL2, An Arabidopsis thaliana Gene Involved in Root Epidermal Cell Differentiation. Absence of Inheritance in the Unstressed Progeny. Int. J. Mol. Sci. 2019, 20, 4446. [Google Scholar] [CrossRef] [Green Version]
- Wolny, E.; Skalska, A.; Braszewska, A.; Mur, L.; Hasterok, R. Defining the Cell Wall, Cell Cycle and Chromatin Landmarks in the Responses of Brachypodium distachyon to Salinity. Int. J. Mol. Sci. 2021, 22, 949. [Google Scholar] [CrossRef]
- Amaral, M.N.D.; Auler, P.A.; Rossatto, T.; Barros, P.M.; Oliveira, M.M.; Braga, E.J.B. Long-term somatic memory of salinity unveiled from physiological, biochemical and epigenetic responses in two contrasting rice genotypes. Physiol. Plant. 2020, 170, 248–268. [Google Scholar] [CrossRef] [PubMed]
- Amraee, L.; Rahmani, F.; Mandoulakani, B.A. 24-Epibrassinolide alters DNA cytosine methylation of Linum usitatissimum L. under salinity stress. Plant Physiol. Biochem. 2019, 139, 478–484. [Google Scholar] [CrossRef] [PubMed]
- Lu, X.; Shu, N.; Wang, J.; Chen, X.; Wang, D.; Wang, S.; Fan, W.; Guo, X.; Guo, L.; Ye, W. Genome-wide analysis of salinity-stress induced DNA methylation alterations in cotton (Gossypium hirsutum L.) using the Me-DIP sequencing technology. Genet. Mol. Res. 2017, 16, 2. [Google Scholar] [CrossRef] [PubMed]
- Eprintsev, A.T.; Fedorin, D.N.; Cherkasskikh, M.V.; Igamberdiev, A.U. Regulation of expression of the mitochondrial and cytosolic forms of aconitase in maize leaves via phytochrome. Plant Physiol. Biochem. 2020, 146, 157–162. [Google Scholar] [CrossRef]
- Igamberdiev, A.U.; Bykova, N.V. Role of organic acids in the integration of cellular redox metabolism and mediation of redox signalling in photosynthetic tissues of higher plants. Free. Radic. Biol. Med. 2018, 122, 74–85. [Google Scholar] [CrossRef]
- Yogev, O.; Yogev, O.; Singer, E.; Shaulian, E.; Goldberg, M.; Fox, T.D.; Pines, O. Fumarase: A Mitochondrial Metabolic Enzyme and a Cytosolic/Nuclear Component of the DNA Damage Response. PLoS Biol. 2010, 8, e1000328. [Google Scholar] [CrossRef] [Green Version]
- Eprintsev, A.T.; Fedorin, D.N.; Karabutova, L.A.; Igamberdiev, A.U. Expression of genes encoding subunits A and B of succinate dehydrogenase in germinating maize seeds is regulated by methylation of their promoters. J. Plant Physiol. 2016, 205, 33–40. [Google Scholar] [CrossRef]
- Eprintsev, A.T.; Fedorin, D.N.; Dobychina, M.A.; Igamberdiev, A.U. Expression and promoter methylation of succinate dehydrogenase and fumarase genes in maize under anoxic conditions. J. Plant Physiol. 2017, 216, 197–201. [Google Scholar] [CrossRef]
- Eprintsev, A.T.; Fedorin, D.N.; Sazonova, O.V.; Igamberdiev, A.U. Expression and properties of the mitochondrial and cytosolic forms of fumarase in sunflower cotyledons. Plant Physiol. Biochem. 2018, 129, 305–309. [Google Scholar] [CrossRef] [PubMed]
- Jayawardhane, J.; Cochrane, D.W.; Vyas, P.; Bykova, N.V.; Vanlerberghe, G.C.; Igamberdiev, A.U. Roles for Plant Mitochondrial Alternative Oxidase Under Normoxia, Hypoxia, and Reoxygenation Conditions. Front. Plant Sci. 2020, 11, 566. [Google Scholar] [CrossRef] [PubMed]
- Bemer, M.; Baroux, C. Unraveling the Complex Epigenetic Mechanisms that Regulate Gene Activity. Adv. Struct. Saf. Stud. 2017, 1675, 205–231. [Google Scholar] [CrossRef]
- Nguyen, H.T.; Leipner, J.; Stamp, P.; Guerra-Peraza, O. Low temperature stress in maize (Zea mays L.) induces genes involved in photosynthesis and signal transduction as studied by suppression subtractive hybridization. Plant Physiol. Biochem. 2009, 47, 116–122. [Google Scholar] [CrossRef]
- Gao, Y.; Jiang, W.; Dai, Y.; Xiaoyi, T.; Zhang, C.; Li, H.; Lu, Y.; Wu, M.; Tao, X.; Deng, D.; et al. A maize phytochrome-interacting factor 3 improves drought and salt stress tolerance in rice. Plant Mol. Biol. 2015, 87, 413–428. [Google Scholar] [CrossRef] [PubMed]
- Eprintsev, A.T.; Fedorin, D.N.; Sazonova, O.V.; Igamberdiev, A.U. Light inhibition of fumarase in Arabidopsis leaves is phytochrome A–dependent and mediated by calcium. Plant Physiol. Biochem. 2016, 102, 161–166. [Google Scholar] [CrossRef]
- Eprintsev, A.T.; Fedorin, D.N.; Cherkasskikh, M.V.; Igamberdiev, A.U. Expression of succinate dehydrogenase and fumarase genes in maize leaves is mediated by cryptochrome. J. Plant Physiol. 2018, 221, 81–84. [Google Scholar] [CrossRef] [PubMed]
- AbdElgawad, H.; Zinta, G.; Hegab, M.M.; Pandey, R.; Asard, H.; Abuelsoud, W. High Salinity Induces Different Oxidative Stress and Antioxidant Responses in Maize Seedlings Organs. Front. Plant Sci. 2016, 7, 276. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moore, A.L.; Proudlove, M.O. Mitochondria and sub-mitochondrial particles. In Isolation of Membranes and Organelles from Plant Cells; Hall, T.L., Moore, A.L., Eds.; Academic Press: London, UK, 1983; pp. 153–184. [Google Scholar]
- Lowry, O.H.; Rosebrough, N.J.; Farr, A.L.; Randall, R.J. Protein measurement with the Folin phenol reagent. J. Biol. Chem. 1951, 193, 265–275. [Google Scholar] [CrossRef]
- Rozen, S.; Skaletsky, H. Primer3 on the WWW for General Users and for Biologist Programmers. Bioinform. Methods Protoc. 2000, 132, 365–386. [Google Scholar] [CrossRef] [Green Version]
- Nicot, N.; Hausman, J.-F.; Hoffmann, L.; Evers, D. Housekeeping gene selection for real-time RT-PCR normalization in potato during biotic and abiotic stress. J. Exp. Bot. 2005, 56, 2907–2914. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of Relative Gene Expression Data Using Real-Time Quantitative PCR and the 2−ΔΔCT Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Li, L.C.; Dahiya, R. MethPrimer: Designing primers for methylation PCRs. Bioinformatics 2002, 18, 1427–1431. [Google Scholar] [CrossRef] [PubMed] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Primer | Sequence |
|---|---|---|
| Aco1 | forward | 5′-TGGAAGGAGATGCTGTCAGT-3′ |
| reverse | 5′-CGTATAGCGCCATCCACATG-3′ | |
| Aco2 | forward | 5′-CAAGTTCTTCAGCCTTCCGG-3′ |
| reverse | 5′-GCAAGGTCTACAACTGCTGG-3′ | |
| Fum1 | forward | 5′-GATTACTTCGATCATTGAGGT-3′ |
| reverse | 5′-ACCAGAACTCGCGGATGTGGC-3′ | |
| Fum2 | forward | 5′-ACCAGAACTCGCGGATGTGGC-3′ |
| reverse | 5′-TGGTTCATTCTCAGGCAGAGA-3′ | |
| ZmCOI6.1 | forward | 5′-GGGTGTTCCTCAAGTACGGG-3′ |
| reverse | 5′-GGGTGGGTACGGTAGCAAAA-3′ | |
| Pif5 | forward | 5′-GAAAGAACCTTTCCGCGTGT-3′ |
| reverse | 5′-AAAGCAGGGCATGTGAACAG-3′ | |
| Pif6 | forward | 5′-TGAATCCAGGTTGCATCCCT-3′ |
| reverse | 5′-CATCCCTGAGGTGACCGATT-3′ |
| Gene | Position of the Studied Cytosine | Primer | Sequence | |
|---|---|---|---|---|
| Aco1 | I | -425 bp | forward M | 5′-TGTATTTTAGAAATAGAGTTTTGCGT-3′ |
| reverse M | 5′-CTACGCACGAATTAAATTCGAAT-3′ | |||
| forward U | 5′-ATGTATTTTAGAAATAGAGTTTTGTGT-3′ | |||
| reverse U | 5′-ATACCTACACACAAATTAAATTCAAA-3′ | |||
| II | -565 bp | forward M | 5′-TAGAGATTATTTTTCGATTCGATTC-3′ | |
| reverse M | 5′-CTACGCACGAATTAAATTCGAAT-3′ | |||
| forward U | 5′-GAGATTATTTTTTGATTTGATTTGGTT-3′ | |||
| reverse U | 5′-ATACCTACACACAAATTAAATTCAAA-3′ | |||
| IIII | -941 bp | forward M | 5′-TTATTAACGTTTAGCGGTAGAGTTC-3′ | |
| reverse M | 5′-CTACGCACGAATTAAATTCGAAT-3′ | |||
| forward U | 5′-ATTAATGTTTAGTGGTAGAGTTTGT-3′ | |||
| reverse U | 5′-ATACCTACACACAAATTAAATTCAAA-3′ | |||
| Aco2 | I | -579 bp | forward M | 5′-ATTCGAAAATAGTGAGAAGTTGTC-3′ |
| reverse M | 5′-TAAAAAACTAACTAACCAAATCGCT-3′ | |||
| forward U | 5′-TTATTTGAAAATAGTGAGAAGTTGTTG-3′ | |||
| reverse U | 5′-TCAAACAAACCAAAACATAATACATACA-3′ | |||
| II | -821 bp | forward M | 5′-GTTTATTGTACGTATAAGGAAGC-3′ | |
| reverse M | 5′-TAAAAAACTAACTAACCAAATCGCT-3′ | |||
| forward U | 5′-GTTTATTGTATGTATAAGGAAGTGG-3′ | |||
| reverse U | 5′-TCAAACAAACCAAAACATAATACATACA-3′ | |||
| III | -1034 bp | forward M | 5′-GGTATTATAAAATTTATTAAGGTCGG-3′ | |
| reverse M | 5′-TAAAAAACTAACTAACCAAATCGCT-3′ | |||
| forward U | 5′-AGTTTGGTATTATAAAATTTATTAAGGTTG-3′ | |||
| reverse U | 5′-TCAAACAAACCAAAACATAATACATACA-3′ | |||
| Fum1 | I | -778 bp | forward M | 5′-TAGGAATAATTTAAATAATACGG-3′ |
| reverse M | 5′-TCTATTATAAAATAATACTTTCCC-3′ | |||
| forward U | 5′-GTAGGAATAATTTAAATAATATGG-3′ | |||
| reverse U | 5′-TCTATTATAAAATAATACTTTCCC-3′ | |||
| II | -717 bp | forward M | 5′-GGTAGGGAATGGTTTGCGT-3′ | |
| reverse M | 5′-TCTATTATAAAATAATACTTTCCC-3′ | |||
| forward U | 5′-GGTAGGGAATGGTTTGTGT-3′ | |||
| reverse U | 5′-TCTATTATAAAATAATACTTTCCC-3′ | |||
| IIII | -552 bp | forward M | 5′-TTGAAGGTTATTTATTTATACGG-3′ | |
| reverse M | 5′-TCTATTATAAAATAATACTTTCCC-3′ | |||
| forward U | 5′-ATTGAAGGTTATTTATTTATATGG-3′ | |||
| reverse U | 5′-TCTATTATAAAATAATACTTTCCC-3′ | |||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Eprintsev, A.T.; Fedorin, D.N.; Cherkasskikh, M.V.; Igamberdiev, A.U. Effect of Salt Stress on the Expression and Promoter Methylation of the Genes Encoding the Mitochondrial and Cytosolic Forms of Aconitase and Fumarase in Maize. Int. J. Mol. Sci. 2021, 22, 6012. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22116012
Eprintsev AT, Fedorin DN, Cherkasskikh MV, Igamberdiev AU. Effect of Salt Stress on the Expression and Promoter Methylation of the Genes Encoding the Mitochondrial and Cytosolic Forms of Aconitase and Fumarase in Maize. International Journal of Molecular Sciences. 2021; 22(11):6012. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22116012
Chicago/Turabian StyleEprintsev, Alexander T., Dmitry N. Fedorin, Mikhail V. Cherkasskikh, and Abir U. Igamberdiev. 2021. "Effect of Salt Stress on the Expression and Promoter Methylation of the Genes Encoding the Mitochondrial and Cytosolic Forms of Aconitase and Fumarase in Maize" International Journal of Molecular Sciences 22, no. 11: 6012. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22116012